Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes - zhangLab
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Resource
Tracing the genetic footprints of vertebrate landing
in non-teleost ray-finned fishes
Graphical abstract Authors
Xupeng Bi, Kun Wang, Liandong Yang, ...,
Min Zhu, Shunping He, Guojie Zhang
Correspondence
clad@ihb.ac.cn (S.H.),
guojie.zhang@bio.ku.dk (G.Z.),
wwang@mail.kiz.ac.cn (W.W.),
zhumin@ivpp.ac.cn (M.Z.)
In Brief
Comparative analyses of divergent
lineages of ray-finned fishes reveal that
that these species exhibit mosaic
genomic features that have facilitated the
adaptive evolution of phenotypes that
contributed to the water-to-land
transition.
Highlights
d De novo reference genome assemblies of four non-teleost
ray-finned fishes
d Basal ray-finned fishes have key limb development
regulatory elements
d Lung-related genes in early ray-finned fishes hold the key for
the lung origin
d Cardio-respiratory systems’ co-evolution during early air
breathing evolution
Bi et al., 2021, Cell 184, 1–15
March 4, 2021 ª 2021 Elsevier Inc.
https://doi.org/10.1016/j.cell.2021.01.046 ll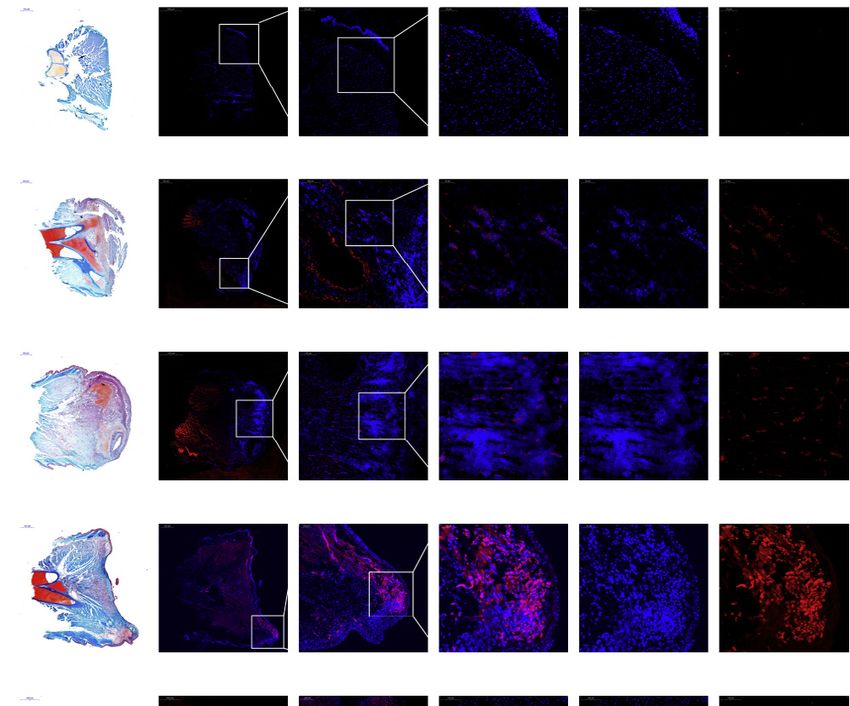
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
Tracing the genetic footprints of vertebrate
landing in non-teleost ray-finned fishes
Xupeng Bi,1,2,17 Kun Wang,3,17 Liandong Yang,1,17 Hailin Pan,2,17 Haifeng Jiang,1,17 Qiwei Wei,4,17 Miaoquan Fang,2
Hao Yu,2 Chenglong Zhu,3 Yiran Cai,2 Yuming He,2 Xiaoni Gan,1 Honghui Zeng,1 Daqi Yu,5,6 Youan Zhu,7,8 Huifeng Jiang,9
Qiang Qiu,3 Huanming Yang,2,10,11 Yong E. Zhang,5,6,12 Wen Wang,3,12,13,* Min Zhu,7,8,14,* Shunping He,1,12,15,*
and Guojie Zhang2,12,13,16,18,*
1State Key Laboratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology, Chinese Academy of Sciences,
Wuhan 430072, China
2BGI-Shenzhen, Shenzhen 518083, China
3School of Ecology and Environment, Northwestern Polytechnical University, Xi’an 710072, China
4Key Laboratory of Freshwater Biodiversity Conservation, Ministry of Agriculture and Rural Affairs, Yangtze River Fisheries Research Institute,
Chinese Academy of Fishery Sciences, Wuhan, China
5Key Laboratory of Zoological Systematics and Evolution and State Key Laboratory of Integrated Management of Pest Insects and Rodents,
Institute of Zoology, Chinese Academy of Sciences, Beijing 100101, China
6University of Chinese Academy of Sciences, Beijing 100049, China
7Key Laboratory of Vertebrate Evolution and Human Origins, Institute of Vertebrate Paleontology and Paleoanthropology, Chinese Academy
of Sciences, 142 Xi-zhi-men-wai Street, Beijing 100044, China
8CAS Center for Excellence in Life and Paleoenvironment, Beijing 100044, China
9Tianjin Institute of Industrial Biotechnology, Chinese Academy of Sciences, Tianjin, China
10James D. Watson Institute of Genome Sciences, Hangzhou, China
11Guangdong Provincial Academician Workstation of BGI Synthetic Genomics, BGI-Shenzhen, Shenzhen 518120, China
12Center for Excellence in Animal Evolution and Genetics, Chinese Academy of Sciences, 32 Jiaochang Donglu, Kunming 650223, China
13State Key Laboratory of Genetic Resources and Evolution, Kunming Institute of Zoology, Chinese Academy of Sciences,
Kunming 650223, China
14College of Earth and Planetary Sciences, University of Chinese Academy of Sciences, Beijing 100049, China
15Institute of Deep-Sea Science and Engineering, Chinese Academy of Sciences, Sanya 572000, China
16Villum Center for Biodiversity Genomics, Section for Ecology and Evolution, Department of Biology, University of Copenhagen,
Copenhagen, Denmark
17These authors contributed equally
18Lead contact
*Correspondence: wwang@mail.kiz.ac.cn (W.W.), zhumin@ivpp.ac.cn (M.Z.), clad@ihb.ac.cn (S.H.), guojie.zhang@bio.ku.dk (G.Z.)
https://doi.org/10.1016/j.cell.2021.01.046
SUMMARY
Rich fossil evidence suggests that many traits and functions related to terrestrial evolution were present long
before the ancestor of lobe- and ray-finned fishes. Here, we present genome sequences of the bichir, pad-
dlefish, bowfin, and alligator gar, covering all major early divergent lineages of ray-finned fishes. Our analyses
show that these species exhibit many mosaic genomic features of lobe- and ray-finned fishes. In particular,
many regulatory elements for limb development are present in these fishes, supporting the hypothesis that
the relevant ancestral regulation networks emerged before the origin of tetrapods. Transcriptome analyses
confirm the homology between the lung and swim bladder and reveal the presence of functional lung-related
genes in early ray-finned fishes. Furthermore, we functionally validate the essential role of a jawed vertebrate
highly conserved element for cardiovascular development. Our results imply the ancestors of jawed verte-
brates already had the potential gene networks for cardio-respiratory systems supporting air breathing.
