Review Article Hair cell regeneration from inner ear progenitors in the mammalian cochlea
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Am J Stem Cells 2020;9(3):25-35
www.AJSC.us /ISSN:2160-4150/AJSC0112726
Review Article
Hair cell regeneration from inner ear
progenitors in the mammalian cochlea
Shasha Zhang1, Ruiying Qiang1, Ying Dong1, Yuan Zhang1, Yin Chen5, Han Zhou5, Xia Gao5, Renjie Chai1,2,3,4,5
1
Key Laboratory for Developmental Genes and Human Disease, Ministry of Education, Institute of Life Sciences,
Southeast University, Nanjing 210096, China; 2Co-Innovation Center of Neuroregeneration, Nantong University,
Nantong 226001, China; 3Institute for Stem Cell and Regeneration, Chinese Academy of Science, Beijing, China;
4
Jiangsu Province High-Tech Key Laboratory for Bio-Medical Research, Southeast University, Nanjing 211189,
China; 5Department of Otolaryngology Head and Neck Surgery, Affiliated Drum Tower Hospital of Nanjing
University Medical School, Jiangsu Provincial Key Medical Discipline (Laboratory), Nanjing 210008, China
Received April 17, 2020; Accepted June 10, 2020; Epub June 15, 2020; Published June 30, 2020
Abstract: Cochlear hair cells (HCs) are the mechanoreceptors of the auditory system, and because these cells
cannot be spontaneously regenerated in adult mammals, hearing loss due to HC damage is permanent. However,
cochleae of neonatal mice harbor some progenitor cells that retain limited ability to give rise to new HCs in vivo. Here
we review the regulatory factors, signaling pathways, and epigenetic factors that have been reported to play roles in
HC regeneration in the neonatal mammalian cochlea.
Keywords: Cochlea, inner ear progenitor, hair cell regeneration, transcription factor, signaling pathway
Introduction tors, regulatory factors and signaling pathways
regulate HC regeneration.
Sensorineural hearing loss, one of the most
common health problems around the world, is Inner ear progenitors in the neonatal cochlea
mainly caused by cochlear hair cell (HC) dam-
age or loss [1]. In non-mammalian vertebrates, In recent years, researchers have found that
such as birds and fish, HCs can be spontane- the SCs of the cochlea have certain ability for
ously regenerated from supporting cells (SCs) proliferation and differentiation, and as de-
after damage [2-4]. However, HCs in the adult scribed above, these cells can first divide and
mammalian cochlea cannot be spontaneously then differentiate into HCs or they can trans-
regenerated, and only neonatal cochlear HCs differentiate directly into HCs [10, 17]. White et
have a limited capacity for regeneration [5, 6]. al. isolated P27+ transgenic neonatal mouse
cochlear SCs and tested the ability of the cell
Damaged mammalian vestibular organs can
cycle re-entry and HC regeneration [10]. The
also generate new HCs from SCs in limited
presence of both BrdU+ and BrdU- regenerated
numbers [7-9]. It has been reported that pro-
HCs indicated that SCs can generate new HCs
genitor cells can be isolated from the auditory
through both direct differentiation and mitotic
and vestibular organs of the inner ear and can
pathways [10, 18].
form spheres and self-renew in vitro [10-13].
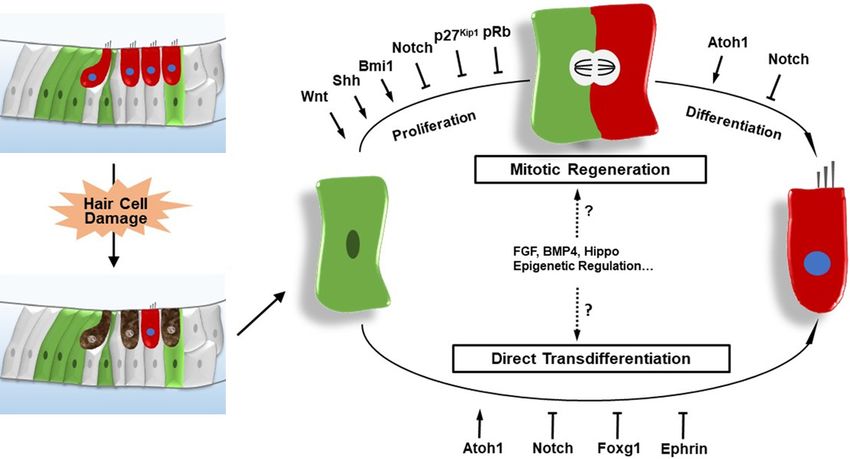
HCs are regenerated through two mechanis- Leucine-rich repeat-containing G-protein cou-
ms. In mitotic regeneration, inner ear progeni- pled receptor 5 (Lgr5), a Wnt signaling down-
tors re-enter the cell cycle, divide mitotically, stream target gene, has been reported to be a
and then differentiate into new HCs. In direct progenitor/stem cell marker in many other tis-
trans-differentiation, inner ear progenitors swi- sues [19, 20]. Chai et al. and Shi et al. both
tch cell fate and directly differentiate into new reported that cochlear Lgr5+ cells, a subset of
HCs [14-16]. We will focus in this review on the SCs including inner pillar cells, inner border
mechanisms through which transcription fac- cells, third-row Deiters’ cells, and the lateral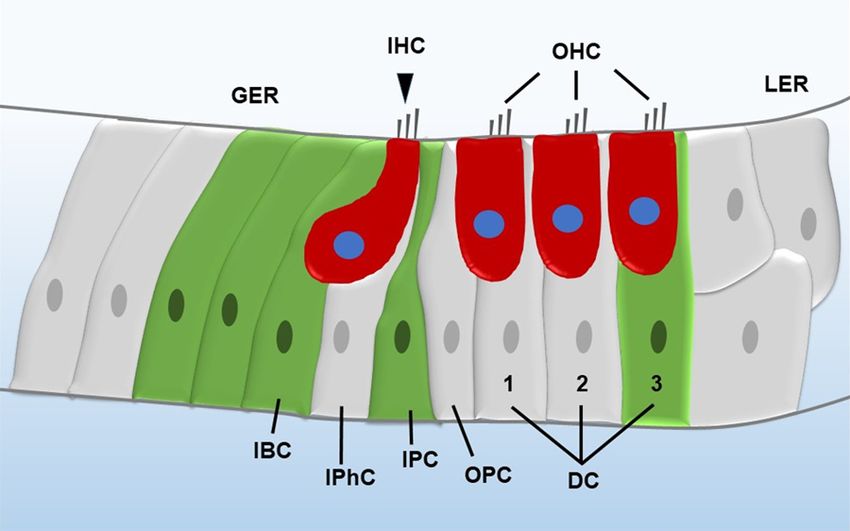
Hair cell regeneration
HCs in vitro in a similar man-
ner as Lgr5+ progenitors [25].
