The Evolutionary Path from C3 to C4 Photosynthesis: A Review
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
International Journal of Current Microbiology and Applied Sciences
ISSN: 2319-7706 Volume 9 Number 1 (2020)
Journal homepage: http://www.ijcmas.com
Review Article https://doi.org/10.20546/ijcmas.2020.901.082
The Evolutionary Path from C3 to C4 Photosynthesis: A Review
Priyanka Upadhyay1*, Neha Agrawal1, Praveen Kumar Yadav2 and Ruby Patel3
1
Department of Plant Breeding and Genetics, Punjab Agricultural University, Ludhiana,
Punjab India, 141004
2
Division of Seed Science and Technology, Indian Agricultural Research Institute, New
Delhi, 110012, India
3
Department of Agricultural Chemistry and Soil Science, Bidhan Chandra Krishi
Vishwavidyalya, Mohanpura, Nadia West Bengal India, 741252
*Corresponding author
ABSTRACT
The C4 photosynthetic carbon cycle can be explained as the elaborated
addition to the C3 photosynthetic pathway. It is a unique series of
biochemical, anatomical and regulatory gene characteristics that
concentrate CO2around the carboxylating enzyme Rubisco, thereby
Keywords
increasing photosynthetic efficiency during high rates of photorespiration.
C4 photosynthetic The C4 photosynthetic pathway has evolved as an adaptation to high light
pathway, C3- intensities, high temperatures and dryness, therefore in the warmer climates
C4transition,
photorespiration, of the tropical and subtropical dominating the grassland floras and biomass
genome production. C4 photosynthesis originated more than 40 times independently
duplications, during angiosperm evolution in over 15 families of angiosperms,
Rubisco
representing convergent evolutionary phenomena. Probably, C4 grasses
Article Info evolved in the early Oligocene about 30 million years ago, while later
Accepted: appeared C4 dicots, less than 20 million years ago. Low atmospheric CO2 is
15 December 2019 pivotal factor forC3- C4 transition, because it is required for high rates of
Available Online:
20 January 2020 photorespiration. Consistently, the increasing global aridification and
declining atmospheric CO2 coincides with periods of the appearance of
C4plants. Genetically, leading mechanism for creation of C4 genome is
duplications of whole genomes, genome segments, or single genes followed
by non-functionalization and neo-functionalization with selection for
carbon conservation traits under high photorespiration promoting
conditions being the ultimate factor behind the origin of C4 photosynthesis.
748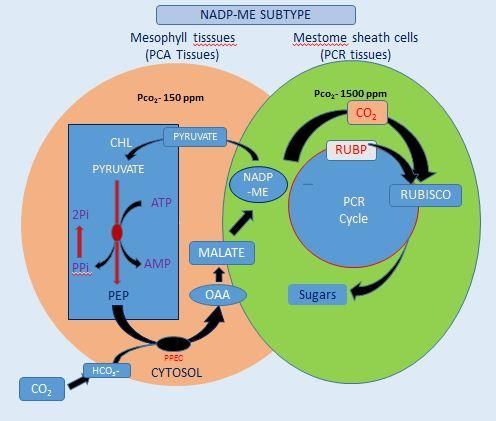
Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
Introduction property adds oxygen to ribulose-1,5-
bisphosphate, resulting in one molecule each
The ability of photosynthetic organisms to of 3-phosphoglycerate and 2-
sequester inorganic CO2 of the atmosphere phosphoglycolate. The metabolic purpose of
into organic carbon of the biosphere via the phosphoglycolate is unknown and in higher
Calvin- Benson-Bassham pathway is pivotal concentrations it is toxic for the plant(a potent
for the existence of life on the earth. The C4 inhibitor of chloroplastic function) (Anderson,
photosynthetic carbon cycle can be explained 1971). Therefore, it has to be processed in a
as the elaborated addition to the C3 metabolic pathway called photorespiration.
photosynthetic pathway. It is a unique series Besides energy demanding, photorespiration
of biochemical, anatomical and regulatory leads to a net loss of CO2. The catalytic
gene characteristics. In most terrestrial C4 activity of Rubisco with O2 as a substrate is
species, it relies on the co-ordinated some 100-fold lower than with CO2 at
functioning of mesophyll and bundle sheath equivalent concentrations of the two gases
cells, except in the chenopods Borszczowia (Tcherkez et al., 2006).Thus, under
aralocaspia and Bientertia cycloptera where unfavorable conditions including high
the C4 photosynthetic mechanism operates in temperatures and dryness the efficiency of
single cells (Freitag and Stichler 2000; photosynthesis can be decreased by 40%
Voznesenskaya et al., 2001, 2002). The C4 (Ehleringer et al., 1991) by decrease in the
photosynthetic pathway has evolved as an affinity of Rubisco for CO2 (Jordan and
adaptation to high light intensities, high Ogren 1984). The remaining reduced level of
temperatures, and dryness. Therefore, CO2 and abundant availability of O2 inside the
grassland floras and biomass production in leaf results in enhanced rates of RuBP
the warmer climates of the tropical and oxygenation.
subtropical regions has been dominated by C4
plants (Edwards et al., 2010). The unfavorable oxygenase reaction of
Rubisco evolved more than 3 billion years
In all plants, the primary photosynthetic CO2 ago when atmospheric CO2 concentrations
reduction reaction is the fixation of CO2 by were high and oxygen concentrations low can
the enzyme RuBP carboxylase/oxygenase be explained as a relict of the evolutionary
(Rubisco). Rubisco is a ubiquitous enzyme in history of this enzyme. Later on, enzyme’s
most autotrophic organisms from prokaryotes alteration or to exchange Rubisco by another
(photosynthetic and chemoautotrophic carboxylase was impossible. Indeed, plants
bacteria, cyanobacteria and archaea), to developed different ways to cope with this
eukaryotes (various algae and higher plants) problem. Perhaps the most successful solution
and even phytoplankton in the sea (providing was C4 photosynthesis. A marked and
more than 45% of global net primary sustained decline in atmospheric pCO2during
production annually). Approximately, the Tertiary period reveals the appearance of
Rubisco comprises up to 50% of the total C4 plants in the fossil record (Ehleringer et
soluble protein in the plant leaf or inside the al., 1991; Sage 2001). Therefore, low pCO2
microbe. might have been an important driving force
for evolution of the pathway.
