Report Genome-wide Association of Yorkie with Chromatin and Chromatin-Remodeling Complexes
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Cell Reports
Report
Genome-wide Association of Yorkie
with Chromatin and Chromatin-Remodeling Complexes
Hyangyee Oh,1 Matthew Slattery,2,3 Lijia Ma,3 Alex Crofts,3 Kevin P. White,3 Richard S. Mann,2 and Kenneth D. Irvine1,*
1Howard Hughes Medical Institute, Waksman Institute, and Department of Molecular Biology and Biochemistry, Rutgers University,
Piscataway, NJ 08854, USA
2Department of Biochemistry and Molecular Biophysics, Columbia University, 701 West 168th Street, HHSC 1104, New York, NY 10032, USA
3Institute for Genomics and Systems Biology and Department of Human Genetics, University of Chicago, 900 East 57th Street, KCBD 10115,
Chicago, IL 60637, USA
*Correspondence: irvine@waksman.rutgers.edu
http://dx.doi.org/10.1016/j.celrep.2013.01.008
SUMMARY to elevate the transcription of target genes, but the mechanism(s)
by which it does so is not understood.
The Hippo pathway regulates growth through the Two key aspects of transcriptional activation in eukaryotes
transcriptional coactivator Yorkie, but how Yorkie are modulation of chromatin structure and recruitment of the
promotes transcription remains poorly understood. core transcriptional machinery. Chromatin modifiers include
We address this by characterizing Yorkie’s associa- ATP-dependent chromatin-remodeling complexes, such as
tion with chromatin and by identifying nuclear part- NURF or SWI/SNF, and histone-modifying enzymes, which
modulate the properties of nucleosomes through posttransla-
ners that effect transcriptional activation. Coimmu-
tional modifications (Li et al., 2007). Recruitment of the core
noprecipitation and mass spectrometry identify transcriptional machinery can involve direct interactions with
GAGA factor (GAF), the Brahma complex, and the core components or interactions with components of a large
Mediator complex as Yorkie-associated nuclear complex called Mediator, which links transcriptional activators
protein complexes. All three are required for Yorkie’s to core subunits of RNA polymerase (Malik and Roeder,
transcriptional activation of downstream genes, and 2010). Chromatin modification and recruitment of core tran-
GAF and the Brahma complex subunit Moira interact scriptional machinery are often thought of as distinct pro-
directly with Yorkie. Genome-wide chromatin- cesses, but they can be mechanistically linked. One transcrip-
binding experiments identify thousands of Yorkie tion factor associated with both processes is GAGA factor
sites, most of which are associated with elevated (GAF, encoded by the Trithorax-like locus, Trl). GAF has been
transcription, based on genome-wide analysis of reported to associate with components of the core transcrip-
tional machinery (Chopra et al., 2008) but was also identified
messenger RNA and histone H3K4Me3 modification.
as a gene required for the normal expression of homeotic
Chromatin binding also supports extensive func- genes, and in this role it is thought to act by influencing
tional overlap between Yorkie and GAF. Our studies chromatin structure (Farkas et al., 1994). Here, we identify
suggest a widespread role for Yorkie as a regulator a direct physical link between Yki and proteins involved in chro-
of transcription and identify recruitment of the chro- matin remodeling, providing insights into how Yki activates
matin-modifying GAF protein and BRM complex as transcription.
a molecular mechanism for transcriptional activation
by Yorkie. RESULTS
Widespread Localization of Yki on Chromatin
INTRODUCTION Although several genes directly regulated by Yki have been
identified, the full constellation of Yki targets is unknown. We
The Hippo pathway is essential for normal development and is identified Yki target genes by chromatin immunoprecipitation
dysregulated in many cancers, reflecting its important role in (ChIP) using Yki antisera and DNA sequencing (ChIP-seq). This
growth control (Zhao et al., 2011). Hippo signaling is regulated identifies candidate direct targets of Yki, whereas most genes
by diverse upstream inputs, which converge on the protein whose transcription is altered by changes in Yki activity might
kinases Hippo and Warts (Wts). Downstream outputs of Hippo be indirectly affected. Yki-associated chromatin was isolated
signaling in Drosophila are mediated through the oncogenic and analyzed from both 8–16 hr embryos and third-instar wing
transcriptional coactivator Yorkie (Yki), which is negatively regu- discs. This identified a large number of loci with significant Yki
lated by Wts (Oh and Irvine, 2010). As a transcriptional coacti- association: 6,491 Yki-bound regions in wing disc chromatin
vator, Yki does not bind DNA directly, but instead interacts and 3,749 in embryonic chromatin (corresponding to an esti-
with DNA-binding proteins, utilizing multiple DNA-binding part- mated 3,899 and 2,416 genes, respectively, when peaks within
ners (Oh and Irvine, 2010). Once recruited to DNA, Yki’s role is 500 bp of the transcription start site or within the transcription
Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors 309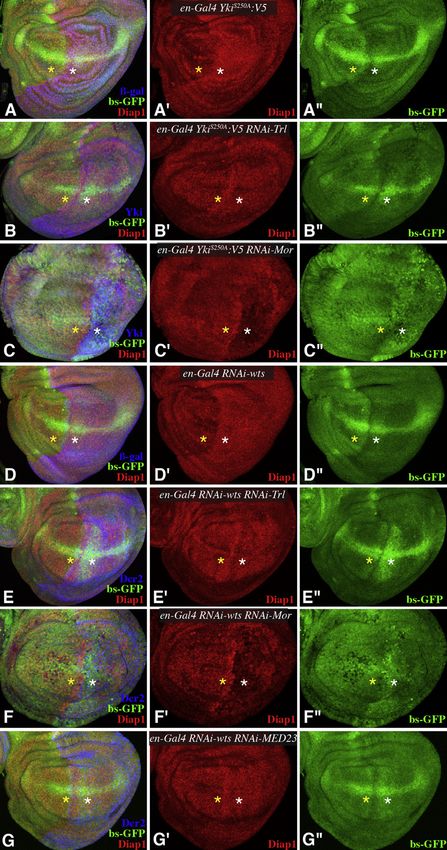
Figure 1. Localization of Yki and GAF on Chromosomes in Wing Discs
(A) Comparison of the percentage DNA near promoters (within 3 kb upstream of a transcription start site), in introns, in exons, and in intergenic regions, within the
whole genome, and within the Yki-bound fraction.
