Measles Virus Infects both Polarized Epithelial and Immune Cells by
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
JOURNAL OF VIROLOGY, May 2008, p. 4630–4637 Vol. 82, No. 9
0022-538X/08/$08.00⫹0 doi:10.1128/JVI.02691-07
Copyright © 2008, American Society for Microbiology. All Rights Reserved.
Measles Virus Infects both Polarized Epithelial and Immune Cells by
Using Distinctive Receptor-Binding Sites on Its Hemagglutinin䌤
Maino Tahara, Makoto Takeda,* Yuta Shirogane, Takao Hashiguchi, Shinji Ohno, and Yusuke Yanagi
Department of Virology, Faculty of Medicine, Kyushu University, Fukuoka 812-8582, Japan
Received 19 December 2007/Accepted 12 February 2008
Measles is one of the most contagious human infectious diseases and remains a major cause of childhood
morbidity and mortality worldwide. The signaling lymphocyte activation molecule (SLAM), also called CD150,
is a cellular receptor for measles virus (MV), presumably accounting for its tropism for immune cells and its
immunosuppressive properties. On the other hand, pathological studies have shown that MV also infects
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
epithelial cells at a later stage of infection, although its mechanism has so far been unknown. In this study, we
show that wild-type MV can infect and produce syncytia in human polarized epithelial cell lines independently
of SLAM and CD46 (a receptor for the vaccine strains of MV). Progeny viral particles are released exclusively
from the apical surface of these polarized epithelial cell lines. We have also identified amino acid residues on
the MV attachment protein that are likely to interact with a putative receptor on epithelial cells. All of these
residues have aromatic side chains and may form a receptor-binding pocket located in a different position from
the putative SLAM- and CD46-binding sites on the MV attachment protein. Thus, our results indicate that MV
has an intrinsic ability to infect both polarized epithelial and immune cells by using distinctive receptor-
binding sites on the attachment protein corresponding to each of their respective receptors. The ability of MV
to infect polarized epithelial cells and its exclusive release from the apical surface may facilitate its efficient
transmission via aerosol droplets, resulting in its highly contagious nature.
Measles remains a major cause of childhood morbidity and cytia are also detected in epithelial tissues in various organs,
mortality worldwide despite the availability of efficacious vac- such as the skin, oral mucosa, pharynx, trachea, esophagus,
cines. Measles virus (MV), an enveloped RNA virus belonging intestines, and urinary bladder (5, 24, 26, 27, 31, 33, 34, 37).
to the genus Morbillivirus in the family Paramyxoviridae, is Epithelial cells do not express SLAM, and wild-type (WT)
transmitted via aerosol droplets and considered to be one of strains of MV, unlike vaccine strains, do not infect epithelial
the most contagious human pathogens. MV has two envelope cell lines. Thus, the mechanism by which MV infects epithelial
glycoproteins, the hemagglutinin (H) and fusion (F) proteins, tissues remains to be determined. Recently, we have reported
which are responsible for receptor binding and membrane that a SLAM⫺ human lung adenocarcinoma cell line, NCI-
fusion, respectively (18). MV enters a cell by membrane fusion H358, supports MV entry, replication, and syncytium forma-
at the cell surface. The attachment of the H protein to a tion independently of SLAM and CD46 (43). Furthermore,
cellular receptor is believed to induce the conformational analyses using several monoclonal antibodies (MAbs) against
change of the H protein, as well as that of the F protein, which the H protein indicated that the receptor-binding site on the H
promotes the fusion of the viral envelope with the host cell protein used to infect NCI-H358 cells is different from those
membrane. MV also causes cell-cell fusion in susceptible cells. used for SLAM and CD46 (43).
The signaling lymphocyte activation molecule (SLAM), also In this study, we show that besides NCI-H358, four human
known as CD150, has been identified as a receptor for MV (10, polarized epithelial cell lines can support WT MV growth and
21, 47). SLAM is expressed on immune cells, such as activated syncytium formation via a SLAM- and CD46-independent
lymphocytes, mature dendritic cells, and macrophages, provid- mechanism.
ing a good explanation for the lymphotropism and immuno- MV is found to be released exclusively from the apical sur-
suppressive nature of MV (4, 51). Although CD46, a ubiqui- face of these polarized cell lines. We have also identified amino
tously expressed complement regulatory molecule, functions as acid residues on the H protein that are likely to interact with a
a receptor for the vaccine strains of MV (8, 30), a great ma- putative receptor on epithelial cells mediating SLAM- and
jority of viruses circulating in measles patients use SLAM, but CD46-independent infection. These residues have aromatic
not CD46, as a receptor (35, 51). A recent study of MV infec- side chains and may form a receptor-binding pocket strategi-
tion in macaque monkeys also identified SLAM⫹ lymphocytes cally located on the H protein. These observations provide new
and dendritic cells as the predominantly infected cell types (6). insight into MV infection of epithelial cells as well as into
However, pathological data from humans and experimen- measles pathogenesis.
tally infected monkeys have shown that MV antigens and syn-
MATERIALS AND METHODS
* Corresponding author. Mailing address: Department of Virology, Cells. The characteristics and culture conditions for the following cell lines
Faculty of Medicine, Kyushu University, 3-1-1 Maidashi, Higashi-ku, used were as described previously: Vero/hSLAM (35), B95a (25), NCI-H157
Fukuoka 812-8582, Japan. Phone: 81-92-642-6138. Fax: 81-92-642- (39), NCI-H1299 (15), A549 (16), NCI-H460 (3), NCI-H358 (3), T84 (28),
6140. E-mail: mtakeda@virology.med.kyushu-u.ac.jp. Calu-3 (12), HT29 (13), Caco-2 (12), PC-3 (23), MDCK (14), LLC-PK1 (22), and
䌤
Published ahead of print on 20 February 2008. Vero C1008 (9).