The RuBP carboxylase reaction catalyzes the
carboxylation of ribulose-1,5-bisphosphate, A complex combination of both biochemical
leading to two molecules of 3- and morphological specialization provides
phosphoglycerate, while its oxygenase elevated pCO2 at the site of Rubisco
749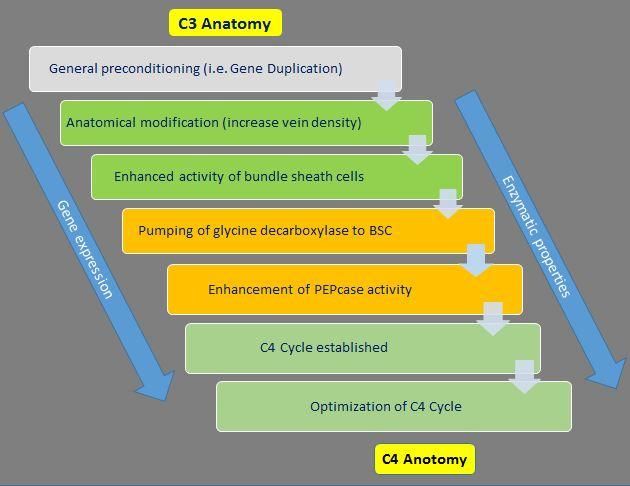
Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
carboxylation in all C4 species. This result in Rubisco refixes the released CO2 in the
suppression of photorespiration and allowing bundle sheath cells. This results in release of
Rubisco to work near to its maximal rate, CO2 and a three-carbon compound, which
such that CO2 assimilation in C4plants is diffuses back to the mesophyll cells. Here, at
effectively CO2 saturated in air (Hatch the end the primary CO2 acceptor PEP is
1987).In most C4 plants a division of labor regenerated by pyruvate orthophosphate
between two distinct, specialized leaf cell dikinase by the consumption of two
types, the mesophyll and the bundle sheath molecules of ATP (Hatch, 1987). In NADP-
cells leads to the CO2 concentration ME subtype of C4 photosynthesis malate is
mechanism except in some species where C4 the dominant transport metabolite while Asp
functions within individual cells(Edwards et can be used in parallel. Malate synthesis
al., 2004). Since, Rubisco works more occurs in the mesophyll chloroplasts and the
efficiently in C4 than C3 plants and can decarboxylation by NADP-ME in the bundle
operate under high CO2 concentrations in the sheath chloroplasts (Fig. 2).
bundle sheath cells. Indeed, C4 species are
also characterized by greater nitrogen and The other two biochemical subtypes are
water use efficiency relative to C3 plants. The differentiated from the NADP-ME type by the
increased nitrogen use efficiency is largely transport metabolites used and the subcellular
accounted for saving nitrogen in Rubisco localization of the decarboxylation reaction.
protein (Evans and von Caemmerer 2000) In NAD-ME plants Asp is used as transport
because C4 plants need less of this enzyme, metabolite which is synthesized in the
however in C3 plants leaves it is the most mesophyll cytosol. Deamination and
abundant protein. Since the rate of reduction of Asp results in malate which is
photosynthesis per unit nitrogen in the leaf is decarboxylated by NAD-ME in the bundle
increased in C4 species (Oaks 1994). Better sheath mitochondria. Plants of the PEPCK
water use efficiency of C4 relative to C3 plants type use Asp as well as malate as transport
accounted from its CO2 concentration metabolites.
mechanism even when keeping their stomata
more closed. Thus reducing the water loss by Asp is synthesized in the cytosol of mesophyll
transpiration (Long 1999). cells and decarboxylated in the cytosol of
bundle sheath cells by the combined action of
Initially, in the mesophyll cells of C4 plants Asp amino transferase and PEPCK. This
CO2 is converted to bicarbonate by carbonic reaction produces NADH that is used in the
anhydrase, which is then fixed by mitochondria to produce the ATP needed to
phosphoenolpyruvate (PEP) carboxylase into drive the PEPCK reaction (Hatch 1987).
C4 acids(oxaloacetate) using PEP as CO2 When Asp is used as transport metabolite,
acceptor. Oxaloacetate is rapidly converted to usually, pyruvate (the three-carbon
the more stable C4 acids malate or Asp that decarboxylation product) is partially
diffuses to the bundle sheath cells and is transported back to the mesophyll cells in
decarboxylated there to supply CO2 for Alaform to maintain the ammonia balance
Rubisco. Three basic biochemical subtypes of between the two cell types (Hatch, 1987).
C4 photosynthesis are defined by one of three
one of three different decarboxylating Resistance of the bundle sheath to CO2
enzymes: NADP-dependent malic enzyme diffusion and the relative biochemical
(NADP-ME), NAD-dependent ME (NAD- capacities of the C3 and C4 cycle determine
ME), and PEP carboxykinase (PEPCK). the efficiency of the C4 pathway and the pCO2
750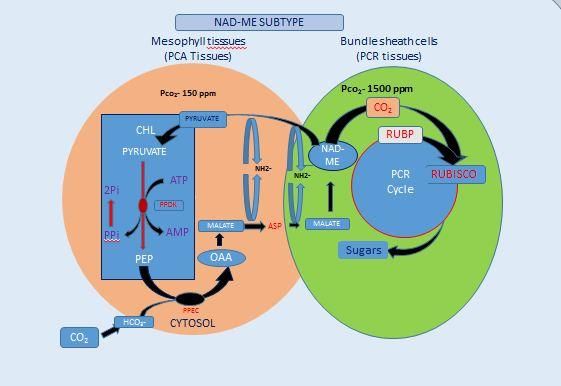
Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
attained in the bundle sheath. The leakiness of Polyphyletic evolution and quantitative
the bundle sheath is intimately linked with the trait of C4 photosynthesis
efficiency of the C4 concentrating
mechanisms. Leakiness is defined as that C4 photosynthesis originated more than 40
fraction of CO2 generated by C4 acid times independently during angiosperm
decarboxylation in the bundle sheath that evolution(Gowik and Westhoff, 2011). Most
subsequently leaks out (Farquhar 1983). of the C4 species occur in the grassesand
Consumption of energy by C4 cycle in ATP sedges approximately 4,600 and 1,600
form during regeneration of PEP, CO2 leakage respectively, whereas only about 1,600
from bundle sheath is an energy cost to the C4 dicots species are known. They are spread
leaf. High energy demand of C4 cycle relative over 15 families with 75% of them clustering
to C3 cycle was demonstrated with quantum in the four families Chenopodiaceae,
yield measurements under varying pCO2, pO2 Amaranthaceae, Euphorbiaceae, and
and temperature by Ehleringer and Bjorkman Asteraceae (Muhaidat et al., 2007),
(1977). These data represented that at low representsing convergent of evolutionary
temperatures C4 species have lower quantum phenomena. Probably, C4 grasses evolved in
yields than C3 species, but at high temperature the early Oligocene about 30 million years
superior quantum yields where in C3 species ago, while later appeared C4 dicots, less than
the quantum yields decrease due to high 20 million years ago (Sage 2004). The
photorespiratory rates. polyphyletic origin of C4 photosynthesis
indicates that only relatively small
The significant parameters of the C4 evolutionary changes were required for the
concentrating mechanism, such as bundle establishment of this photosynthetic pathway.
