Morphohistological analysis and histochemistry of Feijoa sellowiana somatic embryogenesis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Protoplasma (2004) 224: 33–40
DOI 10.1007/s00709-004-0055-5 PROTOPLASMA
Printed in Austria
Morphohistological analysis and histochemistry of Feijoa sellowiana
somatic embryogenesis
G. C. Cangahuala-Inocente, N. Steiner, M. Santos, and M. P. Guerra*
Grupo de Pesquisas em Recursos Geneticos Vegetais, Departamento de Fitotecnia, Centro de Ciências Agrárias, Universidade Federal de Santa
Catarina, Florianópolis, Santa Catarina
Received February 11, 2004; accepted March 5, 2004; published online October 4, 2004
© Springer-Verlag 2004
Summary. Morphohistological analysis and histochemical studies were normally associated with reentry into the mitotic cycle as
carried out during the induction and development of Feijoa sellowiana well as with alterations in the cell division planes. In some
somatic embryos. Zygotic embryos were cultured on LPm medium con-
taining 2,4-dichlorophenoxyacetic acid (20 M) and glutamine (8 mM). cases, however, in vitro morphogenic competence is not
Somatic embryogenesis could be induced from embryogenic cells that directly associated with the level of mitotic activity
originated in meristematic centers or from clusters of cells. The presence (Dolezelova et al. 1992).
of few starch grains and abundant protein bodies was observed in the
globular and early torpedo stages, while in torpedo and cotyledonary-
Somatic embryogenesis is a process through which
stage somatic embryos an enhanced synthesis of starch grains was associ- bipolar embryos develop from a nonzygotic cell without
ated with the accumulation of reserves to be used in the conversion of the vascular connections with the original tissue. Somatic em-
embryos to plantlets. Proteins were predominantly observed in protoderm
bryogenesis is a multistep regeneration process starting
cells, as well as in the meristematic apical region of torpedo and cotyle-
donary-stage somatic embryos. with the formation of proembryogenic cell masses, fol-
lowed by somatic-embryo formation, maturation, desicca-
Keywords: Pineapple guava; Acca sellowiana; Feijoa sellowiana; His- tion, and plant regeneration (von Arnold et al. 2002).
tological analysis; Somatic embryogenesis; Protein body; Starch grain.
Somatic embryos can differentiate either directly or indi-
Abbreviations: 2,4-D 2,4-dichlorophenoxyacetic acid; PAS periodic rectly from the explant (Williams and Maheswaran 1986).
acid-Schiff reaction; TBO toluidine blue O. Indirect somatic embryogenesis arises from undetermined
cells following the formation of a nondifferentiated callus.
Introduction Distinguishing between direct and indirect somatic em-
bryogenesis is, however, a difficult task. In conifers, em-
In plant cell tissue culture, competent cells are recognized bryogenic calluses consist of proembryogenic masses (von
by their responses to external signals that activate specific Arnold et al. 2002), which contradicts the criterion of uni-
developmental pathways (McDaniel 1984). This is demon- cellular origin. This type of indirect embryogenesis is,
strated when an isolated explant that is not intrinsically re- however, rarely found in angiosperms (Haccius 1978).
