Cheats as first propagules: A new hypothesis for the evolution of individuality during the transition from single cells to multicellularity
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Insights & Perspectives
Hypotheses
Cheats as first propagules:
A new hypothesis for the evolution of
individuality during the transition from
single cells to multicellularity
Paul B. Rainey1) and Benjamin Kerr2)
The emergence of individuality during the evolutionary transition from single Introduction
cells to multicellularity poses a range of problems. A key issue is how vari-
ation in lower-level individuals generates a corporate (collective) entity with The panoply of plant and animal form
Darwinian characteristics. Of central importance to this process is the evol- that defines life owes much to the rise of
multicellularity [1]. From a genetically
ution of a means of collective reproduction, however, the evolution of a
diverse range of starting positions, inde-
means of collective reproduction is not a trivial issue, requiring careful con- pendent unicellular lineages have made
sideration of mechanistic details. Calling upon observations from exper- the transition to multicellularity [2]. The
iments, we draw attention to proto-life cycles that emerge via most ancient transitions occurred in
unconventional routes and that transition, in single steps, individuality to the major lineages of large multicellular
eukaryotes approximately 1,000 million
higher levels. One such life cycle arises from conflicts among levels of selec-
years ago [3]. Multicellularity has also
tion and invokes cheats as a primitive germ line: it lays the foundation for arisen in the ciliates, slime molds, dia-
collective reproduction, the basis of a self-policing system, the selective toms, and certain groups of prokaryotes
environment for the emergence of development, and hints at a plausible ori- [2, 4–7]; most recently it has occurred in
gin for a soma/germ line distinction. the volvocine algae [8–11].
.
The evolution of multicellularity
involved a hierarchical shift in
Keywords:
Darwinian individuality during which
biological complexity; conflict; cooperation; experimental evolution; multi-level
individual cells relinquished their
selection capacity to reproduce as independent
units and came to reproduce as part
of a larger whole [12, 13]. Explaining this
shift in selection – from individual cells
to groups of cells – poses a range of
significant problems. Okasha [13] sum-
marizes: ‘‘The challenge is to understand
DOI 10.1002/bies.201000039 [. . .] transitions in Darwinian terms. Why
was it advantageous for the lower-level
1)
New Zealand Institute for Advanced Study and Abbreviations: units to sacrifice their individuality and
Allan Wilson Centre for Molecular Ecology & MLS, multi-level selection; WS, wrinkly spreader. form themselves into a corporate body?
Evolution, Massey University, Auckland,
And how could such an arrangement,
New Zealand
2)
Department of Biology, University of once first evolved, be evolutionarily
Washington, Box 351800 Seattle, Washington stable?’’ Equally, one might focus on
98195, USA the higher level and ask how individu-
ality emerges at the level of the corpor-
*Corresponding author:
Paul B. Rainey ate body. In placing the emphasis on
E-mail: p.b.rainey@massey.ac.nz individuality at the higher level [14]
Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc. www.bioessays-journal.com 1P. B. Rainey and B. Kerr Insights & Perspectives .....
there is recognition that individuality is However, the transition to multicellular- functionality and independent of the
a derived character and one that ity is far more than the evolution of reproductive properties of the individ-
requires an evolutionary explanation cooperation. Critical for the evolution ual cells. While such a scenario
Hypotheses
[15]. The key issue is to explain how of multicellular organisms is the evol- describes plausible changes, the model
variation in lower-level individuals gen- ution of group level adaptations includ- assumes that the capacity to leave group
erates a corporate entity with Darwinian ing group reproduction, mechanisms to offspring is already in place. But how
characteristics [16]. In this context suppress cheating, and the emergence such a new level of reproduction
we argue that the critical problem is of development and differentiation. The emerges requires explanation.
the evolution of a means of collective focus of attention thus shifts from traits
reproduction. that are defined by the properties of
The obvious solution is a life cycle: individual entities to traits that are the
life cycles involving single-cell bottle- properties of groups of cells. This shift
The evolutionary
necks are a ubiquitous feature of multi- marks a significant alteration in emergence of group
cellular life [15, 17, 18]: life cycles allow perspective and a move to the MLS-2 reproduction
collectives to produce offspring. Despite framework [20]. However, in MLS-2,
their biological significance, the evol- group fitness is defined independently From a theoretical perspective the shift
utionary origins of life cycles are unclear of particle fitness. The most successful from MLS-1 to MLS-2 encapsulates an
[15, 19]. Here, informed by experimental groups are those that contribute the evolutionary transition in individuality.
studies, we draw attention to critical greatest number of group offspring to The transition completes when the
issues and mechanistic problems that the next generation irrespective of the higher-level entities become Darwinian
lie at the heart of life cycle evolution. number of cells those groups contain. individuals, that is, when populations
We suggest solutions – albeit of an Thus, fitness in MLS-1 and MLS-2 con- of these organisms display variation,
unconventional sort – and even go so texts is different: in the MLS-1 context, heritability, and reproduction. Thus,
far as to suggest that one route to a fitness is the number of offspring one critical trait that marks individuality
proto-life cycle may have been fueled particles, whereas, in MLS-2, the num- at the higher level is the capacity for
by the tension inherent in levels of selec- ber of offspring collectives defines fit- groups to leave offspring groups.
