A minor population of splenic dendritic cells expressing CD19 mediates IDO-dependent T cell suppression via type I IFN signaling following B7 ligation
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
International Immunology, Vol. 17, No. 7, pp. 909–919 ª The Japanese Society for Immunology. 2005. All rights reserved. doi:10.1093/intimm/dxh271 For permissions, please e-mail: journals.permissions@oupjournals.org A minor population of splenic dendritic cells expressing CD19 mediates IDO-dependent T cell suppression via type I IFN signaling following B7 ligation Babak Baban1*, Anna M. Hansen1*, Phillip R. Chandler1,2, Anna Manlapat1, Adam Bingaman1,3, David J. Kahler1, David H. Munn1,4 and Andrew L. Mellor1,2 1 Immunotherapy Center and 2Department of Medicine, 3Department of Surgery and 4Department of Pediatrics, Medical College of Georgia, 1120, 15th Street, Augusta, GA 30912, USA Keywords: CTLA4-Ig, dendritic cells, interferon, STAT1, T cell suppression Abstract By ligating CD80/CD86 (B7) molecules, the synthetic immunomodulatory reagent CTLA4-Ig (soluble synthetic CTLA4 fusion protein) induces expression of the enzyme indoleamine 2,3-dioxygenase (IDO) in some dendritic cells (DCs), which acquire potent T cell regulatory functions as a consequence. Here we show that this response occurred exclusively in a population of splenic DCs co-expressing the marker CD19. B7 ligation induced activation of the transcription factor signal transducer and activator of transcription (STAT1) in sorted CD191, but not CD19NEG, DCs. STAT1 activation occurred even when DCs lacked receptors for type II IFN (IFNc); however, STAT1 activation and IDO up-regulation were not observed when DCs lacked receptors for type I IFN (IFNab). Thus, IFNa, but not IFNc, signaling was essential for STAT1 activation and IDO up-regulation in CD191 DCs following B7 ligation. Consistent with these findings, B7 ligation also induced sorted CD191, but not CD19NEG, DCs to express IFNa. Moreover, recombinant IFNa induced CD191, but not CD19NEG, DCs to mediate IDO-dependent T cell suppression, showing that IFNa signaling could substitute for upstream signals from B7. These data reveal that a minor population of splenic DCs expressing the CD19 marker is uniquely responsive to B7 ligation, and that IFNa-mediated STAT1 activation is an essential intermediary signaling pathway that promotes IDO induction in these DCs. Thus, CD191 DCs may be a target for regulatory T cells expressing surface CTLA4, and may suppress T cell responses via induction of IDO. Introduction Induced or spontaneous indoleamine 2,3-dioxygenase (IDO) Grohmann and colleagues reported that murine DCs re- activity suppresses adaptive T cell-mediated immunity in sponded to CTLA4-Ig (soluble synthetic CTLA4 fusion pro- murine models of pregnancy, allogeneic tissue transplantation tein)-mediated ligation of B7 (CD80/86) molecules by and inflammatory diseases. These include tumor growth, expressing functional IDO, and that the ability of CTLA4-Ig to asthma and autoimmune diseases, such as colitis, insulin- delay tissue allograft rejection was partly IDO dependent (6). dependent diabetes mellitus, rheumatoid arthritis and multiple Consistent with this, we found that CTLA4-Ig-mediated sclerosis (1, 2). Human and murine dendritic cells (DCs) blockade of destructive donor allospecific T cell responses acquire potent T cell regulatory properties following induction was completely dependent on the ability of CTLA4-Ig to of IDO expression (1, 3, 4). In mice, IDO expression by murine induce IDO in recipient mice (7). However, IDO expression CD8a+ DCs suppressed delayed-type hypersensitivity re- was detected exclusively in specific DC subsets with sponses to tumor-associated peptide antigens (5). Recently, distinct phenotypes (B220+, CD8a+) that acquired potent *These authors contributed equally to this study. Correspondence to: A. L. Mellor; E-mail: amellor@mcg.edu Received 31 January 2005, accepted 20 April 2005 Transmitting editor: E. Simpson Advance Access publication 20 June 2005
910 IDO-expressing CD19+ regulatory dendritic cells
IDO-dependent T cell regulatory functions following in vivo Mixed lymphocyte reactions
exposure to CTLA4-Ig (7, 8). Since B7 molecules are Mixed lymphocyte reactions (MLRs) were performed essen-
expressed widely on a variety of antigen-presenting cells tially as described previously (14). Combinations of respond-
(APCs), these findings suggested that the ability to respond to ers and stimulators were set up in triplicate wells in a total
CTLA4-Ig treatment by expressing functional IDO was re- of 200 ll per well RPMI 1640 medium (catalog no. 15-041-
stricted to specific populations of splenic DCs co-expressing CV; Cellgro, Herndon, VA, USA) supplemented with 10%
B220 and/or CD8a. fetal bovine serum (FBS, Sigma), penicillin (100 IU mlÿ1),
IFNc (IFN type II) is a potent inducer of IDO expression in 100 mg mlÿ1 streptomycin (Cellgro), 2 mM mlÿ1 L-glutamine
multiple cell types, such as cultured cell lines, tumor cells and (Cellgro) and 5 3 10ÿ5 M 2-mercaptoethanol in 96-well round-
physiologic murine splenic CD8a+ DCs (5, 9). IFNc induces bottomed plates (Falcon, Bedford, MA, USA). Responder
IDO gene transcription in vitro via activation of signal trans- T cells were enriched using nylon wool (15) and used at either
ducer and activator of transcription (STAT1), a member of the 1 3 105 or 5 3 104 cells per well together with equal numbers
signal transducer and activator family of transcription factors of fractionated or 53 unfractionated non-irradiated stimula-
(6, 10). However, following in vivo treatment with CTLA4-Ig, tors. Plates were incubated for 72 h at 37C in a humidified 5%
splenic DCs from mice deficient for IFNcR expression CO2 atmosphere. Wells were pulsed with 0.5 lCi [3H]thymi-
[IFNcRa-KO (gene deficient) mice] mediated IDO-dependent dine ([3H]TdR) in 40 ll RPMI 1640 for the last 6 h of the
T cell suppression as efficiently as DCs from wild-type mice, incubation period. TdR incorporation was measured using the
suggesting that IFNc signaling was not essential for induction BetaPlate system (Wallac, Newark, NJ).
of functional IDO activity in DCs following B7 ligation in vivo (8).
In the current study, we show that B7 ligation mediated by
1-Methyl-tryptophan and 103 tryptophan
CTLA4-Ig induced a highly selective pattern of IFNa (IFN type
I) secretion and STAT1 activation restricted to a specific 1-Methyl-D-tryptophan (1mT; Aldrich, Milwaukee, WI, USA)
population of splenic DCs. The principal responsive subset was added to give a final concentration of 100 lM.
comprised a minor DC population expressing the marker L-Tryptophan (L-a-amino-3-indole-propionic acid FW 204.2,
CD19. These DCs appeared thus to be similar to CD19+ Sigma) was used at a final concentration of 245 lM to give
plasmacytoid DCs that we recently identified as the principal 103 the normal concentration used in stock RPMI (24.5 lM
IDO+ regulatory DC population present in tumor-draining final).
lymph nodes (TDLNs) (11).