(B) Comparison of the distribution of messenger RNA levels (blue) and proximal promoter (within 100 bp of transcription start) H3K4me3 modification (yellow)
between Yki target genes and non-Yki targets. Units of expression for RNA-seq are fragments per kilobase of transcript per million mapped reads (FPKM).
H3K4me3 wing ChIP-seq data are from Pérez-Lluch et al. (2011).
(C) Shown is the overlap between Yki- and GAF-binding sites, where numbers indicate the numbers of peaks. Numbers in the overlap differ because one peak for
one protein can overlap two peaks of the other.
(D) Plot of ChIP peaks at three loci regulated by Yki. Transcription units of targets are in purple, and transcription units of neighboring genes are in black.
Regions called Yki peaks are identified by gray bars. Yki-responsive enhancers that have been identified at these loci (Oh and Irvine, 2011; Wu et al.,
2008; Zhang et al., 2008a) are indicated in orange at the bottom, and a previously identified Hth- and Yki-binding region (Peng et al., 2009) is indicated in
light blue.
(legend continued on next page)
310 Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors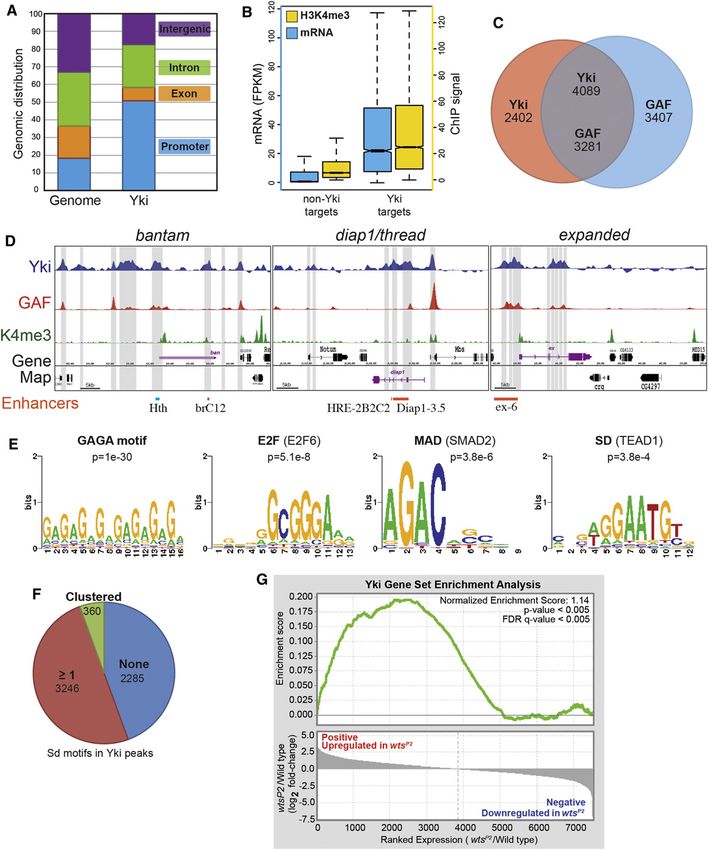
unit are assigned to that gene). Gene ontology analysis suggests Correlation between Yki and GAF Binding
that these genes are linked to a broad range of functions We used de novo motif discovery to find DNA sequence motifs
(Table S1). that are enriched within Yki-bound chromatin. The most en-
Genome annotation revealed that Yki-bound regions are riched motifs comprise GAGA-rich sequences (Figures 1E and
enriched near promoters as compared to intronic, exonic, and S1E; Table S2), which correspond to the DNA recognition motif
intergenic regions (Figure 1A; Figure S1A), consistent with of GAF, which plays crucial roles at multiple steps of transcrip-
Yki’s role as a transcriptional activator. Comparison of transcript tion (Adkins et al., 2006). Earlier studies have implicated Sd,
levels identified a correlation between transcription and Yki Hth, and Mad in the recruitment of Yki to DNA (Oh and Irvine,
binding, as genes with Yki-binding sites were expressed on 2010), and another DNA-binding transcription factor, E2F1,
average at levels over 10-fold higher than genes without Yki- synergizes with Yki to regulate cell-cycle genes (Nicolay
binding sites in wing discs and at roughly 7-fold higher levels et al., 2011). Binding sites for these proteins did not emerge
in embryos (Figures 1B and S1B). Genomic analysis also from de novo motif analysis, but scanning against a database
revealed a correlation between Yki-bound chromatin and peaks of known DNA motifs identified SMAD2 (Mad homolog)-binding
of H3K4me3 modification. Across the genome, the average and TEAD1 (Sd homolog)-binding motifs, and multiple E2F
H3K4me3 ChIP signal near promoters, which is associated motifs, as enriched near the centers of Yki-binding regions (Fig-
with actively transcribed genes (Eissenberg and Shilatifard, ure 1E; Table S2). E2F-related motifs were also overrepre-
2010), was several-fold higher for genes with Yki sites than for sented near the centers of Yki embryo peaks (Figure S1E; Table
genes without Yki sites (Figures 1B and S1B). These obser- S2), and a subset of Yki embryo peaks were enriched for TALE
vations indicate that Yki binding to chromatin is correlated (Hth) motifs (Figure S1E; Table S2). Moreover, when we
with increased transcription, and thus imply that Yki acts as analyzed Yki-bound regions with a more degenerate Sd-
a broad regulator of transcriptional activity at thousands of loci binding motif, 50%–60% of Yki peaks contained one or more
throughout the genome. potential Sd-binding sites (Figures 1F and S1F). These results
As a further test of this, we conducted expression analysis support a model in which Yki is recruited to DNA by multiple
by RNA sequencing (RNA-seq) on wing discs isolated from factors.