INTRODUCTION that occurred during the origin of tetrapods is challenging
because the animals involved in the transitional events are
The water-to-land transition during the Devonian is one of the extinct. However, it has long been postulated that many of these
most prominent events in vertebrate evolution. During this tran- innovations evolved in the ancestors of bony vertebrates (ray-
sition, various organs underwent adaptive changes, particularly finned fishes, lobe-finned fishes, and tetrapods) and thus still
those related to locomotion and respiration systems (Hinchliffe, are present in some living basal groups of actinopterygians
1994; Long and Gordon, 2004). Study of the genomic changes (ray-finned fishes) (Graham et al., 2014; Takeuchi et al., 2009;
Cell 184, 1–15, March 4, 2021 ª 2021 Elsevier Inc. 1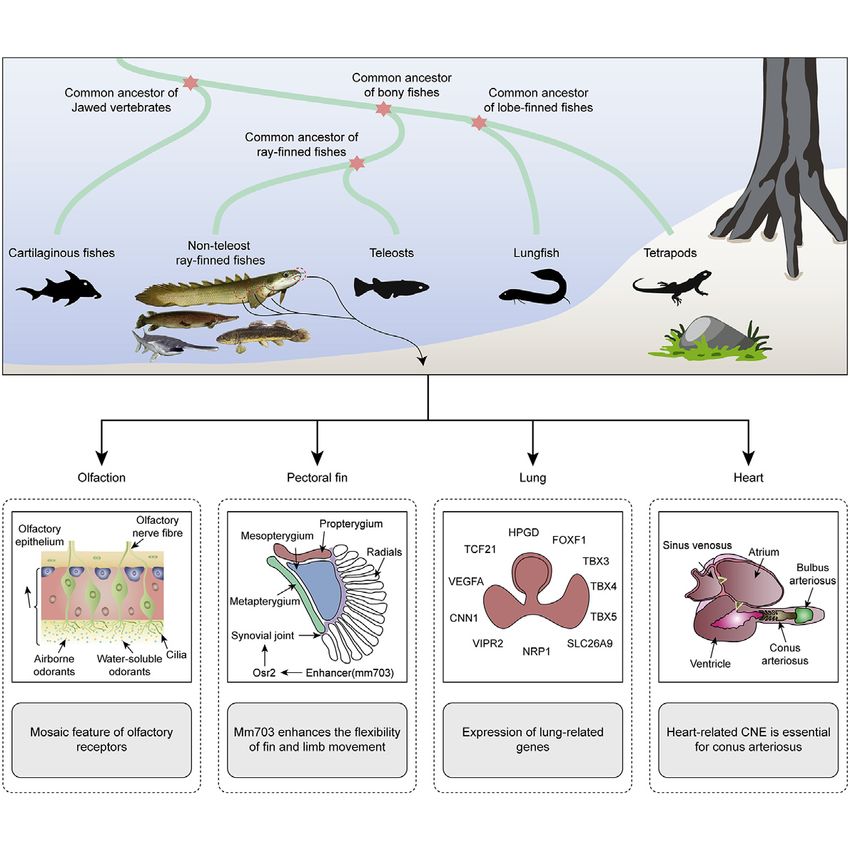
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
Zaccone et al., 2009). For instance, the lung respiratory system features of lobe- and ray-finned fishes, particularly relating to
that tetrapods rely on for survival also exists in several basal regulatory elements for limb development. We then undertook
lobe-finned fishes, such as coelacanths, lungfish, and Polypter- transcriptomic-based exploration to confirm the homology be-
iformes, one of the early divergent ray-finned fish groups. tween the lung and swim bladder and revealed the presence of
Anatomically, the lungs are distributed in pairs on the ventral functionally lung-related genes in early ray-finned fishes. Finally,
side and originate from the pharynx posterior to the gills. While we performed functional experiments to validate the essential
the lungs disappeared and were replaced by the swim bladder role of an element that is highly conserved in jawed vertebrate
in most ray-finned fishes, the presence of the lung or its homol- (gnathostome) ancestors in cardiovascular development.
ogous organs in the stem jawed vertebrates has been proposed
for decades (Denison, 1941; Goujet, 2011; Perry et al., 2001). It is RESULTS
clear that the lungs should have originated by at least the last
common ancestor (LCA) of ray-finned fishes and lobe-finned Genome assembly and annotation
fishes, which is at least in Late Silurian, not very far away from We generated from a bichir 250 Gb of long sequence reads using
the LCA of cartilaginous and bony fishes (presumably Upper Oxford Nanopore Technology and used the data to produce a
Ordovician) (Andreev et al., 2015). Thus, the genetic basis of draft genome assembly. The assembly was polished using Illu-
the traits that evolved during this major transition may be acces- mina short reads and then further improved into a chromo-
sible through comparison of existing developmental pathways some-level assembly with Hi-C data. The final assembly of the
that could mirror the developmental plasticity in the ancestral bichir genome has contig N50 size of 4.53 Mb and scaffold N50
lineage (Amemiya et al., 2013; Gibert, 2017; Muschick et al., size of 189.69 Mb, with 94.3% of the sequences anchored onto
2011; West-Eberhard, 2003). If so, investigation on the genomes 18 chromosomes. The genomes of the American paddlefish,
of extant species from these early divergent lineages of ray- bowfin, and alligator gar were sequenced and assembled at
finned fishes may illuminate the genetic regulatory processes 135–242x coverage with Illumina short reads from libraries with
underlying the trait evolution during the terrestrial transition (Qu inserted sizes ranging from 250 bp to 40 kb (Table S1). From
et al., 2015). 89%–96% of the 2,586 vertebrate core sets of Benchmarking
The ray-finned fishes (Actinopterygii) are an extremely diverse Universal Single-Copy Orthologs (BUSCO) are completely anno-
group of vertebrates with over 32,000 species, thus representing tated in these four species (Simão et al., 2015) (Table S1). We also
about half of all extant vertebrate species (Friedman, 2015; Sal- produced transcriptome data from nine tissues for each species
lan, 2014). Most Actinopterygii (ca. 95%) are teleosts, which to assist the annotation, which resulted in 18,839–23,374 protein
initially diversified through adaptive radiation in the Mesozoic coding genes for these genomes (Table S1).
and underwent further dramatic diversification in the Cenozoic The sizes of the non-teleost actinopterygians genomes studied
(Glasauer and Neuhauss, 2014). The other extant ray-finned to date vary considerably, from 0.95 to 3.67 Gb, which implies
fishes are distributed in three deeply divergent actinopterygian dramatic changes happened to their genomic structures during
groups (Cladistia, Chondrostei, and Holostei), which are often re- the early divergence of ray-finned fishes. Some of this size varia-
garded as ‘‘living fossils’’ due to their ancient evolutionary origins tion is due to differences in their proportions of repeat content,
and possession of traits ascribed to a common ancestor of ray- which range from 14% in the gar to 44% in the bichir (Table
finned fishes and lobe-finned fishes (Hurley et al., 2007; Noack S1). Interestingly, TcMar-Tc1 seems to be the most abundant
et al., 1996). Many species in these groups possess fascinating type of transposon in all basal ray-finned fishes (Figure S1A), in
characteristics that have been associated with air-breathing dur- accordance with previous findings that TcMar-Tc1 transposons
ing vertebrate terrestrialization, such as ventral paired lungs are active in freshwater species (Yuan et al., 2018). In addition,
(Farmer, 1999; Perry and Sander, 2004). Living cladistians (Poly- similar to the genomes of lobe-finned and cartilaginous fishes,
pterus and Erpetoichthys) also bear morphological similarities to these genomes contain abundant LINE/CR1 repeats, which
the lobe-finned fishes, such as elongated body forms, large might have originated from their common ancestor (Figure S1B).