Anti-IFN antibody
Monoclonal rat anti-murine IFNa antibody (catalog no. 22100-
Methods 1) and monoclonal rat anti-murine IFNc antibody (catalog no.
22500-1) were purchased from PBL Biomedical Laboratories.
Mice
F1[CBA 3 B6], IDO-deficient (IDO-KO with F1[CBA 3 B6] Anti-IDO antibody
backgrounds), BALB/c and BM3 (CBA) TCR transgenic mice Polyclonal rabbit anti-murine IDO antibody was prepared by
used in these studies were bred at the Medical College of a commercial supplier (Biosource International, Hopkinton,
Georgia (12, 13). IFNabR-KO, IFNcR-KO and (background MA, USA). Antisera were raised against two synthetic peptides
matched) strain 129 wild-type mice were generous gifts from (KPTDGDKSEEPSNVESRGC and CSAVERQDLKALEKALHD)
D. Moskofidis (Medical College of Georgia). IFNabR-KO mice following conjugation to ovalbumin. Antisera were affinity
with BALB/c backgrounds were generous gifts from W. purified over the first peptide and screened for reactivity by
Portnoy (University of California, Berkeley, CA, USA). All ELISA.
procedures were carried out with the approval of the In-
stitutional Animal Care and Use Committee. Immunohistochemistry
Tissue sections (5 mm) were prepared from formalin-fixed
paraffin-embedded tissues. Following de-paraffinization, sec-
CTLA4-Ig
tions were washed for 10 min in distilled water. Cytospin
Native CTLA4-Ig (non-mutant, catalog no. C4483) and mutant preparations of ~20 000 sorted cells per sample chamber
(catalog no. C4358) isotypes of CTLA4-IgG2a were pur- were centrifuged (700 r.p.m., 5 min), air-dried, fixed in 10%
chased from Sigma (St Louis, MO, USA). Mice were injected formalin and washed twice in PBS. All subsequent procedures
with 100 lg CTLA4-Ig (intra-peritoneally) and DCs were were carried out at room temperature (RT). Endogenous
incubated with 100 lg mlÿ1 of CTLA4-Ig. Unless otherwise peroxidase activity was blocked with hydrogen peroxide
stated in the text, the native CTLA4-Ig isotype was used for (1 : 10 w/PBS, 10 min). Tissue sections were also treated
studies described. with proteinase K (catalog no. S3020; DAKO, Carpentaria, CA,
USA) for 10 min. After two washes in PBS, all preparations
were treated with universal blocking reagent at 1 : 10 in
Recombinant IFN distilled water (catalog no. HK085-5K; BioGenex, San Ramon,
Recombinant mouse IFNa (catalog no. 12100-1) and IFNc CA, USA), rinsed in PBS and incubated with either anti-IDO
(catalog no. 12500-1) were purchased from PBL Biomedical antibody or anti-IFNa antibody (1 : 100 in PBS; 1 h for
Laboratories (Piscataway, NJ, USA). cytospins, 2 h for tissue sections). After two washes in PBS,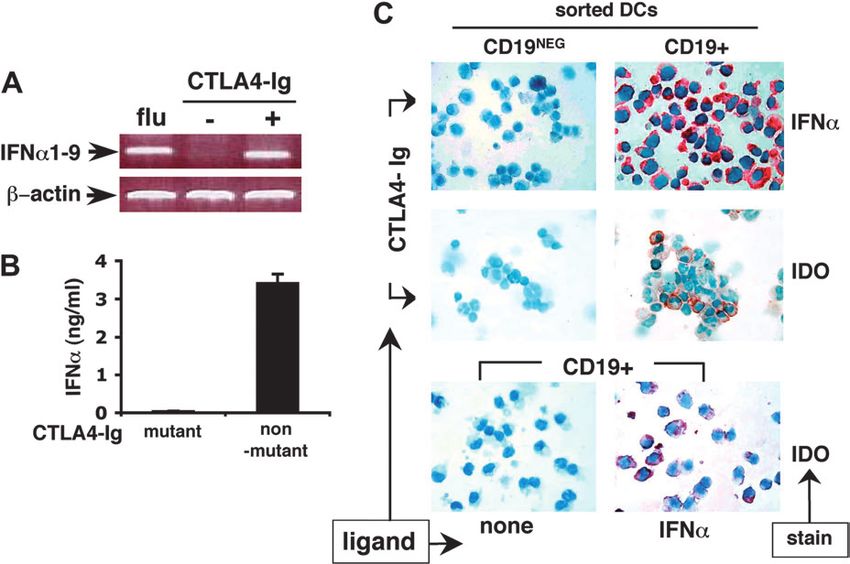
IDO-expressing CD19+ regulatory dendritic cells 911
preparations were treated with biotinylated goat anti- CA, USA) and PE–CD19 (catalog no. 557329, Pharmingen) for
rabbit Ig (catalog no. HK336-9R, BioGenex). After a 5-min 20 min at 4C. Preparative cell sorting was performed as
wash in PBS, slides were incubated for 20 min in peroxidase- described (8), using a Mo-Flo four-way flow cytometer equipped
conjugated streptavidin (catalog no. HK330-9k, BioGenex). with DakoCytomation SummitTM software (DakoCytomation, Ft
IDO-expressing cells were visualized using 3-amino-9- Collins, CO, USA) to select cells of interest. CD11c+ cell
ethylcarbazole chromogen (catalog no. HK121-5K Liquid fractions were selected for high purity (>98%), which was
AEC, BioGenex) for 30 s to 10 min as necessary for optimal achieved by setting sorting gates to collect cells unambiguously
staining. Preparations were counterstained with hematoxylin stained by CD11c mAb (CD11cHIGH). This procedure sacrificed
(catalog no. 7221; Richard-Allan Scientific, Kalamazoo, MI, some DCs with lower CD11c-staining profiles (CD11cLOW), but
USA) and mounted in Faramount (catalog no. S3025, DAKO). avoided contamination with macrophages and other cell types
Anti-IDO antibody pre-incubated with neutralizing peptide whose autofluorescence overlapped the CD11cLOW region. As
(1.2 mg antibody : 10 mg peptide) was used as the specificity shown in Results, essentially all IDO-dependent T cell suppres-
control. sive activities segregated with the unambiguous CD11cHIGH
sorted cells that co-expressed CD19, so it was not necessary to
Immunofluorescence (STAT1 and P-STAT1) staining include ambiguous CD11cLOW DC populations for the purposes
Tissue sections and cytospin preparations were prepared as of this study. Sorting gates for CD19 staining were set between
above. To permeabilize, all preparations were incubated in distinct populations of stained and unstained cells. All sorted
0.2% Triton X-100 for 5 min at RT. All slides were washed three DCs exhibited comparable light scatter properties (FSCHIGH,
times for 5 min at RT and then incubated in blocking buffer SSCHIGH) characteristic of large mononuclear cells.