wild-type and wtsP2 mutant larvae (Justice et al., 1995). Com- To address the significance of GAGA-rich sequences, we
parison of the relative expression levels of genes associated used a GAF antisera to perform ChIP-seq experiments and
with Yki peaks to those not associated with Yki peaks revealed identified GAF-bound chromatin in both 8–16 hr embryos and
that they differed by over 30% at the median expression level. wing discs. A highly significant overlap was identified: over
Among non-Yki-bound genes, the average expression was 60% of Yki sites overlap GAF sites (Figures 1C and S1C),
scored as having decreased, which we think reflects nor- including peaks at known target genes (Figures 1D and S1D).
malization to total genome-wide expression, which should be This suggests that GAF is a frequent partner of Yki throughout
increased if most of the 3,899 Yki-bound genes are affected by the genome for transcriptional regulation of downstream
wtsP2. To provide a statistical test of the relation between Yki genes.
binding and transcriptional changes in wtsP2, we used gene set
enrichment analysis (GSEA) (Subramanian et al., 2005). Indeed, Identification of GAF as a Nuclear Yki-Associated
genes with increased expression in wtsP2 were highly enriched Protein
for Yki target genes as determined by ChIP-seq (p < 0.005) (Fig- To define mechanisms by which Yki activates transcription, we
ure 1G; Table S4). sought to identify nuclear Yki-associated proteins. Yki is nor-
We also examined Yki binding at previously characterized mally predominantly cytoplasmic, reflecting endogenous Hippo
targets of Yki. At three genes for which Yki-responsive pathway activity (Dong et al., 2007; Oh and Irvine, 2008). To
enhancers have been characterized, ban (Oh and Irvine, 2011), avoid cytoplasmic Yki-associated proteins, we isolated nuclear
Diap1 (Wu et al., 2008; Zhang et al., 2008a), and ex (see below), extracts from Drosophila S2 cells and then purified endogenous
Yki-binding peaks in wing discs overlap functionally defined Yki- Yki complexes using anti-Yki serum or, as a control, preim-
response elements (Figure 1D). These genes were all identified mune serum, attached to beads. Coimmunoprecipitated pro-
as Yki targets in imaginal discs, and Yki binding at these loci teins, visualized by silver staining on SDS-PAGE gels, that
was more extensive in wing discs than in embryos (Figures 1D were precipitated using anti-Yki and not preimmune sera (Fig-
and S1D). ure 2A) were then identified by mass spectrometry. This
(E) Motifs and significance scores for DNA-binding proteins at Yki-bound chromatin. De novo motif analysis identified variants on the GAF-binding motif (GAGA)
as the highest scoring. SMAD2 (Mad) and E2F-binding motifs are enriched within all Yki-binding regions, whereas TEAD1 (Sd) motifs are most enriched within
Yki-binding regions without a GAF-binding site.
(F) Analysis of DNA within Yki-bound wing peaks using a degenerate Sd-binding motif consistent with the published literature (Halder and Carroll, 2001): [AT][AG]
[AG]AAT[GT][CT]. A total of 2,285 regions do not contain a Sd motif, and the remainder do. A total of 360 regions contain clustered Sd motifs (two motifs separated
by 20 or fewer base pairs) and 3,246 regions contain one or more isolated motifs.
(G) GSEA of the correlation between relative expression changes and Yki binding. Genes are ordered according to their relative fold change between wild-type
and wtsP2. Log2-transformed fold-change values for expressed genes are plotted in the bottom graph (see Experimental Procedures). The GSEA running
enrichment score is represented in the top graph (green line). Genes that increase in wtsP2 are significantly enriched for Yki target genes (p < 0.005; FDR q-value <
0.005); genes that decrease in wtsP2 are not enriched for Yki targets.
See also Figure S1.
Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors 311Figure 2. Yki Associates with GAF and Mor
(A) Silver-stained gel displaying proteins precipi-
tated from S2 cell nuclear extracts by Yki anti-
bodies or preimmune serum, as indicated. Red,
green, and blue asterisks indicate positions of Yki,
GAF, and Mor bands, as determined by immuno-
blotting or mass spectrometry.
(B and C) Western blots on S2 cell nuclear
extract (input) or proteins immunoprecipitated
from S2 cell nuclear extract by Yki antibodies
or preimmune serum and detected by anti-Yki
(top), anti-GAF (B, bottom), or anti-Mor (C,
bottom). Nuclear Yki was barely detectible in the
input lanes but was enriched by immunopur-
ification, and is normally detected as a doublet.
Three bands of GAF were detected using GAF
antibody.
(D and E) Western blots showing coimmuno-
precipitation of Yki3SA:Flag or Yki-WW3SA:Flag
with GAF:V5 (D) or Mor:V5 (E) from S2 cell
nuclear extracts (GFP:Flag is a negative con-
trol). Upper panels (Input) show blots on nuclear
lysates, and lower panels (V5-IP) show blots
(anti-Flag) on material precipitated by anti-V5
beads.
(F) Western blot showing results of GST pull-down
assays. GST:GAF:V5, GST:Mor:V5, or GST were
immobilized on glutathione-agarose beads and
incubated with bacterially-expressed GFP:V5,
Yki:V5, Yki-WW:V5, or Yki-N:V5. Protein com-
plexes were washed, resolved by SDS-PAGE, and
bound proteins detected by V5 or GST antibodies,
as indicated.