4630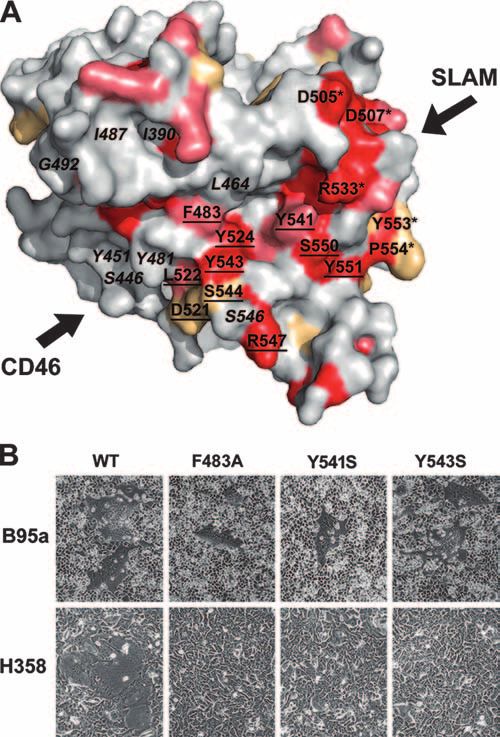
VOL. 82, 2008 MV INFECTION OF POLARIZED EPITHELIAL CELLS 4631
Viruses. All full-length genome plasmids were derived from the p(⫹)MV323
plasmid encoding the antigenomic full-length cDNA of the IC-B WT strain of
MV (44). The p(⫹)MV323-EGFP and p(⫹)MV323-Luci plasmids, which have
an additional transcriptional unit of the enhanced green fluorescent protein
(EGFP) and the Renilla luciferase genes, respectively, were reported previously
(20, 43). Recombinant MV strains were generated from the full-length genome
plasmids as reported previously (29, 41).
Plasmid constructions. The eukaryotic expression vector pCA7 is a derivative
of pCAGGS (32) and has a multiple-cloning site (MCS) for Acc65I, KpnI, SacI,
EcoRI, BsaBI, EcoRV, NotI, XhoI, SphI, and NsiI located downstream from two
promoters, the CAG and T7 promoters (41, 42). The pCA7ps vector is generated
by replacing the MCS of the pCA7 vector with one containing PacI, BstEII,
BsmBI, BmtI, NheI, BglI, PmlI, FseI, NaeI, RsrII, and SpeI (the vector contains
another SpeI site outside the MCS). The pCA7ps-ICH plasmid was generated by
inserting the PacI-SpeI fragment (nucleotide positions 7238 to 9175) of the
full-length genome plasmids derived from p(⫹)MV323 (44) (nucleotide posi-
tions are shown in accordance with the sequence of the IC-B strain genome [45])
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
into the PacI and SpeI sites of the pCA7ps vector. Amino acid substitutions
(N481A, N481R, N481E, N481F, F483A, D521S, L522A, Y524S, Y541S, Y543S,
S544A, R547S, S550A, and Y551S) were introduced into p(⫹)MV323-EGFP,
p(⫹)MV323-Luci, or pCA7ps-ICH by site-directed mutagenesis using the com-
plementary primer pairs. pCA7-ICH-N481Y, pCXN2-KAH-N481F, pCXN2-
KAH-N481S, pCXN2-KAH-N481T, and pCXN2-EdF have been described pre-
viously (38, 50).
Measurement of Renilla luciferase activity. Cells infected with Renilla lucif-
erase-expressing MVs were lysed in Renilla luciferase assay lysis buffer. The
Renilla luciferase activity in the cells was then analyzed by a Renilla luciferase
assay system (Promega, Madison, WI), according to the manufacturer’s instruc-
tion. Chemiluminescence was measured using a Mithras LB940 plate reader
(Berthold Technologies, Pforzheim, Germany).
Cell-cell fusion assay. B95a or NCI-H358 cells cultured in six-well-cluster
plates were cotransfected with each of the H-protein-expressing plasmids (1 g)
and pCXN2-EdF (1 g), using Lipofectamine 2000 (Invitrogen Life Technolo-
gies, Carlsbad, CA). At 2 or 3 days posttransfection, the cells were observed
under a phase-contrast imaging microscope.
Virus titration. The titers of EGFP-expressing viruses were determined as
described previously (20, 42) and expressed in cell infectious units (CIU). The
multiplicity of infection (MOI) was calculated in accordance with the number of
CIU determined on Vero/hSLAM cells. The titers of luciferase-expressing vi-
ruses were determined by plaque titration.
Virus growth. Various cell lines cultured in 96-well-cluster plates were infected
with luciferase-expressing MV strains at an MOI of 1. At various time intervals,
the cells were harvested, and the Renilla luciferase activities were analyzed.
Caco-2, T84, and HT29 cells were seeded at confluence on 24-mm-diameter FIG. 1. WT MV spreads in human polarized epithelial cells.
Transwell filter supports with 0.4-m pores (Corning Inc., Corning, NY) and (A) Subconfluent monolayers of various cell lines were infected with
cultured for 8 to 12 days to produce polarized monolayers. EGFP-expressing MV IC323-EGFP at an MOI of 1. At the indicated days p.i., the distribu-
strains were inoculated into either the apical or basolateral medium and incu- tion of ZO-1 in monolayers was examined using a MAb specific to
bated for 2 h at 37°C. The cells were then washed with phosphate-buffered saline ZO-1, followed by incubation with Alexa Fluor 549-conjugated anti-
(PBS) from both sides and incubated in culture medium at 37°C. At various days mouse secondary antibody. The MV-infected cells were visualized with
postinfection (p.i.), both apical and basolateral media were collected to deter- EGFP autofluorescence. (B) Subconfluent monolayers of various cell
mine the numbers of CIU in them. lines were infected with IC323-Luci at an MOI of 1. The Renilla
Immunofluorescence staining and confocal microscopy. The cells were cul- luciferase activities in cells were determined at the various time points
tured on collagen-coated coverslips. After MV infection, cells were fixed and indicated.
permeabilized with PBS containing 2.5% formaldehyde and 0.5% Triton X-100.