(20% normal donkey serum, 1% BSA, 0.02% NaN3, 13 PBS)
for 45–60 min. Following treatment with the primary antibody Analytical flow cytometry
[phospho-(Y701)-STAT1 (P-STAT1), antibody catalog no. Phenotypic analyses of splenic DCs were performed using
9171; Cell Signaling Technology, Beverly, MA, USA] overnight four-color flow cytometry with dye-conjugated mAbs. DC
at 4C, preparations were then washed three times with Tris- subsets were identified using a cocktail of mAb to CD11c,
buffered saline (TBS) for 5 min each time. All slides were then B220, CD19 and CD8a and cell-surface markers were
incubated with the secondary fluorescence-labeled antibody identified using PE-conjugated mAb to H2Kb, H-2Ak/Ek,
(1 : 100, catalog no 711-166-152; Jackson Immunoresearch CD80 and CD86 (all from BD Biosciences, San Diego, CA,
Laboratories, West Grove, PA, USA) for 1 h in the dark at RT, USA). The CD11c gate was set to match sorting parameters
washed twice in TBS for 5 min each time in the dark and then shown in Fig. 1(A) to permit comparisons with Mo-Flo-sorted
counterstained using bis-Benzimide, Hoechst (catalog no. DC populations and with our previous studies (8).
B-2883, Sigma).
Reverse transcription–polymerase chain reaction
Splenic DC isolation
Analysis of IFNa gene expression in CD11c+-enriched DC
Spleens were harvested into 1% FBS/HBSS. One milliliter of population treated or untreated with 100 lg mlÿ1 CTLA4-Ig
collagenase IV (100 CD units mlÿ1 in 1% FBS/HBSS-CLS-4; was performed using semi-quantitative reverse transcription–
Worthington, Lakewood, NJ, USA) was injected into three polymerase chain reaction (RT–PCR). Total RNA was isolated
areas of each spleen. Injected spleens were then placed in from cells using RNA STAT-60 (catalog no. CS-110; Tel-Test
collagenase IV (1 ml per spleen of 400 CD units mlÿ1 in 1% Inc., Friendswood, TX, USA). A total of 3 ng of RNA was
FBS/HBSS). After incubation (37C, 30 min), spleens were amplified for 40 cycles (IFNa: 94C for 30 s, 65C for 1 min and
made into a single-cell suspension and centrifuged (1300 72C for 2 min; b-actin: 94C for 30 s, 52.5C for 1 min and
r.p.m., 5 min) and erythrocytes lysed (3 min) in 3 ml of ACK 68C for 2 min), following reverse transcription in a one-step
lysing buffer (catalog no. 10-548E; BioWhittaker, Walkersville, reaction (RT–PCR ‘Access’; Promega, Madison, WI, USA). A
MD, USA). Splenocytes were washed twice (10 mM EDTA total of 10 ll of reaction was electrophoresed on a 1% agarose
in Ca/Mg-free PBS) before fractionation (MACS) or sorting gel. Primers for amplification of specific IFNa subtype tran-
(Mo-Flo) as described below. scripts were IFNa1–9, forward CCTGATGGTCTTGGTGGTGATAA
and reverse CAGTTCCTTCATCCCGACCAG (16) and for b-
AutoMACS fractionation actin transcripts were forward AGCAAGAGAGGTATCCTG and
Cell pellets were re-suspended in running buffer (1% BSA, or reverse CTTTACGGATGTCAACGTC. As controls, 2 3 106
2% FCS in 1 mM EDTA in Ca/Mg-free PBS), and anti-murine CD11c+ DC cells were infected with influenza A virus strain X31
CD11c microbeads (catalog no. 130-052-001; Miltenyi, Auburn, (courtesy of Graeme Price) at multiplicity of infection 10 in PBS
CA, USA) were added (50 ll mlÿ1). Following incubation (30 min for 1 h at 37C. Cells were pelleted (5 min, 700 3 g) and re-
on ice) in the dark, cells were washed twice and CD11c+ cells suspended in 0.5 ml RPMI for 4 h. Total RNA was isolated from
were selected using the AutoMACS system. Typically, CD11c+ cells using RNA STAT-60.
cells isolated by this procedure were 80–85% pure, while
CD11cÿ cells were >99% pure. Western blots
A total of 106 CD11c+ DCs were enriched by AutoMACS, treated
Preparative flow cytometry or untreated with 100 lg mlÿ1 CTLA4-Ig in vitro for 5 h, harvested
Splenocyte cell suspensions were incubated with a cocktail of in cell lysis buffer (PBS, 1% NP40, 0.5% sodium deoxycho-
APC–CD11c (catalog no. 550261; Pharmingen, San Diego, late, 0.1% SDS, 150 ng mlÿ1 phenylmethylsulfonylfluoride,912 IDO-expressing CD19+ regulatory dendritic cells
Fig. 1. IDO-competent splenic DCs express CD19. Splenic DCs from F1[CBA 3 B6] mice treated 24 h earlier with PBS (ÿ) or 100 lg CTLA4-Ig (+)
were used as stimulators in MLRs using responder BM3 (H-2Kb specific) T cells. T cell proliferation was assessed after 72 h as described in
Methods. DCs were sorted by (Mo-Flo) rapid flow cytometry (8). FACS profiles show the gating criteria used to select (A) purified DCs (>99%
CD11c+) and (B) sorted CD19+ and CD19NEG DC populations used in MLRs. (C) MLRs contained unfractionated (total) splenocytes, or sorted
CD19+ and CD19NEG DCs as APCs, and responder T cells (nylon wool enriched) from BM3 TCR transgenic mice. Parallel (triplicate) MLRs
contained no additions (black bars), or excess (103 normal) tryptophan (white bars). Arrows in panel C indicate percent inhibition of T cell
proliferation due to IDO activity.
100 ng mlÿ1 aprotinin) and 30 lg of cell protein was levels of CD11c (CD11cHIGH) and excluded the majority of
electrophoresed on 10% polyacrylamide gels overlaid with splenic plasmacytoid DCs, which express relatively low levels
a 5% stacking gel. Protein was quantitated using the of CD11c (17, 18). As we have shown previously, splenic
bicinchonic acid assay (Pierce, Rockford, IL, USA). Anti- CD11cHIGH DC populations sorted in this way from untreated
body against activated STAT1 (P-STAT1, Tyr701, catalog no. mice and from IDO-KO mice treated with CTLA4-Ig stimulated
91H; Cell Signaling Technology) was used in combination vigorous BM3 T cell proliferation (8). In contrast, unfractionated
with standard ECL techniques. (total) splenocytes (Fig. 1A and B) and sorted DC populations
co-expressing B220 or CD8a mediated IDO-dependent T cell
ELISA suppression (8).