sponsive acquires this competence when it is activated by The histological alterations associated with the position
an inductive signal (Finstad et al. 1993). This suggests that and activity of competent cells during the acquisition of so-
cell competence may be acquired through a dedifferentia- matic embryogenic competence has been the subject of sev-
tion process (Torrey 1977). Regenerative competence is eral studies. For example, in hybrid Rosa species, cells on
the periphery of the callus have been observed to undergo
internal segmenting divisions and either form somatic em-
* Correspondence and reprints: Departamento de Fitotecnia, Centro de bryos directly or continue to proliferate forming embryo-
Ciências Agrárias, Universidade Federal de Santa Catarina, C.P. 476,
88034-001 Florianópolis, SC, Brazil. genic calluses (Rout et al. 1998). In Feijoa sellowiana, the
E-mail: mpguerra@cca.ufsc.br formation of a dense layer of meristematic cells originating34 G. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana
in the adaxial face of the cotyledons of zygotic embryos has Material and methods
been described. Two patterns of somatic-embryo differentia-
Plant material
tion were observed: one from single epidermal cells and the
Ripe fruits of Feijoa sellowiana (O. Berg) O. Berg genotype 101 were ob-
other from groups of meristematic cells located near the
tained from the germplasm collection of the São Joaquim Experimental
adaxial surface (Canhoto and Cruz 1996). Station (EPAGRI), Santa Catarina, southern Brazil, and seeds were surface
The plant growth regulators used for embryogenic induction sterilized according to Guerra et al. (2001). The zygotic embryos were ex-
produce alterations in cell polarity and promote subsequent cised in an aseptic chamber and inoculated into test tubes (25 150 mm)
containing 15 ml of induction medium, consisting of basal medium LPm
asymmetric divisions (Ammirato 1983). Carya illinoinensis (von Arnold and Eriksson 1981) supplemented with Morel vitamins (Morel
cultures induced by naphthaleneacetic acid have been reported and Wetmore 1951), glutamine (8 mM), 2,4-D (20 M), sucrose (3%), and
to show embryogenic regions composed of homogeneous, iso- agar-agar (0.7%). The pH was adjusted to 5.8 prior to autoclaving. The cul-
tures were maintained in the dark at 25 C during the induction phase.
diametric, meristematic cells, and the somatic embryos derived
from these cultures generally had a normal morphology. In
contrast, somatic embryos induced in culture media containing Microscopic preparation
2,4-dichlorophenoxyacetic acid (2,4-D) showed abnormalities Zygotic embryos incubated in the induction medium were removed
every 3 days over the 90-day culture period and fixed for 24 h in 0.2 M
(Rodriguez and Wetzstein 1998).
phosphate buffer (pH 7.3) containing 2.5% paraformaldehyde. After fix-
The aim of the present work is to evaluate the morpho- ation, the samples were dehydrated in a graded ethanol series and em-
histology and the histochemical aspects associated with the bedded in historesin (Leica), as described by Arnold et al. (1975).
induction and development of somatic embryos from com- Sections, 5 m thick, were cut with a rotary microtome (Slee Technik)
and fixed onto slides by heating.
petent explants of Feijoa sellowiana cultured in inductive Samples were dehydrated with periodic acid and stained by the periodic
conditions. acid-Schiff reaction (PAS) to reveal starch grain location. Storage proteins
Fig. 1a–d. Histology of F. sellowiana embryogenic cultures induced by 2,4-D (20 M). a Longitudinal section of zygotic embryo after 15 days in
culture showing large cells in the cotyledonary tissues and small, compact cells in the root tissues. b Cell segregation resulting from proliferative
burst of epidermal cells in zygotic-embryo cotyledon after 18 days in culture. c Induction of meristematic cluster originating from parenchyma cells
of cotyledon after 21 days in culture. d Induction of globular somatic embryos after 60 days in culture. a–c Stained with TBO, d stained with PAS. co
Cotyledon, ra root, me apical meristem, seg cell segregation, me-no meristematic nodule, se somatic embryo, se-glo globular-stage somatic embryo,
st starch. Bars: a, 0.300 mm; b–d, 100 mG. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana 35
were stained with Coomassie brilliant blue R250 (Sigma) (Gahan 1984), After 18 days in culture, a proliferative burst in the
and acid polysaccharides and phenols were stained with 0.5% toluidine
epidermis and the beginning of cellular segregation could
blue O (TBO) (O’Brien et al. 1965). Photographs were taken with a stan-
dard Olympus BX 40 microscope. be seen (Fig.1b). Cells originating from this process
were small and isodiametric with a parietal nucleus and
large vacuole and contained phenolic compounds and
Results
starch grains. After 21 days in culture, meristematic cen-
After 15 days on somatic embryogenesis induction medium, ters showing two distinct regions were observed (Fig.1c).
zygotic embryos showed expanded, green cotyledons. In the One region was centrally located with intense mitotic ac-
longitudinal section of the cotyledon, stained with TBO, tivity and protein synthesis, as indicated by Coomassie
large cells with parietal nuclei and cytoplasm and just one brilliant blue R250 staining (data not shown). The second
large vacuole could be observed. In contrast, the root re- region was characterized by the accumulation of phenolic
gions revealed small cells with a high nucleoplasmic ratio, compounds, as revealed by the green metachromatic re-
dense cytoplasm, and a small or absent vacuole (Fig.1a). action (Fig.1c).