(GAF:FLAG). However, both Yki isoforms
were coimmunoprecipitated with GAF
(Figure 2D), suggesting that the WW
domains are not required for GAF bind-
ing. The WW domains were also not
identified several nuclear proteins with known roles in transcrip- required for the interaction between GST-GAF and Yki in
tion, including GAF. Indeed, GAF was identified as the protein bacterial lysates (Figure 2F).
with the lowest log(e) score (an indication of high confidence
in the assignment) and was identified from one of the most Association of BRM and Mediator Complexes with Yki
prominent bands in nuclear extracts (Figure 2A; Table S3), Two additional proteins, Moira (Mor) and Mediator complex
implying that the correlation between Yki and GAF binding to subunit 23 (MED23), were also identified by low log(e) scores
chromatin reflects their physical association. and represented by multiple peptides (Table S3). Mor is a compo-
The identification of GAF as a Yki-interacting protein was nent of the Brahma (BRM) complex, the Drosophila cognate of the
confirmed by coimmunoprecipitation; western blotting re- conserved SWI/SNF chromatin-remodeling complex. Two addi-
vealed that GAF was specifically coprecipitated by anti-Yki tional components of the BRM complex, Brahma (Brm) and
and not by preimmune serum (Figure 2B). To determine Dalao, were also identified as nuclear Yki-associated proteins
whether this interaction is direct, we performed coprecipitation by mass spectrometry (Table S3). MED23 is a component of
assays on proteins expressed in bacteria. GAF was expressed Mediator (reviewed in Malik and Roeder, 2010). Four additional
as a glutathione S-transferase (GST) fusion protein. Yki was Mediator subunits, MED15, MED31, MED19, and MED1, were
pulled down by GST:GAF, but not by GST (Figure 2F). Earlier also identified as nuclear Yki-associated proteins by mass spec-
studies identified an essential role for the WW domains of Yki trometry (Table S3). Previously characterized DNA-binding part-
in transcriptional activation (Oh and Irvine, 2010). An isoform ners of Yki were either not identified (Hth, Mad) or identified but
of Yki in which all three Wts phosphorylation sites were only with low log(e) scores (Sd) (Table S3); their expression in S2
mutated (Yki:V53SA) or an isoform that also contained inacti- cells might be too low for robust identification by this approach.
vating mutations in the WW domains (Yki:V53SA-WW) (Oh and We confirmed the identification of Mor as a Yki-associated
Irvine, 2009) was cotransfected in S2 cells with tagged GAF protein through multiple approaches. Endogenous Mor was
312 Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authorsspecifically coimmunoprecipitated from cultured Drosophila Yki, such as dorsal-ventral boundary Wingless (Wg), were not
cells with Yki antibodies (Figure 2C), and V5-tagged Mor could significantly reduced by Trl RNAi (Figure 3F). We also extended
precipitate FLAG-tagged Yki (Figure 2E). Moreover, bacterially our analysis of GAF by showing that a minimal ban reporter,
expressed Yki was specifically precipitated by GST:Mor (Fig- brC12-lacZ, that responds directly to Yki and Mad (Oh and Irvine,
ure 2F). Thus, Yki and Mor can directly bind to each other, estab- 2011) was also downregulated by Trl RNAi (Figure 3F) and that
lishing a direct link from Yki to a chromatin-remodeling complex. Trl clones also exhibit decreased Diap1 levels (Figures S2K
Both wild-type Yki and Yki-WW also interacted with Mor (Figures and S2L).
2E and 2F), demonstrating that they bind to each other in a WW- These in vivo studies were complemented by transcriptional
domain-independent manner. Although the C-terminal half of Yki assays in S2 cells. We used two previously characterized
(from amino acid [aa] 241) was not expressed well enough in reporters: a ban reporter that is activated by Yki through Mad
bacteria to be assayed, the N-terminal half (Yki-N, up to aa (br2.5-luciferase) (Oh and Irvine, 2011), and a reporter that is
240) was expressed well. Yki-N did not interact well with GST- regulated by a Yki:Gal4 DNA-binding domain fusion protein
GAF but did interact with GST:Mor, suggesting that Mor, but (Yki:Gal4DBD, UAS-luciferase) (Oh and Irvine, 2009). In addition,
not GAF, interacts with the Yki N terminus (Figure 2F). Attempts we identified a Yki-responsive enhancer upstream of ex and
to confirm a physical interaction between individual Mediator used this to create an ex-luciferase reporter. For a non-Yki-
subunits and Yki were unsuccessful (not shown), but homologs responsive reporter, we used a Dpp-pathway responsive
of Yki (TAZ) and MED15 (ARC105) have been reported to coas- enhancer from Ubx (DRE-luciferase) (Oh and Irvine, 2011).
sociate within a complex in mammalian cells (Varelas et al., Reduction of GAF levels by Trl RNAi reduced Yki-mediated acti-
2008). vation of both the ban and ex reporters (Figures 3K and 3L).