The cells were then washed with PBS and incubated with the mouse MAb against
ZO-1 (Invitrogen) for 1 h at 37°C, followed by incubation with Alexa Fluor
594-conjugated secondary antibody (Molecular Probes, Eugene, OR). Thirty not shown). This prompted us to examine the susceptibility of
fluorescence images of the cells were obtained sequentially from the top to the
bottom of the cells by using a confocal microscope (Radiance 2100; Bio-Rad,
several polarized epithelial cell lines to WT MV. Caco-2,
Hercules, CA) and merged by using Lasersharp software (Bio-Rad). HT29, T84, Calu-3, MDCK, LLC-PK1, and Vero C1008 cells
are known to be “polarized cells,” which form tight junctions
between adjacent cells (1, 7, 9, 17, 36, 40, 49). These cell lines
RESULTS
showed a circumferential immunofluorescent staining pattern
Infection of epithelial cells with WT MV. In an attempt to of ZO-1, a major component of the tight junction (40), indi-
understand why WT MV can grow in the NCI-H358 cell line, cating one of their properties as polarized cells (Fig. 1A).
microarray analysis was performed to compare its gene expres- NCI-H358 cells also exhibited an incompletely circumferential
sion pattern with the patterns of other nonsusceptible human staining pattern of ZO-1, suggesting that they may have an
lung carcinoma cell lines. The results showed that the NCI- ability to form tight junctions. By contrast, scattered dots or
H358 cells express several genes encoding molecules involved short fragments of ZO-1 were observed on the surfaces or at
in tight-junction formation or cell adhesion at significantly the margins of nonpolarized cell lines (HeLa, HEK293, 293T,
higher levels than those of the nonsusceptible cell lines (data NCI-H460, NCI-H1299, NCI-H322, A549, NCI-H157, and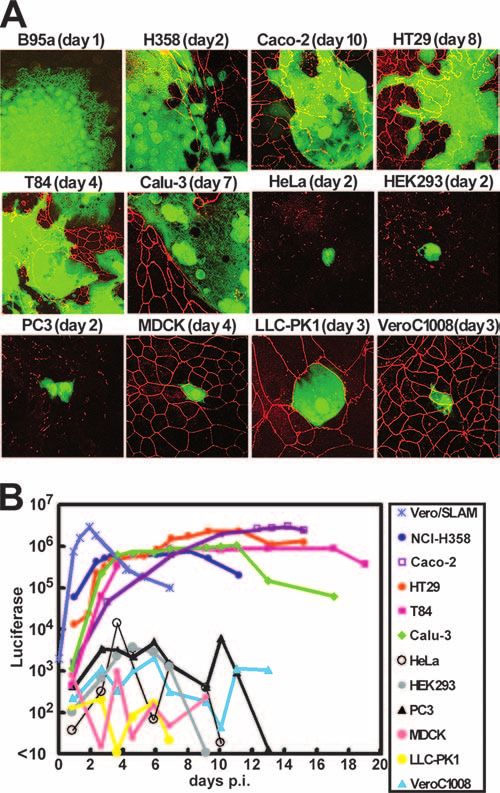
4632 TAHARA ET AL. J. VIROL.
FIG. 2. WT MV buds from the apical surface of polarized epithelial cells. Caco-2 (A) and HT29 (B) cells were cultured at confluence on
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
24-mm-diameter Transwell filter supports with 0.4-m pores. At 8 to 12 days after plating, polarized monolayers of cells were infected with
IC323-EGFP from the apical side. At various time intervals, the numbers of CIU/ml of both the apical and basolateral media were measured (filled
and open circles, respectively). Average numbers of CIU/ml in duplicate experiments are shown.
PC3) (Fig. 1A and data not shown). No signal for ZO-1 was fresh medium at 37°C. At various days p.i., the amounts of the
observed on B-lymphoblastoid B95a cells, which are suscepti- virus in the apical and basolateral media were determined (Fig.
ble to all MV strains. The subconfluent monolayers of these 2A). At 4 days p.i., more than 1 ⫻ 102 CIU/ml of infectious
cell lines were infected with IC323-EGFP, a recombinant WT virus was detected in the apical medium. At 18 days p.i., the vi-
MV strain (based on the IC-B WT strain of MV) expressing rus titer in the apical medium reached 6 ⫻ 104 CIU/ml. The
EGFP (20, 44), and observed daily. The MV-infected cells virus titer in the basolateral medium increased only after 24
were visualized with EGFP autofluorescence. As shown previ- days p.i., when tight junctions were disrupted by an MV-in-
ously (20, 43), IC323-EGFP grew and produced syncytia in duced cytopathic effect. Similar data were obtained with the
B95a (using SLAM as a receptor) and NCI-H358 (using an HT29 (Fig. 2B) and T84 (data not shown) cells. The monolayers
unknown receptor) cells (Fig. 1A). The virus also grew and of the Caco-2 cells were also infected with IC323-EGFP from
produced syncytia in all of the human polarized cell lines the basolateral side. The virus entered the cells much less
examined (Caco-2, HT29, T84, and Calu-3), disrupting tight efficiently than it did from the apical side, and infectious virus
junctions. By contrast, the polarized nonhuman cell lines (dog particles were again released mostly from the apical side (data
MDCK, porcine LLC-PK1, and monkey Vero C1008) and the not shown). Analyses by confocal microscopy showed that H,
nonpolarized human cell lines only rarely supported IC323- F, and matrix proteins were predominantly transported to the
EGFP infection, and no syncytia were detected in them even apical surface of the WT-MV-infected cells (data not shown).
when the cells were observed for 2 weeks (Fig. 1A and data not All these data indicate that WT MV buds almost exclusively
shown). from the apical surface of polarized epithelial cells.