sheath resistance to CO2 diffusion, bundle C4 evolution can be assumed in genetic terms,
sheath pCO2and leakiness of the bundle which raises the question of whether we can
sheath measurement cannot be done directly use the information about the genetic
and these estimates vary widely. It has been architecture and evolution of this pathway and
demonstrated through various models that a introduce modules of C4-ness into present C3
low bundle sheath conductance is pivotal plant and thereby transform them into C3-C4
feature of the C4 photosynthetic pathway intermediate or even C4-like plants (Sheehy et
(Berry and Farquhar, 1978; von Caemmerer al., 2007).
and Furbank, 1999). The conductance across
the mesophyll/bundle sheath interface and the The transition from C3 to C4 photosynthesis
bundle sheath surface area to leaf area ratio pathway
(Sb) are the basis for expression of
conductance bundle sheath. An estimate of Sb The transition from C3 to C4 and the
ranges from 0.6-3.1 m2 m-2 (Apel and Peisker occurrence of C3-C4 intermediate species in
1978; Brown and Byrd 1993). today’s flora provides us an evolutionary
adaptive advantage for the resulting species
Neverthless the conductance to CO2 diffusion independent of whether it will progress
across the mesophyll bundle sheath interface toward the full expression of the C4
is also several-fold relative to equivalent syndrome. Recently most widely accepted
conductance across the cell wall and model of C4 evolution proposes a stepwise
chloroplast interface in C3 species (Evans and sequence of changes leading from C3 to C4
vonCaemmerer 1996; von Caemmerer and plants (Fig. 3).
Furbank 2003).
751Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
Genetically the C4 syndrome may therefore be compartmentalized expression established the
best described as a polygenic, quantitative C4 cycle between mesophyll and bundle
trait. The concept of C4 photosynthesis being sheath cells. Massive changes in gene
as a quantitative trait immediately implies a regulation accompanied all these steps. The
number of questions that what is the genetic kinetic properties of enzymes also involved in
architecture of C4 photosynthesis, i.e. how metabolic pathways and were affected by
many genes are required to establish this these evolutionary changes, adapted to the
phenotypic syndrome? Are the genes new requirements.
organized into functional units giving rise to
distinct subphenotypes? Do these functional Kranz anatomy development
units form gene regulatory networks whose
component genes are regulated coordinately The most significant feature towards C4
and hence may be viewed as separate evolution was the development of the Kranz
regulatory modules? anatomy. Shortest distance between
mesophyll cell and to the next bundle sheath
Here, we only present a short summary and cell is pivotal for establishing an efficient CO2
elucidate how the evolutionary changes might concentrating mechanism. Therefore, in
have been realized through modifications at planar leaves enhancement of the vein density
the molecular/genetic level. is essential. A higher vein density also
increased the mechanical integrity of the
Genetically, C4 evolution began with leaves, which could be beneficial in windy
duplications of whole genomes, genome habitats, or improved the water supply of
segments, or single genes followed by non- leaves in dry and hot biotopes (Sage 2004).
functionalization and neo-functionalization
(Monson 2003). Thus redundant gene copies Considerably leaf architecture may vary in the
prevent deleterious consequences of various mono and dicotyledonous C4 lineages.
evolutionary changes that alter or switch off Typically all C4 plants have a wreath-like
the specific function of a certain gene. The structure of mesophyll and bundle sheath cells
non- and neo-functionalization’s major targets around the vascular bundles (Kranz
are the promoter and enhancer region of genes anatomy).Location of mesophyll cells are
to allow for altered expression and always toward the outer face of the leaf and
compartmentalization, and the coding region so remain in contact with the intercellular air
to alter regulatory and catalytic properties. space, while bundle sheath cells arrangement
are internal to the mesophyll cells and hence
Further at anatomic level, leaf modification close to the vascular tissues. The high
occurred toward Kranz anatomy. A densities of plasmodesmata lead to mesophyll
rudimentary Kranz anatomy resulted from an and bundle sheath cells of C4 species in close
increase in vein density and an enhancement proximity (Dengler and Nelson 1999).
and activation of the bundle sheath cell layer.
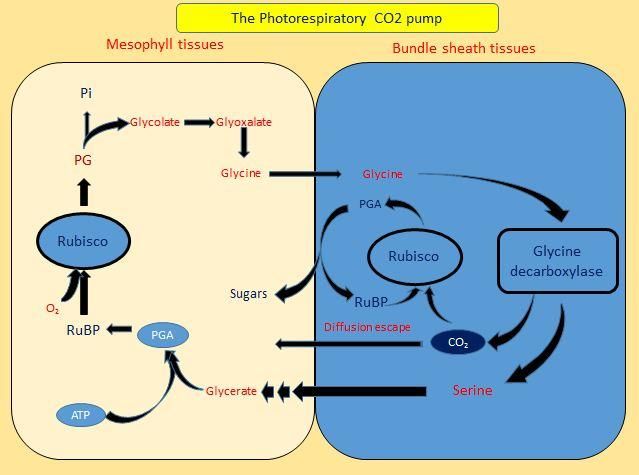
The compartmentation of Gly decarboxylase A comparative analysis of the leaf
in the bundle sheath cells was the next step development in both monocot and dicot C3
which led to a photorespiratory CO2 pump and C4 species revealed that the close vein
(Fig. 4). An elevated phosphoenolpyruvate spacing in leaves of C4 plants is due to
carboxylase activity and subsequently an changes in the initiation frequency and
increase in the other C4 cycle enzymes and pattering of the minor and not the major veins
transporters accompanied by their (Ueno et al., 2006; McKown and Dengler
752Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
2009).The greater vein density observed in C4 photosynthetic CO2 fixation in two ways.