UAS-luciferase was not significantly affected by Trl RNAi (Fig-
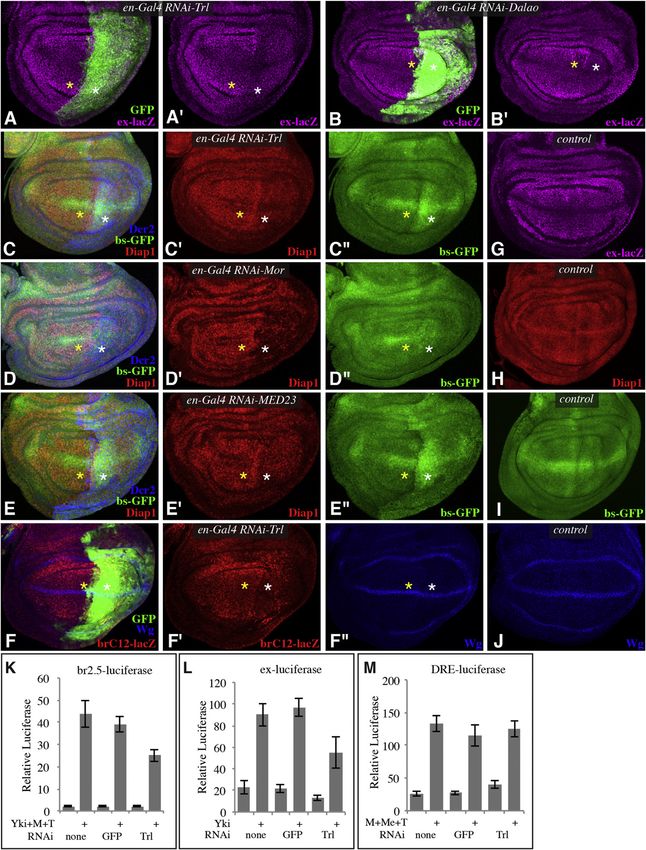
GAF, BRM, and Mediator Are Required for Yki Activity ure S3H), but the ChIP analysis implies that GAF does not act
A major biological function of Yki is to promote growth and at all Yki target genes. The non-Yki-responsive reporter (DRE-
inhibit apoptosis. Mutation or downregulation of Trl, BRM luciferase) was also not affected by Trl RNAi (Figure 3M). To
components, or Mediator subunits can result in growth defects assess requirements for BRM, we assayed the effects of
and reduced cell viability (Elfring et al., 1998; Farkas et al., RNAi-mediated knockdown of the same three subunits exam-
1994; Terriente-Félix et al., 2010) (Figure S2). To determine ined in vivo: Dalao, Mor, and Brm. Significant reductions were
whether this reflects roles in Yki-mediated transcription, three observed for both the ban reporter and UAS-luciferase (Fig-
well-established in vivo targets of Yki, expanded (ex), thread, ure S3I and S3J). These reporter assays further support the
(th, commonly referred to as Diap1), and bantam (ban, a conclusion that GAF and BRM contribute to Yki-mediated tran-
microRNA gene), were assayed in wing discs in which GAF, scriptional activation.
BRM subunits (Mor, Brm, or Dalao), or Mediator subunits
(Kto, MED23, or MED15) were downregulated by RNA interfer- Activated Yki Requires GAF, BRM, and Mediator
ence (RNAi). Expression of ex, monitored using an ex-lacZ To further investigate the requirement for GAF, BRM, and Medi-
reporter, could be reduced by knockdown of GAF (Figure 3A), ator in Yki-mediated transcription, we examined Yki target genes
BRM subunits (Figure 3B), or Mediator subunits (Figures S3A under conditions where Yki activity was elevated, either through
and S3B). Expression of th, monitored using Diap1 antibodies, reduction of Wts levels or through expression of an activated
was also reduced by knockdown of GAF (Figure 3C), BRM form of Yki (YkiS250A) (Oh and Irvine, 2009). The increased
subunits (Figures 3D, S2M, S2N, S3C, and S3D), or Mediator expression of Diap1 or ban normally observed in the presence
subunits (Figures 3E, S3E, and S3F). ban expression was moni- of activated Yki (Figures 4A and 4D) was either partially or
tored using a sensor, bs-GFP, in which ban target sites are completely suppressed when GAF, BRM, or Mediator subunits
present in the 30 UTR of a GFP transgene (Brennecke et al., were downregulated by RNAi (Figures 4 and S4). Thus, GAF,
2003). bs-GFP expression was upregulated by knockdown of BRM, and Mediator all contribute to Yki-mediated transcriptional
GAF or Mediator (Figures 3C, 3E, S3E, and S3F), which indi- activation in vivo. Moreover, these experiments provide genetic
cates that ban expression was reduced, and this effect was evidence that these genes act at or below the level of Yki rather
comparable to that induced by knockdown of Yki (Figure S3G). than on upstream components of the Hippo pathway, consistent
For Brm complex subunits, only subtle effects on bs-GFP with their identification as nuclear cofactors of Yki.
expression were observed when RNAi lines were expressed
under en-Gal4 control (Figures 3D, S3C, and S3D), but DISCUSSION
increased bs-GFP expression was obvious when RNAi lines
were expressed in clones under AyGal4 control (Figures S2M Molecular mechanisms underlying the transcriptional activation
and S2N). In sum, three well-established direct targets of Yki of target genes by Yki have remained ill-defined. We have reme-
are all dependent upon GAF, BRM, and Mediator for their died this by identifying multiple nuclear cofactors of Yki’s tran-
normal expression. scriptional activity: GAF, BRM, and Mediator. Our discovery of
The requirement for GAF in the expression of ex, th, and ban the association of Yki with GAF and BRM establishes a direct
confirms that the overlap of Yki- and GAF-binding regions on connection between Yki and chromatin-remodeling complexes,
chromatin, and their physical interaction, are reflective of a func- thus identifying direct linkage to chromatin-remodeling com-
tional requirement for GAF in the expression of these key Yki plexes as a mechanism by which Yki promotes transcription.
targets. Moreover, other genes that are not direct targets of The linkage of Yki to chromatin remodeling is further supported
Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors 313(legend on next page) 314 Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors
by the observation that wts mutations can influence position Studies of how Yki promotes growth have focused on specific
effect variegation (Figures S1G and S1H). target genes with known roles in promoting growth and cell-
The interaction between Yki and BRM is mediated by the BRM cycle progression and inhibiting apoptosis (Oh and Irvine,
subunit Mor, which binds directly to Yki. BRM is broadly required 2010). However, characterization of Yki’s localization to chro-
for gene activation in Drosophila (Armstrong et al., 2002), and matin suggests that Yki may have thousands of direct targets.