A total of 106 CD11c+ DCs from IDO-WT mice, enriched by IDO-mediated T cell suppression segregated with sorted
AutoMACS, were treated with 100 lg mlÿ1 non-mutant or DC populations expressing CD19 when prepared from
mutant CTLA4-Ig in vitro for 5 h. Media were then harvested CTLA4-Ig-treated mice (Fig. 1C). Lack of T cell proliferation
and measured for IFNa as per manufacturer’s instructions was due to IDO-mediated suppression because underlying
(Mouse IFN Alpha ELISA kit; PBL Biomedical Laboratories). potent stimulatory properties of DCs from CTLA4-Ig-treated
mice became evident when excess tryptophan was added to
cultures. Identical outcomes were obtained when IDO inhibitor
Results 1mT was added to cultures (data not shown). Sorted CD19NEG
DCs from mice exposed to CTLA4-Ig were not suppressive
CD19+ DCs mediate IDO-dependent T cell suppression and stimulated robust BM3 T cell proliferation, though their
In a recent study on murine DCs from TDLNs, we reported that T cell stimulatory properties were slightly enhanced in the
DC populations expressing the surface marker CD19 were the presence of excess tryptophan (Fig. 1C). The suppressive
only cells that mediated IDO-dependent T cell suppression effects of CD19+ DCs were potent and dominant since CD19+
(11). To examine if CD19 also identified the population of DCs DCs were a minor DC population (Fig. 1B), yet they completely
from spleen that mediated IDO-dependent T cell suppression, suppressed T cell proliferation in the presence of CD19NEG
we sorted CD19+ and CD19NEG DCs from untreated and DCs that stimulated potent T cell responses only when
CTLA4-Ig-treated F1[CBA 3 B6] mice using a rapid (Mo-Flo) separated from CD19+ DCs (Fig. 1C). These data revealed
flow cytometer, and then cultured sorted DCs with H-2Kb- that a minor DC population expressing CD19 mediated IDO-
specific CD8+ T cells from BM3 TCR transgenic mice to assess dependent T cell suppression following CTLA4-Ig treatment
their T cell stimulatory properties (Fig. 1). DC populations were in vivo.
gated based on purity criteria (>98% CD11c+, see Methods). We performed multi-color flow cytometric analyses to
This sorting strategy selected DCs expressing relatively high evaluate the phenotypic characteristics of DC populationsIDO-expressing CD19+ regulatory dendritic cells 913
expressing the CD19 marker that mediated potent IDO- Table 1. Phenotypic analyses of splenic DC subsets
dependent T cell suppression. Approximately 50% of total
CD11c+ splenocytes fell within populations gated using the Marker B220+ B220+ B220NEG B220NEG
criteria shown in Fig. 1(A) (CD11cHIGH). Within this gated CD8+ CD8NEG CD8+NEG CD8NEG
(~10%)a (~12%) (~29%) (49%)
CD11cHIGH DC population and consistent with data in Fig.
1(B), CD19 staining was heterogeneous, though highest levels CD19 938 6 39b 1169 6 71 115 6 12 39 6 4
of CD19 expression were detected on minor DC populations MHCI 831 6 32 534 6 38 517 6 28 277 6 10
that also co-expressed B220; these B220+ cells accounted for MHCII 1371 6 94 1401 6 86 771 6 65 844 6 115
CD80 558 6 2 348 6 26 424 6 10 229 6 5
~20% of DCs falling within the CD11cHIGH-gated DC popu- CD86 809 6 29 441 6 47 617 6 41 233 6 39
lation (Table 1). Much lower levels of CD19 were detected on
CD8a+(B220NEG) DC populations and B220NEGCD8aNEG DCs a
Approximate percentage of CD11cHIGH DCs using gates shown in
did not express detectable CD19. CD19+ DCs expressed high Fig. 1(A). bMean fluorescence intensity (from more than three
levels of MHC class I and MHC class II (MHCI, MHCII) and B7 separate experiments).
(CD80, CD86) compared with CD19NEG DCs, suggesting that
CD19+ DCs were mature DCs. Identical outcomes were
obtained when phenotypic analyses were performed on DCs intracellular location of STAT1 in sorted splenic DC populations
from untreated mice and from mice exposed to CTLA4-Ig prior cultured with CTLA4-Ig in vitro (Fig. 2D–I). After culture with
to flow cytometric analyses. Thus, in vivo CTLA4-Ig treatment CTLA4-Ig for 5 h, >80% of MACS-enriched CD19+ DCs
had no detectable effect on the phenotypic characteristics of stained with anti-P-STAT1 antibody (Fig. 2G), and staining co-
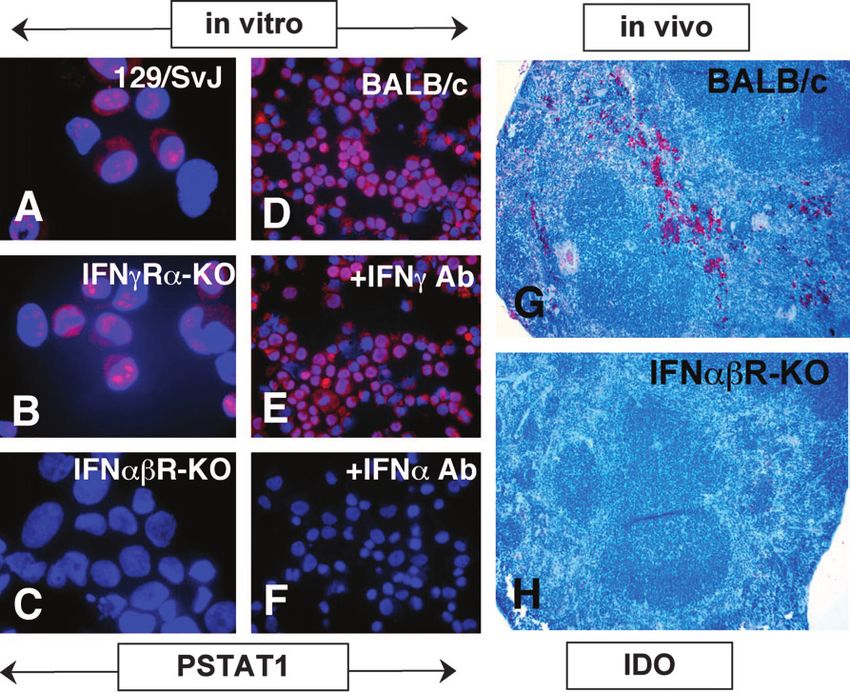
DCs or the relative proportions of DC subsets (data not localized to cell nuclei. In contrast,914 IDO-expressing CD19+ regulatory dendritic cells Fig. 2. B7 ligation selectively activates STAT1 in DC subsets. (A–C) AutoMACS-enriched CD11c+ DCs from an IFNcRa-KO (129/SvJ) mouse were incubated with CTLA4-Ig (100 lg mlÿ1) for 5 h and cytospins were stained with anti-P-STAT1 (Y701-phosphorylated form) antibody (B, red) and counterstained with Hoechst dye (A, blue) to highlight nuclei (C, merged A + B images). (D–I) Splenic CD11c+ DCs from F1[CBA 3 B6] mice were sorted into CD19NEG/CD19+ (D/G), B220NEG/B220+ (E/H) or CD8aNEG/CD8a+ (F/I) subsets, incubated with CTLA4-Ig (100 lg mlÿ1) for 5 h and stained to detect nuclear P-STAT1 as in (A–C). Anti-B220 (E and H) or anti-CD8a mAbs (F and I) conjugated to green dye were used to verify the efficiency of Mo-Flo sorting. (K–M) F1[CBA 3 B6] mice were treated with non-mutant (K and L) or mutant (M) CTLA4-Ig and 24 h later spleen sections were stained with anti-P-STAT1 antibody (red) and Hoechst dye (blue) as before (K, red image only; L and M, merged images;). Lymphoid follicles are outlined by dotted lines in merged images. Original magnifications: 31000 (A–L) or 3100 (K–M). (Fig. 3C). These data suggested that signaling through IFN STAT1 activation following B7 ligation. DCs from F1[CBA 3 B6] type I receptors was essential for STAT1 activation following B7 mice were cultured with CTLA4-Ig alone (Fig. 3D) or in the ligation, while signaling through type II IFN receptors was not presence of CTLA4-Ig and mAbs that neutralized IFNc (Fig. 3E) essential for this response. and IFNa (Fig. 3F), and STAT1 activation was assessed as To test if IFN type I signaling was essential for IDO up- before. While anti-IFNc mAb had no significant effect on the regulation following B7 ligation, we injected CTLA4-Ig into