compared to C3leaves resulted from either Inside the leaf all photorespiratory CO2 is set
modifications of auxin production or free far apart from the outer surface.
allocation and/or modifications of the Therefore several cell layers diffusion is
competency of ground tissue cell to become necessary, before it could escape from the
procambial cells (McKown and leaf. Therefore it enhances chances of refixing
Dengler2009). Since the molecular events the photorespired CO2the plants and
causing the initiation of veins are not clear in minimizes the loss of carbon due to
C3 model plants, so it is presently challenging photorespiration. Since the mitochondria
to predict the changes that led to the C4 concentrate adjacent to the vascular bundles
typical leaf anatomy. thus in some C3-C4 intermediate species this
refixation capacity is supported by the spatial
Typically, C3 plants bundle sheath cells have distribution of the organelles within the
low photosynthetic activity because they bundle sheath cell (Rawsthorne et al., 1998).
possess only a few chloroplasts. Bundle Besides, the Gly shuttle enhances the CO2
sheath to mesophyll cells ratio increases with concentration within thebundle sheath cells.
higher vein densities. Since only the As a consequence, the carboxylation activity
mesophyll cells show high photosynthetic of Rubisco in the bundle sheath cells
activity of a leaf, with a given size decreases. increases, while its oxygenase reaction is
The evolutionary pressure could have led to outcompeted (Bauwe, 2010). Photorespiratory
an increase of the number of chloroplasts in CO2 pumps occur in some two dozen species
the bundle sheath cells to maintain the overall in Alternanthera (Amaranthaceae), Panicum
photosynthetic activity. For metabolizing the and Neurachne (Poaceae), Parthenium
photorespiratoryGly in bundle sheath cells an (Asteraceae), Moricandia (Brassicaceae) and
enhancement of chloroplast numbers in cellsis Flaveria (Asteraceae) (Monson, 1999). It is
necessary, which would also require an assumed that the establishment of such a
increase in the numbers of mitochondria and photorespiratory CO2 pump is an important
peroxisomes. intermediate step on the way toward C4
photosynthesis.
The Photorespiratory CO2 Pump during
Transition of C3-C4 Photosynthesis A photorespiratory CO2 pump can easily be
accomplished at the molecular level. Gly
Photorespiratory metabolites are a carbon decarboxylase multienzyme complex encoded
source that can be exploited to improve the by single gene expression had to be restricted
efficiency of Rubisco in C3 leaves (Hunt et to the bundle sheath cells. This might had
al., 1987; von Caemmerer, 1989; Rawsthorne, been achieved through relatively subtle
1992). A photorespiratoryGly shuttle is a changes in the cis-regulatory elements that
common feature in all extant C3–C4 control the expression of these genes
intermediate that pumps CO2 into the (Akyildiz et al., 2007).
bundlesheath cells (Bauwe, 2010). This is
obtained by localising the Gly In Moricandia arvensis (theC3-C4
decarboxylation reaction to the bundle sheath intermediate species) for example, only the P
mitochondria, thus all Gly produced by subunit of Gly decarboxylaseis restricted to
photorespiration in the mesophyll has to be the bundle sheath. Since the enzyme is
transferred to the bundle sheath cells for inactive without this subunit, Gly cannot be
further processing. The Gly shuttle affects decarboxylated in the mesophyll (Rawsthorne
753Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
et al., 1988). For other C3-C4 intermediates bundle sheath cells. Enhancement of
from the genera Flaveria and Panicum, it was chloroplastic pyruvate orthophosphate
found that the other subunit genes werealso dikinase expression might have occurred for
expressed specifically or at least preferentially allowing an efficient PEP regeneration and to
in the bundle sheath cells (Morgan et al., complete the C4 cycle. In this phase of C4
1993). evolution plants exhibit high activities of C4
cycle enzymes, but still in the mesophyll cells
Relative to C3 species, the intermediates also Rubisco has high activity. Consequently, CO2
exhibit close vein spacing, enlarged bundle is only partially fixed through the C4
sheath cells, increased frequency of pathway.
plasmodesmata between mesophyll and
bundle sheath cells, and increased number of The C4 cycle establishment
organelles in the bundle sheath cells (Brown
et al., 1983; Rawsthorne, 1992). These The spatial separation of the two
developments facilitated efficient function of carboxylation reactions was the key step in
the glycine shuttle by reducing diffusion establishing true C4 photosynthesis and to
distances, increasing intercellular transport, integrate the C4 and Calvin-Benson cycle.
and enhancing metabolic capacity in the PEPC was restricted to the mesophyll and
bundle sheath. In doing so, they established Rubisco to the bundle sheath cells. This step
the anatomical and ultrastructural framework was necessary when the C4 cycle activity
required for the subsequent evolution of C4 increased to such a level that CO2 fixation by
metabolism. PEPC reached the same magnitude as by
Rubisco and hence the C4 and the Calvin-
An increase in the levels of carbonic Benson cycle competed for CO2and ATP
anhydrase and PEPC in the cytosol of the (Monson, 1999). Now photo assimilated CO2
mesophyll cells might have been the next step in the vast majority passed initially through
toward true C4photosynthesis. This would the C4cycle before it was fixed by Rubisco.