paired spiracles dorsally positioned on the head, and developed
lobate portion of the pectoral fins. These similarities either Basic interrelationships of ray-finned fishes
evolved through convergent evolution (Friedman, 2015; Giles Despite extensive analyses on both extinct and extant species
et al., 2017) or were inherited from the primitive forms of gna- since the late nineteenth century (Allen, 1876; Cope, 1878), the
thostomes (Zhu et al., 2013; Zhu and Yu, 2009). These mosaic early evolutionary history of actinopterygians remains controver-
features place the non-teleost actinopterygians in a phylogenetic sial. The major debate regarding extant lineages concerns the
position that is crucial for comparative evolutionary and develop- relationship between Chondrostei and Holostei. While some
mental studies that aim to illuminate some of the key transitions studies have argued they are a monophyletic clade (Inoue
that occurred during the evolution of terrestrial vertebrates. et al., 2003; Venkatesh et al., 2001), others have indicated that
Thus, to elucidate the genetic basis of these features in early Acipenseriformes (sturgeons and paddlefishes), an order of the
diverging ray-finned fishes, we produced a chromosome level Chondrostei subclass, should be treated as a sister group of
assembly of the bichir (Polypterus senegalus) and draft genome the Neopterygii that is composed of Holostei and Teleostei (Kiku-
assemblies for the American paddlefish (Polyodon spathula), the gawa et al., 2004). We identified 359 one-to-one orthologs
bowfin (Amia calva), and the alligator gar (Atractosteus spatula), across 27 chordates (allowing up to one missing) that derived
which together cover all three teleost outgroup lineages. Our an- from the same ancestral chromosomes before the two rounds
alyses showed that these species exhibit many mosaic genomic of whole-genome duplications (WGDs) that occurred at the
2 Cell 184, 1–15, March 4, 2021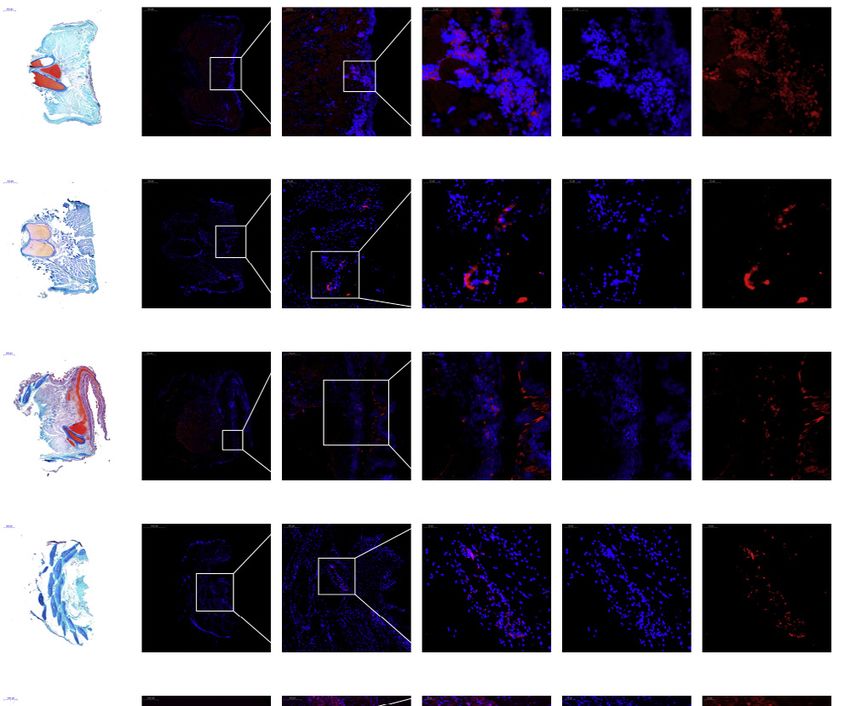
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
(legend on next page)
Cell 184, 1–15, March 4, 2021 3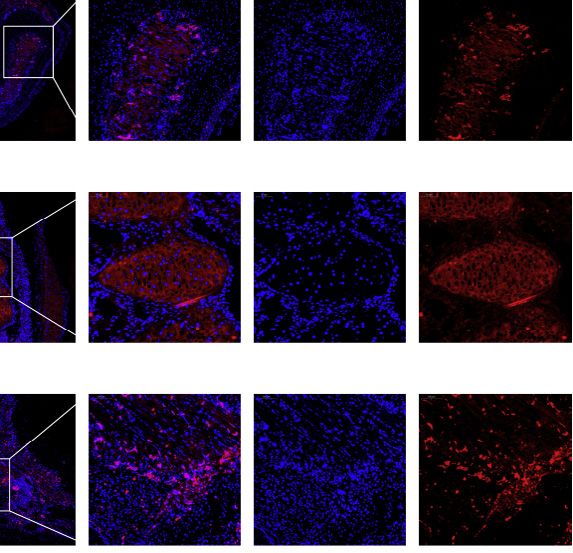
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
base of vertebrates (Sacerdot et al., 2018) (Figure S1C). Phyloge- 3R WGD also occurred in paddlefish (Figure S2C). To investigate
netic analyses with these orthologous genes confirmed that the whether the 3R WGD event occurred in the common ancestor of
Polypteriformes (bichirs) form a sister lineage of all other ray- paddlefish and sterlet, or independently in the two lineages, we
finned fishes and resolved that Acipenseriformes are the sister analyzed the distributions of synonymous substitution rates
lineage of the Neopterygii (Figure 1). This topology is also (Ks) and 4-fold degenerative third-codon transversions (4dTv)
supported by morphological evidence (Gardiner et al., 2005) among the 3,743 paddlefish and 8,297 sterlet paralogs. We
and further confirmed by the phylogenetic trees generated with found that the Ks age distributions of the two species had
23 Mb of genome-wide orthologous sequences and 319 single different ranges, suggesting that independent WGD events
copy genes of 14 selected vertebrate species (Figures S1D occurred in the two lineages (Figure 2C). This pattern was also
and S1E). Molecular dating using the 359 orthologs and cali- confirmed by both the distribution of 4dTv values, and gene phy-
brated using nine fossil records (Table S1) suggests that actino- logeny analyses (Figures S2D and S2E). Based on the diver-
pterygians split from sarcopterygians about 453 million years gence time (135 Mya) between paddlefish and sterlet (Figure 1),
ago (Mya, 95% confidence interval: 445.8–459.8 Mya), which is we estimate that the 3R WGD events in paddlefish and sterlet
close to a previous estimate (422 Mya) based on mitochondrial occurred at 121 Mya and 51 Mya, respectively. Although our
genomes (Lalu et al., 2010). estimated date for the 3R WGD in sterlet is much more recent
than that estimated in a previous study, that prior estimate based
Genome evolution of early ray-finned fishes on a sterlet-Atlantic sturgeon divergence time of 166 Mya, which
The split of actinopterygians and sarcopterygians was a major is much earlier than the divergence time (74 Mya) estimated in
evolutionary step during the transition toward current vertebrate other studies (Rabosky et al., 2013).
diversity (Evans, 2008; Soltis and Soltis, 2012; Volff, 2005). Using The independent duplication of the whole genome in these
the chromosome assemblies of bichir, sterlet, and spotted gar two closely related lineages offers an excellent model for
(Braasch et al., 2016; Du et al., 2020), we reconstructed the studying the evolutionary fate of duplicate genes after WGD.
ancestral karyotype of ray-finned fishes and obtained 38 proto- Unlike the sterlet, which retained eight Hox clusters, the pad-
chromosomes for Actinopterygii (Figure 2A; see Figure S2A for dlefish genome has lost one HoxC cluster since its 3R WGD.
a full description of karyotype reconstruction). Remarkably, Interestingly, convergent loss of this HoxC cluster after 3R
the non-teleost actinopterygians have a higher level of genome WGD also occurred in some teleost fishes, such as fugu (Ta-
synteny with chicken than teleosts, whose genomes have under- kifugu rubripes) and medaka (Oryzias latipes) (Figure S2C). At
gone more fusions, fissions, and rearrangements following the global genome level, based on 8,377 singletons and 3,743
teleost-specific genome duplication (Figures 2B and S2B). For paralogous pairs detected in the paddlefish genome, we esti-
example, we found that 51% of the bichir genome has conserved mated a duplicate gene retention rate after the paddlefish 3R
synteny with the chicken genome. When expanding the analyses WGD of around 31%, which is considerably lower than the
to include more jawed vertebrates, we found that non-teleost ac- retention rate in sterlet (70%), in accordance with the indica-
tinopterygians share 9%–36% synteny with humans, thus higher tions that the 3R WGD was more recent in the sterlet than in
than both the proportions shared by any of the teleosts (6%–7%) the paddlefish. However, the rate in the paddlefish is higher
examined and humans (Table S2). These findings clearly indicate than the corresponding rate (15%–20%) following the teleost
that the early actinopterygians largely preserved the ancestral WGD (Hrbek et al., 2007). Notably, duplicated genes involved
genome structure of bony vertebrates before the teleost-specific in protein turnover, signal transduction, cell proliferation, and
genome duplication. Despite the overall conservation with the development were preferably maintained in the sterlet (Du
ancestral genome, 22 chromosome fusion events appear to et al., 2020), but duplicated genes in the paddlefish are en-
have occurred in the bichir lineage, in which ancestral micro- riched in functions related to biosynthetic process, gene
chromosomes merged with other macro-chromosomes. These expression, and metabolic process (Table S2). These findings
rearrangement processes were accompanied with an elevated suggest that substantially different selection pressures acted
level of transposon element activity, which increases genome on the two lineages.
size in these species.