tion and may have involved cheating ness. While this makes intuitive – and Reproduction of collectives requires
genotypes as propagules. theoretical – sense [13], it does not development and a life cycle, which is
amount to an explanation: just how not something that newly formed
individuality transfers from particles groups are necessarily born with
The multi-level selection to collectives is a profound problem. [15, 19, 30]. When considering the evol-
framework Theoretical studies of Michod and utionary origins of such a capability –
Nedelcu have made important contri- particularly via natural selection – prob-
Multi-level selection (MLS) theory butions, particularly the concept of fit- lems arise. The evolution of traits adap-
[13, 20–22] provides a powerful theoreti- ness decoupling: the need – during an tive at a given level of biological
cal framework within which to consider evolutionary transition – for fitness at organization requires the existence –
major evolutionary transitions. During the higher level to become decoupled at that level – of the necessary prereq-
initial stages of the transition from from the fitness of lower level [29]. uisites for Darwinian individuality
single cells to multicellularity, the focus While being a seminal insight, the [16, 31–34]. When the trait whose origin
is individual cells. Given appropriate mechanism by which it comes about is we wish to explain is reproduction we
ecological conditions [23–25], selection unclear. For example, Michod [14] uses face a dilemma: appeals to natural
favors the evolution of simple undiffer- a simple model for the evolution of mul- selection would seem to presuppose
entiated groups – arising, for example, ticellularity that begins with ‘‘adult’’ the existence of collective reproduction
from the production of adhesive glues organisms comprised of two cell types – the very trait whose evolution requires
[25–28]. The cause of cooperation (pro- (cooperate and defect). Although the explanation. Griesemer foresaw pre-
duction of adhesive glues) is the prop- adult organisms are capable of produc- cisely this problem when he argued that
erty of the individual cells. Selection at ing offspring propagules, the pro- explaining the emergence of a new level
the higher (group) level affects the duction of propagules is not a of organization is necessary before
spread of the trait, but group fitness is consequence of adult functionality, invoking the evolution of adaptations
nothing more than the average (or sum) but rather is dependent on the average specific to that new level [30].
of the fitness of the individual cells that fitness of the individual entities of which Below we outline two adaptive
comprise the group. From a formal each adult is comprised. As Okasha [13] solutions in which individuality
perspective the spread of cooperation remarks, this is ‘‘a sort of gray area emerges at the very same moment that
is readily explained by kin selection between MLS-1 and MLS-2’’. Gradually, the capacity for groups to leave collec-
and traditional group selection theory as the transition proceeds, fitness tive offspring evolves. However, we also
and is encompassed by MLS-1 theory. becomes ‘‘decoupled’’ from the lower recognize the potential for non-adaptive
Within this MLS-1 framework, the fittest level and with this, individuality solutions. A third possibility is that evol-
groups are those that contribute the emerges at the level of the adult, to ution of a means of collective reproduc-
greatest number of individual cells to the point where the capacity to leave tion is not necessary and that selection
the next generation [13, 20]. offspring is a product of adult on group viability alone is sufficient.
2 Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc...... Insights & Perspectives P. B. Rainey and B. Kerr
Hypotheses
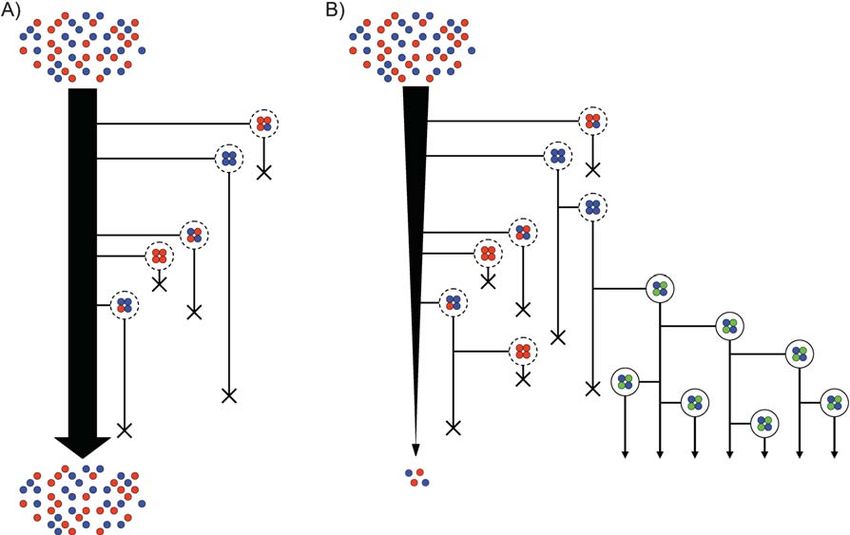
Figure 1. The role of group reproduction in group adaptation. A: A scenario is shown in policing, development, and differen-
which loose groups form from individual cells (given as red and blue circles). These groups tiation.
do not beget new groups, nor do they contribute individual cells back to the cell population. Imagine, however, that the viability
Natural selection can certainly act on these groups. For example, in the picture, groups with
process operates in tandem with a proc-
more blue cells live longer and therefore the frequency of blue cells within groups remains
high (this occurs even though the blue cells are at a frequency equal to the red cells within
ess by which groups are created from
the ‘‘free cell’’ population). However, there is no way for evolutionary innovations at the group the lower-level parts of pre-existing
level to propagate through this form of group viability selection (given finite group lifetimes). groups (Fig. 1B). For selection to work
For example, it is not the case that groups with blue cells are more likely to form in future creatively – and potently – on the higher
generations because they have a viability advantage at the group level. B: A scenario is level it is crucial for groups to beget
shown where group reproduction occurs. This opens the door for fecundity selection at the groups. But this returns us to the para-
level of groups. In this picture, if a group possesses an innovation improving its survival or
doxical situation described above:
reproduction, then the innovation can be passed on to daughter groups. For example, the
production of specialized cell types (shown in green) leads to a proliferation of groups with namely, that the capacity of groups to
these specialized cells. Such a scheme requires both group reproduction and heredity of the beget groups requires groups to have
developmental program. In this figure we surround the constituent cells with a solid outer evolved this capacity.
circle as they now have some of the properties associated with a higher-level individual (i.e.
differentiation of parts and capacity to reproduce). If these groups compete with their free cell
cousins and group formation confers advantages, then this population could shift from lower- Insights from experiments
level individuals to higher-level individuals, thereby accomplishing a major transition.