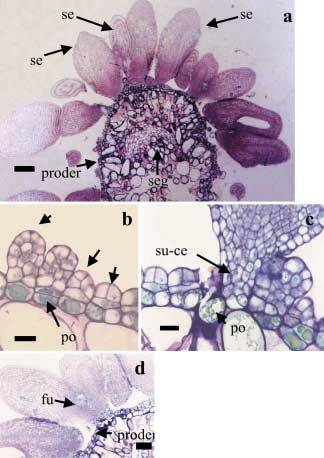
Fig. 2 a–d. Indirect somatic embryogenesis
in F. sellowiana. a Somatic embryos arising
from a layer of embryogenic cells. b Induc-
tion of proembryos. c Group of suspensor
cells. b and c Note the presence of polyphe-
nols (po). d Fusion of somatic embryos. All
sections were stained with TBO. se Somatic
embryo, proder protoderm, seg cell segrega-
tion, su-ce suspensor cells, fu fused somatic
embryos. Bars: a, 150 m; b–d, 50 m36 G. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana
The first visualization of somatic embryogenesis was consequence of this fragmentation, groups of embryonic
possible after 39 days in culture. After 60 days in culture, cells were isolated from the surrounding tissue (Fig. 3a).
histological analysis revealed the development of somatic The cells of somatic embryos in different developmental
proembryos arising from peripheral cells of the meriste- stages showed similar histochemical reactions. However, the
matic centers (Fig.1d). Protein bodies were observed in morphological features were distinct. The cells of globular
the cells of somatic proembryos (data not shown). somatic embryos contained few starch grains and were sur-
An embryogenic cell layer surrounding the meristematic rounded by a layer of protoderm cells. These cells were
centers (Fig. 2a) was competent for somatic-proembryo de- small with high nucleoplasmic ratios and dense cytoplasm
velopment. The first divisions of this cell layer were pericli- (Fig. 3b). In the early torpedo stage, the metachromatic reac-
nal, but subsequent divisions occurred in several planes. The tion of TBO was observed specifically in the basal cells
proembryos developed from clumps of cells (Fig. 2 b, c). (Fig. 3c), similar to the observed pattern in the cells of the
Staining with TBO revealed that the cells of this peripheral peripheral layer surrounding the meristematic centers (see
layer were small and isodiametric, and their vacuoles were Fig.1c). Starch grains were also present in these cells but
filled with polyphenol compounds (Fig. 2 b, c). Once the were absent from the apical region of somatic embryos
meristematic centers acquired embryonic features, fragmen- (Fig. 3d). A positive Schiff reaction also revealed starch
tation of these cellular masses was frequently observed. As a grains in the intracellular domain of basal cells of torpedo
Fig. 3 a–f. Histological sections of F. sell-
owiana somatic embryos. a and b Em-
bryogenic cells forming globular somatic
embryos. Note the presence of polyphenols
(po) and starch granules (st) in the mother
cells. b Globular somatic embryos showing
a well-developed protoderm. c and d Early-
torpedo-stage somatic embryos. e and f
Torpedo stage somatic embryos showing pro-
cambial region. a and c Stained with TBO. b,
d, and e Stained with PAS. f Stained with
Coomassie brilliant blue. proder Protoderm,
procam procambium, pro protein body, seg
cell segregation. Bars: a–e, 50 m; f, 100 mG. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana 37
stage embryos (Fig. 3e). Coomassie brilliant blue staining re- ing protoderm and procambial cells. These somatic em-
vealed protein bodies in all cells at this stage (Fig. 3f). Acid bryos also revealed conspicuous apical and root meristem
polysaccharides could be seen in pre-cotyledonary-stage so- regions. Initially, somatic embryo development was syn-
matic embryos stained with TBO, mainly as constituents chronous (Fig.1d), but continued in an asynchronous
of the cellular wall (Fig. 4a, b). Cotyledonary-stage somatic manner (Fig. 2a). Vascular connections were detected be-
embryos contained protein bodies in the protoderm cells tween the embryos and the peripheral cells (Fig. 2a). Ab-
(Fig. 4d), as well as starch grains in the basal cells (Fig. 4e). normalities were often found in the developing somatic
Somatic embryos in the early torpedo (Fig. 3c), torpedo embryos, such as an altered number of cotyledons and,
(Fig. 3f), pre-cotyledonary (Fig. 4a, b), and cotyledonary most commonly, the presence of fused somatic embryos
stages (Fig. 4c) exhibited differentiated regions contain- (Fig. 2d).