The same number of isolated
Lgr6+ cells generates signifi-
cantly more Myosin7a+ HCs
compared to Lgr5+ progeni-
tors, while Lgr5+ progenitors
form more cell spheres than
Lgr6+ cells in vitro [26], which
suggests that Lgr6+ cells have
greater ability for differentia-
tion and lesser ability for pro-
liferation compared to Lgr5+
progenitors. Another reported
inner ear progenitor marker is
Frizzled9, which is a Wnt re-
Figure 1. Illustration of the mammalian cochlea. The red cells are HCs, and ceptor gene. Frizzled9 is ex-
the green cells are Lgr5+ progenitors. IHC, inner hair cell; OHC, outer hair pressed in inner phalangeal
cell; GER, greater epithelial ridge; LER, lesser epithelial ridge; DC, Deiters’ cells, inner border cells, and
cell; OPC, outer pillar cell; IPC, inner pillar cell; IPhC, inner phalangeal cell;
IBC, inner border cell.
third-row Deiters’ cells in neo-
natal cochlea, and Frizzled9+
cells could regenerate HCs
greater epithelial ridge (Figure 1), are the inner both in vivo and in vitro. Moreover, Frizzled9+
ear progenitors in the neonatal mouse cochlea cells have a similar capacity for proliferation,
[21, 22]. These Lgr5+ progenitors have been differentiation, and HC generation as Lgr5+
shown to regenerate HCs in the neonatal progenitors [27].
cochlea both in vivo and in vitro, and Wnt sig-
naling induction either by Wnt agonists or in In summary, the discovery of inner ear progeni-
β-catenin overexpression transgenic mice pro- tors has provided a new approach for cell trans-
motes the proliferation of Lgr5+ progenitors plantation therapy. As mentioned above, there
and HC regeneration [21, 23]. are two mechanisms for HC regeneration. One
is trans-differentiation in which the inner ear
In another study, Jan et al. used reporter mice progenitors switch cell fate to become HCs, and
for Axin2 gene, which is a downstream negative the other is mitotic regeneration in which inner
feedback gene of the Wnt signaling pathway ear progenitors proliferate and then differenti-
[24], and showed in both in vitro cell culture ate into new HCs. Many transcription factors
and in vivo animal experiments that Axin2+ and signaling pathways are reported to be in-
tympanic border cells have similar characteris- volved in the development of the inner ear, and
tics as cochlear progenitors. These cells can several factors have been shown to be involved
proliferate into cell colonies and can be differ- in HC regeneration in the neonatal mouse
entiated into SCs and HCs. Moreover, the ability cochlea, including Atoh1, p27Kip1, pRb, Foxg1,
of these Axin2+ cells to proliferate and differen- and the Wnt, Notch, Hedgehog, and Ephrin sig-
tiate can be induced by Wnt agonists and sup- naling pathways (Figure 2).
pressed by Wnt inhibitors, similar with Lgr5+
progenitors. Therefore, it is suggested that HC regeneration: transcription factors and
Axin2+ cells might also be a potential source of regulatory factors
progenitors for treating hearing disorders.
Atho1 (also called Math1) is a helix-loop-helix
Recently, two other genes have been reported transcription factor that is essential for HC dif-
to be novel inner ear progenitor markers. The ferentiation. The expression of Atoh1 is visi-
first is Lgr6, which is also a Wnt-signaling do- ble from embryonic day 14.5 in the cochlea.
wnstream target gene. Lgr6+ cells, which only Deletion of the Atoh1 gene leads to the failure
include inner pillar cells in the neonatal mouse of HC formation, while its overexpression in-
cochlea, are a subpopulation of Lgr5+ progeni- duces ectopic HCs [28, 29]. Atoh1 also plays
tors, and Lgr6+ cells can generate Myosin7a+ important roles later during inner ear develop-
26 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
Figure 2. The regulation of HC regeneration in the neonatal mammalian cochlea after HC damage. HCs are regener-
ated through mitotic regeneration-in which progenitors re-enter the cell cycle, mitotically divide, and then differenti-
ate into new HCs-or through direct trans-differentiation in which progenitors switch cell fate and directly differentiate
into new HCs.
ment in HC survival and maturation [30, 31]. neonatal cochlea leads to the proliferation of
In neonatal mice, Atoh1 is also important by pillar cells without cell fate conversion [42-44],
promoting HC regeneration, and ectopic acti- which suggests that other factors are required
vation of Atoh1 induces new HCs generation in to induce the differentiation of SCs into HCs.