proportion of DCs containing intra-nuclear P-STAT1 (30–50% of BALB/c mice with defective expression of IFNabRs, harvested DCs in Fig. 3D and E), anti-IFNa mAb completely blocked spleens 24 h later and stained tissue sections with anti-IDO STAT1 activation in a dose-dependent manner, and no P- antibody (Fig. 3G and H). Consistent with previous studies STAT1+ DCs (among ~5000 cells inspected) were detected using F1[CBA 3 B6] and 129/SvJ mice (7, 8), IDO+ cells were when >50 lg mlÿ1 anti-IFNa mAb was present (Fig. 3F, data not dispersed in splenic red pulp areas of control BALB/c mice shown). These data support the hypothesis that IFNa is an (Fig. 3G). However, no IDO+ cells were detected in spleen of essential intermediary signaling ligand that activates STAT1- IFNabR-KO mice (Fig. 3H). Since STAT1 activation (Fig. 2A–C) mediated IDO up-regulation in DCs following B7 ligation. These and IDO expression (8) following B7 ligation occurred findings also suggested that splenocytes were induced to normally in mice deficient for IFNcR expression, these data express IFNa following B7 ligation in vitro. Since we used suggested that IFNa signaling, but not IFNc signaling, was MACS-enriched CD11c+ DCs, these data suggested that DCs essential for early activation of STAT1 and subsequent IDO up- might be the source of IFNa, though MACS enrichment did not regulation in minor populations of splenocytes. In addition, the completely remove other (CD11cNEG) splenocytes, which might pattern of IDO expression induced in spleen 24 h after in vivo be a source of IFNa. CTLA4-Ig treatment (Fig. 3G) was reminiscent of the pattern of STAT1 activation observed at earlier times (Fig. 2), suggesting that selective activation of STAT1 preceded IDO expression in B7 ligation induces CD19+ DCs to express and the same minor population of cells located in splenic red pulp. secrete IFNa Though cells that responded to B7 ligation were present in To identify cells that produced IFNa following B7 ligation, we genetically manipulated mice lacking IFNcRs, it is possible measured IFNa gene and protein expression by splenocytes. that these cells might not develop in mice lacking IFNabRs. To First, we assessed IFNa gene transcription by RT–PCR address this alternative explanation for failure to activate analysis and IFNa secretion by ELISA following CTLA4-Ig STAT1 and induce IDO expression in IFNabR-KO mice, we treatment in vitro (Fig. 4). Transcripts of the IFNa1–9 genes employed a complementary approach to test the hypothesis were detected in RNA samples prepared from CTLA4-Ig- that IFN type I, but not type II, signaling was essential for treated and influenza virus-infected DCs (Fig. 4A). IFNa1–9
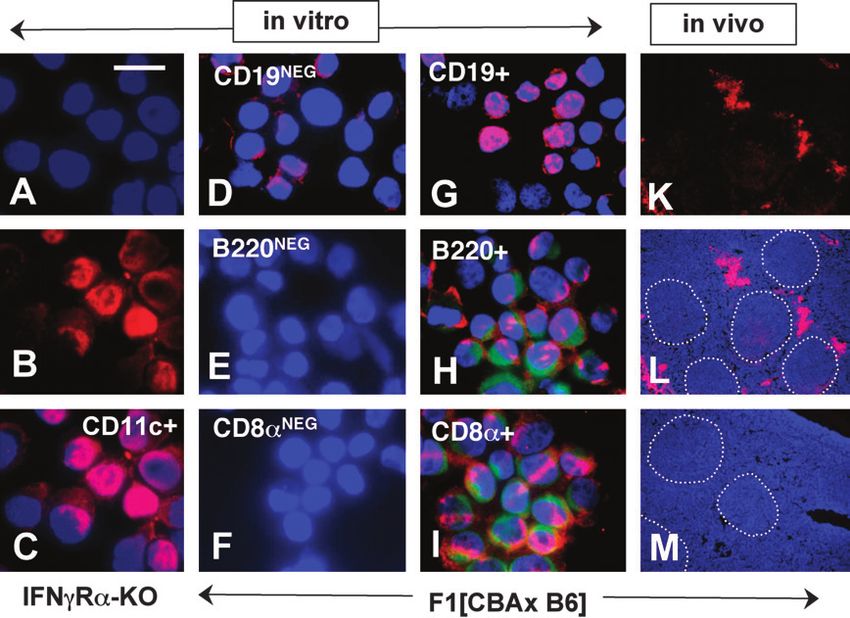
IDO-expressing CD19+ regulatory dendritic cells 915 Fig. 3. B7 ligation induces STAT1 activation and IDO expression via IFN type 1 signaling. (A–C) AutoMACS-enriched CD11c+ DCs from mice indicated were incubated with non-mutant CTLA4-Ig (100 lg mlÿ1) for 5 h and stained with anti-P-STAT1 antibody (red) and Hoechst dye (blue) as in Fig. 2. (D–F) Splenic DCs from BALB/c mice were incubated with non-mutant CTLA4-Ig with no additions (D), 50 lg mlÿ1 anti-IFNc (E) and anti- IFNa (F) antibody and stained to detect nuclear P-STAT1. (G and H) Spleen tissue sections were prepared from mice indicated 24 h after injection with non-mutant CTLA4-Ig (100 lg). Tissues were stained using an anti-IDO antibody (Methods). Original magnifications: 31000 (A–C), 3400 (D–F) and 3100 (G and H). transcripts were not detected in RNA samples prepared from expression induced by STAT1-dependent IFNa signaling were untreated DCs. Consistent with this, DCs secreted IFNa into both confined to the CD19+ DC population. culture media following treatment with non-mutant CTLA4-Ig, while DCs treated with the mutant CTLA4-Ig isotype, which did not induce IDO in DCs (8), did not secrete IFNa (Fig. 4B). IFNa, but not IFNc, promotes IDO-dependent T cell Based on the finding that B7 ligation induced DCs to suppression express IFNa, we hypothesized that IFNa production was Based on data showing that IFNa signaling, but not IFNc restricted to CD19+ DCs following B7 ligation, like STAT1 signaling, was essential for STAT1 activation and IDO up- activation and functional IDO up-regulation (Figs 1 and 2). To regulation in splenic DCs (Figs 3 and 4), we hypothesized that test this hypothesis, we treated purified (Mo-Flo sorted) IFNa produced by CD19+ DCs following B7 ligation signaled CD19+ and CD19NEG (CD11cHIGH) DCs with CTLA4-Ig for CD19+ DCs to acquire potent IDO-dependent T cell regulatory 5 h and stained them with anti-IFNa mAb (Fig. 4C, upper functions. To test this hypothesis, we asked if recombinant panels). Almost all CD19+ DCs contained cytoplasmic IFNa, IFNa could substitute for in vivo CTLA4-Ig treatment as while very few cells expressing IFNa were detected in a stimulus to induce IDO-dependent T cell suppression. Using CD19NEG DCs. To test if IDO expression was also restricted to the experimental system described in Fig. 1, we performed CD19+ DCs following B7 ligation, we cultured Mo-Flo-sorted MLRs using splenocytes from untreated F1[CBA 3 B6] mice DCs for 24 h in the presence of CTLA4-Ig and stained cells for as stimulators and assessed the effect of adding recombinant IDO expression. As expected from functional data reported in IFNa (Fig. 5A) and IFNc (Fig. 5B) on their ability to stimulate Fig. 1(C), IDO+ cells were detected exclusively in the CD19+ BM3 T cell proliferation. T cell proliferation was reduced DC population (Fig. 4C, center panels). IDO was also selec- significantly in cultures containing >150 U mlÿ1 IFNa. This tively induced in the same CD19+ DC population when recom- anti-proliferative effect of IFNa was due to induction of IDO and binant IFNa was added to culture medium (Fig. 4C, lower not an intrinsic anti-proliferative effect of IFNa because T cell panels). These data showed that B7 ligation and IFNa induction proliferative responses recovered to control levels in the were preferentially confined to DC populations expressing presence of the IDO inhibitor (1mT). Also consistent with the CD19. IDO staining after 24 h was more heterogeneous than hypothesis that IFNa signaled IDO induction, addition of IFNa IFNa staining after 5 h in CD19+ DCs, suggesting that the ability to MLRs containing splenocytes from IDO-KO mice had no to express high levels of IDO may not be uniformly distributed effect on their ability to promote T cell proliferation. within the CD19+ DC population. However, these data show In contrast to outcomes obtained with IFNa, addition that IFNa secretion induced following B7 ligation and IDO of recombinant IFNc did not suppress T cell proliferation