Subconfluent monolayers of these cell lines were also in- Identification of residues on the H protein critical for fusion
fected with IC323-Luci, a recombinant WT MV strain express- of NCI-H358 cells induced by MV H and F proteins. Amino
ing the Renilla luciferase (43). At various time intervals, the acid residues on the H protein that are important in supporting
Renilla luciferase activity in cells was quantified (Fig. 1B). In cell-cell fusion in epithelial cells induced by MV glycoproteins
Vero/hSLAM cells (Vero cells constitutively expressing human were examined. Our previous study suggested that WT MV
SLAM), IC323-Luci replicated efficiently, showing the maxi- uses a novel receptor-binding site on the H protein which is
mum luciferase activity at 45 h p.i. Although slower than that different from those for SLAM and CD46 to infect epithelial
in the Vero/hSLAM cells, the luciferase activities increased in cells (43). That receptor-binding site is, however, likely to
all four of the human polarized cell lines (Caco-2, HT29, T84, overlap that for CD46, as the N481Y substitution, which en-
and Calu-3) and in the NCI-H358 cells after infection with ables WT MV to use CD46 as a receptor, also enhances the
IC323-Luci. The maximum luciferase activities in the Caco-2 ability of WT MV to grow and induce syncytia in the NCI-
and HT29 cells were as high as that in the Vero/hSLAM cells. H358 epithelial cells via a CD46-independent mechanism (43).
By contrast, the luciferase activity did not increase significantly Asn at position 481 of the H protein (of the IC-B WT strain)
in the nonhuman polarized or human nonpolarized cell lines. was replaced with Phe, Ser, Thr, Ala, Arg, Glu, or Tyr, and the
These data indicate that WT MV has an ability to grow and mutated H proteins were transiently expressed, together with
form syncytia in human polarized epithelial cells as well as in the F protein, in B95a and NCI-H358 cells. None of the sub-
NCI-H358 cells. stitutions affected the cell-cell fusion of B95a cells (Fig. 3 and
Exclusive budding of WT MV at the apical surface of polar- data not shown), consistent with the observation that the res-
ized cells. The confluent monolayers of the Caco-2 cells form- idue at position 481 is not involved in the H protein interaction
ing tight junctions were prepared on the filters of Transwell with SLAM (48). Similarly, none of the substitutions inhibited
permeable supports, and 1 ⫻ 104 CIU of IC323-EGFP was the cell-cell fusion of NCI-H358 cells, indicating that the res-
inoculated into the apical medium. After 2 h of incubation with idue at position 481 is not critical for the H protein to support
the virus, the cells were washed with PBS and then cultured in cell-cell fusion of the cells (Fig. 3 and data not shown). How-VOL. 82, 2008 MV INFECTION OF POLARIZED EPITHELIAL CELLS 4633
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
FIG. 3. Role of residues at position 481 of the H protein in cell-cell
fusion of NCI-H358 cells. NCI-H358 or B95a cells cultured in six-well-
cluster plates were transfected with the WT or mutant-H-protein-
expressing plasmid (1 g) together with the F-protein-expressing plas-
mid (1 g). –, no H protein plasmid. At 2 (B95a) or 3 (NCI-H358) days
posttransfection, the cells were observed under a phase-contrast im-
aging microscope.
FIG. 4. Identification of critical amino acid residues on the H pro-
tein to support cell-cell fusion of NCI-H358 cells. (A) The conserved
residues on H proteins of seven morbilliviruses (measles, rinderpest,
ever, the H protein with the N481F substitution, which cannot peste-des-petits-ruminants, canine distemper, dolphin distemper, por-
support CD46-dependent cell-cell fusion (50), enhanced the poise distemper, and phocine distemper viruses). The H protein mono-
cell-cell fusion of the NCI-H358 cells as efficiently as that with mer is illustrated as observed almost from the top. Red, identical;
the N481Y substitution (Fig. 3). These results suggest that salmon pink, strong similarity; wheat yellow, weak similarity; gray, little
similarity. The residues at the putative SLAM-binding sites are shown
aromatic residues (Tyr and Phe) at position 481 positively with asterisks, and those at the putative CD46-binding sites are de-
modulate the interaction of the H protein with the putative noted in italics. The underlined residues, which are conserved among
receptor on NCI-H358 cells. morbilliviruses and reside outside the putative SLAM- and CD46-
Recently, we have determined the crystal structure of the binding sites, were studied by mutagenesis. (B) NCI-H358 and B95a
cells cultured in six-well-cluster plates were transfected with the WT or
MV H protein, which allowed us to precisely locate the amino mutant-H-protein-expressing plasmid (1 g) together with the F-pro-
acid residues presumed to be interacting with SLAM and tein-expressing plasmid (1 g). At 2 (B95a) or 3 (NCI-H358) days
CD46 (19). The structure also revealed that there are many posttransfection, cells were observed under a phase-contrast imaging
residues on the surface of the molecule which are well con- microscope.
served among different morbilliviruses and reside outside the
putative SLAM- and CD46-binding sites (Fig. 4A). We spec-
ulated that these conserved residues may be involved in Y551S substitution inhibited the cell-cell fusion of B95a cells
SLAM- and CD46-independent infection of epithelial cells. To but not that of NCI-H358 cells (data not shown). Importantly,
test this possibility, the H proteins with substitutions at these the F483A, Y541S, and Y543S substitutions caused the H
residues (Phe483, Asp521, Leu522, Tyr524, Tyr541, Tyr543, protein to lose the ability to support syncytium formation in
Ser544, Arg547, Ser550, and Tyr551) were examined for their NCI-H358 cells but not in B95a cells (Fig. 4B).
ability to support the cell-cell fusion of NCI-H358 cells induced Infection of human epithelial cells with recombinant MV
by H and F proteins. The L522A and Y524S substitutions strains bearing the H protein with specific substitutions. Sub-
caused the H protein to lose the ability to support cell-cell stitutions in the H protein that enhanced (N481Y and N481F)
fusion in both B95a and NCI-H358 cells, whereas the D521S, or suppressed (F483A and Y543S) syncytium formation in
S544A, R547S, and S550A substitutions exhibited little effect NCI-H358 cells were introduced into the genomes of infec-
on the cell-cell fusion of either cell line (data not shown). The tious MVs by reverse genetics techniques (29, 41, 44). The4634 TAHARA ET AL. J. VIROL.
tent with the findings obtained with plasmid-mediated fusion
analysis (Fig. 3). These mutant viruses also induced larger
syncytia in the four human polarized epithelial cell lines,
HT29, Calu-3, Caco-2, and T84, than the parental virus did
(Fig. 5A). On the other hand, the mutant viruses with the
F483A or Y543S substitution neither grew well nor induced
syncytia in NCI-H358 cells (Fig. 5A). The strains with the
F483A or Y543S substitution did not induce syncytia in HT29
and Calu-3 cells, but they did produce syncytia in Caco-2 and
T84 cells (Fig. 5A).