young postnatal mice [32, 33]. Moreover, in the
young adult deafened guinea pig model, forc- pRb is a retinoblastoma protein encoded by the
ed expression of Atoh1 induces HC regenera- retinoblastoma gene Rb1 and plays important
tion and decreases the hearing threshold [34]. roles in cell cycle exit, differentiation, and sur-
However, only a subset of these cells is able to vival [45, 46]. And it has been shown that dele-
give rise to new HCs, and they do so only at tion of Rb1 gene leads to the cell-cycle re-entry
early postnatal stages. of both embryonic and postnatal mammalian
HCs [47-49]. In neonatal mice, inactivation of
Cyclin-dependent kinase inhibitors (CKIs) are pRb in SCs results in cell cycle re-entry of both
divides into two families, the Cip/Kip family and pillar and Deiters’ cells and an increase in the
the Ink4 family, which play roles in governing number of pillar cells. The nuclei of Rb-/- mitotic
cell cycle transitions and maintaining postmi- pillar and Deiters’ cells were observed to mi-
totic state of numerous cell types [35, 36]. grate toward the HC layer and these cells divide
p19Ink4d (Cdkn2d) and p21Cip1 (Cdkn1a) have near the epithelial surface, similar to the SCs
been shown to be required in maintenance of in the regenerating avian cochlea. However,
the postmitotic state of HCs [37, 38]. p27Kip1 there are no newly regenerated HCs, and SC
(Cdkn1b), begins to be expressed in prosenso- death followed by HC loss occurs [50].
ry cells during the embryonic development of
the mammalian cochlea, and it persists at high Foxg1 (formerly called BF-1), one of the fork-
levels in SCs of the mature organ of Corti [39, head box family proteins, is involved in mor-
40]. Deletion of the p27Kip1 gene in the mouse phogenesis, cell fate determination, and prolif-
cochlea results in continuous cell proliferation eration in many tissues, especially in the brain
in the postnatal and adult mouse cochlea and [51-55]. Foxg1 knockout mice die in the peri-
to the appearance of supernumerary HCs and natal period and show shortened cochleae wi-
SCs [39, 41]. Deletion of p27Kip1 in SCs of the th multiple extra rows of HCs and SCs along
27 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
with vestibular defects [56, 57]. It was recently SC mitotic division and direct trans-differentia-
reported that conditional knockdown of Foxg1 tion. In contrast, Notch activation maintains
in SCs and progenitors in neonatal mice induc- SCs in a quiescent state, thereby inhibiting
es their direct trans-differentiation, but not regeneration of HCs [67, 68]. In the mamma-
their proliferation, and subsequently leads to lian postnatal cochlea, the Notch inhibition by
extra HCs [58]. γ-secretase inhibitor upregulates Atoh1 expres-
sion and results in the trans-differentiation of
HC regeneration: signaling pathways adjacent SCs into HCs [69, 70]. Li et al. report-
ed a direct interaction between the Notch and
During cochlear development, the canonical Wnt signaling pathways, that Notch inhibition
Wnt/β-catenin signaling pathway regulates cell induces mitotically generated HCs in mamma-
proliferation, cell fate decision, and HC differ- lian cochleae via activating the Wnt pathway
entiation, and Wnt signaling activation induces [71]. In addition, Notch and Wnt co-regulation
inner ear progenitor proliferation and HC regen- promotes SC proliferation and HC regenerati-
eration in both mammalian and non-mammali- on in both the cochlea and utricle in neonatal
an vertebrates [59, 60]. The inhibition of Wnt mice [72, 73]. A particularly exciting finding is
signaling in the embryonic mouse cochlea by that a genetic reprogramming process involv-
small molecule inhibitors or in transgenic mice ing β-catenin activation, Notch1 deletion, and
reduces the proliferation of prosensory cells Atoh1 overexpression is established and can
[61]. Conversely, Wnt signaling activation pro- promote extensive SC proliferation followed by
motes the prosensory domain formation and mitotic HC regeneration [74].
increases the number of HCs [62]. As men-
tioned above, Lgr5 and Lgr6, the Wnt signaling Hedgehog signaling is important for the forma-
downstream targets, are expressed in embry- tion of the dorsoventral axis of the inner ear,
onic and neonatal inner ear progenitors [22, and plays important roles in the prosensory
25]. And these progenitors can act as inner domain formation [75], the progenitor prolifera-
ear progenitors both in vivo and in vitro due to tion, and HC differentiation during inner ear
their ability of self-renew, proliferation, and dif- development [76]. The cell fate of progenitors,
ferentiation into HCs [21, 23, 63, 64]. In neona- whether differentiate into vestibular cells or
tal cochlea, both Wnt agonists treatment and auditory cells, is depend on the balance be-
β-catenin overexpression promote the prolifer- tween Wnt and Hedgehog signaling [77, 78].
ative capacity of Lgr5+ progenitors and subse- A few studies have reported the roles of Hed-
quent HC formation, whereas Wnt antagonists gehog signaling in mammalian HC regenera-
treatment reduce the proliferation and HC re- tion. Hedgehog signaling induces SC prolifera-
generation ability of Lgr5+ progenitors [23, 62, tion and HC regeneration in the postnatal rat
65]. Wnt activation also causes the Axin2+ tym- cochlea after neomycin treatment [79], and
panic border cells to proliferate and differenti- Sonic Hedgehog recombinant protein effecti-
ate into HCs and SCs in newborn mice [24]. vely promotes in vitro sphere formation, prolif-
The combined expression of β-catenin and At- eration, and differentiation of Lgr5+ progeni-
oh1 in Lgr5+ cells increases the HC regenera- tors isolated from the neonatal cochlea. Hed-
tion capacity of the postnatal cochlea by ten- gehog signaling was also proved to induce SC
fold, and these newly regenerated HCs can sur- proliferation and HC regeneration in neomyc-
vive until adulthood [66]. However, the com- in damaged cochlea by using transgenic R26-
bined expression of β-catenin and Atoh1 can- SmoM2 mice which constitutively activate Hed-
not induce HC regeneration in the adult mam- gehog signaling in the SCs leads to [80].
malian cochlea.