Recent analyses have shown that a third round of WGD (3R Genetic regulation related to limb flexibility
WGD) occurred in the sterlet (Acipenser ruthenus) (Du et al., The evolution of terrestrial locomotion, one of the major transi-
2020), a species closely related to the paddlefish. We detected tions in vertebrate evolution, involved the appearance of derived
seven Hox clusters in the paddlefish genome, indicating that a supporting appendicular structures (Carroll and Holmes, 2007;
Figure 1. Interrelationship and divergence times of a broad selection of living and extinct chordates
Amphioxus was used as an outgroup, and a species tree was generated using RAxML and ASTRAL on the basis of 359 orthologs. The divergence times of extant
species were estimated with the MCMCTree program under a correlated molecular clock and general time reversible (GTR) substitution model. Extinct species
representing taxa in each of the major clades were added to the phylogenetic tree using information in previous literature (Coates et al., 2017; Morris, 2008; Dupret
et al., 2014; Giles et al., 2015; 2017; Grande et al., 2002; Maisey, 2005; Olsen and McCune, 1991; Shu et al., 1999; 2003; Stensiö, 1927; Xu et al., 2018; Young,
1980; Zhu et al., 2013; 2009). The black rectangles at the nodes represent 95% confidence intervals of the corresponding estimated divergence times. The ‘‘y’’
symbols indicate branches leading to extinct species with fossil records (dated to times indicated by the ends of the respective branches). First appearances of
the jaw, lung, and swim bladder are marked on the tree, and geological periods are indicated at the bottom (from left to right: Cryogenian, Ediacaran, Cambrian,
Ordovician, Silurian, Devonian, Carboniferous, Permian, Triassic, Jurassic, Cretaceous, Paleogene, Neogene, and Quaternary).
4 Cell 184, 1–15, March 4, 2021
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
A
B C
Figure 2. Reconstruction of proto-chromosomes for the common ancestor of ray-finned fishes
(A) 38 Actinopterygii proto-chromosomes (color-coded) were constructed using chicken as an outgroup.
(B) Circos plots showing conserved synteny between bichir and sterlet (left) and bichir and spotted gar (right).
(C) Ks distributions of 3,743 paralogous genes in paddlefish (purple), 8,297 paralogous genes in sterlet (orange), and 10,729 orthologs between paddlefish and
sterlet (green). Times of putative 3R WGD events are indicated by the x axis.
Long and Gordon, 2004; Neyt et al., 2000). Some of these To test this hypothesis, we first searched for the presences
anatomical features are also present in the basal ray-finned of homologs for 264 genes related to limb morphogenesis
fishes (Clack, 2009). For example, the humerus is a proximal development, limb bud formation, and limb joint morphogen-
basal element of the sarcopterygian forelimb, which evolved esis in all jawed vertebrates. However, we did not detect
from the metapterygium and putatively appeared in the common much difference in the copy numbers of these genes when
ancestor of cartilaginous and bony fishes but was completely comparing teleosts with other vertebrates, nor did we detect
lost in the teleost pectoral fin (Davis et al., 2004; Tanaka, 2016; specific gene loss in teleosts, suggesting the loss of movement
Wagner and Chiu, 2001; Woltering et al., 2020). A previous study flexibility in teleost might not be through the significant change
on the fleshy pectoral fins of coelacanths showed that the flex- over the protein-coding sequences (Table S3). Therefore, we
ible and movable elbow joint connects the humerus and ulna next screened for the presence of conserved enhancer se-
with two crisscrossed ligaments (Miyake et al., 2016). Mean- quences that have been functionally validated in mice. Among
while, Polypterus possess preaxial and postaxial muscles in 14 tissues investigated, teleosts showed the lowest numbers of
both pectoral and pelvic fins that resemble those in the lobe- conserved enhancers with mice in all tissue types, which might
finned fishes Latimeria and Neoceratodus (Molnar et al., 2017). partially be due to the higher genomic turnover rates after the
We reason that an ancestral developmental potential for the flex- 3R WGD in this group. Interestingly, we found there was a
ible movement of pectoral fins through the joint connections much higher number of conserved enhancers that specifically
might have been present in the crown jawed vertebrates but function in limbs, in non-teleost actinopterygians, lobe-finned
was lost in teleosts. fishes, and cartilaginous fishes, than in teleosts. Specifically,
Cell 184, 1–15, March 4, 2021 5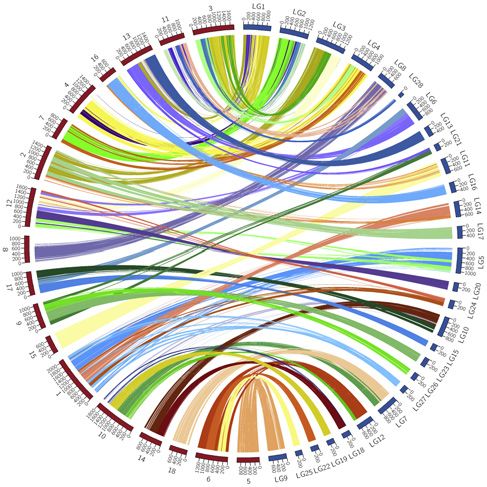
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
A
B
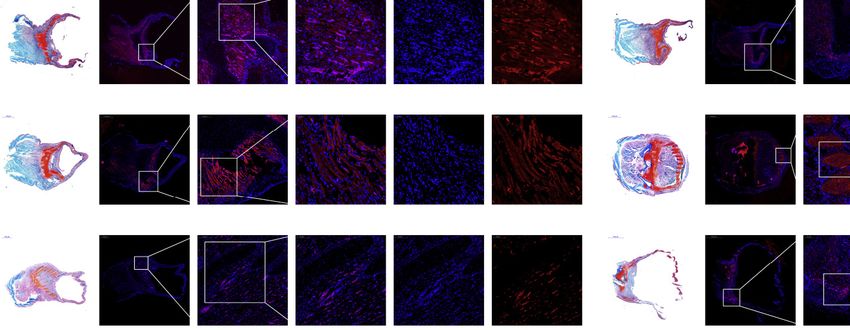
Figure 3. Early limb development and in situ expression of Osr2 in the pectoral fin of bichir
(A) Annotation results for the 40 limb-related enhancers in 25 chordate species. The mm703 enhancer originated after emergence of crown gnathostomes and is
preserved in lobe-finned fishes, tetrapods, and basal ray-finned fishes (as indicated by yellow bars).
(B) Pectoral fin and endoskeleton regeneration in bichir. Panels to the left show Safranin O/Fast green staining at indicated days after amputation (see also
Figure S3B), with cartilage and bone stained red and blue, respectively. The left side in these images is near the base of the pectoral fin. Middle and right panels
show results of in situ hybridization of the Osr2 gene at different magnifications, revealing that Osr2 is weakly expressed in blastema at 1 dpa (days post
amputation) and highly expressed from 30 dpa. The blue dot is DAPI staining, showing the morphology and location of the nucleus. Pink around the blue dot
indicates that the gene is expressed here. Abbreviations: ms, mesopterygium; mt, metapterygium; pr, propterygium; rd, radials (rd). Black arrows in the 70 dpa
image indicate synovial joints in the pectoral fin.
across the six teleost genomes we investigated, only one Strikingly, we detected a highly conserved limb enhancer
species (zebrafish) contains one limb-specific enhancer. In (mm703) in all five cartilaginous fishes and all bony vertebrates
contrast, we identified 3–6 in non-teleost actinopterygian spe- except teleosts. This enhancer is located in the downstream re-
cies and 3–40 in sarcopterygians (Figure 3A). Notably, all of gion of Osr2. A previous study in mice has revealed that mm703
the seven limb enhancers detected in all non-teleost actino- is a cis-regulatory module of the Lmx1b transcription factor that
pterygian species are also present in the coelacanth genome could regulate the expression of the Osr2 gene (Haro et al.,
(Table S3). 2017). Study of the Osr2 transcription factor in mice has shown
6 Cell 184, 1–15, March 4, 2021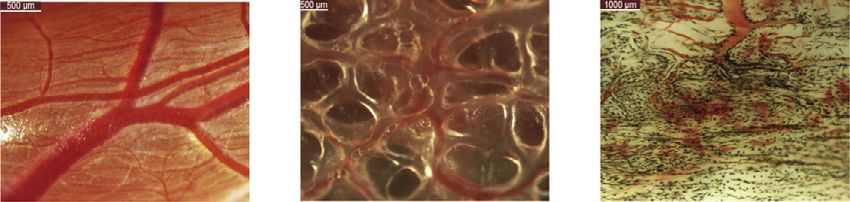
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
Figure 4. Properties of olfactory receptors
in jawed vertebrates
‘‘Air’’ (yellow circles) and ‘‘water’’ (blue circles)
refer to the detection of airborne and water-solu-
ble odorants, respectively. Sizes of the circles
indicate numbers of intact OR genes, which
possess a full-length OR protein coding sequence
that is at least 250 amino acids long.
the regeneration process (Figures 3B and
S3B). Though further functional experi-
ments would be valuable to reveal the
detail mechanism of how mm703 and
Osr2 mediate joint modification, our find-
ings suggest an ancient role of Osr2 in
synovial joints, which might have evolved
in early vertebrates.