916 IDO-expressing CD19+ regulatory dendritic cells
Fig. 4. B7 ligation induces CD19+ DCs to secrete IFNa, which induces IDO expression. (A) RT–PCR analyses of RNA from MACS-enriched DCs
infected with influenza virus (flu), exposed to PBS (ÿ) or non-mutant CTLA4-Ig (+, 100 lg mlÿ1) for 5 h. (B) A total of 106 MACS-enriched DCs were
cultured with mutant or non-mutant CTLA4-Ig for 5 h and culture media analyzed for IFNa by ELISA. (C) Mo-Flo-sorted (>99% CD11c+) CD19NEG
and CD19+ DCs were incubated with non-mutant CTLA4-Ig and stained with anti-IFNa antibody (upper panels) or anti-IDO antibody (center
panels) after 5 or 24 h, respectively. Sorted CD19+ DCs were incubated with or without IFNa for 24 h and stained with anti-IDO antibody as
indicated (lower panels).
(Fig. 5B), and addition of IDO inhibitor did not enhance T cell splenic CD19+ DCs are unique in their ability to produce IFNa
responses when recombinant IFNc or no exogenous IFN was and respond to IFNa-mediated signaling by acquiring potent
added to MLRs (data not shown). These data revealed that T cell regulatory functions via STAT1 activation and IDO up-
IFNa induced IDO-dependent T cell suppression, while IFNc regulation in response to B7 ligation.
had no effect in this system. The rationale for studying CD19+ DCs in spleen was based
To examine if IFNa acted to induce IDO-dependent T cell on our previous discovery that CD19+ DCs constituted the
suppression selectively in the CD19+ DC population, we principal cell population that mediated IDO-dependent T cell
repeated the previous experiment using purified CD19+ and suppression in TDLNs (11). CD19 is a component of signaling
CD19NEG DC populations sorted by flow cytometry (Fig. 5C complexes expressed by B cells, and has been widely used to
and D, respectively). When CD19+ DCs were used as APCs, separate B cells from DCs; partly for this reason, splenic
addition of recombinant IFNa to MLRs induced potent IDO- CD19+ DCs may not have been recognized previously.
dependent T cell suppression, which was reversed in the CD19+ DCs from TDLNs shared certain characteristics with
presence of 1mT or excess tryptophan in MLRs. In contrast, the B cell lineage, including D–J region Ig gene rearrange-
addition of recombinant IFNc had no significant effect on T cell ments and expression of B220 and Pax5 (11). Similar links to
proliferation (Fig. 5C). Moreover, neither IFNa nor IFNc the B cell lineage have been reported in plasmacytoid DC
addition had any effect on the robust T cell stimulatory activity subsets from other studies (20). In spleens of F1[CBA 3 B6]
of CD19NEG DCs (Fig. 5D). These outcomes confirmed that mice, we found that CD19+ DCs constituted ~20% of sorted
IFNa was the relevant upstream signaling ligand that induced splenic DCs expressing relatively high levels of CD11c and,
functional IDO expression in CD19+ DC populations since like CD19+ DCs from TDLNs of B6 mice, these cells co-
IFNa substituted for B7 ligation in promoting IDO-dependent expressed B220 and many also expressed CD8a. Murine
T cell regulatory functions of CD19+ DCs. plasmacytoid DCs have been reported to express B220 and
120G8 and low/intermediate levels of CD11c, display imma-
ture phenotypes with respect to MHC and B7 expression
Discussion
levels and have relatively weak T cell stimulatory functions
In the current study, we identified a small population of splenic associated with T cell suppressive and tolerogenic outcomes
CD19+ DCs as the principal cell type that mediated IDO- (17, 18, 21–23). In the current study, sorted CD19+ DCs that
dependent T cell suppression following CTLA4-Ig treatment mediated IDO-dependent T cell suppression expressed
in vivo. CD19+ DCs selectively responded to B7 ligation by relatively high levels of CD11c, had mature phenotypes and
secreting IFNa and activating STAT1. Since IFNa, but not IFNc, were potent T cell stimulators. However, when IDO activity was
could substitute for B7 ligation to promote IDO-dependent induced following in vivo CTLA4-Ig, or in vitro IFNa, treatment
T cell suppression, these data support the hypothesis that these DCs became strongly suppressive. Hence, CD19IDO-expressing CD19+ regulatory dendritic cells 917 Fig. 5. IFNa, but not IFNc, signals IDO-dependent T cell suppression in CD19+ DCs. (A and B) Total splenocytes from F1[CBA 3 B6] (closed symbols) or IDO-KO mice (open symbols) were used to stimulate BM3 T cells in the presence of recombinant IFNa (A) or IFNc (B). T cell proliferation was assessed after 72 h (squares), as in Fig. 1(C). IDO inhibitor, 1mT (100 lM), was added to parallel MLRs (circles). Asterisks indicate significant reduction in T cell proliferation (P < 0.01). (C and D) As above, except that Mo-Flo-sorted CD19+ (C) and CD19NEG (D) DCs were used as APCs. Parallel MLRs contained no additions (black bars), 1mT (white bars) or excess tryptophan (striped bars) with or without recombinant IFNa or IFNc, as indicated. expression appears to identify the population of DCs that can Though we identified CD19+ DCs as the principal DC be induced to acquire potent T cell regulatory functions via population that mediated functional T cell suppression in the IDO. These DCs appear distinct from typical plasmacytoid present study, other DCs might also express non-functional DCs defined previously by others, although they share certain immunoreactive