Ephrins and their receptors Ephs also play role
Because Notch signaling pathway plays impor- in HC regeneration. EphA4 receptor is expres-
tant roles in HC differentiation during inner ear sed in HCs, while Ephrin-B2 is present in SCs,
development, many researchers have exam- and this complementary pattern of expression
ined its roles in HC regeneration in postnatal is necessary for the establishment of the com-
cochlea. In both the zebrafish lateral line and partment boundary between HCs and SCs [81].
mature avian basilar papilla, inhibition of Notch Jean Defourny et al. demonstrated that mam-
signaling increases HC regeneration through malian HCs can be directly generated from SCs
28 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
by inhibition of ephrin-B2 signaling. Using ei- Future perspectives
ther ephrin-B2 conditional knockout mice, shR-
NA-mediated gene silencing, or soluble inhibi- Although HC regeneration can be induced by
tors, they found that downregulation of ephrin- many factors and signaling pathways in the
B2 signaling at late embryonic stages after HC neonatal mammalian cochlea, HCs cannot be
production, results in translocation of SC into regenerated in the adult mammalian cochlea
HC layers and subsequent cell fate switch from and current technologies are still quite far from
SC to HC [81]. Interestingly, throughout inner restoring hearing functions in the HC-damaged
ear development, Ephrin-B2 and Notch are ex- mammalian cochlea. Thus, further research is
pressed in similar SC types [82]. Moreover, needed to find ways to induce HC regenerati-
Ephrin-B2, whose expression is induced by on in both the neonatal and adult mammalian
Notch signaling, is reported to be a direct Not- cochlea.
ch signaling downstream target [83]; therefo-
re, Ephrin-B2 might be required following Not- First, more pathways and important factors,
ch lateral inhibition in order to segregate the including those that might regulate the prolif-
SCs from adjacent HCs. eration and differentiation of stem cells and
progenitors, such as FGF, BMP4, and Hippo sig-
HC regeneration: epigenetic regulation naling pathway, should be explored in the study
of HC regeneration. The FGF signaling pathway
Epigenetic factors have recently emerged as has been shown to be important in inner ear
important regulators in both inner ear develop- development and to be related to the otic plac-
ment and in HC regeneration. In the neuro- ode induction and the otic vesicle development
masts of developing zebrafish larva, inhibition [85-87]. Deletion of the FGF receptor 1 (Fgfr1)
of the histone-modifying enzyme lysine-specific gene in the inner ear results in decrease of the
demethylase 1 (LSD1) disrupts cell prolifera- number of proliferative prosensory cells and
tion, induces apoptosis, and reduces the num- subsequent decrease of the numbers of HCs
bers of sensory HCs and SCs [84]. And epigen- and SCs [88, 89]. The roles of the FGF signaling
etic regulation of Atoh1 was reported to guide pathway in HC regeneration has been explored
HC development in the developing mouse co- in the utricles of chickens and the lateral lines
chlea [10]. Inhibition of histone acetyltransfer- of zebrafish [90-93]. Many reports has shown
ase activity reduces H3K9 acetylation at the that BMP4 plays important roles in mammali-
Atoh1 locus and therefore prevents Atoh1 an and non-mammalian inner ear development
mRNA increase and subsequent HC differen- [94-100], and it is recently reported that BMP4
tiation. Interestingly, the H3K4me3/H3K27me3 can also antagonize HC regeneration in the
bivalent chromatin structure, observed in pro- avian auditory epithelium [101]. The Hippo/Yap
genitors, persists at the Atoh1 locus in perina- signaling pathway plays important roles in de-
tal SCs [10], suggesting the important roles of velopment, homeostasis, and regeneration in
such structures in HC regeneration. many tissues and cancer cells [102-106], and
it has been reported that Hippo/Yap controls
Histone deacetylase (HDAC) inhibitor treatment proliferation and differentiation of lung and
of HC-damaged chicken utricles reduces prolif- plays key roles in regeneration and fibrogenes-
eration of SCs, but does not affect HC regener- is after kidney injury. In zebrafish lateral line,
ation [63]. Similarly, inhibition of HDAC activity Yap1 plays important roles in HC differentia-
in HC-damaged zebrafish larvae also reduces tion. Knockdown of Yap1 in developing zebraf-
SC proliferation and subsequent HC regenera- ish affects development of the lateral line sys-
tion [23]. Bmi1, a Polycomb group protein and a tem and recapitulates the Prox1a deficiency in
component of the Polycomb repressive com- mechanosensory cells of neuromast [107]. All
plex 1, maintains the proliferative capacity of of the above factors and signaling pathways
SCs by sustaining high levels of Wnt signaling in can be used as good candidates for further HC
the neonatal mouse cochlea. In neonatal Bmi1- regeneration study in the mammalian inner ear.