The growth of the mutant viruses possessing the F483A or
Y543S substitution in the H protein was also examined by
using recombinant MVs expressing Renilla luciferase (Fig. 5B).
These mutant viruses grew in B95a cells as efficiently as the
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
virus carrying the WT H protein. The virus possessing the
Y543S substitution grew very poorly in NCI-H358, HT29, and
Calu-3 cells but replicated well in Caco-2 and T84 cells. The
replication of the virus with the F483A substitution was also
greatly reduced in NCI-H358, HT29, Calu-3, and Caco-2 cells
but less so in T84 cells. These data suggest that although the
same residues on the H protein may play an important role in
WT MV infection of epithelial cells, the virus-receptor inter-
actions are somehow different among different epithelial cell
lines.
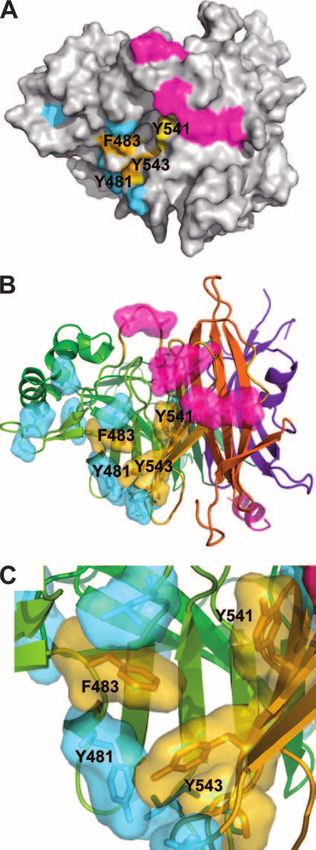
Taken together, our results indicate that Phe483, Tyr541,
and Tyr543 on the H protein are important for WT MV to
infect NCI-H358 and polarized epithelial cell lines. The resi-
due at position 481 is not critical, but the presence of aromatic
residues at that position greatly enhances WT MV infection of
epithelial cells. These four positions are indicated on the three-
dimensional structure of the H protein, together with putative
SLAM- and CD46-binding sites (19) (Fig. 6A and B). (The
structure shown in Fig. 6 is based on the H protein of the
Edmonston strain, but all indicated residues except that at
position 481 are conserved between the Edmonston and WT
IC-B strains.) The aromatic side chains of these residues may
form a receptor-binding pocket interacting with a putative
receptor allowing MV infection of epithelial cells (Fig. 6C).
FIG. 5. Infection of human epithelial cells with recombinant MV
strains bearing the H protein with specific substitutions. (A) EGFP DISCUSSION
autofluorescence in MV-infected cells. B95a, H358, HT29, Calu-3,
Caco-2, and T84 cells were infected with recombinant MVs at an MOI CD46 is the first identified MV receptor (8, 30). The ubiq-
of 0.5. The panels show representative images with a fluorescence uitous expression of CD46 had been thought to explain the
microscope at 2 days (B95a and H358 cells), 6 days (HT29, Calu-3, and findings that MV can infect various types of cells in vivo and in
Caco-2 cells), or 14 days (T84 cells) p.i. (B) The monolayers of B95a,
H358, HT29, Calu-3, Caco-2, and T84 cells were infected with recom-
vitro. Then, SLAM was identified as a receptor for both the
binant MVs at an MOI of 1.0. At 2 (for B95a) or 3 (for other cell lines) WT and vaccine strains of MV, and the use of CD46 was shown
days p.i., the Renilla luciferase activities in the cells were determined. to be limited mostly to the vaccine strains of MV (47), making
The bar graph shows the percent Renilla luciferase activities (average us rethink measles pathogenesis. From the known distribution
and standard deviation) of the cells infected with recombinant MVs of SLAM in the body, the first targets for MV are likely to be
bearing the WT H protein (dark gray) or mutant H protein with the
Y543S (white) or F483A (light gray) substitution. The Renilla lucifer- dendritic cells, lymphocytes, and/or macrophages in the lym-
ase activities of the respective cell lines infected with IC323-Luci bear- phoid tissues of the respiratory tract (51). After the initial
ing the WT H protein were set to 100%. infection of those cells, the virus enters the blood and repli-
cates in lymphoid tissues and organs throughout the body,
causing profound immunosuppression. At a later stage of in-
recombinant MV with the N481Y substitution in the H protein fection, infected dendritic cells may transmit MV to epithelial
was reported previously (38). All of the recombinant viruses cells (6). Previous studies using a panel of cell lines showed
with the mutated H proteins efficiently replicated and pro- that only SLAM⫹ cells support efficient WT MV infection and
duced syncytia in B95a cells (Fig. 5A). The viruses with the syncytium formation (11). Epithelial cells do not express
N481Y or N481F substitution induced much larger syncytia in SLAM, and WT MV uses SLAM but not CD46 as a receptor.