IDO protein following B7 ligation, or other features, such as B220 expression and the ability to produce treatments. Consistent with this, the proportion of MACS- IFNa (8, 11, 18). Hence, CD19 may not be a distinct DC enriched (CD11c+) DCs containing activated intra-nuclear lineage marker but rather identifies these DC populations P-STAT1 (~30–50%) following B7 ligation was higher than the with particular functional characteristics, including the ability proportion of CD19+ DCs (~10%). Previously, we detected to respond to B7 ligation by up-regulating IDO. IDO expression in several different DC subsets, including DCs The responsiveness of DCs to CTLA4-Ig isotypes may differ co-expressing CD8a, B220 and the NK-DC marker DX5, all of between mouse strains. Grohmann and colleagues reported which expressed immunoreactive IDO protein by immunohis- that a different CTLA4-Ig isotype (CTLA4-IgG3) partially tochemistry after B7 ligation in vivo (7). However, these earlier blocked T cell-mediated rejection of pancreatic islet allografts studies did not include assays to measure T cell stimulatory transplanted into B6 mice, and showed that this reagent functions of sorted DC subsets. It is known that IDO can be induced functional IDO expression in isolated B6 DCs (6). expressed in non-functional form in both murine and human With our CTLA4-Ig preparation (CTLA4-IgG2a) we found DC subsets (3, 24). Thus, the functional analyses of T cell that DCs from B6 mice were unresponsive, while DCs from stimulatory functions performed in the current study were CBA, BALB/c and 129/SvJ mice responded by up-regulating critically important in identifying biologically relevant popula- IDO (our unpublished data). Thus, the CTLA4-Ig reagent we tions of IDO-expressing DCs. used may have failed to induce IDO in B6 mice for technical Several recent reports revealed that IDO enzyme activity in reasons, perhaps related to the Fc domain structure. In this DCs has potent inhibitory effects on T cell responses in vitro regard, it may be important that a mutant isotype of CTLA4- and in vivo (5, 7, 8, 11, 25). In mice, IDO expression was first IgG2a, engineered to reduce complement factor C1q and associated with CD8a+ DCs in response to IFNc treatment FcR binding, also failed to induce IDO. Since CD19+ DCs (5, 24). More recently, we identified B220+ DCs in spleen and constituted the principal cell subset mediating IDO-mediated TDLNs as potent mediators of IDO-dependent T cell suppres- suppression in tumor-bearing B6 mice (11), and CD19+ DCs sion (8, 11). B220+ and CD8a+ DC subsets may overlap to were detected in comparable proportions in CBA, 129/SvJ some extent as CD8a is expressed by some plasmacytoid and F1[CBA 3 B6] mice (our unpublished data), it is likely DCs (20–22), as discussed above. However, in our system, the that the presence of CD19+ DCs is not strain dependent. CD19 marker gave the best segregation of IDO-dependent
918 IDO-expressing CD19+ regulatory dendritic cells
T cell suppressor functions, STAT1 activation and IFNa pro- Abbreviations
duction in distinct populations of splenic DCs. APC antigen-presenting cell
The role of IFNa in our system was unexpected. Plasmacy- CTLA4-Ig soluble synthetic CTLA4 fusion protein
toid DCs are known to produce IFNa in response to microbial DC dendritic cell
infections (17), most likely via signals transmitted through Toll- FBS fetal bovine serum
IDO indoleamine 2,3-dioxygenase
like receptors, but IFNc is known to be a more potent IDO IRF IFN regulatory factor
inducer than IFNa (19). However, we found that IFNa signaling KO gene deficient
was required to induce IDO expression in CD19+ DCs and that 1mT 1-methyl-D-tryptophan
recombinant IFNa could re-capitulate the response to B7 MLR mixed lymphocyte reaction
ligation, leading to IDO-dependent T cell suppression. P-STAT1 Phospho-(Y701)-STAT1
RT room temperature
The unique signaling processes that confer the highly RT–PCR reverse transcription–polymerase chain reaction
selective link between B7 ligation and IDO induction in distinct STAT1 signal transducer and activator of transcription
DC populations are not fully defined. Grohmann and TBS Tris-buffered saline
colleagues showed that IFNc was an essential upstream TDLN tumor-draining lymph node
TdR thymidine
ligand required for IDO induction in unfractionated splenic
CD11c+ DCs following CTLA4-Ig treatment in vitro (6).
However, we developed different experimental approaches
to address the specific question of which DC populations were References
principally responsible for IDO-mediated suppression when 1 Mellor, A. L. and Munn, D. H. 2004. IDO expression in dendritic
DCs were exposed to CTLA4-Ig in vivo. We found that IFNc cells: tolerance and tryptophan catabolism. Nat. Rev. Immunol.
signaling was not essential for this process, while IFNa 4:762.
2 Seo, S. K., Choi, J. H., Kim, Y. H. et al. 2004. 4-1BB-mediated
signaling was essential. STAT1 activation appears to be an immunotherapy of rheumatoid arthritis. Nat. Med. 10:1088.
obligate event preceding IDO expression since IDO was not 3 Munn, D. H., Sharma, M. D., Lee, J. R. et al. 2002. Potential
induced in STAT1-deficient mice (6, 26). Previous reports have regulatory function of human dendritic cells expressing indole-
also shown that IFNa can induce IDO expression via STAT1- amine 2,3-dioxygenase. Science 297:1867.
dependent signaling, though IFNa is considerably less potent 4 Grohmann, U., Fallarino, F. and Puccetti, P. 2003. Tolerance, DCs
and tryptophan: much ado about IDO. Trends Immunol. 24:242.
as an IDO inducer than IFNc in most cell types studied (9, 19). 5 Grohmann, U., Fallarino, F., Silla, S. et al. 2001. CD40 ligation
Our findings are consistent with the hypothesis that STAT1 ablates the tolerogenic potential of lymphoid dendritic cells.
activation is a selective response to IFNa by specific DCs, J. Immunol. 166:277.
including minor DC populations expressing CD19. 6 Grohmann, U., Orabona, C., Fallarino, F. et al. 2002. CTLA-4-Ig
regulates tryptophan catabolism in vivo. Nat. Immunol. 3:1097.