Early origination of air olfactory
receptors
Vertebrate genomes contain numerous
olfactory receptor (OR) genes, predomi-
nantly expressed in the olfactory epithe-
lium of the nasal cavity where they play
key roles in detection of odorous mole-
cules in the environment (Zhang and Fire-
stein, 2002). We characterized OR genes
and found cartilaginous fishes have much
smaller (3) numbers of intact OR genes,
while the numbers have been expanded
to 100 and 200 in ray-finned fishes
and lobe-finned fishes, respectively.
Group a and g OR genes are required to
detect airborne odorants and are the
dominant OR type in tetrapods but ab-
sent in teleost fishes, which instead
have a large number of OR genes that
participate in detection of water-soluble
odorants (Niimura, 2009). Interestingly,
most non-teleost actinopterygians have
that this gene is essential for formation of synovial joints (Gao mosaic OR features with a mixture of both types of OR genes,
et al., 2011), which greatly enhances the flexibility of limb move- in accordance with the air-breathing characteristics of some
ment and thus has a presumed association with vertebrate ter- species in these lineages (Figure 4; Table S4). Among them,
restrialization (Askary et al., 2016). Our Hi-C data also revealed the bichir has the highest number of both a and g ORs, which
a close interaction between this enhancer and Osr2 in bichir, is consistent with previous finding that spiracle-mediated aspira-
and the pattern was conserved with that in mice, implying a tion accounts for up to 93% of all air breaths in Polypterus (Gra-
conserved regulatory function might also be present in the bichir ham et al., 2014). It should be noted that both a and g ORs had
(Figure S3A). Consistent with this, it has been shown that the bi- already appeared in bony vertebrates and expanded separately
chir can regenerate pectoral fins with remarkable accuracy, in different lineages in a later period (Figure S4A). Nevertheless,
resembling the limb regeneration process found in amphibians there is no evidence so far that Acipenseriformes can breathe in
(Cuervo et al., 2012). To further infer the potential role of Osr2 the air. Accordingly, there are fewer OR genes in this group than
in the bichir pectoral fin, we monitored the expression profile of in other non-teleost actinopterygians. Notably, previous studies
Osr2 during regeneration of the bichir pectoral fin (as detailed have detected no OR gene related to air-breathing in lampreys or
in the STAR methods section), a homologous structure to the elephant sharks (Niimura, 2009), but we found a g gene in the
front limb in tetrapods. We found that Osr2 was mainly ex- bamboo shark (Chiloscyllium punctatum) and whale shark
pressed at joints in the pectoral fin, especially between the meta- (Rhincodon typus) (Figure 4). The presence of a and g in some
pterygium and radials, and its expression level increased during of the fishes that cannot breathe air remains mysterious. We
Cell 184, 1–15, March 4, 2021 7
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
A B
D
C
E
F
(legend on next page)
8 Cell 184, 1–15, March 4, 2021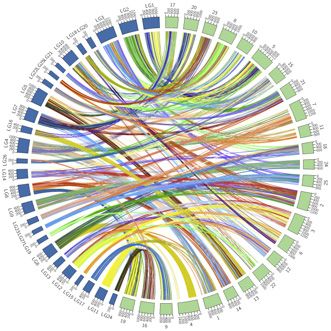
Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
speculate that these ORs might primitively have been function- We also assessed the phylogenetic relationships between
ally associated with water-soluble odorants but later specialized lung and swim bladder samples based on transcriptome dis-
toward interactions with airborne odors in cooperation with aerial tances that were evaluated as 1 Spearman’s correlation coeffi-
olfaction during the fish-tetrapod transition. However, further cient. We found the swim bladders were nested within the lung
functional experiments are needed to verify this hypothesis. samples, which supports the hypothesis that the swim bladder
In addition to the main olfactory system, many vertebrates also evolved from the lung, with a locational transition from the ventral
use the vomeronasal system as an accessory olfactory system to the dorsal side of the esophagus (Sagai et al., 2017) after
for pheromone detection. Most terrestrial vertebrates have a divergence of Polypteriformes and other ray-finned lineages
higher copy number of vomeronasal type-1 receptors (V1Rs) (Figures 5B and S5B). Moreover, the swim bladder of the alligator
than type-2 receptors (V2Rs), which respectively detect small gar clustered with the bichir lung rather than the swim bladder of
volatile molecules diffusing in air and molecules that are soluble paddlefish, indicating that the alligator gar’s swim bladder
in aquatic environments (Silva and Antunes, 2017). Although retains the function of the ancestral lungs that evolved in the
basal ray-finned fishes have more V2Rs than V1Rs like teleosts, common ancestor of ray-finned fishes and lobe-finned fishes
we found that the bichir has significantly higher copy numbers of (Figure 5B). This is consistent with the anatomical structures of
V1Rs than other teleost fishes (Figure S4B). The exact roles of the species’ lungs and swim bladders, as surfaces of the bichir’s
V1Rs in this aquatic fish thus would be interesting for further lung and the alligator gar’s swim bladder are highly vascularized,
experimental study. unlike the surface of the paddlefish swim bladder (Figures 5C
and S5C).
Transcriptional profiles reveal the evolution of A Kolmogorov-Smirnov test showed that expression levels of
vertebrate lungs 461 genes (after quantile normalization) were significantly higher
To address the evolution of lungs and their relationship with the (p < 0.01 after correction) in the swim bladders than in all lung
swim bladders of fish, a question that has intrigued researchers samples. We found that some of the most strongly expressed
for centuries, we sequenced transcriptomes from 355 samples genes in swim bladder (e.g., Ruvbl1, Amot, and Mmp2) are asso-
representing 10 tissues of eight vertebrate species (three ciated with cell migration (Figure 5B), which might have been
lobe-finned fishes, three basal ray-finned fishes, and two tele- involved in the ventral to dorsal transition during swim bladder
osts) with five biological replications per tissue per species development. We also identified 466 genes that are more
(Table S5). An expression level matrix based on 5,046 1:1 or- strongly expressed in air-breathing than in non-air-breathing ver-
thologs of the eight species was generated and standardized tebrates. These genes are significantly enriched in functions
with quantile normalization. The principal-component analysis related to development of alveoli and alveolar lamellar bodies
(PCA) clustered the lung and swim bladder expression profiles, (Table S5). Among them we found 24 genes related to angiogen-
something also supported by the neighbor-joining (NJ) tree of esis, which is crucial for formation of blood vessels at the surface
all tissues based on gene expression levels after normalization of alveoli (Carmeliet and Jain, 2011), and 15 of these were among
(Figure 5A). This result is in accordance with Darwin’s hypoth- the top 50 most significantly differentially expressed genes (Fig-
esis that they are homologous organs (Darwin, 1859) and the ures 5B and 5D). These findings support the above-described
previous anatomic analysis of pulmonary arteries in non-teleost differences in anatomical structure between lungs and swim
actinopterygians that also showed lungs and swim bladders bladders.
are homologous organs(Longo et al., 2013). Additionally, the to- We further compared the lung expression profiles with those of
pology of the tree generated with transcriptome data all other tissues in the bichir and the three lobe-finned species
strongly resembles the species phylogeny, demonstrating the and identified 11 genes exhibiting lung-specific expression (Fig-
high correlation between genomic and transcriptomic evolution ure 5E). Interestingly, all of them reportedly participate in lung
(Figure S5A). development and function (Anagnostopoulou et al., 2012; Arora
Figure 5. Analyses of basal ray-finned species illuminating the evolution of lung-related gene expression
(A) Score plot obtained from PCA of 355 transcriptome samples based on expression profiling of 5,046 orthologs. The NJ tree in the top right corner is based on
expression data and indicates a homologous relationship between lung and swim bladder. The red dotted circle represents the clustering of lungs and swim
bladders.