Our experimental work uses popu-
lations of the bacterium Pseudomonas
fluorescens. When propagated in a
This final option we consider unlikely ancestral state (which is readily envis- spatially structured environment, the
and explain why in the next section. aged), then selection will favor the most ancestral bacterium diversifies produc-
viable groups (Fig. 1A). Although such ing a range of niche specialist genotypes
groups are seen by selection, the con- [35]. Among the numerous emergent
The inadequacy of viability nection between the consequences of forms is a class of genotypes collectively
selection selection at the level of groups at one known as wrinkly spreader (WS), which
point in time and the properties of form a self-supporting mat at the air-
The absence of a means of collective groups at a latter point in time is lack- liquid interface (Fig. 2).
reproduction does not mean that selec- ing. The only connection is via the WS genotypes arise from a wide
tion cannot act on collectives, but its lower-level entities. It is difficult to see range of simple mutations that result
capacity to do so is limited to selection how viability selection alone could in over-activation of adhesive factors
at the level of collective viability. result in the evolution of true group- (a cellulosic polymer and a protein-
Provided that simple undifferentiated level traits such as the capacity for aceous factor) [36–38]. The overproduc-
groups can evolve repeatedly from the group reproduction, let alone, self- tion of ‘‘glues’’ causes cells to remain
Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc. 3P. B. Rainey and B. Kerr Insights & Perspectives .....
requires that the cells within each group
periodically switch off traits that deter-
mine social behavior and then reactivate
Hypotheses
their expression to form new groups.
This requires the existence of develop-
mental control – a group-level trait – the
evolution of which raises the problems
discussed above. In the absence of a
means of regulating social behavior,
newly formed groups are driven extinct
Figure 2. The rise, fall, and destruction of a simple undifferentiated group. Left: The wrinkly
spreader mat is the cumulative product of the cooperative interactions of millions of cells. By
by selfish types.
working together the cells in the mat colonize the air-liquid interface – a niche that is unavail- One way forward would be for group
able for the ancestral (broth-colonizing) type. In colonizing this new niche the cells of the mat reproduction to be effected by an exter-
are rewarded with an abundance of oxygen. Middle: When the mat becomes too heavy, it nal factor, for example, stochastic
collapses into the broth (it is not buoyant). The collapse is hastened by the presence of disturbance of microcosms. Indivi-
cheating genotypes that grow like a cancer within the mat adding no structural strength, but duality of a kind would therefore be
reaping the benefits (access to oxygen). Right: A mat is far more than the sum of the individ-
endowed to the groups, but it is difficult
ual parts. This photo was taken immediately after disturbing (with a brief shake) a microcosm
with an intact mat. The mat breaks into many pieces (just visible on the bottom) and does
to see how this haphazard means of
not spontaneously reform. While a mat will eventually re-emerge, it will do so by a process of reproduction would be effective.
growth and development from a limiting inoculum. Dawkins [34] comes to a similar con-
clusion regarding the difficulty of organ-
ismal adaptation given reproduction
through a type of slapdash fissioning.
attached after cell division. While there evolution of cheating (selfish) types is to
is a significant fitness cost to each indi- be expected. Such types evolve and
vidual WS mutant [25, 39, 40], WS cells grow as a cancer within the mat. Life cycles: Solutions and
nonetheless increase in frequency ulti- Cheats do not produce adhesive poly- transitions
mately out-competing the ancestral gen- mers and therefore grow rapidly – they
otype. They achieve this because the are also highly motile. Provided they For Dawkins, adaptive evolution at the
cost to individual cells is traded against arise within the fabric of the mat then level of the multicellular organism
a benefit that accrues to the group of WS they reap the benefits of group member- requires a developmental cycle (e.g.
cells. It works as follows: the production ship (access to oxygen) while forgoing multicellular differentiation from a
of adhesive glues means that upon the cost of polymer production: in doing single-cell origin each generation).
binary fission, daughter cells remain so they make no contribution to the net- However, to avoid the pitfall of invoking
linked. Continuing cell division causes work of polymeric strands required for group reproduction as a precondition
the population of cells to expand in a maintenance of mat integrity. As might for its own evolution any adaptive
single-cell layer across the air-liquid be anticipated, the cancerous growths solution to the evolution of a life cycle
interface ultimately joining and becom- compromise the WS mat, and it ulti- would appear to require the emergence
ing attached to the edge of the glass vial. mately collapses [25] (Fig. 2): a classic of a life cycle concomitant with the tran-
Once the surface is colonized, the mat tragedy of the commons [43]. sition in individuality. While seemingly
grows in thickness, becoming a robust The emergence of groups leads to improbable, we outline two scenarios,
structure that is the cumulative product questions as to their further evolution. the first arising directly from experimen-
of the cooperative interactions of many At this point, standard (MLS-1) group tal studies.