deficient cochleae, SCs fail to re-enter the cell As mentioned above, many epigenetic regula-
cycle in response to HC damage, and the in tors such as LSD1, histone modifications, and
vitro sphere-forming ability of Bmi1-deficient HDAC inhibitors, which have been studied in
cochlear progenitors is also reduced [11]. inner ear development and HC regeneration in
29 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
non-mammalian organisms, are also very good gram of China (2017YFA01039000), the Str-
candidates for studying HC regeneration in the ategic Priority Research Program of the Chinese
mammalian inner ear. Academy of Science (XDA16010303), the Na-
tional Natural Science Foundation of China
Second, the interactions of multiple pathways (Nos. 81970892, 81970882), the Jiangsu Pro-
in cell proliferation and HC differentiation sh- vince Natural Science Foundation (BK2019-
ould be explored. As mentioned above, some 0062, BE2019711), Boehringer Ingelheim Ph-
research has studied the cross talk between arma GmbH, the Fundamental Research Funds
two or more signaling pathways and factors for the Central Universities (2242020R40137),
[72-74], but these studies are far from regener- the Excellence Project of Southeast University,
ating HCs and repairing inner ear damage in the Open Research Fund of the State Key
adult mammals. Laboratory of Genetic Engineering, Fudan
And lastly, the maturation and survival of new- University (No. SKLGE1809) and Shenzhen
ly generated HCs and HC regeneration in adult Fundamental Research Program (JCYJ2019-
mammals still remains a challenge. Bradley 0814093401920).
Walters et al. found that combining p27Kip1 dele-
Disclosure of conflict of interest
tion with ectopic Atoh1 expression surmounts
age-related decline of HC regeneration from None.
SCs, leading to conversion of SCs to HCs in
mature mouse cochleae and after noise dam- Address correspondence to: Dr. Renjie Chai, Co-
age [108]. Moreover, co-activation of GATA3 or Innovation Center of Neuroregeneration, Nantong
Pou4f3 and Atoh1 promoted conversion of SCs University, Nantong 226001, China; Key Laboratory
to HCs in adult mice and activation of Pou4F3 for Developmental Genes and Human Disease,
alone also converted mature SCs to HCs in vivo Ministry of Education, Institute of Life Sciences,
[108]. In another recent report, Yilai Shu et al. Southeast University, Nanjing 210096, China. Tel:
reported that transient co-activation of cell cy- +86-25-83790971; Fax: +86-25-83790971; E-mail:
cle activator Myc and inner ear progenitor gene renjiec@seu.edu.cn; Dr. Xia Gao, Department of
Notch1 induces proliferation of diverse adult Otorhinolaryngology Head and Neck Surgery, Nan-
cochlear sensory epithelial cell types, and en- jing Drum Tower Hospital, 321 Zhongshan Road,
ables adult SCs to respond to transcription Nanjing 210096, China. Tel: +86-25-83304616-
factor Atoh1 and efficiently trans-differentiate 61131; Fax: +86-25-83304616-61131; E-mail:
into HC-like cells [109]. Although it is excited xiagao@aliyun.com
to see these two recent reports that HC could
now be regenerated from SCs in adult mice by References
genes and signaling regulation, the regenera-
tion efficiency and the maturation of regener- [1] Wagner EL and Shin JB. Mechanisms of hair
ated HCs remains still a problem. More efforts, cell damage and repair. Trends Neurosci 2019;
such as other genes and signaling co-regula- 42: 414-424.
tion, apoptosis inhibition and maturation in- [2] Corwin JT and Oberholtzer JC. Fish n’ chicks:
duction of newly regenerated HCs, should be model recipes for hair-cell regeneration? Neu-
made in the future. ron 1997; 19: 951-954.
[3] Tucci DL and Rubel EW. Physiologic status of
In summary, much effort has been put into regenerated hair cells in the avian inner ear
exploring the mechanisms of HC regeneration following aminoglycoside ototoxicity. Otolaryn-
in the mammalian inner ear, and many factors gol Head Neck Surg 1990; 103: 443-450.
[4] Cotanche DA and Dopyera CE. Hair cell and
and signaling pathways have been shown to
supporting cell response to acoustic trauma in
play important roles in the neonatal cochlea.
the chick cochlea. Hear Res 1990; 46: 29-40.
However, these studies are still far from re- [5] Bermingham-McDonogh O and Rubel EW. Hair
generating HCs and repairing HC damage in cell regeneration: winging our way towards a
adult mammals, which is the ultimate research sound future. Curr Opin Neurobiol 2003; 13:
objective in this field. 119-126.
[6] Brigande JV and Heller S. Quo vadis, hair cell
Acknowledgements regeneration? Nat Neurosci 2009; 12: 679-
685.
This work was supported by grants from the [7] Forge A, Li L, Corwin JT and Nevill G. Ultrastruc-
Major State Basic Research Development Pro- tural evidence for hair cell regeneration in the
30 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
mammalian inner ear. Science 1993; 259: [21] Shi F, Kempfle JS and Edge AS. Wnt-responsive
1616-1619. Lgr5-expressing stem cells are hair cell pro-
[8] Warchol ME, Lambert PR, Goldstein BJ, Forge genitors in the cochlea. J Neurosci 2012; 32:
A and Corwin JT. Regenerative proliferation in 9639-9648.
inner ear sensory epithelia from adult guinea [22] Chai R, Xia A, Wang T, Jan TA, Hayashi T, Berm-
pigs and humans. Science 1993; 259: 1619- ingham-McDonogh O and Cheng AG. Dynamic
1622. expression of Lgr5, a Wnt target gene, in the
[9] Burns JC and Stone JS. Development and re- developing and mature mouse cochlea. J As-
generation of vestibular hair cells in mammals. soc Res Otolaryngol 2011; 12: 455-469.