(B) NJ tree of lung and swim bladder samples based on transcriptome distances, clearly separating actinopterygians and sarcopterygians. Schematic
morphological diagrams of lungs and swim bladders of each species are displayed under the tree. The heatmap below shows expression profiles of 21 of the
most significantly highly expressed genes in the air-breathing and swim bladder groups.
(C) Anatomic illustration of the lungs of the bichir and swim bladders of the American paddlefish and alligator gar.
(D) The angiogenesis process involves three consecutive steps. First an endothelial cell tip cell is selected to ensure blood vessel branching formation. Second, tip
cells navigate in response to guidance signals and adhere to the extracellular matrix to migrate, while stalk cells proliferate and elongate. Finally, neighboring
branches fuse, and a new vessel forms. Eleven highly expressed genes in the air-breathing group participate during these three processes. Genes in the 50 most
highly expressed sets are marked in blue.
(E) The presence and absence of lung-related genes in representative vertebrates.
(F) Results of in situ hybridization analysis of five representative genes in lungs of the bichir and the mouse and swim bladder of the zebrafish, showing that these
genes are expressed in the region of the alveolar cells (indicated by the arrowhead). Zebrafish have two copies of TBX3, NRP1, and VEGFA, one copy of a VIPR2
gene, but completely lost SLC26A9. Zebrafish-a and -b show the expression pattern of the paralogous copies when the paralogous genes exist. The red signals
show the expression of the targeted gene.
Cell 184, 1–15, March 4, 2021 9Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
A C
D
B
E
Figure 6. Genomic sequences specifically shared by non-teleost actinopterygians and lobe-finned fishes
(A) Distributions of conserved synteny blocks in the bichir genome shared with sarcopterygians or teleosts. The gene alignment map in the middle shows the
syntenic map for the Hand2 gene.
(B) VISTA plot showing the presence of a heart-related CNE upstream of the Hand2 gene across all jawed vertebrates except the Neoteleostei. Peaks (blue,
exons; red, non-coding regions) indicate regions with conserved sequences to the human counterpart. The Hand2 CNE is highlighted in pale yellow.
(C) Hematoxylin and eosin-stained transverse sections of E16.5 hearts. Hand2 CNE/ mutant hearts showed slightly reduced right ventricular compact
myocardium of right ventricle body wall (4 out of 11) and significantly decreased chamber volume (6 out of 11). Numbers in the bottom indicate the hearts with
(legend continued on next page)
10 Cell 184, 1–15, March 4, 2021Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
et al., 2012; Hong et al., 2007; Morrisey and Hogan, 2010; Quag- fishes, non-teleost actinopterygians, basal teleosts, and am-
gin et al., 1999; Vaidya et al., 2017). Most of the 11 genes are also phibians; and has been presumably absorbed into the right
expressed in swim bladders of teleosts, suggesting that they still ventricle (RV) in amniotic animals and completely lost in the
play functional roles in both swim bladders and lungs. One of more advanced teleosts (Icardo, 2006; Maldanis et al., 2016).
these genes (Tcf21) has been apparently present since the com- This tissue contains compact myocardium and functions as an
mon ancestor of chordates, and two (Foxf1 and Hpgd) appar- extra ventricle or accessory pumping chamber that minimizes
ently originated with the common ancestor of vertebrates. The pressure fluctuations and regulates the outflow of blood from
others appeared after the split of jawed and jawless vertebrates the ventricle to the ventral aorta (Icardo et al., 2002a; 2002b; Lor-
and are present in all jawed vertebrates. It is striking to observe enzale et al., 2018; Satchell and Jones, 1967).
such a high proportion of lung-specific expression genes in the Through genome synteny comparisons, we found that over
bichir. These include two well-known lung development genes 15% of the bichir genome has conserved genomic synteny
(Tbx4 and Tbx5) that were apparently present in the common with the lobe-finned fish genomes but only 6% with teleosts
ancestor of cartilaginous and bony fishes. It is also consistent (Figure 6A). Interestingly, gene ontology (GO) enrichment anal-
with a previous report on an important lung-related regulatory ysis for genes shared the same synteny between non-teleost
enhancer (lung mesenchyme-specific enhancer, LME) of Tbx4, actinopterygians, and lobe-finned fishes showed significantly
which is present in cartilaginous fishes and also exists in the bi- enriched in the cardiac development-related GO terms (Fig-
chir, bowfin, and spotted gar (Hara et al., 2018; Tatsumi et al., ure S6A), supporting the different cardiac functions displayed
2016) (Figure S5D). In situ hybridization experiments with five between the two groups. It also pinpointed that these highly
of these genes confirmed that they are all highly active in alveolar conserved synteny blocks over long evolutionary history might
cells of both the mouse and the bichir but weakly expressed in be favored by natural selection and possibly maintain the
the swim bladder surface of the zebrafish (Figure 5F). Previous similar function through conserved enhancer regulation that
analysis of fossils has shown that the pharynx has been drasti- present in the same genomic regulatory blocks (Kikuta
cally rebuilt during the origin of gnathostomes, and the lungs et al., 2007).
were derivatives of the pharynx posterior to the gills (Perry From the specifically conserved synteny between non-
et al., 2001). Considering that some of these lung-related genes teleost actinopterygians and sarcopterygians, we found a highly
(such as Tbx3) are also expressed in the pharyngeal region (Mes- conserved non-coding element (CNE) in all jawed vertebrates,
bah et al., 2008), we hypothesize that they might have appeared except the Neoteleostei, located near the Hand2 gene (Fig-
in the common ancestor of cartilaginous and bony fishes and ure 6B). Hand2 is essential for cardiac morphogenesis, espe-
play a role in forming the distinctive pharynx in gnathostome (Du- cially formation of the RV. Its expression is also positively corre-
pret et al., 2014; Kuratani and Ahlberg, 2018). lated with cardiomyocyte proliferation (Han et al., 2019). To
understand the function of this CNE, we knocked out the element
Concomitant changes in circulatory systems in mice, which resulted in perinatal lethality for homozygous
Vertebrates have evolved sophisticated cardio-respiratory sys- mutated animals (Figures S6B and S6C). The homozygous
tems. The respiratory system provides an adequate supply of knockout mice developed a thin, small RV and hence congenital
oxygen to maintain proper cardiac function (under conditions heart defects (Figures 6C and S6D). We also detected weaker
that are not excessively stressful), while the circulatory system expression of Hand2 in the RV of homozygous knockout than
efficiently transfers the oxygen to other organs. During the in the heterozygous knockouts (Figure 6D). Thus, our knockout
evolution of air-breathing respiration, selection pressures may experiment suggested that conserved CNE has a regulatory
have driven co-adaptive changes in the circulatory system. In role in Hand2 expression patterns. We also detected this CNE
the lung circulatory system, the oxygen-depleted blood is in all selected non-Neoteleostei teleosts (Elopomorpha, Osteo-
pumped away from the heart to lungs through the pulmonary ar- glossomorpha, Otocephala, and Protacanthopterygii) but not
tery. After oxygenation, the blood flows back to heart through other crown teleost groups (Figure 6E). This is consistent with
the pulmonary veins. However, most air-breathing fish did not the anatomical observation that a conus arteriosus-like structure
evolve the specialized chamber to separate the two streams of is present in the cardiac outflow tracts of basal teleost lineages
blood and thus mix deoxygenated and oxygenated blood in their but has been completely lost in advanced teleosts, which have
circulatory system (Ishimatsu, 2012). It has been hypothesized a single chamber (bulbus arteriosus) in these tracts (Icardo,
that dominance of the conus arteriosus was an ancestral charac- 2006). Thus, we speculate that loss of this CNE in the Neoteleos-
teristic of the vertebrate cardiac outflow tract (Icardo, 2006; tei might explain the complete absence of conus arteriosus in
Lorenzale et al., 2018). The conus arteriosus emerged in the these fishes, which have developed a diversified cardiovascular
ancestor of jawed vertebrates; is preserved in cartilaginous system that co-appeared with the gas bladder respiratory
defect out of the total hearts analyzed. The cell nuclei are stained blue with hematoxylin, and the extracellular matrix and cytoplasm pink with eosin. LA, left atrium;
LV, left ventricle; RA, right atrium; RV, right ventricle; C.A, conus arteriosus; B.A, bulbus arteriosus; VE, ventricle.
(D) Results of qRT-PCR analysis of Hand2 gene expression in cardiomyocytes dissected from E16.5 embryonic mouse hearts. Data are mean ± SEM (*p < 0.05,
t test).
