Gene amplification is a relatively frequent event leading to ZBTB7A (Pokemon) overexpression in non-small cell lung cancer
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Pathology
J Pathol 2007; 213: 294–302
Published online 28 September 2007 in Wiley InterScience
(www.interscience.wiley.com) DOI: 10.1002/path.2222
Original Paper
Gene amplification is a relatively frequent event leading
to ZBTB7A (Pokemon) overexpression in non-small cell
lung cancer
K Apostolopoulou,1 IS Pateras,1 K Evangelou,1 PK Tsantoulis,1 M Liontos,1 C Kittas,1 DG Tiniakos,1 A Kotsinas,1
C Cordon-Cardo2 and VG Gorgoulis1 *
1 Molecular Carcinogenesis Group, Laboratory of Histology–Embryology, Medical School, University of Athens, Athens, Greece
2 Department of Pathology, Memorial Sloan–Kettering Cancer Center, Sloan-Kettering Institute, 1275 York Avenue, New York, NY 10021, USA
*Correspondence to: Abstract
VG Gorgoulis, Antaiou 53 Str.,
Lamprini, Ano Patissia 111 46, ZBTB7A (Pokemon) is a member of the POK family of transcriptional repressors. Its main
Athens, Greece. function is the suppression of the p14ARF tumour suppressor gene. Although ZBTB7A
E-mail: histoclub@ath.forthnet.gr expression has been found to be increased in various types of lymphoma, there are no
reports dealing with its expression in solid tumours. Given that p14ARF inhibits MDM2,
No conflicts of interest were
the main negative regulator of p53, we hypothesized that overexpression of ZBTB7A could
declared.
lead indirectly to p53 inactivation. To this end, we examined the status of ZBTB7A and
its relationship with tumour kinetics (proliferation and apoptosis) and nodal members
of the p53 network in a panel of 83 non-small cell lung carcinomas (NSCLCs). We
observed, in the majority of the samples, prominent expression of ZBTB7A in the cancerous
areas compared to negligible presence in the adjacent normal tissue elements. Gene
amplification (two- to five-fold) was found in 27.7% of the cases, denoting its significance as
a mechanism driving ZBTB7A overproduction in NSCLCs. In the remaining non-amplified
group of carcinomas, analysis of the mRNA and protein expression patterns suggested
that deregulation at the transcriptional and post-translational level accounts for ZBTB7A
overexpression. Proliferation was associated with ZBTB7A expression (p = 0.033) but not
apoptosis. The association with proliferation was reflected in the positive correlation between
ZBTB7A expression and tumour size (p = 0.018). The overexpression of ZBTB7A in both
p53 mutant and p53 wild-type cases, implies either a synergistic effect or that ZBTB7A exerts
its oncogenic properties independently of the p14ARF –MDM2–p53 axis. The concomitant
expression of ZBTB7A with p14ARF (p = 0.039), instead of the anticipated inverse relation,
supports the latter notion. In conclusion, regardless of the pathway followed, the distinct
expression of ZBTB7A in cancerous areas and the association with proliferation and tumour
size pinpoints a role for this novel cell cycle regulator in the pathogenesis of lung cancer.
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John
Received: 27 April 2007 Wiley & Sons, Ltd.
Revised: 21 June 2007
Keywords: ZBTB7A; Pokemon; p14ARF ; amplification; overexpression; differential PCR;
Accepted: 6 July 2007
lung cancer
Introduction a NH2-terminal POZ/BTB domain (poxvirus zinc
finger/broad-complex tramtrack, bric-à-brac) and a
Pokemon (POK erythroid myeloid ontogenic fac- COOH-terminal domain, consisting of Kruppel-type
tor), also known as FBI-1 (factor that binds to zinc fingers. The POZ/BTB domain is responsi-
IST, the HIV-1 inducer of short transcripts), LRF ble for homodimerization/heterodimerization and the
(leukaemia/lymphoma-related factor — mouse homo- recruitment of corepressor complexes, such as histone
logue) and OCZF (osteoclast-derived zinc finger — rat deacetylases, whereas Kruppel-type zinc fingers are
homologue) [1], is encoded by the ZBTB7A gene, responsible for specific DNA recognition and binding
which resides in the 19p13.3 locus (Figure 1A) and [3] (Figure 1A).
belongs to the POK family of transcriptional repres- A recent in vitro study showed that mouse embryo
sors. This family participates in embryonic develop- fibroblasts (MEFs) lacking ZBTB7A resisted oncogene-
ment and differentiation, but has also been recently mediated transformation. This resistance was restored
implicated in carcinogenesis [2]. Members of the POK when ZBTB7A was co-expressed with various onco-
family have a characteristic structure and contain genes, such as EIA + RasV12 , Myc + RasV12 and
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.