have accompanied in recapturing the The evolving C4 pathway was further
photorespiratoryCO2 that escaped from the optimized by compartmentalizing other
bundle sheath into the mesophyll cells. Also enzymes of both the C4 and Calvin-Benson
this evolutionary step is reflected by C3-C4 cycles, by adapting the light reaction of
intermediate species of the genus Flaveria, photosynthesis and by strongly increasing
which contain significantly higher levels in carbonic anhydrase activity in the cytosol of
PEPC transcript and protein amounts as mesophyll cells. Characteristic of the C4
compared to Flaveria species (C3 plant) which photosynthetic pathway are determined by the
do not exhibit C4cycle activity yet (Ku et al., extensive shuffling of metabolites within
1991; Engelmann et al., 2003). mesophyll and bundle sheath cells of
organelles and the cytosol respectively. C4
The remaining C4 cycle enzymes must have pathway evolution requires the proper
been elevated to establish a limited C4 cycle establishment of transport capacity. For
activity. It is suggested that even in C3 plants instance, in NADP-ME type plants for every
the activity of the decarboxylating enzymes fixation of CO2 molecule, one molecule of
NADP-ME and NADME is massively pyruvate and oxaloacetate have to be
increased in vascular tissues (Hibberd and transported into the mesophyll chloroplasts
Quick 2002). Therefore the related genes and in a countermove PEP and malate have to
expression must have been shifted to the be translocated to the cytosol. On the other
754Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
hand, in the bundle sheath cells entry of about 18% of the genes in maize (Zea mays)
malate leads to exit of pyruvate from are differentially expressed between
chloroplast matching the CO2 assimilation mesophyll and bundle sheath cells. It showed
rate. Transcriptome and proteome analyses at that the establishment of C4 photosynthesis
large scale shows that other pathways related involved a dramatic redesign and
to sulfur, nitrogen, and carbon metabolism restructuring of leaf functions. At the
were also altered with respect to either overall molecular level, most of the quantitative and
activity or to mesophyll/bundle sheath qualitative changes in gene expression are not
compartmentation (Friso et al., 2010; yet understood and only a few have been
Brautigam et al., 2011). It was might be analyzed in great detail. These things
because of energy supply difference and demonstrate the flexible nature in achieving
reduction equivalents in the different tissues the desired goal, i.e. different alteration
and for optimization of overall integration of pattern for different genes for their adaption
the various metabolic pathways. and functioning in the C4 pathway (Hibberd
and Covshoff, 2010).
Alterations at Gene Expression level
Transcriptional control can help to achieve
C4 photosynthesis evolution was accompanied cell-specific gene expression. For example,
by massive alteration in quantitative and specific gene expression of the photosynthetic
spatial gene expression. The quantitative PEPC of the mesophyll, the C4 plant Flaveria
alterations in C4 evolution can be observed in trinervia ppcA depends on a cis-regulatory
Cleome species. When the transcriptomes of element, the MESOPHYLL EXPRESSION
mature leaves of the C4 plant Cleome MODULE1, whose location is about 1,900 bp
gynandra and the closely related C3 species upstream of the transcriptional initiation site
Cleome spinosa were quantitatively compared (Gowik et al., 2004). In the C3Flaverias a very
by a RNA-Seq-based digital gene expression similar element was found in the promoters of
approach, then about 2.8% of the detected the orthologous ppcA genes; however, direct
transcripts differed significantly in their mesophyll specificity was lacking in these
abundance between the two species elements. Thus, for a gene with no apparent
(Brautigam et al., 2011). It can be expected expression specificity into a mesophyll, slight
that the expression levels of genes in the C4 modifications within a cis-regulatory element
cycle, thephotorespiratory pathway, and the were sufficient to convert them into a
photosynthetic light reactions including mesophyll-specific gene (Akyildiz et al.,
several other pathways also changed. It can be 2007).
seen in the C4 Cleome, which showed reduced
steady-state levels transcripts for the In contrast, regulation mainly at the
shikimate pathway, and amino acid posttranscriptional level was reported for the
metabolism (Brautigam et al., 2011). The C4 bundle sheath-specific expression of one of
species cytosolic and plastidic protein the genes encoding the small subunit of
synthesis machinery encoding components Rubiscoin the C4 plant Flaveria bidentis
genes are down-regulated, while the genes (Patel et al., 2006). Most likely, in mesophyll
involved in starch metabolism, cofactor and bundle sheath cells the FbRcS1
synthesis, and nitrogen metabolism showed transcripts are differentially stable. This is
higher steady-state transcript levels in C4 leaf controlled by stability determinants that are
(Brautigam et al., 2011). The Spatial gene located in the 5’ and 3’ untranslated regions
expression alteration patterns can be seen in of the mRNA (Patel et al., 2006).
Maize. Sawers et al., (2007) reported that
755Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
Figure.1 C4 Photosynthesis
Figure.2 A. The NADP-ME grass Themadatriandra leaf structure and C4 metabolic pathways of
with the PCR tissue in the mestome sheath (left panel) and B. The NAD-ME grass Panicum
effusum with the PCR tissue in the bundle sheath layer that resides outside
the mestome sheath (right panel)
A. B.
Figure.3 Transition from C3 to C4 Photosynthetic Pathway
756Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
Figure.4 The photorespiratory CO2 pump
The transition from C3 to C4 photosynthes is it can be concluded that the regulating
associated with the massive changes in gene networks for developmental and metabolic
expression was associated with the fact that processes operated already in C3 ancestral
C4 evolution must have been easy in genetic angiosperms and could serve as a platform for
terms implies that preexisting gene regulatory the establishment of C4 leaf anatomy and
networks in C3 plants were probably the metabolism.