(E) Hypothetical transitions of cardiac outflow anatomic structure. Dominance of the conus arteriosus in the cardiac outflow tract evolved in the common ancestor
of jawed vertebrates and was preserved in chondrichthyans, sarcopterygians, and non-teleost actinopterygians. A conus arteriosus-like structure can still be
detected in some basal teleosts but has been completely lost in other teleosts.
Cell 184, 1–15, March 4, 2021 11Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
system in adaptation to aquatic environments with low oxygen oratory of Freshwater Ecology and Biotechnology, Institute of Hydrobiology,
tensions (Harter and Brauner, 2017). Chinese Academy of Sciences and from the National Natural Science Founda-
tion of China (41876179 and 31972866) to S.H. and funded by the Strategic Pri-
ority Research Program of the Chinese Academy of Sciences grants (no.
DISCUSSION XDB13000000 and no. XDB31020000) and a Villum Investigator grant (no.
25900) from The Villum Foundation to G.Z. It was also supported by the
The high-quality genome assemblies of four basal ray-finned Guangdong Provincial Academician Workstation of BGI Synthetic Genomics
fishes obtained in this study have filled gaps in knowledge of (no. 2017B090904014) to H. Yang and Youth Innovation Promotion Associa-
tion, Chinese Academy of Sciences to L.Y. (http://www.yicas.cn).
early evolutionary processes of bony fish genomes. We find
these fishes possess many ancestral genomic features of the
bony fishes. Particularly, it is remarkable that many limb- and AUTHOR CONTRIBUTIONS
lung-related genes and associated regulatory elements are pre-
G.Z., S.H., M.Z., and W.W. conceived the study. L.Y. and Haifeng Jiang
sent in cartilaginous and bony fishes, as validated by a number of collected the materials. L.Y., Haifeng Jiang, X.G., and H.Z. performed the
experiments, suggesting that the ancestral regulatory networks morphological laboratory work. X.B. and H. Yu performed the genome assem-
that latterly used to participate in the terrestrial locomotion and bly and genome annotation. H.P. and X.B. designed evolutionary analyses.
air-breathing functions have evolved in early jawed vertebrates. X.B., H.P., K.W., M.F., C.Z., Y.C., and Y.H. performed evolutionary analyses.
Our study suggests some of these ancestral developmental S.H., M.Z., W.W., H.P., Q.W., Y.E.Z., H. Yang, Q.Q., Huifeng Jiang, Y.Z., and
regulation networks have been preserved in non-teleost actino- D.Y. participated in discussions and provided suggestions. G.Z., X.B., and
K.W. wrote the manuscript with the input from all co-authors.
pterygians, which might explain their mosaic biological features
between lobe-finned fishes and ray-finned fishes. These ances-
tral developmental potentials have also facilitated the adaptive DECLARATION OF INTERESTS
evolution of many phenotypes that contributed to the vertebrate
The authors declare no competing interests.
landing.
Received: July 15, 2020
STAR+METHODS Revised: November 11, 2020
Accepted: January 27, 2021
Published: February 4, 2021
Detailed methods are provided in the online version of this paper
and include the following:
REFERENCES
d KEY RESOURCES TABLE
d RESOURCE AVAILABILITY Allen, J.A. (1876). The American bisons, living and extinctVol. 10 (Univer-
sity Press).
B Lead contact
B Materials availability Amemiya, C.T., Alföldi, J., Lee, A.P., Fan, S., Philippe, H., Maccallum, I.,
Braasch, I., Manousaki, T., Schneider, I., Rohner, N., et al. (2013). The African
B Data and code availability
coelacanth genome provides insights into tetrapod evolution. Nature 496,
d EXPERIMENTAL MODEL AND SUBJECT DETAILS 311–316.
B Source organism
Anagnostopoulou, P., Riederer, B., Duerr, J., Michel, S., Binia, A., Agrawal, R.,
d METHOD DETAILS Liu, X., Kalitzki, K., Xiao, F., Chen, M., et al. (2012). SLC26A9-mediated chlo-
B Genome sequencing, assembly and annotation ride secretion prevents mucus obstruction in airway inflammation. J. Clin.
B Phylogeny reconstruction Invest. 122, 3629–3634.
B Reconstruction of ancestral karyotypes Andreev, P.S., Coates, M.I., Shelton, R.M., Cooper, P.R., Smith, M.P., and
B Synteny analysis Sansom, I.J. (2015). Upper Ordovician chondrichthyan-like scales from North
B Whole genome duplication events analysis America. Palaeontology 58, 691–704.
B Limb enhancer identification Apweiler, R., Attwood, T.K., Bairoch, A., Bateman, A., Birney, E., Biswas, M.,
B Fin regeneration and Osr2 in situ expression Bucher, P., Cerutti, L., Corpet, F., Croning, M.D., et al. (2001). The InterPro
B Identification of OR and VR genes database, an integrated documentation resource for protein families, domains
and functional sites. Nucleic Acids Res. 29, 37–40.
B Transcriptome analysis
B In situ hybridization of lung-specific genes Arora, R., Metzger, R.J., and Papaioannou, V.E. (2012). Multiple roles and in-
teractions of Tbx4 and Tbx5 in development of the respiratory system. PLoS
B Knockout of the Hand2 CNE
Genet. 8, e1002866.
d QUANTIFICATION AND STATISTICAL ANALYSIS
Askary, A., Smeeton, J., Paul, S., Schindler, S., Braasch, I., Ellis, N.A., Post-
lethwait, J., Miller, C.T., and Crump, J.G. (2016). Ancient origin of lubricated
Supplemental information joints in bony vertebrates. eLife 5, e16415.
Supplemental Information can be found online at https://doi.org/10.1016/j.
cell.2021.01.046. Birney, E., Clamp, M., and Durbin, R. (2004). GeneWise and Genomewise.
Genome Res. 14, 988–995.
ACKNOWLEDGMENTS Braasch, I., Gehrke, A.R., Smith, J.J., Kawasaki, K., Manousaki, T., Pasquier,
J., Amores, A., Desvignes, T., Batzel, P., Catchen, J., et al. (2016). The spotted
We thank Qiwu Xu, Guangyi Fan, and Xiaoli Chen for Hi-C library construction gar genome illuminates vertebrate evolution and facilitates human-teleost
and sequencing, and also thank the support from the Fish10K project and Wu- comparisons. Nat. Genet. 48, 427–437.
han Frasergen Bioinformatics Co., Ltd. We thank Tom Gilbert for the helpful Carmeliet, P., and Jain, R.K. (2011). Molecular mechanisms and clinical appli-
suggestions. This research was supported by grants from the State Key Lab- cations of angiogenesis. Nature 473, 298–307.
12 Cell 184, 1–15, March 4, 2021Please cite this article in press as: Bi et al., Tracing the genetic footprints of vertebrate landing in non-teleost ray-finned fishes, Cell (2021),
https://doi.org/10.1016/j.cell.2021.01.046
ll
Resource
Carroll, R.L., and Holmes, R. (2007). Evolution of the appendicular skeleton of Giles, S., Xu, G.-H., Near, T.J., and Friedman, M. (2017). Early members
amphibians. In Fins into Limbs: Evolution, Development, and Transformation, of ‘living fossil’ lineage imply later origin of modern ray-finned fishes. Nature
B.K. Hall, ed. (The University of Chicago Press), pp. 185–224. 549, 265–268.
Chauve, C., and Tannier, E. (2008). A methodological framework for the recon- Glasauer, S.M., and Neuhauss, S.C. (2014). Whole-genome duplication
struction of contiguous regions of ancestral genomes and its application to in teleost fishes and its evolutionary consequences. Mol. Genet. Genomics
mammalian genomes. PLoS Comput Biol. 4, e1000234. 289, 1045–1060.
Chen, Y., Chen, Y., Shi, C., Huang, Z., Zhang, Y., Li, S., Li, Y., Ye, J., Yu, C., Li, Goldman, N., and Yang, Z. (1994). A codon-based model of nucleotide substi-
Z., et al. (2018). SOAPnuke: a MapReduce acceleration-supported software tution for protein-coding DNA sequences. Mol. Biol. Evol. 11, 725–736.
for integrated quality control and preprocessing of high-throughput
Goujet, D. (2011). ‘‘Lungs’’ in Placoderms, a persistent palaeobiological myth
sequencing data. Gigascience 7, 1–6.
related to environmental preconceived interpretations. Comptes Rendus Pale-
Clack, J.A. (2009). The fin to limb transition: new data, interpretations, and hy- vol 10, 323–329.
potheses from paleontology and developmental biology. Annual Review of
Graham, J.B., Wegner, N.C., Miller, L.A., Jew, C.J., Lai, N.C., Berquist, R.M.,
Earth and Planetary Sciences 37, 163–179.