www.pathsoc.org.uk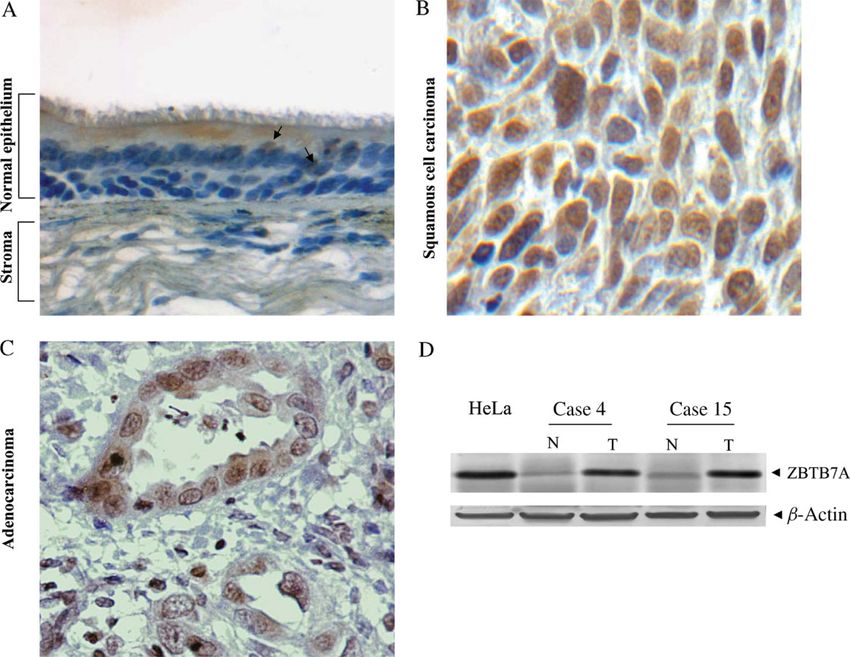
ZBTB7A/Pokemon gene amplification in NSCLC 295
Figure 1. (A) Schematic diagrams depicting: (i) the structure and 19p13.3 chromosomal localization of the ZBTB7A/POKEMON
gene (arrows indicate the location of primer pairs employed in ZBTB7A D-PCR and RT–PCR analysis); and (ii) ZBTB7A protein
structure. The NH2-terminus contains a POZ/BTB domain and the COOH-terminus four Kruppel-type zinc fingers. (B) The
ZBTB7A–ARF network. ZBTB7A suppresses p14ARF expression through direct binding to its promoter [4]. P14ARF , an important
sensor of aberrant proliferation [16], interacts with Mdm2, leading to p53 stabilization [6]
SV40 large T antigen + RasV12 , rapidly leading MEFs [8], thus affecting the activity of p53, we hypoth-
to overt oncogenic transformation [4,5]. The poten- esized that ZBTB7A could disturb the balance of
tial of ZBTB7A as a genuine proto-oncogene was the p14ARF –MDM2–p53 network by increasing its
also examined in vivo in transgenic mice that over- levels in cancer. This could represent an alternative
expressed ZBTB7A in immature T and B lymphoid mechanism leading to p53 inactivation. To this end,
cells. These mice developed thymic lymphomas and we examined the status of ZBTB7A and its rela-
tumour infiltration of bone marrow [4] and their sur- tionship with p14ARF , tumour kinetics and eventually
vival was significantly lower than that of wild-type clinico-pathological features, in an expanded panel of
mice. Despite the accumulation of evidence from non-small cell lung carcinomas (NSCLCs), previously
in vitro and animal studies, studies of ZBTB7A in analysed for the status of the E2F1–p53 network [9].
human cancer specimens are lacking and are nec- To the best of our knowledge, this is the first compre-
essary to elucidate its role in human malignan- hensive study of ZBTB7A expression in lung cancer.
cies.
The main mode of action of ZBTB7A seems to
be through the direct binding and transcriptional Materials and methods
repression of p14ARF , a major tumour suppressor
[4]. Upon culture shock, MEFs lacking ZBTB7A Tissue samples
(ZBTB7A−/− ) had higher p19ARF protein levels than We used frozen and formalin-fixed paraffin-embedded
wild-type MEFs. In addition, the proliferation defects tissue from 83 NSCLCs, including adjacent normal
and the inability of ZBTB7A−/− MEFs to be trans- lung tissue (Table 1). The tumours were classified
formed, were fully rescued by CDKN2A (p19ARF ) loss according to the World Health Organization criteria
[4]. (TNM system) and staged appropriately. Patients did
Knowing that the activity of p14ARF , the nega- not receive chemo-, radio- or immunotherapy prior to
tive regulator of MDM2 [6,7], can be modulated surgery. The study was performed with the approval
by the opposing actions of ZBTB7A [4] and E2F1 of the Ethics Committee of the University of Athens
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.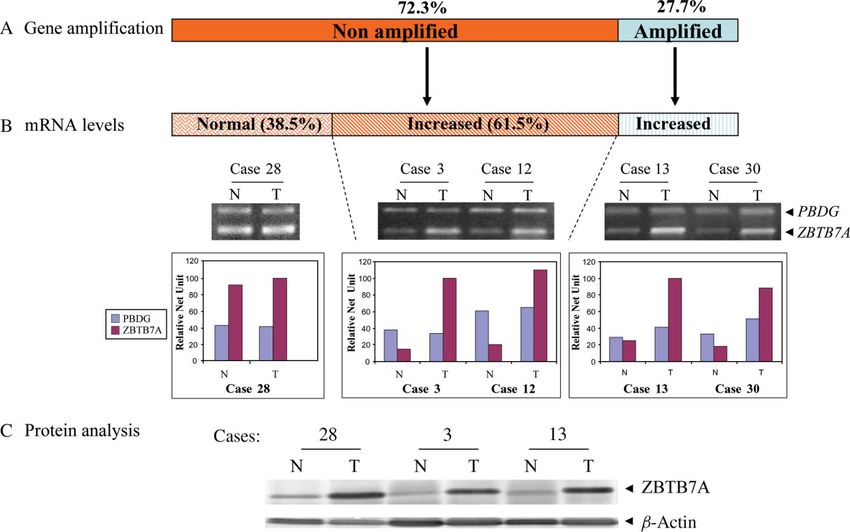
296 K Apostolopoulou et al
Table 1. Summary of clinicopathological features in our NSCLC randomly chosen, high-power fields (×400). Stained
series (n = 83 cases) cells were considered positive irrespective of staining
Median age in years (IQR) 67.5 (7.0) intensity. Slide evaluation was performed by four
independent observers (KA, IP, KE and VG), with
Sex (n)
Males 75 minimal interobserver variability.