Semin Cell Dev Biol 2017; 65: 96-105. [23] Chai R, Kuo B, Wang T, Liaw EJ, Xia A, Jan TA,
[10] White PM, Doetzlhofer A, Lee YS, Groves AK Liu Z, Taketo MM, Oghalai JS, Nusse R, Zuo J
and Segil N. Mammalian cochlear supporting and Cheng AG. Wnt signaling induces prolifera-
cells can divide and trans-differentiate into tion of sensory precursors in the postnatal
hair cells. Nature 2006; 441: 984-987. mouse cochlea. Proc Natl Acad Sci U S A 2012;
[11] Li H, Liu H and Heller S. Pluripotent stem cells 109: 8167-8172.
from the adult mouse inner ear. Nat Med [24] Jan TA, Chai R, Sayyid ZN, van Amerongen R,
2003; 9: 1293-1299. Xia A, Wang T, Sinkkonen ST, Zeng YA, Levin JR,
[12] Oshima K, Senn P and Heller S. Isolation of Heller S, Nusse R and Cheng AG. Tympanic bor-
sphere-forming stem cells from the mouse in- der cells are Wnt-responsive and can act as
ner ear. Methods Mol Biol 2009; 493: 141- progenitors for postnatal mouse cochlear
162. cells. Development 2013; 140: 1196-1206.
[13] Oshima K, Grimm CM, Corrales CE, Senn P, [25] Zhang Y, Chen Y, Ni W, Guo L, Lu X, Liu L, Li W,
Martinez Monedero R, Geleoc GS, Edge A, Holt Sun S, Wang L and Li H. Dynamic expression of
JR and Heller S. Differential distribution of Lgr6 in the developing and mature mouse co-
stem cells in the auditory and vestibular or- chlea. Front Cell Neurosci 2015; 9: 165.
gans of the inner ear. J Assoc Res Otolaryngol [26] Zhang Y, Guo L, Lu X, Cheng C, Sun S, Li W,
2007; 8: 18-31. Zhao L, Lai C, Zhang S, Yu C, Tang M, Chen Y,
[14] Franco B and Malgrange B. Concise review: re- Chai R and Li H. Characterization of Lgr6+ cells
generation in mammalian cochlea hair cells: as an enriched population of hair cell progeni-
help from supporting cells transdifferentiation. tors compared to Lgr5+ cells for hair cell gen-
Stem Cells 2017; 35: 551-556. eration in the neonatal mouse cochlea. Front
[15] Lu X, Shu Y, Tang M and Li H. Mammalian co- Mol Neurosci 2018; 11: 147.
chlear hair cell regeneration and ribbon syn- [27] Zhang S, Liu D, Dong Y, Zhang Z, Zhang Y, Zhou
apse reformation. Neural Plast 2016; 2016: H, Guo L, Qi J, Qiang R, Tang M, Gao X, Zhao C,
2523458. Chen X, Qian X and Chai R. Frizzled-9+ sup-
[16] Atkinson PJ, Huarcaya Najarro E, Sayyid ZN and porting cells are progenitors for the generation
Cheng AG. Sensory hair cell development and of hair cells in the postnatal mouse cochlea.
regeneration: similarities and differences. De- Front Mol Neurosci 2019; 12: 184.
velopment 2015; 142: 1561-1571. [28] Pan N, Jahan I, Kersigo J, Kopecky B, Santi P,
[17] Sinkkonen ST, Chai R, Jan TA, Hartman BH, Johnson S, Schmitz H and Fritzsch B. Condi-
Laske RD, Gahlen F, Sinkkonen W, Cheng AG, tional deletion of Atoh1 using Pax2-Cre results
Oshima K and Heller S. Intrinsic regenerative in viable mice without differentiated cochlear
potential of murine cochlear supporting cells. hair cells that have lost most of the organ of
Sci Rep 2011; 1: 26. Corti. Hear Res 2011; 275: 66-80.
[18] Savoy-Burke G, Gilels FA, Pan W, Pratt D, Que J, [29] Gubbels SP, Woessner DW, Mitchell JC, Ricci
Gan L, White PM and Kiernan AE. Activated AJ and Brigande JV. Functional auditory hair
notch causes deafness by promoting a sup- cells produced in the mammalian cochlea by in
porting cell phenotype in developing auditory utero gene transfer. Nature 2008; 455: 537-
hair cells. PLoS One 2014; 9: e108160. 541.
[19] Barker N, van Es JH, Kuipers J, Kujala P, van [30] Chonko KT, Jahan I, Stone J, Wright MC, Fuji-
den Born M, Cozijnsen M, Haegebarth A, Korv- yama T, Hoshino M, Fritzsch B and Maricich
ing J, Begthel H, Peters PJ and Clevers H. Iden- SM. Atoh1 directs hair cell differentiation and
tification of stem cells in small intestine and survival in the late embryonic mouse inner ear.
colon by marker gene Lgr5. Nature 2007; 449: Dev Biol 2013; 381: 401-410.
1003-1007. [31] Cai T, Seymour ML, Zhang H, Pereira FA and
[20] Jaks V, Barker N, Kasper M, van Es JH, Snip- Groves AK. Conditional deletion of Atoh1 re-
pert HJ, Clevers H and Toftgard R. Lgr5 marks veals distinct critical periods for survival and
cycling, yet long-lived, hair follicle stem cells. function of hair cells in the organ of Corti. J
Nat Genet 2008; 40: 1291-1299. Neurosci 2013; 33: 10110-10122.
31 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
[32] Liu Z, Dearman JA, Cox BC, Walters BJ, Zhang shRNA lentiviral transduction. J Assoc Res Oto-
L, Ayrault O, Zindy F, Gan L, Roussel MF and laryngol 2013; 14: 495-508.