Frank, L.R., and Long, J.A. (2014). Spiracular air breathing in polypterid fishes
Coates, M.I., Gess, R.W., Finarelli, J.A., Criswell, K.E., and Tietjen, K. (2017). and its implications for aerial respiration in stem tetrapods. Nat. Commun.
A symmoriiform chondrichthyan braincase and the origin of chimaeroid fishes. 5, 3022.
Nature 541, 208–211.
Grande, L., Jin, F., Yabumoto, Y., and Bemis, W.E. (2002). Protopsephurus liui,
Morris, S.C. (2008). A redescription of a rare chordate, metaspriggina walcotti
a well-preserved primitive paddlefish (Acipenseriformes: Polyodontidae) from
Simonetta and Insom, from the Burgess shale (middle Cambrian), British
the Lower Cretaceous of China. Journal of Vertebrate Paleontology 22,
Columbia, Canada. Journal of Paleontology 82, 424–430.
209–237.
Cope, E.D. (1878). Descriptions of New Extinct Vertebrata from the Upper Ter-
Han, X., Zhang, J., Liu, Y., Fan, X., Ai, S., Luo, Y., Li, X., Jin, H., Luo, S., Zheng,
tiary and Dakota Formations. (Harvard University).
H., et al. (2019). The lncRNA Hand2os1/Uph locus orchestrates heart develop-
Cuervo, R., Hernández-Martı́nez, R., Chimal-Monroy, J., Merchant-Larios, H., ment through regulation of precise expression of Hand2. Development 146,
and Covarrubias, L. (2012). Full regeneration of the tribasal Polypterus fin. dev176198.
Proc. Natl. Acad. Sci. USA 109, 3838–3843.
Hara, Y., Yamaguchi, K., Onimaru, K., Kadota, M., Koyanagi, M., Keeley, S.D.,
Darwin, C. (1859). On the Origin of Species. (Harvard University Press).
Tatsumi, K., Tanaka, K., Motone, F., Kageyama, Y., et al. (2018). Shark ge-
Davis, M.C., Shubin, N.H., and Force, A. (2004). Pectoral fin and girdle devel- nomes provide insights into elasmobranch evolution and the origin of verte-
opment in the basal actinopterygians Polyodon spathula and Acipenser trans- brates. Nat. Ecol. Evol. 2, 1761–1771.
montanus. J. Morphol. 262, 608–628.
Haro, E., Watson, B.A., Feenstra, J.M., Tegeler, L., Pira, C.U., Mohan, S., and
Denison, R.H. (1941). The soft anatomy of Bothriolepis. Journal of Paleon- Oberg, K.C. (2017). Lmx1b-targeted cis-regulatory modules involved in limb
tology 15, 553–561. dorsalization. Development 144, 2009–2020.
Du, K., Stöck, M., Kneitz, S., Klopp, C., Woltering, J.M., Adolfi, M.C., Feron, R., Harris, R.S. (2007). Improved pairwise Alignment of genomic DNA. PhD thesis
Prokopov, D., Makunin, A., Kichigin, I., et al. (2020). The sterlet sturgeon (Pennsylvania State University).
genome sequence and the mechanisms of segmental rediploidization. Nat.
Ecol. Evol. 4, 841–852. Harter, T.S., and Brauner, C.J. (2017). The O2 and CO2 Transport System in
Teleosts and the Specialized Mechanisms That Enhance Hb–O2 Unloading
Dudchenko, O., Batra, S.S., Omer, A.D., Nyquist, S.K., Hoeger, M., Durand,
to Tissues. In Fish Physiology, A.K. Gamperl, T.E. Gillis, A.P. Farrell, and
N.C., Shamim, M.S., Machol, I., Lander, E.S., Aiden, A.P., and Aiden, E.L.
C.J. Brauner, eds. (Elsevier), pp. 1–106.
(2017). De novo assembly of the Aedes aegypti genome using Hi-C yields
chromosome-length scaffolds. Science 356, 92–95. Hinchliffe, J.R. (1994). Evolutionary developmental biology of the tetrapod
limb. Dev. Suppl. 1994, 163–168.
Dupret, V., Sanchez, S., Goujet, D., Tafforeau, P., and Ahlberg, P.E. (2014).
A primitive placoderm sheds light on the origin of the jawed vertebrate face. Hong, T.-M., Chen, Y.-L., Wu, Y.-Y., Yuan, A., Chao, Y.-C., Chung, Y.-C., Wu,
Nature 507, 500–503. M.-H., Yang, S.-C., Pan, S.-H., and Shih, J.-Y. (2007). Targeting neuropilin 1 as
an antitumor strategy in lung cancer. Clin. Cancer Res. 13, 4759–4768.
Edgar, R.C. (2004). MUSCLE: a multiple sequence alignment method with
reduced time and space complexity. BMC Bioinformatics 5, 113. Hrbek, T., Seckinger, J., and Meyer, A. (2007). A phylogenetic and biogeo-
Evans, B.J. (2008). Genome evolution and speciation genetics of clawed frogs graphic perspective on the evolution of poeciliid fishes. Molecular Phyloge-
(Xenopus and Silurana). Front. Biosci. 13, 4687–4706. netics and Evolution 43, 986–998.
Farmer, C.G. (1999). Evolution of the vertebrate cardio-pulmonary system. Hurley, I.A., Mueller, R.L., Dunn, K.A., Schmidt, E.J., Friedman, M., Ho, R.K.,
Annu. Rev. Physiol. 61, 573–592. Prince, V.E., Yang, Z., Thomas, M.G., and Coates, M.I. (2007). A new time-
scale for ray-finned fish evolution. Proc. Biol. Sci. 274, 489–498.
Frazer, K.A., Pachter, L., Poliakov, A., Rubin, E.M., and Dubchak, I. (2004).
VISTA: computational tools for comparative genomics. Nucleic Acids Res. Icardo, J.M. (2006). Conus arteriosus of the teleost heart: dismissed, but not
32, W273-9. missed. Anat. Rec. A Discov. Mol. Cell. Evol. Biol. 288, 900–908.
Friedman, M. (2015). The early evolution of ray-finned fishes. Palaeontology Icardo, J.M., Colvee, E., Cerra, M.C., and Tota, B. (2002a). Structure of the
58, 213–228. conus arteriosus of the sturgeon (Acipenser naccarii) heart. I: the conus valves
Gao, Y., Lan, Y., Liu, H., and Jiang, R. (2011). The zinc finger transcription fac- and the subendocardium. Anat. Rec. 267, 17–27.
tors Osr1 and Osr2 control synovial joint formation. Dev. Biol. 352, 83–91. Icardo, J.M., Colvee, E., Cerra, M.C., and Tota, B. (2002b). The structure of the
Gardiner, B.G., Schaeffer, B., and Masserie, J.A. (2005). A review of the lower conus arteriosus of the sturgeon (Acipenser naccarii) heart: II. The myocar-
actinopterygian phylogeny. Zoological Journal of the Linnean Society 144, dium, the subepicardium, and the conus-aorta transition. Anat. Rec. 268,
511–525. 388–398.
Gibert, J.M. (2017). The flexible stem hypothesis: evidence from genetic data. Inoue, J.G., Miya, M., Tsukamoto, K., and Nishida, M. (2003). Basal actino-
Dev. Genes Evol. 227, 297–307. pterygian relationships: a mitogenomic perspective on the phylogeny of the
Giles, S., Coates, M.I., Garwood, R.J., Brazeau, M.D., Atwood, R., Johanson, ‘‘ancient fish’’. Mol. Phylogenet. Evol. 26, 110–120.
Z., and Friedman, M. (2015). Endoskeletal structure in Cheirolepis (Osteich- Ishimatsu, A. (2012). Evolution of the cardiorespiratory system in air-breathing
thyes, Actinopterygii), An early ray-finned fish. Palaeontology 58, 849–870. fishes. Aqua-BioScience Monographs 5, 1–28.
Cell 184, 1–15, March 4, 2021 13You can also read