Females 8
Smoking (n) ZBTB7A, p14ARF Strictly nuclear immunopositivity
Yes 72
No 9
was estimated. Corresponding labelling indices were
No available data 2 determined as the percentage of cancer cells with
positive nuclear immunostaining.
Histology (n)
Squamous 42
Adenocarcinoma 28
Adenosquamous 6
p53, E2F1 and Ki67 Labelling indices were esti-
Large cell 3 mated as described elsewhere [9,11].
Other subtypes 4
Stage (n)
I 44
Total protein extraction and western blot analysis
II 27
III 7
Total protein extracts from 16 matched, frozen, nor-
No available data 5 mal and tumour tissues, were obtained according to
protocols described elsewhere [10].
and in accordance with the revised (1983) Helsinki
Declaration of 1975. Antibodies and controls
The following antibodies were used for western blot
Immunohistochemistry analysis: anti-Pokemon (ZBTB7A; rabbit polyclonal,
Antibodies ABCAM ab36606; Antisel, Athens, Greece; 1 : 400);
anti-actin (AC-15; class, IgG1 mouse monoclonal;
The following antibodies were used for immuno-
Abcam; AntiSel, Athens, Greece; 1 : 1000). Actin was
histochemistry (IHC): anti-Pokemon (ZBTB7A; rab-
used to assess equal loading of total protein per
bit polyclonal, ABCAM ab36606; Antisel, Athens,
sample. Extracts from a HeLa cervical cancer cell line
Greece; 1 : 100); anti-p14 (rabbit polyclonal, Neomark-
were used as the positive control [12].
ers RB-1554; Bioanalytica, Athens, Greece; 1 : 100);
anti-Ki67 (MIB1; class IgG1 mouse monoclonal;
epitope, Ki67 nuclear antigen; DAKO, Kalifronas, Gel electrophoresis, blotting and signal development
Athens, Greece; 1 : 100); anti E2F1 (KH95; class
IgG2a mouse monoclonal; epitope, Rb-binding domain Gel electrophoresis, transfer to PVDF membrane
of E2F1 p60; Santa Cruz, Bioanalytica Athens, (NEN; AlterChem, Greece) and blocking were per-
Greece; 1 : 100); anti-p53 (DO7; class IgG2b mouse formed according to a previously described protocol
monoclonal; epitope, residues 1–45; DAKO, Kali- [10]. Signal development was performed with mem-
fronas, Athens, Greece; 1 : 100). brane incubation in nitro blue tetrazolium/5-bromo-4-
chloro-3-indolylphosphate (NBT/BCIP) solution
Method (Promega, SB Biotechnology, Athens, Greece). All
results were validated twice.
IHC was performed as previously described [10].
Controls Microdissection and DNA extraction
Human skin squamous cell carcinoma and cervical Microdissection and DNA extraction were performed
carcinoma tissue sections were employed as posi- following previously published protocols [9,10].
tive controls for ZBTB7A and p14ARF IHC staining,
respectively, according to the manufacturers’ instruc-
tions. For E2F1 and p53, previously described controls Differential polymerase chain reaction (D-PCR)
were used [9,11]. No staining was observed when
ZBTB7A gene amplification was assessed using the
the primary antibody was omitted (negative controls).
D-PCR method [9] and results were evaluated as
Adjacent normal lung parenchyma staining was used
previously described [9]. Primers, annealing temper-
as an internal control.
atures and product sizes are given in Table 2. Each
sample was verified three times. Neoplastic samples
Evaluation
scoring ≥ two-fold difference from the corresponding
The labelling indices were calculated by formally normal counterpart were indicative of ZBTB7A gene
counting immunopositive and negative cells in six amplification.
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.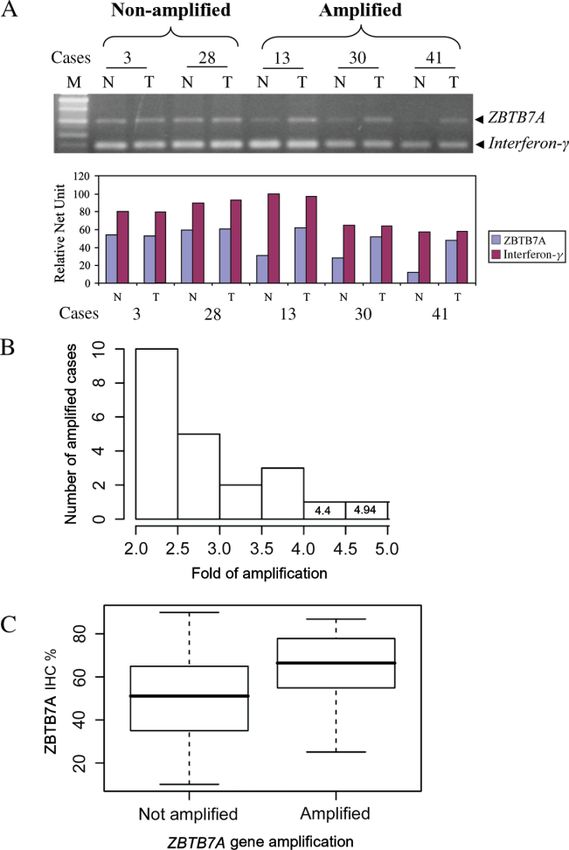
ZBTB7A/Pokemon gene amplification in NSCLC 297
Table 2. Primers and annealing temperatures
Primers Product Annealing
Locus (application) Sequence length (bp) temperature (◦ C)
ZBTB7A D-PCR
Fw 5 -GAA CGA GGG TTT AGT GCA-3 299 57
Rv 5 -CGA GCT GTT CTG GAG AGA-3
RT–PCR
Fw 5 -TCT GCG AGA AGG TCA TCC-3 198 59
Rv 5 -CGT AGT TGT GGG CAA AGG-3
IFN-γ D-PCR
Fw 5 -CTC TTT TCT TTC CCG ATA GGT-3 151 57
Rv 5 -CTG GGA TGC TCT TCG ACC TCG-3
PBDG RT–PCR
Fw 5 -TGA ACG GCG GAA GAA AAC A-3 315 59
Rv 5 -GCA GAT GGC TCC GAT GGT G-3
Fw, forward; Rv, reverse; IFN-γ , interferon-γ ; PBDG, porphobilinogen deaminase gene; D-PCR, differential polymerase chain
reaction; RT–PCR, reverse transcription polymerase chain reaction.