foundation for multiple evolutionary changes
toward C4photosynthesis (Matsuoka 1995). In Since our understanding of gene regulatory
C3 plants the gene regulatory networks assure networks controlling the development and
a coordinated response of genes involved in anatomy of a typical leaf of a C3 angiosperm
photosynthesis and related metabolic is not explicit. We know little about the
pathways (Mentzen and Wurtele, 2008). The molecular nature of cis-and trans-regulatory
promoters driving mesophyll or bundle sheath factors that regulate gene expression in the
specific gene expression in C4 species partly mesophyll and bundle sheath cells of both C3
maintain their cell preference of expression in and C4 plants except the things discussed
C3 species (Matsuoka et al., 993; Engelmann above. The only exceptions are the
et al., 2008), suggesting that the C4 plants GOLDEN2-LIKE (GLK) transcription factors
gene regulatory networks controlling the GLK1 and GLK2. In all land plants this pair
development and differentiation of mesophyll of transcription factors occurs. The GLK
and bundle sheath cells are not fundamentally proteins are largely redundant in Arabidopsis
different from those of C3 species. Therefore, and control the expression of more than 100
757Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
genes. These genes are mainly connected with An Ala to Ser exchange in the C-terminal part
photosynthesis. In maize the two GLK genes of the enzyme is closely related to the lower
are expressed differentially with GOLDEN2 affinity for the substrate PEP (Blasing et al.,
in the mesophyll and bundle sheath, 2000). It is found in all C4 PEPCs analyzed
specifically affecting only chloroplast so far but lacking in nonphotosynthetic or
development in the bundle sheath cells Crassulacean acid metabolism PEPC isoforms
(Waters and Langdale, 2009). Therefore, in (Gowik and Westhoff, 2010). However,
mesophyll/bundle sheath differentiation in the within the grass family C4 PEPCs
C4 plant maize one of the important independently evolved at least eight times,
components of the gene regulatory network then also the resulting enzymes show high
appears as the GLK proteins. degree of similarity . 21 amino acid positions
showed a strong positive selection(Christin et
Enzyme properties optimization al., 2007). Out of these 21 amino acid
positions only two of them are also important
The non-photosynthetic isoforms gave rise to for the evolution of dicot C4 PEPCs. This
all C4 cycle enzymes. The C4 pathway ensures suggests the special requirements for grass C4
high fluxes, as compared to the original PEPCs in comparison to dicot C4 PEPCs.
metabolic environment the concentration of Alternatively, within grasses this can also be
substrates and effector metabolites is elevated inferred that in comparison to the first origins
in the ancestral C3 species. Accordingly, the of C4 photosynthesis most of the dicot C4
C4 isoforms evolution involved changes in lineages are very young (Ehleringer et al.,
their kinetic and regulatory properties. 1997; Sage, 2004). Thus it indicates that the
Perhaps the C4 isoform of PEPC is the best- C4 PEPCs of the grass family are much more
documented example for these evolutionary optimized for their role in C4 photosynthesis
processes (Gowik and Westhoff, 2010). In than their dicot counterparts. Within the
comparison to the nonphotosynthetic PEPCs, photosynthetic PEPCs of the grasses the
C4 PEPCs bind PEP with a lower affinity, higher degree of convergence can be
however their affinity to the other substrate, understood through this.
i.e. bicarbonate, is enhanced. The allosteric
inhibitors of the C4 PEPC isoforms are Asp The unique kinetic and regulatory properties
and malate, towards which they are more were also acquired by the C4 NADP-ME
tolerant and have strongly affected by the during their evolution from nonphotosynthetic
allosteric activators Glc-6-P or Gly. isoforms. The malate and differences in
Relatively small changes in primary enzyme tetramerization of the enzyme leading to an
structure were responsible for these altered pH dependent inhibition in distinct
differences in enzymatic properties. In enzyme regions could be identified (Detarsio
Flaveria trinervia (C4) and Flaveria pringlei et al., 2007). An alteration in the cellular
(C3) the pair of orthologous ppcA PEPCs location of the enzyme also involve in
shares 96% identical amino acid positions. adaptation of C4 enzymes to the new
This was used as an experimental system to metabolic context of the C4 pathway. For
identify some of the evolutionary changes at instance, the photosynthetic carbonic
the amino acid level resolution (Westhoff and anhydrase gene of F. bidentis (FbCA3).
Gowik, 2004). Certain constraints were Highly expressed gene in the mesophyll cells
subjected at the molecular alteration level that (Tetu et al., 2007) evolved from a chloroplast-
is given by the enzyme’s properties. targeted ancestral carbonic anhydrase gene.
The ancestral enzyme mutation in the
758Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
chloroplast transit peptide, the C4 isoform vein density, or enlarged bundle sheath cells.
changed to a cytosolic enzyme (Tanz et al., The transcriptomes, proteomes, and
2009). However, higher expression of this metabolomes analysis of different
ancestral carbonic anhydrase gene was developmental stages of C4 leaves will help in
already reported in leaves, reflecting the understanding the regulation of C4 leaf
minor importance of the intracellular differentiation and the establishment of Kranz
localization of the protein and during anatomy. The transcriptomes level
evolution they were altered. During C4 comparison of closely related C3 and C4
evolution, so far it is not explicitly understood species from genera like Flaveria or Cleome
to the extent the modification of indirect (Brautigam et al., 2011) illuminates the
related enzymes of theC4 pathway. evolutionary trajectories of C4 photosynthesis
and reveals the gene repertoire requirement
Change of C3 crops into C4 photosynthesis for the transition of a C3 into a C4 plant.
Ensuring food security and protecting the In conclusion, the current scenario’s
environment for the world is a continuing complication is the manipulation of the
challenge (Evans, 1998) and requires a second biosphere by human beings. Particularly,
Green Revolution. For covering the energy increase in the atmospheric CO2 could halt the
demands green energy from plant biomass is rise of new C4 life forms and may lead to the
being developed and that might compete with reduction of existing ones (Edwards et al.,
food production for feeding the 21st century 2001). However, other global variables such
growing population for terrain and resources as climatechange, global warming and
in the future. Adequate increase of crop deforestation favorscertain C4 species (Sage
production in a sustainable manner will be and Kubien 2003). Thus, rise in CO2 may
challenging both in terms of harvestable yield threat many C4 species but C4 photosynthesis
and total biomass. Since C4 plants has high as a functional type should not be threatened
photosynthetic capacity and better nitrogen in the near term (Sage et al., 1999b).
and water resources use efficiency. In recent
years C4 photosynthesis has received greater Another avenue for the rise of novel C4
interest and thus it is being considered to species is under way by humanity namely the
transfer C4 photosynthesis into current C3 of C4 photosynthesis into C3 crops (Sheehy et
crops (Sheehy et al., 2007). Currently in rice, al., 2000; Miyao 2003). Research in the
attempts to implement a C4-CO2 concentration natural pathways for C4 evolution may be an
pathwayare under way. important endeavor for overcoming the
developmental barriers to C4 photosynthesis.