Fig. 4 a–f. Histological sections of F. sello-
wiana somatic embryos. a and b Pre-cotyle-
donary somatic embryos showing protoderm
and procambial strands. c–e Cotyledonary
somatic embryos. f Cell agglomerates with
starch grains. a–c Stained with TBO. d Stained
with Coomassie brilliant blue. e and f Stained
with PAS. proder Protoderm, procam procam-
bium, co cotyledon, po polyphenols, pro pro-
tein bodies, st starch granule. Bars: a–e,
100 m; f, 50 m38 G. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana
Discussion Storage products
Reserve compounds play an important role in in vitro mor-
Development of somatic embryogenesis
phogenesis. For example, high levels of polysaccharides at
In the present work, somatic embryos differentiated de the beginning of the in vitro developmental process have
novo from the segregation of cotyledon cells of zygotic been reported (Branca et al. 1994), and the consumption of
embryos. This process occurred in two steps: first, cellular these compounds has been correlated with the onset of
segregation originating in meristematic centers; second, organogenesis and somatic embryogenesis (Mangat et al.
formation of a peripheral cell layer surrounding the meri- 1990, Martin et al. 2000).
stematic centers, with every cell of this layer showing In Carya illinoinensis, the formation of embryogenic pro-
competence for somatic embryogenesis. tuberances is preceded by the accumulation of starch gran-
The cells resulting from the segregation were isodiamet- ules in the subepidermal cell layers of the explant. Starch is
ric, with a parietal nucleus and a large vacuole, and con- rapidly consumed during the formation of embryogenic re-
tained phenolic compounds and starch grains. It has been gions and is absent from globular and heart-shaped embryos
previously reported that single cells can produce few-celled (Rodriguez and Wetzstein 1998). Our results are in agree-
proembryos, referred to as embryogenic units in Zea mays ment with these findings since the meristematic centers con-
(Fransz and Schel 1991) or proembryonic cell masses in tained abundant starch grains which were heavily depleted
Pennsisetum glaucum (Taylor and Vasil 1996) and Quercus in the proembryonic cell clumps. Starch is considered to be
suber L. (Puigderrajols et al. 2001). the primary source of energy for cellular proliferation and
In the present work we observed the presence of thick cell growth. The consumption of these starch grains, therefore,
walls surrounding the proembryo cells. Small globular clus- should provide energy for the development of the somatic
ters without visible polarity were associated with earlier embryos, suggesting an active regulation of starch accumu-
proembryos, whereas globular clusters in which polarity was lation as has been proposed by Martin et al. (2000). Canhoto
already established were associated with later developmental and Cruz (1996) could not detect starch grains in meriste-
stages. Similar morphogenetic features have been described matic layers of F. sellowiana; although, they were present in
for Guinea grass by Karlsson and Vasil (1986) and for cork proembryos. This suggests that starch is rapidly metabolized
oak by Puigderrajols et al. (2001). in embryogenic tissues, providing energy for the intense
The development of zygotic embryos is well under- metabolic and mitotic activity (Stamp 1987).