Figure 2. Representative results of ZBTB7A protein expression in normal and tumour lung tissues. (A) ZBTB7A IHC analysis
showing nuclear immunostaining in very few parabasal intermediate cells (depicted by arrows) of the normal lung epithelium (×200).
(B) Nuclear immunopositivity in squamous cell carcinoma (case 4; ×600). (C) Nuclear immunoreactivity in adenocarcinoma (case
15; ×400). (D) Western blot analysis of ZBTB7A in total protein extracts from matched normal (N)/tumour (T) samples (cases 4
and15). A band of 86 kDa is clearly overexpressed in tumour tissue. HeLa, positive control; β-actin, loading control. Colour signal
from NBT/BCIP treatment was discarded during image processing
RNA extraction and cDNA synthesis Comparative reverse transcriptase reaction
(RT–PCR)
RNA extractions from cancerous material with more
than 90% tumour cells and cDNA syntheses were per- ZBTB7A and porphobilinogen deaminase gene
formed according to a previously published protocol (PBDG) mRNA levels were assessed using a semi-
[10]. quantitative multiplex RT–PCR method [10] and
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.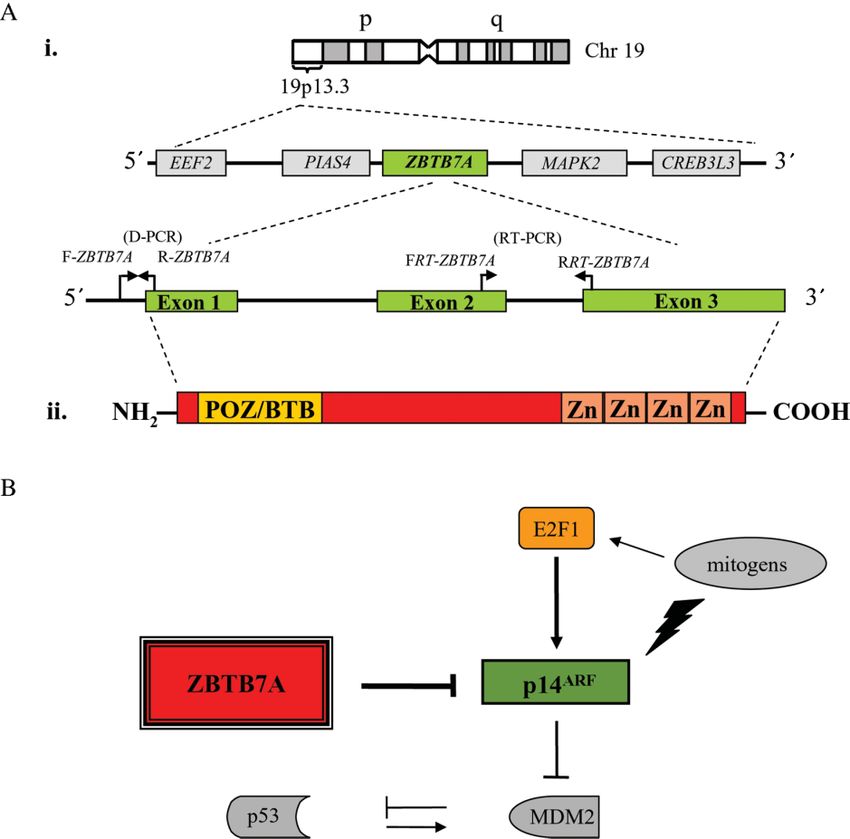
298 K Apostolopoulou et al
evaluation was carried out as previously published
[9]. Primers, annealing temperatures and product sizes
are given in Table 2. Each sample was verified three
times. Neoplastic samples scoring ≥ two-fold differ-
ence from the corresponding normal counterpart were
characterized as overexpressed.
Statistical analysis
Data analysis was performed using the R language
for statistics. Non-parametric tests (Spearman and
Kendall rank correlations, Wilcoxon and Kruskal rank
sum tests) were used accordingly and adjustment for
multiple comparisons was made where applicable.
Results were considered significant when p < 0.05
[13].