millions of cells. By working together, selection models are invoked, but it Consider the model Pseudomonas
the cells in the mat colonize a niche becomes apparent that such models fail populations: the moment the number
unavailable to the ancestral type. In col- to fit with the biological reality of newly of WS cells become sufficient to form
onizing this new niche the cells of the formed WS groups. Standard group a mat the stage is set for the evolution
mat are rewarded with an abundance of selection models effectively explain of cheating types. Cheats, while being
oxygen [25]. the maintenance of cooperation in the the nemesis of the mat, are also its
The evolution of a WS mat involves face of selfish types that emerge as a potential savior. Cheats have character-
the evolution of cooperation – de novo consequence of selection at the lower istics of propagules: they can disperse
and in real time – from an ancestral level. In the absence of population from the mat – like a germ line they can
state that is asocial and unicellular. structure, selfish types ultimately out- regenerate WS, albeit upon further
The spread of polymer production is compete cooperating types causing their mutation (Fig. 3). Indeed, in the case
readily explained by kin selection [41, extinction. If population structure of Pseudomonas, the modular nature
42]. Baring mutation, clonal reproduc- exists, then cooperating types can be of the genetic architecture underlying
tion means that WS mats are comprised maintained provided there is periodic the evolution of WS genotypes provides
of individuals whose relatedness is com- dispersal of cells into a global popu- considerable evolutionarily flexibility
plete, the mat being a clone of geneti- lation, reassortment, followed by the [46, 47]. Ancestral genotypes readily
cally identical cells. Given mutation, the formation of new groups [44, 45]. This give rise to WS genotypes, which in turn
4 Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc...... Insights & Perspectives P. B. Rainey and B. Kerr
(and phenotypic) state inaccessible to
the ancestral genotype [52]. While adap-
2 3 4 tive in the new environment, the trait is
5
Hypotheses
not environmentally responsive. Critical
evolutionary events are thus required to
‘‘rewire’’ the organism, such that mat
formation comes under developmental
control. We suggest that continued
selection in an environment that favors
6 alternate phenotypic states (mat-form-
ing and cheating) provides such an
opportunity. In outlining this scenario
we recognize both parallels and differ-
ences with traditional and emerging
ideas surrounding the evolution of
developmental control [19, 49, 51, 53–
1 56].
7
Additional scenarios for the evol-
ution of life cycles that might effect the
transition from MLS-1 to MLS-2 can be
envisaged. Before considering non-adap-
Figure 3. A putative life cycle for mat-forming bacteria. We start with a single bacterium tive models for life cycle evolution, we
(given in blue) capable of producing an extracellular adhesive. (1) It reproduces at the inter- describe an alternative hypothesis in
face between liquid and air (in the case shown, starting at the inner surface of a glass tube). which, unlike the model above where
Daughter cells stick together because of the adhesive they produce. (2, 3) The resulting mat
the ‘‘germ line’’ is interrupted by
spreads over the liquid’s surface as a single-cell layer. (4) Due to prime access to oxygen, a
robust mat forms. Mutation generates ‘‘cheats’’ (green cells that do not produce any
mutation, here the germ line is uninter-
adhesive polymer and grow faster as a consequence). (5) These cheats spread like a cancer rupted by mutation. From the outset such
within the mat and contribute to (6) the collapse of the mat. Because the cheats do not a model is appealing because it removes
produce the adhesive, they are liberated from the mat upon collapse. (7) Back mutation from the potentially restrictive requirement of
one of these cheats to a mat-producing cell completes the life cycle. Of course, we do not mutation for the transition between
imagine such a life cycle playing out in an environment where only a single mat can form (like stages of the life cycle.
a single tube). Rather, the back mutants from the liberated cheats could establish mats in
Once again we make use of the
different locations from their parent mat. Here the cell type leading to the death of the group
also leads to its rebirth. The cheats amount to propagules (‘‘germ line’’), arising de novo from model Pseudomonas populations as a
the mat-forming ‘‘soma’’ of an incipient multicellular individual. vehicle for our ideas, but this time we
take as the focus of interest the lower-
level (cheating) entities. Consider the
cheating type as a totipotent germ line.
lose the mat-forming phenotype by necessary for the eventual integration Imagine that during the course of its
simple mutations that suppress pro- of ‘‘life cycle’’ phases within a single growth it produces, by chance
duction of the adhesive glues. The cohesive organism (Boxes 1 and 2). mutation, a cell type with which it
effects of these suppressor mutations Indeed, a recent experiment in which interacts, either directly, or indirectly,
can be readily reversed by mutations P. fluorescens cells were ‘‘forced’’ to and which, via that interaction, aids its
at additional loci [48]. Thus, from the transition between groups gives reason own reproductive output. We might
tension among levels of selection, a for optimism. After just four cycles, in consider this a ‘‘helper’’ type; indeed,
proto-life cycle emerges spontaneously two (of twelve) replicate lines, geno- we might consider the WS genotype an
(given appropriate ecological con- types arose that evolved the capacity exemplar of such a helper, although in
ditions) and with no requirement to to switch stochastically between states so doing we add a level of complexity
invoke group-level reproduction as a by an epigenetic mechanism [48]. (and selection) that is not necessary:
precondition. The emergence of such phenotype the helper may be any kind of repro-
A life cycle that requires mutation to switching is a critical event in the evol- ductive altruist. An interesting
transition the emerging ‘‘organism’’ ution of developmental control [49, 50]. example is provided by the suicidal
between phenotypic states is a far cry While the end product remains to be altruists of Salmonella typhimurium
from a developmentally regulated life experimentally realized, we envisage that die while preparing the ground
cycle; however, its existence is sufficient developmental control emerging as a for infection [57].
to allow selection to operate at the level multi-step process, the first stage being Nonetheless, returning to the
of the collective. Indeed, we suggest that the realization of a novel phenotypic familiar WS: as the mat forms it becomes
the proto-life cycle might provide the state (the mat-forming phenotype) – infiltrated by cells of the germ line
basis for the evolutionary emergence the result of selection in an ‘‘extraordi- which reap the advantage that accrues
of development – a ‘‘kick-start’’ – that nary environment’’ [51]. Mutation brings from growth at the air-liquid interface.
establishes the ecological conditions the existing pathway to an expression Eventually the mat collapses and the WS
Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc. 5P. B. Rainey and B. Kerr Insights & Perspectives .....