Mechanisms that confer selective IDO expression exclu- 7 Mellor, A. L., Baban, B., Chandler, P. et al. 2003. Cutting edge:
sively in CD19+ DCs have not been defined. Presumably, induced indoleamine 2,3 dioxygenase expression in dendritic cell
selective induction of IFNa expression following B7 ligation is subsets suppresses T cell clonal expansion. J. Immunol. 171:1652.
controlled by factors in DCs that modulate downstream 8 Mellor, A. L., Chandler, P. R., Baban, B. et al. 2004. Specific
signals generated following B7 ligation, though the nature of subsets of murine dendritic cells acquire potent T cell regulatory
functions following CTLA4-mediated induction of indoleamine 2,3
these mechanisms is not known. Similarly, selective IFNa- dioxygenase. Int. Immunol. 16:1391.
mediated STAT1 activation in CD19+ DCs is probably con- 9 Taylor, M. W. and Feng, G. 1991. Relationship between interferon-c,
trolled by factors downstream of IFNabRs, since many cell indoleamine 2,3-dioxygenase, and tryptophan catabolism.
types express these receptors. One speculative possibility is FASEB J. 5:2516.
10 Levy, D. E. and Darnell, J. E., Jr. 2002. Stats: transcriptional control
that IFN regulatory factors (IRFs), such as IRF-2 and IRF-7, and biological impact. Nat. Rev. Mol. Cell Biol. 3:651.
which are differentially expressed in distinct DC populations, 11 Munn, D. H., Sharma, M. D., Hou, D. et al. 2004. Expression of
might regulate responses to IFNs differently in distinct DC indoleamine 2,3-dioxygenase by plasmacytoid dendritic cells in
subsets (10, 27, 28). Elucidating these signaling mechanisms tumor-draining lymph nodes. J. Clin. Invest. 114:280.
will be critical for understanding why CD19+ DCs selectively 12 Tarazona, R., Sponaas, A.-M., Mavria, G. et al. 1996. Effects of
different antigenic microenvironments on the course of CD8+ T cell
produce IFNa in response to B7 ligation and express IDO in responses in vivo. Int. Immunol. 8:351.
response to IFNa, while most DCs do not respond in this way, 13 Baban, B., Chandler, P., McCool, D., Marshall, B., Munn, D. H. and
even though they express B7 molecules. The key point to Mellor, A. L. 2004. Indoleamine 2,3-dioxygenase expression is
emerge from the current study, however, is that certain minor restricted to fetal trophoblast giant cells during murine gestation
and is maternal genome specific. J. Reprod. Immunol. 61:67.
populations of splenic DCs, best identified by the expression 14 Simpson, E. and Chandler, P. 1986. Analysis of cytotoxic T cell
of CD19 in our system, are selectively programmed to respond responses. In Weir, D. M., Herzenberg, L. A. and Herzenberg,
to B7 ligation by inducing IDO, and acquiring potent T cell L. A., eds, Handbook of Experimental Immunology, p. 68. Black-
regulatory functions as a consequence. well Scientific, Oxford.
15 Julius, M. H., Simpson, E. and Herzenberg, L. A. 1973. A rapid
method for the isolation of functional thymus-derived murine
Acknowledgements lymphocytes. Eur. J. Immunol. 3:645.
16 Fung, M. C., Sia, S. F., Leung, K. N. and Mak, N. K. 2004. Detection
We thank the manager of the MCG Flow Cytometry Core facility, of differential expression of mouse interferon-alpha subtypes by
Jeanine Pihkala, for expert assistance with flow cytometry and Anita polymerase chain reaction using specific primers. J. Immunol.
Wylds, Doris McCool and Erika Thompson for technical assistance Methods 284:177.
with multiple aspects of studies reported here. This work was 17 Asselin-Paturel, C., Boonstra, A., Dalod, M. et al. 2001. Mouse type
supported by NIH grants to A.L.M. (HD41187, AI063402) and I IFN-producing cells are immature APCs with plasmacytoid
D.H.M. (CA103320, CA096651). morphology. Nat. Immunol. 2:1144.IDO-expressing CD19+ regulatory dendritic cells 919
18 Asselin-Paturel, C., Brizard, G., Pin, J. J., Briere, F. and Trinchieri, differentiation and support function of T cells with regulatory
G. 2003. Mouse strain differences in plasmacytoid dendritic cell properties. Immunology 108:481.
frequency and function revealed by a novel monoclonal antibody. 24 Fallarino, F., Vacca, C., Orabona, C. et al. 2002. Functional ex-
J. Immunol. 171:6466. pression of indoleamine 2,3-dioxygenase by murine CD8alpha(+)
19 Hassanain, H. H., Chon, S. Y. and Gupta, S. L. 1993. Differential dendritic cells. Int. Immunol. 14:65.
regulation of human indoleamine 2,3-dioxygenase gene expres- 25 Fallarino, F., Asselin-Paturel, C., Vacca, C. et al. 2004. Murine
sion by interferons-gamma and -alpha. Analysis of the regulatory plasmacytoid dendritic cells initiate the immunosuppressive
region of the gene and identification of an interferon-gamma- pathway of tryptophan catabolism in response to CD200 receptor
inducible DNA-binding factor. J. Biol. Chem. 268:5077. engagement. J. Immunol. 173:3748.
20 Corcoran, L., Ferrero, I., Vremec, D. et al. 2003. The lymphoid past 26 Gurtner, G. J., Newberry, R. D., Schloemann, S. R., McDonald,
of mouse plasmacytoid cells and thymic dendritic cells.
K. G. and Stenson, W. F. 2003. Inhibition of indoleamine 2,3-
J. Immunol. 170:4926.
dioxygenase augments trinitrobenzene sulfonic acid colitis in
21 Martin, P., Del Hoyo, G. M., Anjuere, F. et al. 2002. Characterization
of a new subpopulation of mouse CD8alpha+ B220+ dendritic mice. Gastroenterology 125:1762.
cells endowed with type 1 interferon production capacity and 27 Honda, K., Yanai, H., Mizutani, T. et al. 2004. Role of a trans-
tolerogenic potential. Blood 100:383. ductional-transcriptional processor complex involving MyD88 and
22 O’Keeffe, M., Hochrein, H., Vremec, D. et al. 2002. Mouse IRF-7 in Toll-like receptor signaling. Proc. Natl Acad. Sci. USA
plasmacytoid cells: long-lived cells, heterogeneous in surface 101:15416.
phenotype and function, that differentiate into CD8(+) dendritic 28 Honda, K., Mizutani, T. and Taniguchi, T. 2004. Negative regulation
cells only after microbial stimulus. J. Exp. Med. 196:1307. of IFN-alpha/beta signaling by IFN regulatory factor 2 for
23 Bilsborough, J., George, T. C., Norment, A. and Viney, J. L. 2003. homeostatic development of dendritic cells. Proc. Natl Acad. Sci.
Mucosal CD8alpha+ DC, with a plasmacytoid phenotype, induce USA 101:2416.You can also read

















