Results
The transcriptional repressor ZBTB7A is
overexpressed in NSCLCs
The immunohistochemical signal of ZBTB7A was
nuclear and staining was clearly prevalent in the can-
cerous nests, as evident in Figure 2 and Supplementary
Figure 1 (available at: http://www.interscience.wiley.
com/jpages/0022-3417/suppmat/path.2222.html). In
the adjacent normal bronchial epithelium, a limited
number of parabasal intermediate cells were stained
(Figure 2A). Few tumour-associated stromal cells
exhibited ZBTB7A immunopositivity (Figure 2C). Figure 3. ZBTB7A gene amplification in NSCLCs. (A)
ZBTB7A immunoreactivity was noticed in all carci- Differential PCR (D-PCR) analysis in five representative cases
(Interferon-γ ; reference gene; M, 100bp marker). Absence
nomas examined in 10–90% of tumour cells (mean of ZBTB7A amplification in cases 3 and 28 and presence
52%). Over half of our cases had immunohistochem- of amplification in cases 13, 30 and 41, as shown by gel
ical ZBTB7A expression > 50%. We did not detect electrophoresis and densitometry (presented as histogram).
any histological subtype predominance in ZBTB7A (B) Histogram depicting the range of ZBTB7A fold amplification
expression (Figure 2B, C). The in situ findings were in relation to the number of amplified cases. (C) Box
plot showing the significant correlation between ZBTB7A
confirmed by immunoblotting (Figure 2D). In HeLa amplification and IHC expression (t-test, p = 0.043)
cells and representative matched normal–tumour sam-
ples, a band of approximately 86 kDa was revealed,
which corresponds to the expected detection size of
cases showed increased mRNA and protein levels,
ZBTB7A (Figure 2D). Although the protein’s calcu-
lated molecular weight is 61.5 kDa, it has been shown suggesting that the amplification may have functional
that it runs at an apparent molecular mass of 86 kDa consequences (Figure 4). Supporting this, the carci-
[12,14]. nomas with ZBTB7A amplification demonstrated sig-
nificantly higher ZBTB7A labelling indices than the
non-amplified ones (t-test, p = 0.043; Figure 3C).
Gene amplification is a relatively frequent In the non-amplified cases, increased ZBTB7A
mechanism of ZBTB7A overexpression mRNA levels were observed in 37/60 samples
According to our working hypothesis, the predominant (61.5%), whereas in the remaining group the mRNA
expression of ZBTB7A in cancerous areas implies levels were normal (Figure 4A, B). The concordance
an oncogenic role in lung cancer. Since the other between mRNA and protein expression was not
oncogene of the network, MDM2 (Figure 1B), exerts absolute, since immunoblotting in all cases with
its transformation effects via gene amplification [6,7], normal ZBTB7A mRNA showed higher protein levels
we decided to examine whether the same mechanism in the cancerous area compared to the adjacent
stands for ZBTB7A as well. normal tissue (Figure 4C). This finding suggests
Indeed, D-PCR analysis revealed gene amplifica- that, besides gene amplification, epigenetic, post-
tion of ZBTB7A in 23/83 informative cases (27.7%) transcriptional and post-translational mechanisms may
(Figure 3A). The levels of amplification were in the be implicated in the overexpression of the ZBTB7A
range two- to five-fold (Figure 3B). All amplified gene in NSCLC.
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.ZBTB7A/Pokemon gene amplification in NSCLC 299
Figure 4. Frequency of ZBTB7A gene amplification and overexpression in NSCLCs. (A) Incidence of ZBTB7A amplification
in NSCLCs. (B) Frequency of ZBTB7A transcriptional overexpression and relationship with distribution of gene amplification
in NSCLC. Representative cases with (cases 3, 12, 13 and 30) and without (case 28) mRNA overexpression, accompanied
by histograms showing densitometric evaluations of RT–PCR analysis in matched normal (N)/tumour (T) samples (PBDG,
porphobilinogen deaminase gene, i.e. reference gene). (C) Frequent ZBTB7A overexpression, as assessed by western blot analysis,
even in cases with absence of gene amplification and/or transcriptional overexpression (β-actin, loading control; colour signal from
NBT/BCIP treatment was discarded during image processing)
ZBTB7A expression is associated with ZBTB7A expression correlates positively with the
proliferation and tumour size status of p14ARF
The positive correlation with proliferation and tumour
Bearing in mind that ZBTB7A is a key player in size led us to investigate whether this effect was
a nodal cell cycle control network (Figure 1B), we mediated by a negative correlation of ZBTB7A with
investigated whether its overexpression was associ- its tumour-suppressor target p14ARF , as initially shown
ated with tumour kinetics and clinocopathological fea- in diffuse large B cell lymphomas [4].
tures of the carcinomas. In the subsequent analy- For this purpose, a serial section immunohisto-
ses, ZBTB7A expression was stratified according to chemical analysis of p14ARF was employed in our
tumour stage, in order to negate potential confounding set of carcinomas. The staining of p14ARF (range
influences. 5–85%, mean value 32.96 ± 22.45) was confined
Proliferation, as assessed by Ki67 staining (range mainly to the cancerous areas. In the respiratory
2–95%, mean value 33 ± 17%), was associated with epithelium, few differentiated bronchial cells gave a
ZBTB7A expression (ANOVA, p = 0.033; Figure positive signal. This finding is in agreement with pre-
5A), but not apoptosis, as determined by TUNEL assay viously published results [15]. In contrast to what
(range 0.1–6%, mean value 1.64 ± 1.1; ANOVA, we expected, the in situ analysis revealed a concor-
p = 0.176). The association with the proliferation dance between the expression patterns of ZBTB7A
index was reflected in the positive correlation between and p14ARF (Figure 6). This in situ observation was
ZBTB7A and tumour size (ANOVA, p = 0.018). also confirmed statistically. By stratifying ZBTB7A
ZBTB7A immunohistochemical expression progres- according to tumour stage, it was positively associ-
sively increased from T1 (mean 41%) to T2 (mean ated with p14ARF expression (ANOVA, p = 0.039).