stood since they originate from the fusion of two haploid Our histochemical evaluations also revealed that embryo-
cells. The origin of somatic embryos has been associated genic cells resulting from cell segregation contain protein
with two pathways: unicellular or multicellular. Unicellu- bodies, which were also observed in the meristematic cen-
lar somatic embryogenesis results from the development ters. Most seed storage proteins are secretory proteins syn-
of single cells, whereas multicellular somatic embryogen- thesized from a peptide that is cleaved as the protein is
esis results from the association of embryogenic cells or transported into the lumen of the endoplasmic reticulum
evolves from embryogenic cell clusters (Michaux-Ferrière (Shewry et al. 1995). Storage proteins found in vacuoles are
and Schwendiman 1993). spherical protein bodies that are degraded during germina-
Our results suggest that the somatic embryos in this tion to provide carbon and nitrogen for the growing seedling
study had both unicellular and multicellular origins, as has (Shotwell and Larkins 1989). It has been suggested that the
been described for other dicotyledonous species (Colby presence of proteins in the embryogenic cells is associated
et al. 1991). For Panicum maximum, it has been demon- with the formation of proembryonic cell groups. Meriste-
strated that somatic embryos arise from single cells and matic centers formed of isodiametric cells with prominent
closely resemble the developmental morphology of zygotic nucleoli and high mitotic activity have been observed in
embryos of grasses (Botti and Vasil 1984, Lu and Vasil Eucalyptus urophylla. A well-defined surrounding cellular
1985). Canhoto and Cruz (1996) have shown that somatic region could be stained with naphthol blue-black, revealing
embryos of F. sellowiana can arise directly from multi- sites of protein synthesis (Arruda et al. 2000).
cellular cell clumps on the epidermal adaxial surface A remarkable feature of the meristematic centers that pro-
of zygotic embryo cotyledons. Somatic embryos of F. duced the somatic embryos was the presence of polyphenolic
sellowiana have also been observed to arise directly from compounds. The cultures also showed an enhanced produc-
the cotyledonary tissues of zygotic embryos after sixteen tion of brown exudates of polyphenol origin. These com-
days in culture (Guerra et al. 2001). pounds appeared to inhibit hyperhydricity, thereby serving asG. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana 39
precursors of lignin synthesis as has been observed in Euca- References
lyptus nitens (Bandyopadhyay and Hamill 2000). Cells that Ammirato PV (1983) Embryogenesis. In: Evans DA, Sharp WR, Ammirato
stain green with the metachromatic stain TBO are reported to PV, Yamada Y (eds) Handbook of plant cell culture, vol 1. Macmillan,
contain polyphenols or lignin (Feder and O’Brien 1968). New York, pp 82–123
Arnold W, Mitrenga D, Mayersbach H-V (1975) Freeze-drying and em-
bedding in glycol methacrylate: results of histochemical reactions. Acta
Histochem Suppl 14: 271–277
Histochemistry of somatic embryos
Arruda SCC, Souza GM, Almeida M, Gonçalves AN (2000) Anatomical
and biochemical characterization of the calcium effect on Eucalyptus
As stated previously, we could observe four stages of
urophylla callus morphogenesis in vitro. Plant Cell Tissue Organ Cult
somatic-embryo development. The beginning of histogene- 63: 143–154
sis could be seen at the globular stage with the formation Bandyopadhyay S, Hamill JD (2000) Ultrastructural studies of somatic
of protoderm surrounding the globular embryo (Yeung embryos of Eucalyptus nitens and comparisons with zygotic embryos
found in mature seeds. Ann Bot 86: 237–244
1995). This feature could be clearly observed and was as- Borisjuk L, Panitz H-W, Manteffel R, Wobus U (1995) Embryogenesis
sociated with the presence of few starch grains but abun- of Vicia faba L.: histodifferentiation in relation to starch and storage
dant protein bodies. protein synthesis. Plant Physiol 143: 203–218
Botti C, Vasil IK (1984) The ontogeny of somatic embryos of Pennisetum
The methods used to identify the different types of poly- americanum (L.) K. Schum. II: in cultured immature inflorescences.