Box 1 example, the interrupted model carries
with it the initially burdensome require-
ment for mutation to mediate the tran-
Model for the development of a single mat
Hypotheses
sition between different stages of the life
cycle, whereas the uninterrupted model
Here we develop a simple discrete-time model to track differentiation within a requires only one-way mutation (to
mat and its eventual collapse. The model follows two cell types: mat formers dead-end helper cells). The uninter-
and cheats. We begin by describing the population dynamics within a single rupted model thus seems to offer a lower
mat. Assuming that every mat is initialized by a single mat former cell, then over hurdle for an evolutionary transition.
time, mutation generates cheats. Let m(t) and c(t) be the sizes of the mat However, things get more complex
former and cheat populations, respectively, in a single mat at time t. when one considers a second dis-
Populations within a mat grow according to the following branching tinguishing feature, namely the origin
process [70]: of multicellular differentiation. The
uninterrupted model requires the emer-
mðtÞ
X cðtÞ
X gence of extreme altruism via mutation
mðt þ 1Þ ¼ ½Xi Fi ðXi Þ þ Gj ðYj Þ; (1)
in the presence of would-be cheats. On
i¼1 j¼1
the other hand, the interrupted model
cðtÞ
X mðtÞ involves nothing more than the advent
X
cðt þ 1Þ ¼ Yj Gj ðYj Þ þ Fi ðXi Þ: (2) of cheats in the face of cooperation. We
j¼1 i¼1 do, however, note that for both models,
the reliance on mutation sets a lower
The sets {X1, X2, X3, . . .} and {Y1, Y2, Y3, . . .} contain independent and limit to the number of cells that com-
identically distributed (i.i.d.) Poisson-distributed random variables prise each collective.
(XPoisson(bm) and YPoisson(bc)). The ith mat former has Xi offspring cells, In outlining these two models our
whereas the jth cheat has Yj offspring cells. In this model, bm and bc are the intention has been to portray possible
average number of offspring cells per mat-forming cell and cheat, respectively, scenarios for the evolution of life cycles,
per unit of time (b0 s are birth factors). Because cheats reproduce without particularly the selective conditions
contributing to the integrity of the mat, we assume that these cells have a birth favoring ecologically distinct pheno-
rate advantage, i.e. bc > bm. types, that might eventually evolve to
The sets {F1, F2, F3, . . .} and {G1, G2, G3, . . .} contain i.i.d. binomially- come under regulatory (developmental)
distributed random variables (F(n)Binomial(n,mm,c) and G(n)Binomial control. The molecular details by which
(n,mc,m)). Of its Xi offspring, the ith mat former has Fi cheating mutants, and such control could emerge are unknown
of its Yj offspring, the jth cheat has Gj mat former mutants. For simplicity, we let but are likely to depend on non-adaptive
the probability of mutation from mat former to cheat (mm,c) and from cheat to processes such as mutation and genetic
mat former (mc,m) be equal: mm,c ¼ mc,m ¼ m (Box 2). drift [59], opportunities for co-option
The cell dynamics within a microbial mat are given by equations (1) and (2). [60, 61] (facilitated by mutation and
In addition, we assume that any mat has a finite lifetime (t ). The probability that drift) and the existence of plasticity
a mat collapses at time t ¼ T is given by: [49, 62]. Under some circumstances it
is even possible that the plasticity
Prðt ¼ T Þ ¼ 1expfðam mðT Þ þ ac cðT ÞÞg (3)
inherent in the genomic and regulatory
Thus, as the number of cells in a mat increase, the mat is more likely to organization of certain unicellular enti-
collapse. Again, because cheats do not contribute to mat integrity, they have a ties might be sufficient to produce a
disproportionate negative effect on the lifetime of the mat, i.e. ac > am. simple life cycle with minimal involve-
ment from selection. For example,
single cells driven to group formation
as a mechanism of predation-avoidance
lineage goes extinct; nonetheless, the term advantage gained from coloniza- might – given an appropriately organ-
germ line remains and in time gives rise tion of the oxygen-replete air-liquid ized and pre-prepared regulatory system
to further WS types which it again interface). From another perspective, – be capable of utilizing gradients
exploits for its own advantage. Such a the WS is an unfortunate pawn, sacri- generated across the colony as a means
scenario captures aspects of an earlier ficed by the germ line. of, for example, regulating the tran-
hypothesis in which the germ line Thus from different starting pos- sition between clumping and dispersing
originates as a consequence of ‘‘other itions we arrive at essentially the same behaviors [63].
cell lineages altruistically removing end point: in both, interrupted and An idea like this involving co-
themselves from the reproductive line uninterrupted models, there exists option of a life history gene has been
to perform some somatic benefit to the potential for the evolution of a life cycle suggested to explain the evolution of
organism’’ [58]. From one perspective, and with that exists potential to arrest reproductive altruism in the higher vol-
the WS is an extreme altruist, sacrificing in the germ line stage: individuality vocine algae [64]. The central idea is
its life for the germ line (altruism being in an MLS-2 sense is apparent. There that in the ancestral (unicellular) state
an indirect consequence of the short- are, however, some differences. For expression of the life history gene is
6 Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc...... Insights & Perspectives P. B. Rainey and B. Kerr
Box 2 pressure to lengthen mat generation time to maximize the
absolute number of cheat cells produced by a mat. We use
the model from Box 1 to identify the optimal mutation rate
Hypotheses
Adaptive developmental programs under r- and K-selection.
in mat populations As in Box 1, assume a given mat collapses at t. There
are c(t) cheats at this time, which we label c. Under
A life cycle initially dependent upon mutation and fueled by r-selection, we maximize growth rate of mats within a
conflicts among levels of selection appears understand- mat population. To do this, we consider the joint distri-
ably restrictive. In particular, there is the thorny issue of bution of t and c. Specifically, for any mat, we have:
heritability, which arises from the mutational lottery that Prðt ¼ T and c ¼ C Þ ¼ pðT ; C Þ
determines the fate of cells. However, this problem is not Armed with this distribution, the long-term growth rate
as great as it may first seem: the critical issue is the rate of (r) of a mat population with a specified developmental
transition between states and this rate is heritable. Indeed, program is given by the solution to the Euler-Lotka
the way a mat consigns cells to different categories via equation [71–73]:
mutation defines its developmental program. In turn, this X1 X 1
yields the life history of the mat. Thus, we focus on how pðT ; CÞmCe rT ¼ 1
changes in mutation rate (m) affect mat fitness. Here we T ¼0 C¼0
show how the developmental program can be adaptively For simplicity, we assume that a fraction m of the cheats
tuned to specific ecological conditions. mutate back to mat formers directly after the mat collapses.