55%) to T3 (mean 68%) (Figure 5B). However, Since p14ARF is considered an important sensor
there was no relation with lymph node stage (N; of aberrant proliferation [16], we assumed that the
ANOVA, p = 0.790). The overall pathological stage prominent ZBTB7A/p14ARF co-staining could be a
(IA–IIIA) was associated with ZBTB7A labelling result of various p14ARF -inducing signals overcom-
index (LI; ANOVA, p = 0.014). Somewhat unexpect- ing the suppressive activity of ZBTB7A. Among
edly, ZBTB7A expression was much higher in smok- the factors opposing the p14ARF -suppressive activity
ers (mean 55%) than in non-smokers (mean 30%) and of ZBTB7A was E2F1, previously shown by us to
the difference was significant (t-test, p = 0.011). be overexpressed in NSCLCs [9]. Staining of E2F1
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.300 K Apostolopoulou et al
a role in the pathophysiology of lung cancer. Its
key position as a specific negative regulator of the
nodal tumour-suppressor gene p14ARF [4] implies
that increased expression of ZBTB7A could inacti-
vate the p53 pathway by increasing the levels of
MDM2 (Figure 1B). In addition, it could comple-
ment the oncogenic properties of MDM2, which is
frequently aberrantly overexpressed in lung cancer
[11,17]. In a limited number of mutant p53 cases
we observed co-expression of ZBTB7A and MDM2,
supporting this notion (unpublished data). Moreover,
the high LI of ZBTB7A in carcinomas with mutated
p53 implies either that it targets p53-independent path-
ways [5] and/or that carcinomas harbouring both alter-
ations could posses a selective advantage in their
evolutionary trajectory. Another POK family mem-
ber, HIC-1 (hypermethylated i n cancer 1 ), which
forms a feedback loop with p53 [18,19], was shown to
develop a more aggressive malignant phenotype when
inactivated together with p53 [20]. The possibility
that ZBTB7A inactivates p53-independent targets was
based on the observation that it was overexpressed in
an unpublished series of bladder and colon carcinomas
[4], in which the p14ARF –p53 pathway is frequently
compromised [21,22]. The recently revealed ability of
POZ/BTB domain-containing proteins to interact with
cullin-based E3 ligase complexes could well constitute
a novel property of ZBTB7A, unrelated to p53. This
feature would enable aberrant ZBTB7A expression to
Figure 5. ZBTB7A expression is associated with tumour deregulate various cellular targets, via protein ubiq-
proliferation and tumour size. (A) Scatter plot depicting a uitination, as in the overexpression of other ubiquitin
direct relationship between ZBTB7A IHC expression and Ki67
IHC expression. (B) Box plot showing the positive association ligases [23].
between ZBTB7A IHC expression and tumour size Although aberrant overproduction of ZBTB7A
seems to characterize cancerous tissues, there are no
in our extended series (range 7–99%, mean value clues about the mechanism leading to its overexpres-
34.5 ± 18.32) showed in the serial section analysis sion. Based on the fact that MDM2, the other onco-
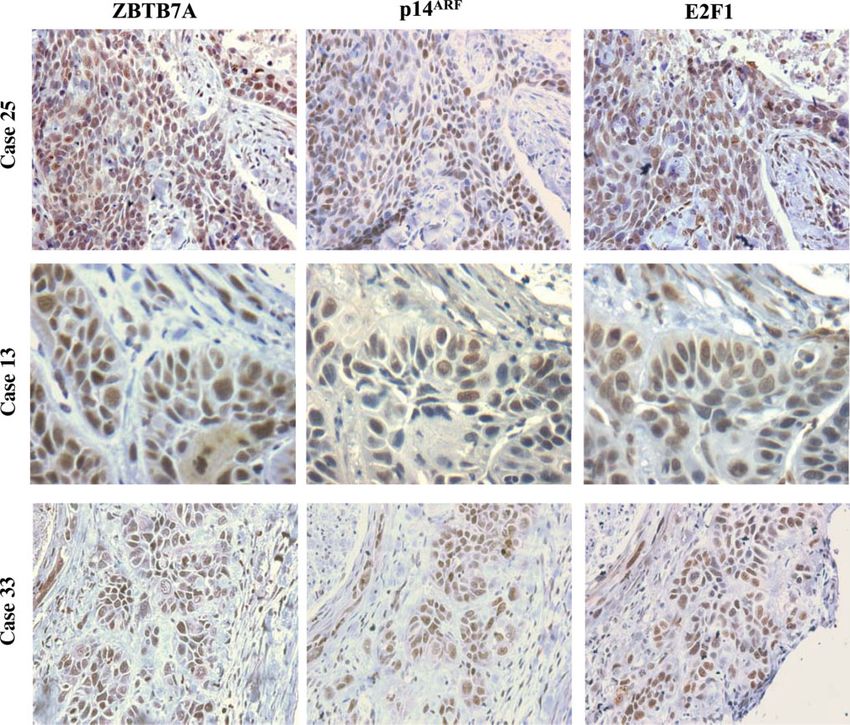
a remarkable concomitant expression with ZBTB7A gene of the p14ARF –p53–MDM2 network, is ampli-
and p14ARF , providing a putative explanation for the fied in various malignancies [6,7], it was tempting to
positive relationship between the two latter molecules speculate that a similar phenomenon, at least in part,
(Figure 6). may drive ZBTB7A overexpression as well. Moreover,
Subsequently, we examined ZBTB7A in relation ZBTB7A resides in the chromosomal locus 19p13.3, a
with p53 expression. From our previous studies, a gene-rich region (Figure 1A), frequently mutated in
strong correlation between p53 immunoreactivity and cancer [5,24], and a site involved in one of the most
the presence of p53 mutations was found [9,11,17]. We common cryptic translocations t(14;19)(p32;p13.3)
observed that ZBTB7A overexpression was equally found in B-cell non-Hodgkin’s lymphoma [25]. In
prevalent in both p53 mutant and p53 wild-type addition, 19p13 has been characterized as a rare fragile
tumours (p = 0.377), implying that ZBTB7A may also site (FRA19B) (NCBI Entrez Gene, Gene ID 2470),
act outside the p14ARF –MDM2–p53 axis. and is known to contain repeats that could facilitate
the process of gene amplification. Indeed, a signif-
icant proportion of the NSCLCs examined (27.7%)
Discussion showed a two- to five-fold amplification of ZBTB7A.