saccharides revealed different regions of acid and neutral Can J Bot 62: 1629–1635
compound accumulation in F. sellowiana somatic embryos. Branca C, Torelli A, Fermi P, Altamura MM, Bassi M (1994) Early phases
in in vitro culture of tomato cotyledons: starch accumulation and protein
Acid polysaccharides, indicated by the metachromatic stain
pattern in relation to the hormonal treatment. Protoplasma 182: 59–64
TBO, were observed mainly as constituents of the cellular Canhoto JM, Cruz GS (1996) Histodifferentiation of somatic embryos in
wall, whereas neutral polysaccharides, indicated by the cotyledons of pineapple guava (Feijoa sellowiana Berg.). Protoplasma
Schiff reaction, were mainly observed as reserve substances 191: 34–45
Colby SM, Juncosa AM, Stamp J, Meredith CP (1991) Developmental
in the intracellular domain. The Schiff reaction identifies anatomy of direct shoot organogenesis from leaf petiole of Vitis
neutral polysaccharides due to its requirement for the pres- vinifera (Vitaceae). Am J Bot 78: 260–269
ence of 1,2-glycol groups that are oxidized by periodic acid Dolezelova M, Dolezel J, Nesticky M (1992) Relationship of embryo-
genic competence in maize (Zea mays L.) leaves to mitotic activity,
(Trick and Pueschel 1990). cell cycle and nuclear DNA content. Plant Cell Tissue Organ Cult 31:
Histochemical evaluations of the torpedo and cotyle- 215–221
donary-stage somatic embryos revealed that starch grains Feder N, O’Brien TP (1968) Plant microtechnique: some principles and
new methods. Am J Bot 55: 123–142
occurred only in the root region, while protein bodies were
Finstad K, Brown DCW, Joy K (1993) Characterization of competence
observed in the protoderm as well as in the apical meri- during induction of somatic embryogenesis in alfalfa tissue culture.
stem. Borisjuk et al. (1995) have reported the presence of Plant Cell Tissue Organ Cult 34: 125–132
small and large starch grains in the protoderm and cotyle- Fransz PF, Schel JHN (1991) Cytodifferentiation during the development
of friable embryogenic callus of maize (Zea mays). Can J Bot 69: 26–33
donary cells, respectively, of Vicia faba zygotic embryos. Gahan PB (1984) Plant histochemistry and cytochemistry: an introduction.
The morphological abnormalities of the somatic embryos Academic Press, London
observed in this work could be due to the 2,4-D supplement Guerra MP, Dal Vesco LL, Ducroquet JPHJ, Nodari RO, Reis MS (2001)
Somatic embryogenesis in Feijoa sellowiana: genotype response, aux-
in the induction medium. The high incidence of abnormal
inic shock and synthetic seeds. Braz J Plant Physiol 13: 117–128
embryos in this species has also been attributed to their mul- Haccius B (1978) Question of unicellular origin of non-zygotic embryos
ticellular origin (Canhoto and Cruz 1996). Somatic embryos in callus cultures. Phytomorphology 28: 74–81
of Carya illinoinensis induced with 2,4-D have also been Karlsson SB, Vasil IK (1986) Morphology and ultrastructure of embryo-
genic cell suspension cultures of Panicum maximum (Guinea grass) and
observed to show a high incidence of abnormalities includ- Pennisetum purpureum (Napier grass). Am J Bot 73: 894–901
ing fasciated, fan-shaped, and tubular embryos (Rodriguez Lu C, Vasil IK (1985) Histology of somatic embryogenesis in Panicum
and Wetztein 1998). maximum (Guinea grass). Am J Bot 12: 1908–1913
Mangat BS, Pelekis MK, Cassells AC (1990) Changes in the starch con-
In conclusion, the results of the present work show that so- tent during organogenesis in in vitro cultured Begonia rex stem ex-
matic embryos of F. sellowiana can be induced from embryo- plants. Physiol Plant 79: 267–274
genic cells arising in meristematic centers or from clusters of Martin AB, Cuadrado Y, Guerra H, Gallego P, Hita O, Martin L, Dorado A,
Villalobos N (2000) Differences in the contents of total sugars, starch
isolated cells. The presence of few starch grains and abundant
and sucrose in embryogenic and non-embryogenic calli from Medicago
protein bodies was a distinctive feature of globular somatic arborea L. Plant Sci 154: 143–151
embryos, while cells in the torpedo and cotyledonary stages McDaniel CN (1984) Competence, determination and induction in plant
contained abundant starch grains. This could be associated development. In: Malacinski GM, Bryant SV (eds) Pattern formation.