NCI-H358 cells than the parental virus did (Fig. 5A), consis- How then does MV infect epithelial cells in vivo? AlthoughVOL. 82, 2008 MV INFECTION OF POLARIZED EPITHELIAL CELLS 4635
previous studies have shown that a low level of SLAM-inde-
pendent WT MV infection occurs in various cell lines (the
efficiency is 100 to 1,000 times lower than that using SLAM),
this type of infection does not produce syncytia in infected cells
(20), except in primary cultures of human small-airway epithe-
lial cells and endothelial cells (2, 46).
Here and in our previous paper (43), we have demonstrated
that WT MV has the ability to infect and produce syncytia in
certain epithelial cell lines. The infection is independent of
SLAM and CD46. Furthermore, we have identified residues on
the attachment H protein that presumably interact with the
putative epithelial cell receptor. Those residues are in a loca-
tion different from putative SLAM- and CD46-binding sites on
the H protein. Thus, our results indicate that MV has an
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
intrinsic ability to infect both polarized epithelial and immune
cells using distinctive receptor-binding sites on the H protein
corresponding to respective receptors.
All of the identified amino acid residues relevant for infec-
tion of epithelial cells are those with aromatic side chains
(Phe483, Try541, and Try543), suggesting that hydrophobic
interactions play an important role in the binding of the H
protein to the putative receptor on epithelial cells. This is also
supported by the finding that the presence of aromatic residues
at position 481, which is located close to the aforementioned
three positions, enhances cell-cell fusion in epithelial cells after
expression of the H and F proteins or recombinant virus in-
fection (Fig. 3 and 5). These results suggest that there exists a
cluster of hydrophobic residues on the putative epithelial cell
receptor, which interacts with the aforementioned aromatic
residues on the H protein. We have previously shown that
residues at the SLAM-binding site on the H protein are highly
conserved among different morbilliviruses (19). Tyr541 and
Tyr543 are also highly conserved among morbilliviruses.
Phe483 is found only in MV, but all other morbilliviruses have
a hydrophobic Leu residue at that position. Thus, it is likely
that morbilliviruses other than MV also use the same molecule
as a receptor to infect epithelial cells. These relevant aromatic
residues are located upward from the viral envelope (as por-
trayed in Fig. 6), because of the tilted orientation of the mol-
ecules forming the H protein dimer (19). Thus, they should be
able to interact readily with the putative receptor on epithelial
cells.
Why does MV infect epithelial cells? Polarized epithelial
cells form tight junctions covering the external epithelial sur-
face. The tight junctions prevent the passage of most dissolved
molecules from one side of the epithelium to the other. Thus,
it is possible that MV cannot efficiently release progeny virions
into the external surface through its ability to infect immune
cells alone. At present, the identity of the receptor which MV
uses to infect epithelial cells is unknown, but our results
FIG. 6. Cluster of aromatic residues at the putative receptor-bind- showed that progeny virions are selectively released into the
ing site of the H protein. (A) Surface presentation of the H protein
seen from the top. The amino acid residues assumed to interact with
apical side of polarized epithelial cells (the luminal side). This
SLAM and CD46 are shown in magenta and cyan, respectively. Res- suggests that the ability to infect epithelial cells is necessary for
idues important for infection of epithelial cells (F483, Y541 and Y543) MV to spread efficiently from person to person, rather than to
are shown in orange. (B) Ribbon and stick model at the same angle as spread only within an individual host. This may explain why
that used in panel A. The head of the H protein exhibits a six-bladed
human immunodeficiency virus, which shares a similar tropism
-propeller fold. The colors blue, green, light green, yellow, orange,
and red represent each propeller from -sheets 1, 2, 3, 4, 5, and 6, for immune cells with MV, is transmitted exclusively via sexual
respectively. (C) Magnification of the putative receptor-binding site on contact or blood, whereas MV is transmitted efficiently via
the H protein used to infect epithelial cells. aerosol droplets.4636 TAHARA ET AL. J. VIROL.
ACKNOWLEDGMENTS (SLAM) is a receptor for a lymphotropic strain of measles virus and may
account for the immunosuppressive properties of this virus. Virology 279:
We thank M. Sato, T. Yano, Y. Fujinaga, Y. Ito, S. Naito, and T. 9–21.
Harada for providing cell lines; T. Seya and T. A. Sato for antibodies; 22. Hull, R. N., W. R. Cherry, and G. W. Weaver. 1976. The origin and charac-
and K. Maenaka for helpful discussions. We also thank C. Tsuda teristics of a pig kidney cell strain, LLC-PK. In Vitro 12:670–677.
(Tomy Digital Biology Co., Ltd.) for the analysis of microarray data. 23. Kaighn, M. E., K. S. Narayan, Y. Ohnuki, J. F. Lechner, and L. W. Jones.
This work was supported by grants from the Ministry of Education, 1979. Establishment and characterization of a human prostatic carcinoma
cell line (PC-3). Invest. Urol. 17:16–23.
Culture, Sports, Science and Technology and the Ministry of Health,
24. Kimura, A., K. Tosaka, and T. Nakao. 1975. Measles rash. I. Light and
Labor and Welfare of Japan. M. Tahara is supported by research electron microscopic study of skin eruptions. Arch. Virol. 47:295–307.
fellowships from the Japan Society for the Promotion of Science for 25. Kobune, F., H. Sakata, and A. Sugiura. 1990. Marmoset lymphoblastoid cells
Young Scientists. as a sensitive host for isolation of measles virus. J. Virol. 64:700–705.
26. Lightwood, R., and R. Nolan. 1970. Epithelial giant cells in measles as an acid
REFERENCES in diagnosis. J. Pediatr. 77:59–64.
1. Anderson, J. M., C. M. Van Itallie, M. D. Peterson, B. R. Stevenson, E. A. 27. Moench, T. R., D. E. Griffin, C. R. Obriecht, A. J. Vaisberg, and R. T.
Carew, and M. S. Mooseker. 1989. ZO-1 mRNA and protein expression Johnson. 1988. Acute measles in patients with and without neurological
during tight junction assembly in Caco-2 cells. J. Cell Biol. 109:1047–1056. involvement: distribution of measles virus antigen and RNA. J. Infect. Dis.