All amplified cases were accompanied by increased
The expression of ZBTB7A has been well examined mRNA and protein levels, strongly suggesting that
in various types of lymphoma where it was found to the amplification process may have functional conse-
be increased [4,5], but detailed information regarding quences. This finding is of great interest because gene
its status in solid tumours is lacking. amplification pinpoints candidate oncogenes or onco-
The predominant expression of ZBTB7A in NSCL- genic pathways that stem from a selection process that
cancerous areas, as assessed by immunohistochem- aims to promote tumour growth. This notion was fur-
istry and immunoblotting, suggests that it may play ther supported by the observation that the proliferation
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.ZBTB7A/Pokemon gene amplification in NSCLC 301
Figure 6. IHC analysis of ZBTB7A, p14ARF and E2F1 expression in consecutive sections of three representative NSCLCs. Note
the concordance between the expression patterns of ZBTB7A, p14ARF and E2F1 (case 13, ×400; cases 25 and 33, ×200)
index and tumour size of the carcinomas was pro- sensor of aberrant proliferation [16], a possible expla-
portional to ZBTB7A immunoexpression, which was nation for this intriguing observation is that p14ARF is
higher in the amplified cases (Figures 3, 5). up-regulated by various incoming oncogenic signals
Although gene amplification could explain the over- overcoming the suppressive activity of ZBTB7A. In
production of ZBTB7A in a subset of our cases, this context, E2F1, a potent positive regulator of
the remaining subgroup of non-amplified carcinomas p14ARF [8] frequently found overexpressed in lung
(72.3%) showed overexpression of ZBTB7A as well. cancer [9], was co-expressed with p14ARF (Figure 6).
As shown in Figure 4, approximately 62% of these In conclusion, overexpression of ZBTB7A is a
samples had increased mRNA levels, possibly as a prominent phenomenon in NSCLCs and is frequently
result of enhancement at the transcriptional level. This driven by gene amplification. In practice, this could
could be achieved by constant stimulation from the confer a growth advantage, as demonstrated by the
upstream signalling pathways, which regulate ZBTB7A positive association with proliferation and tumour size.
expression and are relatively less well studied. Puta-
Acknowledgements
tive stimulators could be either growth factor receptors
or adhesion molecules similar to those controlling the This work was co-funded from the European Social Fund and
POK family member Kaiso [2]. Recently, ZBTB7A National Resources–(EPEAEK-II) PYTHAGORAS II (Grant
No. 7953) and SARG, University of Athens (Grant Nos
was reported to be up-regulated upon fibronectin- 73/17/06 and 70/4/4281).
mediated β1-integrin ligation in precursor B leukaemia
cells, providing some evidence about its upstream
signalling [26]. In the remaining group with normal Supplementary material
mRNA levels, decreased protein degradation could be
the reason for high ZBTB7A levels. Supplementary material may be found at the web
address: http://www.interscience.wiley.com/jpages/
An unexpected finding of the present study was the
0022-3417/suppmat/path.2222.html
positive relationship between ZBTB7A and p14ARF
expression. This was particularly clear in the serial
section analysis (Figure 6). Given that ZBTB7A References
suppresses the transcription of ARF [4], an inverse 1. Stogios PJ, Chen L, Prive GG. Crystal structure of the BTB
expression pattern between the two molecules was domain from the LRF/ZBTB7 transcriptional regulator. Protein
anticipated. Since p14ARF is considered a sensitive Sci 2007;16:336–342.
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.302 K Apostolopoulou et al
2. Kelly KF, Daniel JM. POZ for effect — POZ-ZF transcrip- p19ARF/p16β is frequently altered in non-small cell lung
tion factors in cancer and development. Trends Cell Biol cancer and correlates with p53 overexpression. Oncogene
2006;16:578–587. 1998;17:2779–2785.
3. Schubot FD, Tropea JE, Waugh DS. Structure of the POZ domain 16. Lowe SW, Cepero E, Evan G. Intrinsic tumour suppression.
of human LRF, a master regulator of oncogenesis. Biochem Nature 2004;432:307–315.