Mat-level fitness is the ability of the mat to generate We use a Monte Carlo simulation approach to generate
offspring mats and is proportional to the number of cheats the joint distribution p. Specifically, we generate 50,000
contained in the mat upon its collapse. This fitness metric points (t, c)i using equations (1)–(3). An example of this
is fully adequate if mats always have the same generation joint distribution is shown in Fig. 4. In the figure we see the
time. However, the generation time of a mat is specified (at life history tradeoff faced by the mat: higher fecundity
least probabilistically) by its developmental program requires a longer generation time. With this joint distri-
[Eq. (3) in Box 1]. bution, we solve the following equation for r:
All else being equal, a shorter generation time is X mc e rti
50;000
i
beneficial within a growing population of mats. However, ¼1
50; 000
because cheats simultaneously contribute to mat repro- i¼1
duction and expiration, all else is not equal. For instance, if We then look for the mutation rate (m) that maximizes r.
a slightly longer-lived mat can have many more cheats For K-selection, we search for the mutation rate that
upon collapse, then it may be advantageous to live longer. maximizes c. We employ the same Monte Carlo approach
Different ecological circumstances will favor different to generate 50,000 c values, then we look for the mutation
developmental programs. Here, we consider two ecologi- rate that maximizes the average c value.
cal conditions. In the first (r-selection), sites for mat for- Figure 5 shows the results of this analysis: under r-
mation are always available, so there is a premium on a selection, high mutation rates are favored; under K-selec-
short mat generation time. Production of cheats should be tion, lower mutation rates are favored. Under r-selection,
adjusted as to maximize growth rate within an expanding longevity is sacrificed for a quick investment in cheats allow-
population of mats. In the second condition (K-selection), ing a rapid explosion of mats. Under K-selection, longer-
sites for mat formation are rarely encountered and there is lived mats are selectively favored to maximize cheat output.
Figure 4. The joint distribution of mat lon- Figure 5. Optimal mutation rates in mat development. A: Long-term growth (in an
gevity and mat fecundity. These points were r-selected environment) is shown as a function of the mutation probability. Here we see
generated from simulations of mat develop- higher mutation rates yielding faster growth of a lineage of mats. B: Mat fecundity (favored
ment given by Eqs. (1), (2), and (3) (bm ¼ in a K-selected environment) is maximized at lower rates of mutation. In parts A and B, the
4.0, bc ¼ 6.0, am ¼ 106, ac ¼ 105). parameters of the model are the same as those in Fig. 4.
Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc. 7P. B. Rainey and B. Kerr Insights & Perspectives .....
conditioned on an environmental cue, cycle, the basis of a self-policing system 9. Herron MD, Michod RE. 2008. Evolution of
complexity in the volvocine algae: transitions
but during the transition to multicellu- (Boxes 1 and 2), and ecological circum- in individuality through Darwin’s eye.
larity it evolves to come under the con- stances possibly conducive to the event- Evolution 62: 436–51.
Hypotheses
trol of spatial (developmental) signals. ual emergence of development. In 10. Bonner JT. 1998. The origins of multicellular-
ity. Integr Biol 1: 27–36.
Such a scenario makes a good deal of addition, the hypothesis provides a
11. Sachs JL. 2008. Resolving the first steps
sense and is even supported by studies plausible scenario for the origin of a to multicellularity. Trends Ecol Evol 5:
of regA expression (a regulator of chlor- soma/germ line distinction, and for 245–8.
oplast expression) in unicellular versus sequestration of the germ line by soma 12. Maynard Smith J, Szathmary E. 1995. The
Major Transitions in Evolution. Oxford:
multicellular volvocine algae [64]. – the latter arising from the fact that WS Freeman.
However, just how such a change ‘‘soma’’ is under strong selection to 13. Okasha S. 2006. Evolution and the Levels of
comes about – particularly the change check increased replication of cheating Selection. Oxford: Oxford University Press.
14. Michod RE. 1999. Darwinian Dynamics:
necessary to bring differentiation under germ line types. In this context it is Evolutionary Transitions in Fitness and
the control of endogenous signals – interesting to note recent ideas on the Individuality. Princeton: Princeton University
still requires an evolutionary expla- evolution of ageing as a deprivation syn- Press.
15. Buss LW. 1987. The Evolution of Individuality.
nation (see Ref. [65, 66] for a possible drome driven by the tension between
Princeton: Princeton University Press.
mechanism based on a viability- soma and germ line [69] – a tension that 16. Dennett DC. 1995. Darwin’s Dangerous Idea:
fecundity tradeoff). perhaps, at least for some evolutionary Evolution and the Meanings of Life. London:
transitions, may have an ancient past. Penguin Books. p 586.
17. Wolpert L. 1990. The evolution of develop-
ment. Biol J Linn Soc 39: 109–24.
Conclusion 18. Wolpert L, Szathmary E. 2002.
Acknowledgments Multicellularity: evolution and the egg.
Darwinian transitions in individuality, Nature 420: 745.
particularly those originating from a fra- This work is supported by the Marsden 19. Minelli A, Fusco G. 2010. Developmental
ternal alliance among lower-level entities Fund Council from government funding plasticity and the evolution of animal complex
administered by the Royal Society of life cycles. Phil Trans R Soc Lond, B 365: 631–
[58], pose some of the most tantalizing 40.
problems in biology. Here we have drawn New Zealand. We thank members of 20. Damuth J, Heisler IL. 1988. Alternative for-
attention to the need to explain, in mech- the Rainey and Kerr labs and particu- mulations of multi-level selection. Biol Phil 3:
anistic terms, how variation in lower- larly Katrin Hammerschmidt, Eric 407–30.