The prerequisite for the success of this The identification of the key regulators of C4
endeavor is the knowledge about the genetic traits, and their integration and generation of a
architecture of C4 photosynthesis and the strategy of how the C3 plant rice must be
underlying gene regulatory networks. genetically altered to introduce the C4
Different approaches are needed for pathway should become a milestone in the
elucidating these networks. Large scale relatively young field of synthetic biology.
forward-genetic (with mutagenized rice and
Sorghum bicolor) as well as reverse-genetic References
approaches are being carried out for
identifying the genes involved in C4 subtraits Akyildiz M, Gowik U, Engelmann S, Koczor
like a reduced CO2 compensation point, high M, Streubel M, Westhoff P (2007)
Evolution and function of a cis-regulatory
759Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
module for mesophyll specific gene Cousins AB, Duvall MR, Fox DL,
expression in the C4 dicot Flaveria Freckleton RP, et al., (2010) The origins
trinervia. Plant Cell19:3391-402. of C4 grasslands: integrating evolutionary
Anderson LE (1971) Chloroplast and and ecosystem science. Science328:587-
cytoplasmic enzymes. II. Pea leaf triose 91.
phosphate isomerases. Biochim Biophys Edwards GE, Furbank RT, Hatch MD, Osmond
Acta, 235:237-44. CB. (2001) What does it take to be C4?
Apel P and Peisker M (1978) Einflusshoher Lessons from the evolution of C4
Sauerstoffkonzentrationen auf den CO2- photosynthesis. Plant Physiol., 125:46-
kompensationspunkt von C4-Pflanzen. 49.
Kulturpflanze26:99-103. Ehleringer J. and Bjorkman O. (1977) Quantum
Berry JA & Farquhar GD (1978) The CO2 yields for CO2 uptake in C3 and C4 plants.
concentrating function of C4 Dependence on temperature, CO2 and O2
photosynthesis a biochemical model. In concentration.Plant Physiol59:86-90.
The Proceedings of the Fourth Ehleringer JR, Cerling TE, Helliker BR (1997)
International Congress on Photosynthesis C4 photosynthesis, atmospheric CO2, and
(eds D. Hall, J. Coombs & T. Goodwin), climate.Oecologia112:285-99.
pp 119-31. Biochemical Society of Ehleringer JR, Sage RF, Flanagan LB, Pearcy
London, London, UK. RW (1991) Climate change and the
Blasing OE, Westhoff P, Svensson P (2000) evolution of C4 photosynthesis. Trends
Evolution of C4phosphoenolpyruvate EcolEvol6:95-99.
carboxylase in Flaveria, a conserved Engelmann S, Blasing OE, Gowik U, Svensson
serine residue in the carboxyl-terminal P, Westhoff P (2003) Molecular evolution
part of the enzyme is a major determinant of C4phosphoenolpyruvate carboxylase in
for C4- specific characteristics. J the genus Flaveria—a gradual increase
BiolChem275:27917-23. from C3 to C4 characteristics.
Brautigam A, Kajala K, Wullenweber J, Planta217:717-25.
Sommer M, Gagneul D, Weber KL, Carr Engelmann S, Wiludda C, Burscheidt J, Gowik
KM, Gowik U, Maß J, Lercher MJ, et al., U, Schlue U, Koczor M, Streubel M,
(2011) An mRNA blueprint for C4 Cossu R, Bauwe H, Westhoff P (2008)
photosynthesis derived from comparative The gene for the subunit of glycine
transcriptomics of closely related C3 and decarboxylase from the C4 species
C4 species. Plant Physiol155:142-56. Flaveria trinervia: analysis of
Brown RH and Byrd GT (1993) Estimation of transcriptional control in transgenic
bundle sheath cell conductance in C4 Flaveria bidentis (C4) and Arabidopsis
species and O2 insensitivity of (C3). Plant Physiol., 146:1773-85.
photosynthesis. Plant Physiol103:1183- Evans J.R. & von Caemmerer S. (2000) Would
88. C4 rice produce more biomass than C3
Dengler NG, Nelson T (1999) Leaf structure rice? In Redesigning Rice Photosynthesis
and development in C4 plants. In RF to Increase Yield (eds P.L. Sheehy, P.L.
Sage, RK Monson, eds, C4 Plant Biology. Mitchell and B. Hardy), pp. 293. Elsevier
Academic Press, San Diego, pp 133-72. Science B.V. and International Rice
Detarsio E, Alvarez CE, Saigo M, Andreo CS, Research Institute, Amsterdam, The
Drincovich MF (2007) Identification of Netherlands.
domains involved in tetramerization and Evans J.R. and von Caemmerer S. (1996)
malate inhibition of maize C4-NADP- Carbon dioxide diffusion inside leaves.
malic enzyme. J BiolChem282:6053-60. Plant Physiol., 110:339-46.
Edwards EJ, Osborne CP, Stromberg CA, Farquhar G.D. (1983) On the nature of carbon
Smith SA, Bond WJ, Christin PA, isotope discrimination in C4 species.
760Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
Australian J Plant Physiol10:205-26. Long SP (1999) Environmental responses. In
Freitag H. and Stichler W. (2000) A remarkable RF Sage, RK Monson, eds, C4 Plant
new leaf type with unusual photosynthetic Biology. Academic Press, San Diego, pp
tissue in a central Asiatic genus of 215-49.
Chenopodiaceae. Plant Biol2:154-60. Matsuoka M (1995) The gene for pyruvate,
Friso G, Majeran W, Huang M, Sun Q, van orthophosphate dikinase in C4 plants:
Wijk KJ (2010) Reconstruction of structure, regulation and evolution. Plant
metabolic pathways, protein expression, Cell Physiol 36:937-43.
and homeostasis machineries across Matsuoka M, Tada Y, Fujimura T, Kano-
maize bundle sheath and mesophyll Murakami Y (1993) Tissues pecific light-
chloroplasts: large-scale quantitative regulated expression directed by the
proteomics using the first maize genome promoter of a C4 gene, maize pyruvate,
assembly. Plant Physiol152:1219-50. orthophosphate dikinase, in a C3 plant,
Gowik U and Westhoff P (2011)The Path from rice. ProcNatlAcadSci90:9586-90.
C3 to C4 photosynthesis. Plant Mentzen WI, Wurtele ES (2008) Regulon
Physiol155:56-63. organization of Arabidopsis. BMC Plant
Gowik U, Burscheidt J, Akyildiz M, Schlue U, Biol8:99.