Biophys Res Commun 2006;351:1–6. 17. Gorgoulis VG, Zacharatos P, Kotsinas A, Liloglou T, Kyroudi A,
4. Maeda T, Hobbs RM, Merghoub T, Guernah I, Zelent A, Cordon- Veslemes M, et al. Alterations of the p16–pRb pathway and the
Cardo C, et al. Role of the proto-oncogene Pokemon in cellular chromosome locus 9p21–22 in non-small-cell lung carcinomas:
transformation and ARF repression. Nature 2005;433:278–285. relationship with p53 and MDM2 protein expression. Am J Pathol
5. Maeda T, Hobbs RM, Pandolfi PP. The transcription factor 1998;153:1749–1765.
Pokemon: a new key player in cancer pathogenesis. Cancer Res 18. Wales MM, Biel MA, el Deiry W, Nelkin BD, Issa JP, Cave-
2005;65:8575–8578. nee WK, et al. p53 activates expression of HIC-1, a new candidate
6. Deb SP. Cell cycle regulatory functions of the human oncoprotein tumour suppressor gene on 17p13.3. Nat Med. 1995;1:570–577.
MDM2. Mol Cancer Res 2003;1:1009–1016. 19. Chen WY, Wang DH, Yen RC, Luo J, Gu W, Baylin SB. Tumour
7. Iwakuma T, Lozano G. MDM2, an introduction. Mol Cancer Res suppressor HIC1 directly regulates SIRT1 to modulate p53-
2003;1:993–1000. dependent DNA-damage responses. Cell 2005;123:437–448.
8. Komori H, Enomoto M, Nakamura M, Iwanaga R, Ohtani K. 20. Chen W, Cooper TK, Zahnow CA, Overholtzer M, Zhao Z,
Distinct E2F-mediated transcriptional program regulates p14ARF Ladanyi M, et al. Epigenetic and genetic loss of Hic1 function
gene expression. EMBO J 2005;24:3724–3736. accentuates the role of p53 in tumourigenesis. Cancer Cell
9. Gorgoulis VG, Zacharatos P, Mariatos G, Kotsinas A, Bouda M, 2004;6:387–398.
Kletsas D, et al. Transcription factor E2F-1 acts as a growth- 21. Le Frere-Belda MA, Gil Diez de Medina S, Daher A, Martin N,
promoting factor and is associated with adverse prognosis in non- Albaud B, Heudes D, et al. Profiles of the 2 INK4a gene products,
small cell lung carcinomas. J Pathol 2002;198:142–156. p16 and p14ARF, in human reference urothelium and bladder
10. Pateras IS, Apostolopoulou K, Koutsami M, Evangelou K, Tsan- carcinomas, according to pRb and p53 protein status. Hum Pathol
toulis P, Liloglou T, et al. Down-regulation of the KIP family 2004;35:817–824.
members p27(KIP1) and p57(KIP2) by SKP2 and the role of 22. Lee M, Sup Han W, Kyoung Kim O, Hee Sung S, Sun Cho M,
methylation in p57(KIP2) inactivation in non-small cell lung can- Lee SN, et al. Prognostic value of p16INK4a and p14ARF gene
cer. Int J Cancer 2006;119:2546–2556. hypermethylation in human colon cancer. Pathol Res Pract
11. Gorgoulis VG, Zacharatos P, Kotsinas A, Mariatos G, Liloglou T, 2006;202:415–424.
Vogiatzi T, et al. Altered expression of the cell cycle regulatory 23. Bashir T, Pagano M. Aberrant ubiquitin-mediated proteolysis of
molecules pRb, p53 and MDM2 exert a synergetic effect on cell cycle regulatory proteins and oncogenesis. Adv Cancer Res
tumour growth and chromosomal instability in non-small cell lung 2003;88:101–144.
carcinomas (NSCLCs). Mol Med 2000;6:208–237. 24. Peng WX, Shibata T, Katoh H, Kokubu A, Matsuno Y, Asamura
12. Morrison DJ, Pendergrast PS, Stavropoulos P, Colmenares SU, H, et al. Array-based comparative genomic hybridization analysis
Kobayashi R, Hernandez N. FBI-1, a factor that binds to the HIV- of high-grade neuroendocrine tumours of the lung. Cancer Sci
1 inducer of short transcripts (IST), is a POZ domain protein. 2005;96:661–667.
Nucleic Acids Res 1999;27:1251–1262. 25. Gozzetti A, Davis EM, Espinosa R III, Fernald AA, Anastasi J,
13. R Development Core Team. R: A Language and Environment for Le Beau MM. Identification of novel cryptic translocations
Statistical Computing. R Foundation for Statistical Computing: involving IGH in B-cell non-Hodgkin’s lymphomas. Cancer Res
Vienna, 2005. 2002;62:5523–5527.
14. Pessler F, Pendergrast PS, Hernandez N. Purification and char- 26. Astier AL, Xu R, Svoboda M, Hinds E, Munoz O, de Beau-
acterization of FBI-1, a cellular factor that binds to the human mont R, et al. Temporal gene expression profile of human
immunodeficiency virus type 1 inducer of short transcripts. Mol precursor B leukaemia cells induced by adhesion receptor:
Cell Biol 1997;17:3786–3798. identification of pathways regulating B-cell survival. Blood
15. Vonlanthen S, Heighway J, Tschan MP, Borner MM, Alter- 2003;101:1118–1127.
matt HJ, Kappeler A, et al. Expression of p16INK4a/p16α and
J Pathol 2007; 213: 294–302 DOI: 10.1002/path
Copyright 2007 Pathological Society of Great Britain and Ireland. Published by John Wiley & Sons, Ltd.You can also read

















