2. Andres, O., K. Obojes, K. S. Kim, V. ter Meulen, and J. Schneider-Schaulies. 158:433–442.
2003. CD46- and CD150-independent endothelial cell infection with wild- 28. Murakami, H., and H. Masui. 1980. Hormonal control of human colon
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guest
type measles viruses. J. Gen. Virol. 84:1189–1197. carcinoma cell growth in serum-free medium. Proc. Natl. Acad. Sci. USA
3. Brower, M., D. N. Carney, H. K. Oie, A. F. Gazdar, and J. D. Minna. 1986. 77:3464–3468.
Growth of cell lines and clinical specimens of human non-small cell lung 29. Nakatsu, Y., M. Takeda, M. Kidokoro, M. Kohara, and Y. Yanagi. 2006.
cancer in a serum-free defined medium. Cancer Res. 46:798–806. Rescue system for measles virus from cloned cDNA driven by vaccinia virus
4. Cocks, B. G., C.-C. J. Chang, J. M. Carballido, H. Yssel, J. E. de Vries, and Lister vaccine strain. J. Virol. Methods 137:152–155.
G. Aversa. 1995. A novel receptor involved in T-cell activation. Nature 30. Naniche, D., G. Varior-Krishnan, F. Cervoni, T. F. Wild, B. Rossi, C.
376:260–263. Rabourdin-Combe, and D. Gerlier. 1993. Human membrane cofactor protein
5. Craighead, J. E. 2000. Rubeola (measles), p. 397–410. In J. E. Craighead (CD46) acts as a cellular receptor for measles virus. J. Virol. 67:6025–6032.
(ed.), Pathology and pathogenesis of human viral disease. Elsevier, London, 31. Nii, S., J. Kamahora, Y. Mori, M. Takahashi, S. Nishimura, and Y. Okuno.
United Kingdom. 1964. Experimental pathology of measles in monkeys. Biken J. 6:271–297.
6. de Swart, R. L., M. Ludlow, L. de Witte, Y. Yanagi, G. van Amerongen, S. 32. Niwa, H., K. Yamamura, and J. Miyazaki. 1991. Efficient selection for
McQuaid, S. Yuksel, T. B. Geijtenbeek, W. P. Duprex, and A. D. Osterhaus. high-expression transfectants with a novel eukaryotic vector. Gene 108:193–
2007. Predominant infection of CD150(⫹) lymphocytes and dendritic cells 199.
during measles virus infection of macaques. PLoS Pathog. 3:e178. 33. Nommensen, F. E., and N. W. Dekkers. 1981. Detection of measles antigen
7. Dharmsathaphorn, K., J. A. McRoberts, K. G. Mandel, L. D. Tisdale, and H. in conjunctival epithelial lesions staining by lissamine green during measles
Masui. 1984. A human colonic tumor cell line that maintains vectorial virus infection. J. Med. Virol. 7:157–162.
electrolyte transport. Am. J. Physiol. 246:G204–G208. 34. Olding-Stenkvist, E., and B. Bjorvatn. 1976. Rapid detection of measles
8. Dörig, R. E., A. Marcil, A. Chopra, and C. D. Richardson. 1993. The human virus in skin rashes by immunofluorescence. J. Infect. Dis. 134:463–469.
CD46 molecule is a receptor for measles virus (Edmonston strain). Cell 35. Ono, N., H. Tatsuo, Y. Hidaka, T. Aoki, H. Minagawa, and Y. Yanagi. 2001.
75:295–305. Measles viruses on throat swabs from measles patients use signaling lym-
9. Earley, E., and K. Johnson. 1988. The lineage of the Vero, Vero 76 and its phocytic activation molecule (CDw150) but not CD46 as a cellular receptor.
clone C1008 in the United States, p. 26–29. In B. Simizu, and T. Terasima. J. Virol. 75:4399–4401.
(ed.), Vero cells: origin, properties and biomedical applications. Chiba Uni- 36. Polak-Charcon, S., and Y. Ben-Shaul. 1979. Degradation of tight junctions in
versity, Tokyo, Japan. HT29, a human colon adenocarcinoma cell line. J. Cell Sci. 35:393–402.
10. Erlenhoefer, C., W. J. Wurzer, S. Löffler, S. Schneider-Schaulies, V. ter 37. Sakaguchi, M., Y. Yoshikawa, K. Yamanouchi, T. Sata, K. Nagashima, and
Meulen, and J. Schneider-Schaulies. 2001. CD150 (SLAM) is a receptor for K. Takeda. 1986. Growth of measles virus in epithelial and lymphoid tissues
measles virus but is not involved in viral contact-mediated proliferation of cynomolgus monkeys. Microbiol. Immunol. 30:1067–1073.
inhibition. J. Virol. 75:4499–4505. 38. Seki, F., M. Takeda, H. Minagawa, and Y. Yanagi. 2006. Recombinant
11. Erlenhöfer, C., W. Duprex, B. Rima, V. ter Meulen, and J. Schneider- wild-type measles virus containing a single N481Y substitution in its haemag-
Schaulies. 2002. Analysis of receptor (CD46, CD150) usage by measles virus. glutinin cannot use receptor CD46 as efficiently as that having the haemag-
J. Gen. Virol. 83:1431–1436. glutinin of the Edmonston laboratory strain. J. Gen. Virol. 87:1643–1648.
12. Fogh, J., J. M. Fogh, and T. Orfeo. 1977. One hundred and twenty-seven 39. Sherwin, S. A., J. D. Minna, A. F. Gazdar, and G. J. Todaro. 1981. Expres-
cultured human tumor cell lines producing tumors in nude mice. J. Natl. sion of epidermal and nerve growth factor receptors and soft agar growth
Cancer Inst. 59:221–226. factor production by human lung cancer cells. Cancer Res. 41:3538–3542.