Macmillan, New York, pp 393–412
with the strategy of storing reserve compounds to be used in Michaux-Ferrière N, Schwendiman J (1993) Initiation sequence patterns
the further conversion of the embryos to plantlets. for somatic embryos originating cells. Acta Bot Gallica 140: 603–61340 G. C. Cangahuala-Inocente et al.: Somatic embryos of Feijoa sellowiana Morel GM, Wetmore RH (1951) Tissue culture of monocotyledons. Am Stamp JA (1987) Somatic embryogenesis in cassava: the anatomy and J Bot 38: 138–140 morphology of the regeneration process. Ann Bot 57: 451–459 O’Brien TP, Feder N, McCully ME (1965) Polychromatic staining of Taylor MG, Vasil IK (1996) The ultrastructure of somatic embryo devel- plant cell walls by toluidine blue O. Protoplasma 59: 368–373 opment in pearl millet (Pennsietum glaucum; Poaceae). Am J Bot 83: Puigderrajols P, Mir G, Molinas M (2001) Ultrastructure of early em- 28–44 bryogenesis by multicellular and unicellular pathways in cork oak Torrey JG (1977) Cytodifferentiation in cultured cells and tissues. (Quercus suber L.). Ann Bot 87: 179–189 HortScience 12: 14–15 Rodriguez APM, Wetzstein HY (1998) A morphological and histological Trick H, Pueschel C (1990) Cytochemistry of pit plugs in Bossiela cali- comparison of the initiation and development of pecan (Carya illinoinen- fornica (Corallinales, Rhodophyta). Phycologia 29: 403–409 sis) somatic embryogenesis cultures induced with naphthaleneacetic acid von Arnold S, Eriksson T (1981) In vitro studies of adventitious shoot or 2,4-dichlorophenoxyacetic acid. Protoplasma 204: 71–83 formation in Pinus contorta. Can J Bot 59: 870–874 Rout GR, Samantaray S, Mottley J, Yokoya K, Mandegaran Z, Sarasan V, – Sabala I, Bozhkov P, Dyachok J, Filonova L (2002) Developmental Kandasamy K, Roberts AV, Das P (1998) Histology of in vitro organo- pathways of somatic embryogenesis. Plant Cell Tissue Organ Cult 69: genesis and somatic embryogenesis in callus cultures of Rosa. Biologia 233–249 (Bratislava) 53: 121–126 Williams EG, Maheswaran G (1986) Somatic embryogenesis: factors influ- Shewry PR, Napier JA, Tatham AS (1995) Seed storage proteins: struc- encing coordinated behavior of cells as an embryogenic group. Ann Bot tures and biosynthesis. Plant Cell 7: 945–956 57: 443–462 Shotwell MA, Larkins BA (1989) The molecular biology and biochemistry Yeung EC (1995) Structural and developmental patterns in somatic em- of seed storage proteins. In: Marcus A (ed) The biochemistry of plants: a bryogenesis. In: Yeung EC (ed) In vitro embryogenesis in plants. comprehensive treatise, vol 15. Academic Press, San Diego, pp 297–345 Kluwer Academic, Dordrecht, pp 205–248
You can also read

















