homologous SWI/SNF complexes are broadly required for tran- Yki is directly associated with thousands of chromosomal loci,
scriptional activation in other eukaryotes (Martens and Winston, and both global transcriptional analysis and H3K4me3-modifica-
2003). These complexes promote transcription by remodeling tion patterns indicate that the bulk of this Yki localization is corre-
nucleosomes. Our biochemical and genetic studies identify the lated with increased transcription. Thus, we infer that Yki is
recruitment of BRM as a crucial aspect as Yki’s transcriptional involved in transcriptional activation on a genome-wide scale.
activity, and this link between them is intriguing in light of the This suggests a distinct perspective on how Yki effects its core
biological processes to which both chromatin regulators and biological function of promoting growth. We speculate that in
Yki/Yap activity have been linked, including the growth of addition to activating select growth loci, Yki’s role as a growth
diverse tumors, stem cell maintenance and pluripotency, and promoter might also stem from an ability to induce a broad
regeneration. increase in cellular transcription. Increased growth requires an
The requirement for GAF further stresses the importance of increase in cellular mass, which implies an increased need for
chromatin remodeling to Yki activity and suggests additional thousands of cellular constituents. Indeed, the crucial role of
mechanisms by which Yki promotes transcription. GAF is Myc in promoting growth is thought to stem in part from its
a multifunctional cofactor: it has been linked to chromatin re- role in promoting the expression of ribosomal proteins (van Rig-
modeling through interactions with NURF (Tsukiyama and gelen et al., 2010), and thereby increasing cellular mass through
Wu, 1995) and FACT (Shimojima et al., 2003), and it has also a global increase in cellular translation. Moreover, recent studies
been linked to the general transcriptional machinery (Chopra of Myc have also indicated that it induces a global increase in
et al., 2008) and to looping (Agelopoulos et al., 2012). GAF transcriptional activation (Lin et al., 2012; Nie et al., 2012). A
may combine multiple functions in a single regulatory structure. widespread increase in cellular transcription, induced by Yki,
Indeed, a model was proposed in which nucleosome displace- might similarly promote growth by contributing to the increased
ment by GAF would make DNA recognition sites of activators cellular mass necessary to sustain increased growth rates.
or repressors accessible, and then these proteins would act
in conjunction with GAF to recruit transcriptional machinery EXPERIMENTAL PROCEDURES
(Lehmann, 2004). Although GAF has only been intensively
Drosophila Crosses
studied in Drosophila, the vertebrate c-Krox/Th-POK has
Flies were cultured at 25 C or 29 C. Ectopic expression was induced in eyes
been proposed as functional homolog (Matharu et al., 2010), using GMR-Gal4 and in posterior cells using en-Gal4 UAS-GFP or en-Gal4 bs-
and it will be interesting to see if it has a role in vertebrate GFP with or without UAS-dcr2 and with or without UAS-lacZ. For flip-out
Hippo pathways. ectopic expression clones, UAS-transgenes with y w hs-FLP[122] were
Considering that Yki interacts with protein complexes involved crossed to w; Act > y+ > Gal4 bs-GFP (AyGal4-bs-GFP) or AyGal4,UAS-
at different steps of activation, Yki may play multiple roles to GFP. For MARCM experiments, y w hs-FLP[122] tub-Gal4 UAS-GFP;tub-
Gal80 FRT80B was crossed to hs-FLP[122]; UAS-P35; P[lacW]Trls2325
coordinate and integrate these steps. We propose that Yki is re-
FRT80B/TM6B.
cruited to specific loci through sequence-specific DNA-binding
partners. Yki and GAGA may reinforce each other’s localization Histology and Imaging
to chromosomal sites through their binding to each other and Imaginal discs were fixed and stained as described previously (Cho and Irvine,
also through their recruitment of chromatin-remodeling com- 2004) using as primary antibodies rabbit anti-Yki (1:400) (Oh and Irvine, 2008),
plexes, which could make DNA-binding sites more accessible. mouse anti-Wg (1: 800, DSHB), rabbit anti-Dcr2 (1:1,600, Abcam), goat anti-
b-gal (1:400, Biogenesis), active caspase3 (1:400, Cell Signaling), and mouse
Yki and GAF could then each contribute to recruitment of addi-
anti-Diap1 (1:400, gift of Bruce Hay). Fluorescent stains were captured on
tional transcriptional coactivators, Mediator, and the general a Leica TCS SP5 confocal.
transcriptional machinery, with this recruitment further facilitated Additional experimental details are in the Extended Experimental
by the nucleosome-remodeling activities of BRM and NURF. Procedures.
Figure 3. Influence of GAF, BRM, and Mediator on Yki Target Genes
(A–J) Projections through three to five confocal sections of wing discs; panels marked by prime symbols show separated channels. Yellow asterisks identify
regions with normal gene expression, and white asterisks identify regions with expression of RNAi lines and altered target gene expression. (A and B) en-
Gal4 UAS-GFP ex-lacZ UAS-Dcr2, with (A) UAS-RNAi-Trl[vdrc106433] (B) UAS-RNAi-Dalao[TRiP.JF02116] showing expression of ex-lacZ (magenta) and with
posterior cells marked by GFP (green). (C–E) en-Gal4 bs-GFP; UAS-Dcr2 and with (C) UAS-RNAi-Trl[vdrc106433], (D) UAS-RNAi-Mor[vdrc110712], (E) UAS-
RNAi-MED23[vdrc105247], showing expression of Diap1 (red) and bs-GFP (green), and with posterior cells marked by Dcr2 (blue). (F) en-Gal4 UAS-GFP; UAS-
RNAi-Trl[vdrc106433] UAS-Dcr2 brC12-lacZ, showing expression of brC12-lacZ (red) and Wg (blue) and with posterior cells marked by GFP (green). (G) Wild-type
control showing expression of ex-lacZ (magenta). (H) Wild-type control showing expression of Diap1 (red). (I) Wild-type control showing expression of bs-GFP
(green). (J) Wild-type control showing expression of Wg (blue).