13. Fogh, J., and G. Trempe. 1975. New human tumor cell lines, p. 115–160. In 40. Stevenson, B. R., J. M. Anderson, D. A. Goodenough, and M. S. Mooseker.
J. Fogh (ed.), Human tumor cells in vitro. Plenum Publishing Corp., New 1988. Tight junction structure and ZO-1 content are identical in two strains
York, NY. of Madin-Darby canine kidney cells which differ in transepithelial resistance.
14. Gaush, C. R., W. L. Hard, and T. F. Smith. 1966. Characterization of an J. Cell Biol. 107:2401–2408.
established line of canine kidney cells (MDCK). Proc. Soc. Exp. Biol. Med. 41. Takeda, M., S. Ohno, F. Seki, K. Hashimoto, N. Miyajima, K. Takeuchi, and
122:931–935. Y. Yanagi. 2005. Efficient rescue of measles virus from cloned cDNA using
15. Giaccone, G., J. Battey, A. F. Gazdar, H. Oie, M. Draoui, and T. W. Moody. SLAM-expressing Chinese hamster ovary cells. Virus Res. 108:161–165.
1992. Neuromedin B is present in lung cancer cell lines. Cancer Res. 52: 42. Takeda, M., S. Ohno, F. Seki, Y. Nakatsu, M. Tahara, and Y. Yanagi. 2005.
S2732–S2736. Long untranslated regions of the measles virus M and F genes control virus
16. Giard, D. J., S. A. Aaronson, G. J. Todaro, P. Arnstein, J. H. Kersey, H. replication and cytopathogenicity. J. Virol. 79:14346–14354.
Dosik, and W. P. Parks. 1973. In vitro cultivation of human tumors: estab- 43. Takeda, M., M. Tahara, T. Hashiguchi, T. A. Sato, F. Jinnouchi, S. Ueki, S.
lishment of cell lines derived from a series of solid tumors. J. Natl. Cancer Ohno, and Y. Yanagi. 2007. A human lung carcinoma cell line supports
Inst. 51:1417–1423. efficient measles virus growth and syncytium formation via a SLAM- and
17. Gopalakrishnan, S., M. A. Hallett, S. J. Atkinson, and J. A. Marrs. 2003. CD46-independent mechanism. J. Virol. 81:12091–12096.
Differential regulation of junctional complex assembly in renal epithelial cell 44. Takeda, M., K. Takeuchi, N. Miyajima, F. Kobune, Y. Ami, N. Nagata, Y.
lines. Am. J. Physiol. Cell Physiol. 285:C102–C111. Suzaki, Y. Nagai, and M. Tashiro. 2000. Recovery of pathogenic measles
18. Griffin, D. E. 2007. Measles virus, p. 1551–1585. In D. M. Knipe, P. M. virus from cloned cDNA. J. Virol. 74:6643–6647.
Howley, D. E. Griffin, R. A. Lamb, M. A. Martin, B. Roizman, and S. E. 45. Takeuchi, K., N. Miyajima, F. Kobune, and M. Tashiro. 2000. Comparative
Straus (ed.), Fields virology, 5th ed. Lippincott Williams & Wilkins, Phila- nucleotide sequence analysis of the entire genomes of B95a cell-isolated and
delphia, PA. Vero cell-isolated measles viruses from the same patient. Virus Genes 20:
19. Hashiguchi, T., M. Kajikawa, N. Maita, M. Takeda, K. Kuroki, K. Sasaki, D. 253–257.
Kohda, Y. Yanagi, and K. Maenaka. 2007. Crystal structure of measles virus 46. Takeuchi, K., N. Miyajima, N. Nagata, M. Takeda, and M. Tashiro. 2003.
hemagglutinin provides insight into effective vaccines. Proc. Natl. Acad. Sci. Wild-type measles virus induces large syncytium formation in primary hu-
USA 104:19535–19540. man small airway epithelial cells by a SLAM(CD150)-independent mecha-
20. Hashimoto, K., N. Ono, H. Tatsuo, H. Minagawa, M. Takeda, K. Takeuchi, nism. Virus Res. 94:11–16.
and Y. Yanagi. 2002. SLAM (CD150)-independent measles virus entry as 47. Tatsuo, H., N. Ono, K. Tanaka, and Y. Yanagi. 2000. SLAM (CDw150) is a
revealed by recombinant virus expressing green fluorescent protein. J. Virol. cellular receptor for measles virus. Nature 406:893–897.
76:6743–6749. 48. Vongpunsawad, S., N. Oezgun, W. Braun, and R. Cattaneo. 2004. Selectively
21. Hsu, E., C. Iorio, F. Sarangi, A. Khine, and C. Richardson. 2001. CDw150 receptor-blind measles viruses: identification of residues necessary forVOL. 82, 2008 MV INFECTION OF POLARIZED EPITHELIAL CELLS 4637
SLAM- or CD46-induced fusion and their localization on a new hemagglu- 50. Xie, M.-F., K. Tanaka, N. Ono, H. Minagawa, and Y. Yanagi. 1999. Amino
tinin structural model. J. Virol. 78:302–313. acid substitutions at position 481 differently affect the ability of the measles
49. Wan, H., H. L. Winton, C. Soeller, G. A. Stewart, P. J. Thompson, D. C. virus hemagglutinin to induce cell fusion in monkey and marmoset cells
Gruenert, M. B. Cannell, D. R. Garrod, and C. Robinson. 2000. Tight co-expressing the fusion protein. Arch. Virol. 144:1689–1699.
junction properties of the immortalized human bronchial epithelial cell lines 51. Yanagi, Y., M. Takeda, and S. Ohno. 2006. Measles virus: cellular receptors,
Calu-3 and 16HBE14o. Eur. Respir. J. 15:1058–1068. tropism and pathogenesis. J. Gen. Virol. 87:2767–2779.
Downloaded from http://jvi.asm.org/ on March 24, 2021 by guestYou can also read

















