Koczor M, Streubel M, Westhoff P Miyao M. (2003) Molecular evolution and
(2004) cis-Regulatory elements for genetic engineering of C4 photosynthetic
mesophyll-specific gene expression in the enzymes. J Exp Bot 54:179-89.
C4 plant Flaveriatrinervia, the promoter Monson RK (1999) The origins of C4 genes and
of the C4phosphoenolpyruvate evolutionary pattern in the C4 metabolic
carboxylase gene. Plant Cell16:1077-90. phenotype. In RF Sage, RK Monson, eds,
Gowik U, Westhoff P (2010) C4- C4Plant Biol. Academic Press, San Diego,
phosphoenolpyruvate carboxylase. In AS pp 377-410.
Raghavendra, RF Sage, eds, C4 Monson RK (2003) Gene duplication,
Photosynthesis and Related CO2 neofunctionalization, and the evolution of
Concentrating Mechanisms. Springer C4 photosynthesis. Int J Plant Sci (Suppl)
Verlag, Heidelberg-Berlin, pp 257-75. 164:S43-S54.
Hatch MD (1987) C4 photosynthesis: a unique Morgan CL, Turner SR, Rawsthorne S (1993)
blend of modified biochemistry, anatomy Coordination of the cell specific
and ultrastructure. Biochim Biophys Acta, distribution of the four subunits of glycine
895:81-106. decarboxylase and of serine
Hibberd JM, Covshoff S (2010) The regulation hydroxymethyl transferase in leaves of
of gene expression required for C4 C3-C4 intermediate species from different
photosynthesis. Annu Rev Plant genera.Planta190:468-73.
Biol61:181-207. Muhaidat R, Sage RF, Dengler NG (2007)
Hibberd JM, Quick WP (2002) Characteristics Diversity of kranz anatomy and
of C4 photosynthesis in stems and petioles biochemistry in C4eudicots.Am J Bot.,
of C3 flowering plants. Nature415:451- 94:362-81.
54. Oaks A (1994) Efficiency of nitrogen utilization
Jordan DB, Ogren WL (1984) The CO2/O2 in C3 and C4 cereals. Plant Physiol,
specificity of ribulose 1,5- bisphosphate 106:407-14.
carboxylase/oxygenase. Planta161:308- Patel M, Siegel AJ, Berry JO (2006)
13. Untranslated regions of FbRbcS1 mRNA
Ku MSB,Wu J, Dai Z, Scott RA, Chu C, mediate bundle sheath cell-specific gene
Edwards GE (1991) Photosynthetic and expression in leaves of a C4 plant. J
photorespiratory characteristics of BiolChem281:25485-91.
flaveria species. Plant Physiol96:518-28. Rawsthorne S, Hylton CM, Smith AM,
761Int.J.Curr.Microbiol.App.Sci (2020) 9(1): 748-762
Woolhouse HW (1988) Distribution of bisphosphate carboxylases may be nearly
photorespiratory enzymes between perfectly optimized. ProcNatlAcadSci
bundle-sheath and meso- phyll cells in USA 103:7246-51.
leaves of the C3-C4 intermediate species Tetu SG, Tanz SK, Vella N, Burnell JN,
Moricandia arvensis (L.) DC. Ludwig M (2007) The Flaveriabidentis
Planta176:527-32. beta-carbonic anhydrase gene family
Sage RF (2004) The evolution of C4 encodes cytosolic and chloroplastic
photosynthesis. New Phytol161:341-70. isoforms demonstrating distinct organ-
Sage RF, Kubien DS (2003)Quo vadisC4?An specific expression patterns. Plant
ecophysiological perspective on global Physiol.,144:1316-27.
change and the future of C4 plants. Ueno O, Kawano Y, Wakayama M, Takeda T
Photosynth Res 77:209-25. (2006) Leaf vascular systems in C3 and
Sawers RJ, Liu P, Anufrikova K, Hwang JT, C4 grasses: a two-dimensional analysis.
Brutnell TP (2007) A multitreatment Ann Bot (Lond) 97:611-21.
experimental system to examine vonCaemmerer S and Furbank RT (1999)
photosynthetic differentiation in the Modeling of C4 photosynthesis. In The
maize leaf. BMC Genomics, 8:12. Biology of C4 Photosynthesis (ed. R.F.
Sheehy JE, Mitchell PL, Hardy B (2007) Sage), pp. 169-207. Academic Press, San
Charting New Pathways to C4Rice.World Diego, CA, USA.
Scientific, Singapore. Voznesenskaya EV, Franceschi VR, Kiirats O,
Sheehy JE, Mitchell PL, Hardy B, eds. Artyusheva EG, Freitag H & Edwards
(2000)Redesigning rice photosynthesis to GE. (2002) Proof of C4 photosynthesis
increase yields. Amsterdam, the without Kranz anatomy in Bienertia
Netherlands: Elsevier Science. cycloptera (Chenopodiaceae). Plant J
Tanz SK, Tetu SG, Vella NG, Ludwig M 31:649-62.
(2009) Loss of the transit peptide and an Voznesenskaya EV, Franceschi VR, Kiirats O,
increase in gene expression of an Freitag H. and Edwards GE. (2001) Kranz
ancestral chloroplastic carbonic anatomy is not essential for terrestrial C4
anhydrase were instrumental in the plant photosynthesis. Nature 414:543-46.
evolution of the cytosolic C4 carbonic Waters MT, Langdale JA (2009) The making of
anhydrase in Flaveria. Plant Physiol., a chloroplast. EMBO J., 28:2861-73.
150:1515-29. Westhoff P, Gowik U (2004) Evolution of
Tcherkez GG, Farquhar GD, Andrews TJ C4phosphoenolpyruvate carboxylase-
(2006) Despite slow catalysis and genes and proteins: a case study with the
confused substrate specificity, all ribulose genus Flaveria. Ann Bot., 93:1-1.
How to cite this article:
Priyanka Upadhyay, Neha Agrawal, Praveen Kumar Yadav and Ruby Patel. 2020. The
Evolutionary Path from C3 to C4 Photosynthesis: A Review. Int.J.Curr.Microbiol.App.Sci.
9(01): 748-762. doi: https://doi.org/10.20546/ijcmas.2020.901.082
762You can also read

















