(K–M) Histograms showing results of luciferase assays (depicted as average firefly/renilla ratio from triplicate experiments, where error bars indicate SD) using (K)
br2.5-luciferase, (L) ex-luciferase, or (M) DRE-luciferase reporters in S2 cells transfected to express Yki, TkvQ235D (T), Mad (M), or Medea (Me) as indicated.
Double-stranded RNAs for RNAi against the specific genes were also added as indicated.
See also Figures S2 and S3.
Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors 315Figure 4. GAF, BRM, and Mediator Are
Required for Yki Activity
Projections through three to five confocal sections
of wing discs; panels marked by prime symbols
show separated channels. Yellow asterisks iden-
tify regions with normal gene expression, and
white asterisks identify regions with expression of
RNAi lines and altered target gene expression.
(A–C) en-Gal4 bs-GFP; UAS-Yki:V5S250A UAS-
Dcr2 and with (A) UAS-lacZ (control) (B) UAS-RNAi-
Trl[vdrc106433], (C) UAS-RNAi-Mor[vdrc110712],
showing expression of Diap1 (red) and bs-GFP
(green) and with posterior cells marked by Yki
(blue).
(D–G) en-Gal4 bs-GFP; UAS-RNAi- wts[vdrc9928]
UAS-Dcr2 and with (D) UAS-lacZ (E) UAS-RNAi-
Trl[vdrc106433], (F) UAS-RNAi-Mor[vdrc110712],
or (G) UAS-RNAi-MED23[vdrc105247], showing
expression of Diap1 (red) and bs-GFP (green) and
with posterior cells marked by Dcr2 (blue).
See also Figure S4.
316 Cell Reports 3, 309–318, February 21, 2013 ª2013 The AuthorsACCESSION NUMBERS Farkas, G., Gausz, J., Galloni, M., Reuter, G., Gyurkovics, H., and Karch, F.
(1994). The Trithorax-like gene encodes the Drosophila GAGA factor. Nature
ChIP-seq and RNA-seq data are available under GEO accession number 371, 806–808.
GSE38594. Halder, G., and Carroll, S.B. (2001). Binding of the Vestigial co-factor switches
the DNA-target selectivity of the Scalloped selector protein. Development 128,
SUPPLEMENTAL INFORMATION 3295–3305.
Justice, R.W., Zilian, O., Woods, D.F., Noll, M., and Bryant, P.J. (1995). The
Supplemental Information includes four figures, four tables, and Extended Drosophila tumor suppressor gene warts encodes a homolog of human
Experimental Procedures and can be found with this article online at http:// myotonic dystrophy kinase and is required for the control of cell shape and
dx.doi.org/10.1016/j.celrep.2013.01.008. proliferation. Genes Dev. 9, 534–546.
Lehmann, M. (2004). Anything else but GAGA: a nonhistone protein complex
LICENSING INFORMATION reshapes chromatin structure. Trends Genet. 20, 15–22.
Li, B., Carey, M., and Workman, J.L. (2007). The role of chromatin during tran-
This is an open-access article distributed under the terms of the Creative scription. Cell 128, 707–719.
Commons Attribution-NonCommercial-No Derivative Works License, which
Lin, C.Y., Lovén, J., Rahl, P.B., Paranal, R.M., Burge, C.B., Bradner, J.E., Lee,
permits non-commercial use, distribution, and reproduction in any medium,
T.I., and Young, R.A. (2012). Transcriptional amplification in tumor cells with
provided the original author and source are credited.
elevated c-Myc. Cell 151, 56–67.
Malik, S., and Roeder, R.G. (2010). The metazoan Mediator co-activator
ACKNOWLEDGMENTS
complex as an integrative hub for transcriptional regulation. Nat. Rev. Genet.
11, 761–772.
We thank B. Hay, C.P. Verrijzer, T. Xu, the DSHB, and the Bloomington Stock
Center for antibodies and Drosophila stocks; and Flybase, Dibyendu Kumar, Martens, J.A., and Winston, F. (2003). Recent advances in understanding
and the Waksman Genomics Core for bioinformatics and sequencing support. chromatin remodeling by Swi/Snf complexes. Curr. Opin. Genet. Dev. 13,
This research was supported by HHMI and by NIH grants GM078620 136–142.
(to K.D.I.), 5R01GM054510 (to R.S.M.), 5P50GM081892 (to K.P.W.), and Matharu, N.K., Hussain, T., Sankaranarayanan, R., and Mishra, R.K. (2010).
3U01HG004264 (to K.P.W.). Vertebrate homologue of Drosophila GAGA factor. J. Mol. Biol. 400, 434–447.
Nicolay, B.N., Bayarmagnai, B., Islam, A.B.M.M.K., Lopez-Bigas, N., and
Received: July 27, 2012 Frolov, M.V. (2011). Cooperation between dE2F1 and Yki/Sd defines a distinct
Revised: October 29, 2012 transcriptional program necessary to bypass cell cycle exit. Genes Dev. 25,
Accepted: January 11, 2013 323–335.
Published: February 7, 2013
Nie, Z., Hu, G., Wei, G., Cui, K., Yamane, A., Resch, W., Wang, R., Green,
D.R., Tessarollo, L., Casellas, R., et al. (2012). c-Myc is a universal amplifier
REFERENCES of expressed genes in lymphocytes and embryonic stem cells. Cell 151,
68–79.
Adkins, N.L., Hagerman, T.A., and Georgel, P. (2006). GAGA protein: a multi-
faceted transcription factor. Biochem. Cell Biol. 84, 559–567. Oh, H., and Irvine, K.D. (2008). In vivo regulation of Yorkie phosphorylation and
localization. Development 135, 1081–1088.