Zuo J. Age-dependent in vivo conversion of [44] Oesterle EC, Chien WM, Campbell S, Nellimar-
mouse cochlear pillar and Deiters’ cells to im- la P and Fero ML. p27(Kip1) is required to
mature hair cells by Atoh1 ectopic expression. maintain proliferative quiescence in the adult
J Neurosci 2012; 32: 6600-6610. cochlea and pituitary. Cell Cycle 2011; 10:
[33] Kelly MC, Chang Q, Pan A, Lin X and Chen P. 1237-1248.
Atoh1 directs the formation of sensory mosa- [45] Classon M and Harlow E. The retinoblastoma
ics and induces cell proliferation in the postna- tumour suppressor in development and can-
tal mammalian cochlea in vivo. J Neurosci cer. Nat Rev Cancer 2002; 2: 910-917.
2012; 32: 6699-6710. [46] Lipinski MM and Jacks T. The retinoblastoma
[34] Izumikawa M, Minoda R, Kawamoto K, Abrash- gene family in differentiation and develop-
kin KA, Swiderski DL, Dolan DF, Brough DE and ment. Oncogene 1999; 18: 7873-7882.
Raphael Y. Auditory hair cell replacement and [47] Sage C, Huang M, Vollrath MA, Brown MC,
hearing improvement by Atoh1 gene therapy in Hinds PW, Corey DP, Vetter DE and Chen ZY.
deaf mammals. Nat Med 2005; 11: 271-276. Essential role of retinoblastoma protein in
[35] Cunningham JJ and Roussel MF. Cyclin-depen- mammalian hair cell development and hear-
dent kinase inhibitors in the development of ing. Proc Natl Acad Sci U S A 2006; 103: 7345-
the central nervous system. Cell Growth Differ 7350.
2001; 12: 387-396. [48] Sage C, Huang M, Karimi K, Gutierrez G, Voll-
[36] Vidal A and Koff A. Cell-cycle inhibitors: three rath MA, Zhang DS, Garcia-Anoveros J, Hinds
families united by a common cause. Gene PW, Corwin JT, Corey DP and Chen ZY. Prolifera-
2000; 247: 1-15. tion of functional hair cells in vivo in the ab-
[37] Laine H, Doetzlhofer A, Mantela J, Ylikoski J, sence of the retinoblastoma protein. Science
Laiho M, Roussel MF, Segil N and Pirvola U. 2005; 307: 1114-1118.
p19(Ink4d) and p21(Cip1) collaborate to main- [49] Mantela J, Jiang Z, Ylikoski J, Fritzsch B, Zack-
senhaus E and Pirvola U. The retinoblastoma
tain the postmitotic state of auditory hair cells,
gene pathway regulates the postmitotic state
their codeletion leading to DNA damage and
of hair cells of the mouse inner ear. Develop-
p53-mediated apoptosis. J Neurosci 2007; 27:
ment 2005; 132: 2377-2388.
1434-1444.
[50] Yu Y, Weber T, Yamashita T, Liu Z, Valentine
[38] Chen P, Zindy F, Abdala C, Liu F, Li X, Roussel
MB, Cox BC and Zuo J. In vivo proliferation of
MF and Segil N. Progressive hearing loss in
postmitotic cochlear supporting cells by acute
mice lacking the cyclin-dependent kinase in-
ablation of the retinoblastoma protein in neo-
hibitor Ink4d. Nat Cell Biol 2003; 5: 422-426.
natal mice. J Neurosci 2010; 30: 5927-5936.
[39] Chen P and Segil N. p27(Kip1) links cell prolif-
[51] Tian C, Gong Y, Yang Y, Shen W, Wang K, Liu J,
eration to morphogenesis in the developing
Xu B, Zhao J and Zhao C. Foxg1 has an essen-
organ of Corti. Development 1999; 126: 1581-
tial role in postnatal development of the den-
1590. tate gyrus. J Neurosci 2012; 32: 2931-2949.
[40] Ruben RJ. Development of the inner ear of the [52] Xuan S, Baptista CA, Balas G, Tao W, Soares VC
mouse: a radioautographic study of terminal and Lai E. Winged helix transcription factor
mitoses. Acta Otolaryngol 1967; Suppl 220: BF-1 is essential for the development of the
221-244. cerebral hemispheres. Neuron 1995; 14:
[41] Lowenheim H, Furness DN, Kil J, Zinn C, Gultig 1141-1152.
K, Fero ML, Frost D, Gummer AW, Roberts JM, [53] Huh S, Hatini V, Marcus RC, Li SC and Lai E.
Rubel EW, Hackney CM and Zenner HP. Gene Dorsal-ventral patterning defects in the eye of
disruption of p27(Kip1) allows cell proliferation BF-1-deficient mice associated with a restrict-
in the postnatal and adult organ of corti. Proc ed loss of shh expression. Dev Biol 1999; 211:
Natl Acad Sci U S A 1999; 96: 4084-4088. 53-63.
[42] Liu Z, Walters BJ, Owen T, Brimble MA, Steigel- [54] Adesina AM, Veo BL, Courteau G, Mehta V, Wu
man KA, Zhang L, Mellado Lagarde MM, Valen- X, Pang K, Liu Z, Li XN and Peters L. FOXG1
tine MB, Yu Y, Cox BC and Zuo J. Regulation of expression shows correlation with neuronal
p27Kip1 by Sox2 maintains quiescence of in- differentiation in cerebellar development, ag-
ner pillar cells in the murine auditory sensory gressive phenotype in medulloblastomas, and
epithelium. J Neurosci 2012; 32: 10530- survival in a xenograft model of medulloblas-
10540. toma. Hum Pathol 2015; 46: 1859-1871.