21. Heisler IL, Damuth J. 1987. A method for
level individuals generates a corporate Libby, Peter Meintjes, Jennifer
analyzing selection in hierarchically structured
entity with Darwinian characteristics Nemhauser, and Caroline Rose for val- populations. Am Nat 130: 582–602.
[16]. Our emphasis on this issue stems uable discussion. Brett Calcott and Kim 22. Sober E, Wilson DS. 1998. Unto Others: The
Sterelny provided insightful comments Evolution and Psychology of Unselfish
from the recognition that any explanation Behavior. Cambridge, MA: Harvard
for the evolution of multicellularity from on drafts of the manuscript. We also University Press.
unicells – for the transition between MLS- thank Andrew Moore and three anony- 23. Bell G. 1985. The origin and early evolution of
mous referees for constructive criticism germ cells as illustrated by the Volvocales. In
1 and MLS-2 – is dependent upon explain- Halvorson H, Mornoy A, ed; The Origin and
ing how collectives evolve the capacity leading to a significant improvement of Evolution of Sex. New York: Alan R. Liss. p
to leave collective offspring. The life the essay. 221–56.
24. Boraas ME, Seale DB, Boxhorn JE. 1998.
cycle, we argue, is the critical innovation: Phagotrophy by a flagellate selects for colo-
life cycles decouple fitness – they tran- nial prey: a possible origin of multicellularity.
sition individuality. References Evol Ecol 12: 153–64.
The unconventional life cycles that 25. Rainey PB, Rainey K. 2003. Evolution of
1. Conway Morris S. 1998. The Crucible of cooperation and conflict in experimental bac-
span the MLS-1 to MLS-2 juncture are Creation. Oxford: Oxford University Press. terial populations. Nature 425: 72–4.
founded in experimental reality. The 2. Bonner JT. 2000. First Signals: The Evolution 26. Velicer GJ, Yu YTN. 2003. Evolution of novel
interrupted life cycle model can operate of Multicellular Development. Princeton: cooperative swarming in the bacterium
Princeton University Press. Myxococcus xanthus. Nature 425: 75–8.
in experimental Pseudomonas popu- 3. Wray GA. 2001. Dating branches on the tree 27. Rokas A. 2008. The origins of multicellularity
lations and, via its operation, WS mats of life using DNA. Genome Biol 3: 1–7. and the early history of the genetic toolkit for
can assume the role of ‘‘organisms’’ – 4. Keim CN, Martins JL, Abreu F, et al. 2004. animal development. Annu Rev Genet 42:
Multicellular life cycle of magnetotactic 235–51.
organisms whose fitness is measured,
prokaryotes. FEMS Microbiol Lett 240: 28. Sebe-Pedros A, Roger AJ, Lang FB, et al.
not by the number of bacterial cells 203–8. 2010. Ancient origin of the integrin-mediated
within each mat, but by the number 5. Shapiro JA. 1998. Thinking about bacterial adhesion and signaling machinery. Proc Natl
of mat offspring left by parents. In advo- populations as multicellular organisms. Annu Acad Sci USA 107: 10142–47.
Rev Microbiol 52: 81–104. 29. Michod RE, Nedelcu AM. 2003. On the reor-
cating this model as one route to a 6. Tomitani A, Knoll AH, Cavanaugh CM, et al. ganization of fitness during evolutionary tran-
proto-life cycle we recognize the irony. 2006. The evolutionary diversification of cya- sitions in individuality. Integr Comp Biol 43:
Tensions between levels of selection are nobacteria: molecular-phylogenetic and pale- 64–73.
ontological perspectives. Proc Natl Acad Sci 30. Griesemer J. 2000. The units of evolutionary
typically viewed as significant impedi- USA 103: 5442–7. transition. Selection 1: 67–80.
ments to evolutionary transitions 7. Kaiser D. 2001. Building a multicellular organ- 31. Lewontin RC. 1970. The units of selection.
[12, 67, 68], but our altered perspective ism. Annu Rev Genet 35: 103–23. Ann Rev Ecol Syst 1: 1–18.
reveals a creative role for conflict. This 8. Kirk DL. 1998. Volvox: Molecular Genetic 32. Maynard Smith J. 1988. Evolutionary prog-
Origins of Multicellularity and Cellular ress and the levels of selection. In Nitecki MH,
conflict generates in a single step a Differentiation. Cambridge: Cambridge ed; Evolutionary Progress. Chicago:
means of collective reproduction, a life University Press. University of Chicago Press. p 219–30.
8 Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc...... Insights & Perspectives P. B. Rainey and B. Kerr
33. Hull DL. 1980. Individuality and selection. Ann 47. Beaumont HJE, Kassen R, Knight CG, et al. 60. True JR, Carroll SB. 2002. Gene co-option in
Rev Ecol Syst 11: 311–32. 2006. The genetics of phenotypic innovation. physiological and morphological evolution.
34. Dawkins R. 1982. The Extended Phenotype. In Logan N, ed; Prokaryotic Diversity: Annu Rev Cell Dev Biol 18: 53–80.
Oxford: Oxford University Press. Mechanisms and Significance. Cambridge: 61. Wu J, Zhao F, Wang S, et al. 2007. cTFbase:
Hypotheses
35. Rainey PB, Travisano M. 1998. Adaptive Cambridge University Press. p 91–104. a database for comparative genomics of tran-
radiation in a heterogeneous environment. 48. Beaumont HJ, Gallie J, Kost C, et al. 2009. scription factors in cyanobacteria. BMC
Nature 394: 69–72. Experimental evolution of bet hedging. Nature Genomics 8: 104.