Agelopoulos, M., McKay, D.J., and Mann, R.S. (2012). Developmental regula-
tion of chromatin conformation by Hox proteins in Drosophila. Cell Rep. 1, Oh, H., and Irvine, K.D. (2009). In vivo analysis of Yorkie phosphorylation sites.
350–359. Oncogene 28, 1916–1927.
Armstrong, J.A., Papoulas, O., Daubresse, G., Sperling, A.S., Lis, J.T., Scott, Oh, H., and Irvine, K.D. (2010). Yorkie: the final destination of Hippo signaling.
M.P., and Tamkun, J.W. (2002). The Drosophila BRM complex facilitates Trends Cell Biol. 20, 410–417.
global transcription by RNA polymerase II. EMBO J. 21, 5245–5254. Oh, H., and Irvine, K.D. (2011). Cooperative regulation of growth by Yorkie and
Brennecke, J., Hipfner, D.R., Stark, A., Russell, R.B., and Cohen, S.M. (2003). Mad through bantam. Dev. Cell 20, 109–122.
bantam encodes a developmentally regulated microRNA that controls cell Peng, H.W., Slattery, M., and Mann, R.S. (2009). Transcription factor choice in
proliferation and regulates the proapoptotic gene hid in Drosophila. Cell 113, the Hippo signaling pathway: homothorax and yorkie regulation of the micro-
25–36. RNA bantam in the progenitor domain of the Drosophila eye imaginal disc.
Cho, E., and Irvine, K.D. (2004). Action of fat, four-jointed, dachsous Genes Dev. 23, 2307–2319.
and dachs in distal-to-proximal wing signaling. Development 131, 4489– Pérez-Lluch, S., Blanco, E., Carbonell, A., Raha, D., Snyder, M., Serras, F., and
4500. Corominas, M. (2011). Genome-wide chromatin occupancy analysis reveals
Chopra, V.S., Srinivasan, A., Kumar, R.P., Mishra, K., Basquin, D., Docquier, a role for ASH2 in transcriptional pausing. Nucleic Acids Res. 39, 4628–4639.
M., Seum, C., Pauli, D., and Mishra, R.K. (2008). Transcriptional activation Shimojima, T., Okada, M., Nakayama, T., Ueda, H., Okawa, K., Iwamatsu, A.,
by GAGA factor is through its direct interaction with dmTAF3. Dev. Biol. 317, Handa, H., and Hirose, S. (2003). Drosophila FACT contributes to Hox gene
660–670. expression through physical and functional interactions with GAGA factor.
Dong, J., Feldmann, G., Huang, J., Wu, S., Zhang, N., Comerford, S.A., Genes Dev. 17, 1605–1616.
Gayyed, M.F., Anders, R.A., Maitra, A., and Pan, D. (2007). Elucidation of Subramanian, A., Tamayo, P., Mootha, V.K., Mukherjee, S., Ebert, B.L.,
a universal size-control mechanism in Drosophila and mammals. Cell 130, Gillette, M.A., Paulovich, A., Pomeroy, S.L., Golub, T.R., Lander, E.S., and
1120–1133. Mesirov, J.P. (2005). Gene set enrichment analysis: a knowledge-based
Eissenberg, J.C., and Shilatifard, A. (2010). Histone H3 lysine 4 (H3K4) meth- approach for interpreting genome-wide expression profiles. Proc. Natl.
ylation in development and differentiation. Dev. Biol. 339, 240–249. Acad. Sci. USA 102, 15545–15550.
Elfring, L.K., Daniel, C., Papoulas, O., Deuring, R., Sarte, M., Moseley, S., Terriente-Félix, A., López-Varea, A., and de Celis, J.F. (2010). Identification of
Beek, S.J., Waldrip, W.R., Daubresse, G., DePace, A., et al. (1998). Genetic genes affecting wing patterning through a loss-of-function mutagenesis
analysis of brahma: the Drosophila homolog of the yeast chromatin remodeling screen and characterization of med15 function during wing development.
factor SWI2/SNF2. Genetics 148, 251–265. Genetics 185, 671–684.
Cell Reports 3, 309–318, February 21, 2013 ª2013 The Authors 317Tsukiyama, T., and Wu, C. (1995). Purification and properties of an ATP- Wu, S., Liu, Y., Zheng, Y., Dong, J., and Pan, D. (2008). The TEAD/TEF family
dependent nucleosome remodeling factor. Cell 83, 1011–1020. protein Scalloped mediates transcriptional output of the Hippo growth-regula-
tory pathway. Dev. Cell 14, 388–398.
van Riggelen, J., Yetil, A., and Felsher, D.W. (2010). MYC as a regulator of
Zhang, L., Ren, F., Zhang, Q., Chen, Y., Wang, B., and Jiang, J. (2008a). The
ribosome biogenesis and protein synthesis. Nat. Rev. Cancer 10, 301–309.
TEAD/TEF family of transcription factor Scalloped mediates Hippo signaling
Varelas, X., Sakuma, R., Samavarchi-Tehrani, P., Peerani, R., Rao, B.M., in organ size control. Dev. Cell 14, 377–387.
Dembowy, J., Yaffe, M.B., Zandstra, P.W., and Wrana, J.L. (2008). TAZ Zhao, B., Tumaneng, K., and Guan, K.L. (2011). The Hippo pathway in organ
controls Smad nucleocytoplasmic shuttling and regulates human embryonic size control, tissue regeneration and stem cell self-renewal. Nat. Cell Biol.
stem-cell self-renewal. Nat. Cell Biol. 10, 837–848. 13, 877–883.
318 Cell Reports 3, 309–318, February 21, 2013 ª2013 The AuthorsYou can also read

















