36. Spiers AJ, Bohannon J, Gehrig SM, et al. 462: 90–93. 62. West-Eberhard MJ. 1989. Phenotypic
2003. Biofilm formation at the air-liquid inter- 49. West-Eberhard MJ. 2003. Developmental plasticity and the origins of diversity. Ann
face by the Pseudomonas fluorescens Plasticity and Evolution. Oxford: Oxford Rev Ecol Syst 20: 249–278.
SBW25 wrinkly spreader requires an acetyl- University Press. 63. Hochberg ME, Rankin DJ, Taborsky M.
ated form of cellulose. Mol Microbiol 50: 15– 50. Schlichting CD, Pigliucci M. 1993. Control of 2008. The coevolution of cooperation and dis-
27. phenotypic plasticity via regulatory genes. Am persal in social groups and its implications for
37. Spiers AJ, Kahn SG, Bohannon J, et al. Nat 142: 366–70. the emergence of multicellularity. BMC Evol
2002. Adaptive divergence in experimental 51. Lande R. 2009. Adaptation to an extraordi- Biol 8: 238.
populations of Pseudomonas fluorescens. I. nary environment by evolution of phenotypic 64. Nedelcu AM, Michod RE. 2006. The evol-
Genetic and phenotypic bases of wrinkly plasticity and genetic assimilation. J Evolution utionary origin of an altruistic gene. Mol Biol
spreader fitness. Genetics 161: 33–46. Biol 22: 1435–1446. Evol 23: 1460–144.
38. Spiers AJ, Rainey PB. 2005. The 52. Bantinaki E, Kassen R, Knight C, et al. 2007. 65. Michod RE, Viossat Y, Solari CA, et al. 2006.
Pseudomonas fluorescens SBW25 wrinkly Adaptive divergence in experimental popu- Life-history evolution and the origin of multi-
spreader biofilm requires attachment factor, lations of Pseudomonas fluorescens. III. cellularity. J Theor Biol 239: 257–272.
cellulose fibre and LPS interactions to main- Mutational origins of wrinkly spreader diver- 66. Michod RE. 2006. The group covariance
tain strength and integrity. Microbiology 151: sity. Genetics 176: 441–453. effect and fitness trade-offs during evolution-
2829–39. 53. Snell-Rood EC, Van Dyken JD, ary transitions in individuality. Proc Natl Acad
39. Knight CG, Zitzmann N, Prabhakar S, et al. Cruickshank T, et al. 2010. Toward a popu- Sci USA 103: 9113–9117.
2006. Unravelling adaptive evolution: how a lation genetic framework of developmental 67. Michod RE. 1996. Cooperation and conflict in
single point mutation affects the protein co- evolution: the costs, limits, and con- the evolution of individuality. 2. Conflict
regulation network. Nat Genet 38: 1015–22. sequences of phenotypic plasticity. mediation. Proc R Soc B 263: 813–822.
40. MacLean RC, Bell G, Rainey PB. 2004. The Bioessays 32: 71–81. 68. Jablonka E, Lamb MJ. 2006. The evolution of
evolution of a pleiotropic fitness tradeoff in 54. Pigliucci M, Murren CJ, Schlichting CD. information in the major transitions. J Theor
Pseudomonas fluorescens. Proc Natl Acad 2006. Phenotypic plasticity and evolution by Biol 239: 236–246.
Sci USA 101: 8072–7. genetic assimilation. J Exp Biol 209: 2362–7. 69. Heininger K. 2002. Aging is a deprivation
41. Hamilton WD. 1964. The genetical evolution 55. Schmalhausen II. 1949. Factors of Evolution: syndrome driven by a germ–soma conflict.
of social behavior. 1. J Theor Biol 7: 1–16. The Theory of Stabilizing Selection. Chicago: Aging Res Rev 1: 481–536.
42. Hamilton WD. 1964. The genetical evolution University of Chicago Press. 70. Haccou P, Jagers P, Vatutin V. 2005.
of social behavior. 2. J Theor Biol 7: 17–52. 56. Waddington CH. 1942. The canalization of Branching Processes: Variation, Growth,
43. Hardin G. 1968. The tragedy of the commons. development and genetic assimilation of and Extinction of Populations. New York:
Science 162: 1243–1248. acquired characters. Nature 150: 1008–23. Cambridge University Press.
44. Wilson DS. 1975. A theory of group selection. 57. Ackermann M, Stecher B, Freed NE, et al. 71. Euler L. 1760. Recherches générales sur la
Proc Natl Acad Sci USA 72: 143–146. 2008. Self-destructive cooperation mediated mortalité: la multiplication du genre humain.
45. Maynard Smith J. 1964. Group selection and by phenotypic noise. Nature 454: 987–990. Memoires de l’academie des sciences de
kin selection. Nature 201: 1145–6. 58. Queller DC. 2000. Relatedness and the fra- Berlin 16: 144–64.
46. McDonald MJ, Gehrig SM, Meintjes PL, ternal major transitions. Phil Trans R Soc Lond 72. Lotka AJ. 1925. Elements of Physical Biology.
et al. 2009. Adaptive divergence in exper- B 355: 1647–1655. Baltimore: Williams & Watkins.
imental populations of Pseudomonas fluores- 59. Lynch M. 2007. The frailty of adaptive hypoth- 73. Fisher RA. 1930. The Genetical Theory of
cens. IV. Genetic constraints guide eses for the origins of organismal complexity. Natural Selection. Oxford: Oxford University
evolutionary trajectories in a parallel adaptive Proc Natl Acad Sci USA 104 (Suppl 1): 8597– Press.
radiation. Genetics 183: 1041–53. 8604.
Bioessays 00: 000–000,ß 2010 WILEY Periodicals, Inc. 9You can also read

















