[43] Maass JC, Berndt FA, Canovas J and Kukuljan [55] Pratt T, Tian NM, Simpson TI, Mason JO and
M. p27Kip1 knockdown induces proliferation Price DJ. The winged helix transcription factor
in the organ of Corti in culture after efficient Foxg1 facilitates retinal ganglion cell axon
32 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
crossing of the ventral midline in the mouse. and Atoh1. J Neurosci 2015; 35: 10786-
Development 2004; 131: 3773-3784. 10798.
[56] Pauley S, Lai E and Fritzsch B. Foxg1 is re- [67] Daudet N, Gibson R, Shang J, Bernard A, Lewis
quired for morphogenesis and histogenesis of J and Stone J. Notch regulation of progenitor
the mammalian inner ear. Dev Dyn 2006; 235: cell behavior in quiescent and regenerating
2470-2482. auditory epithelium of mature birds. Dev Biol
[57] Hwang CH, Simeone A, Lai E and Wu DK. Foxg1 2009; 326: 86-100.
is required for proper separation and forma- [68] Ma EY, Rubel EW and Raible DW. Notch signal-
tion of sensory cristae during inner ear devel- ing regulates the extent of hair cell regenera-
opment. Dev Dyn 2009; 238: 2725-2734. tion in the zebrafish lateral line. J Neurosci
[58] Zhang S, Zhang Y, Dong Y, Guo L, Zhang Z, 2008; 28: 2261-2273.
Shao B, Qi J, Zhou H, Zhu W, Yan X, Hong G, [69] Korrapati S, Roux I, Glowatzki E and Doetzl-
Zhang L, Zhang X, Tang M, Zhao C, Gao X and hofer A. Notch signaling limits supporting cell
Chai R. Knockdown of Foxg1 in supporting plasticity in the hair cell-damaged early post-
cells increases the trans-differentiation of sup- natal murine cochlea. PLoS One 2013; 8:
porting cells into hair cells in the neonatal e73276.
mouse cochlea. Cell Mol Life Sci 2020; 77: [70] Mizutari K, Fujioka M, Hosoya M, Bramhall N,
1401-1419. Okano HJ, Okano H and Edge AS. Notch inhibi-
[59] Romero-Carvajal A, Navajas Acedo J, Jiang L, tion induces cochlear hair cell regeneration
Kozlovskaja-Gumbriene A, Alexander R, Li H and recovery of hearing after acoustic trauma.
and Piotrowski T. Regeneration of sensory hair Neuron 2013; 77: 58-69.
cells requires localized interactions between [71] Li W, Wu J, Yang J, Sun S, Chai R, Chen ZY and
the notch and wnt pathways. Dev Cell 2015; Li H. Notch inhibition induces mitotically gener-
34: 267-282. ated hair cells in mammalian cochleae via ac-
[60] Jacques BE, Montgomery WH 4th, Uribe PM, tivating the Wnt pathway. Proc Natl Acad Sci U
Yatteau A, Asuncion JD, Resendiz G, Matsui JI S A 2015; 112: 166-171.
and Dabdoub A. The role of Wnt/beta-catenin [72] Wu J, Li W, Lin C, Chen Y, Cheng C, Sun S, Tang
signaling in proliferation and regeneration of M, Chai R and Li H. Co-regulation of the Notch
the developing basilar papilla and lateral line. and Wnt signaling pathways promotes sup-
Dev Neurobiol 2014; 74: 438-456. porting cell proliferation and hair cell regenera-
[61] Jacques BE, Puligilla C, Weichert RM, Ferrer- tion in mouse utricles. Sci Rep 2016; 6: 29418.
Vaquer A, Hadjantonakis AK, Kelley MW and [73] Ni W, Zeng S, Li W, Chen Y, Zhang S, Tang M,
Dabdoub A. A dual function for canonical Wnt/ Sun S, Chai R and Li H. Wnt activation followed
beta-catenin signaling in the developing mam- by Notch inhibition promotes mitotic hair cell
malian cochlea. Development 2012; 139: regeneration in the postnatal mouse cochlea.
4395-4404. Oncotarget 2016; 7: 66754-66768.
[62] Shi F, Hu L, Jacques BE, Mulvaney JF, Dabdoub [74] Ni W, Lin C, Guo L, Wu J, Chen Y, Chai R, Li W
A and Edge AS. β-catenin is required for hair- and Li H. Extensive supporting cell prolifera-
cell differentiation in the cochlea. J Neurosci tion and mitotic hair cell generation by in vivo
2014; 34: 6470-6479. genetic reprogramming in the neonatal mouse
[63] Wang T, Chai R, Kim GS, Pham N, Jansson L, cochlea. J Neurosci 2016; 36: 8734-8745.
Nguyen DH, Kuo B, May LA, Zuo J, Cunningham [75] Driver EC, Pryor SP, Hill P, Turner J, Ruther U,
LL and Cheng AG. Lgr5+ cells regenerate hair Biesecker LG, Griffith AJ and Kelley MW.
cells via proliferation and direct transdifferen- Hedgehog signaling regulates sensory cell for-
tiation in damaged neonatal mouse utricle. mation and auditory function in mice and hu-
Nat Commun 2015; 6: 6613. mans. J Neurosci 2008; 28: 7350-7358.
[64] Cox BC, Chai R, Lenoir A, Liu Z, Zhang L, Nguy- [76] Zarei S, Zarei K, Fritzsch B and Elliott KL. Sonic
en DH, Chalasani K, Steigelman KA, Fang J, hedgehog antagonists reduce size and alter
Rubel EW, Cheng AG and Zuo J. Spontaneous patterning of the frog inner ear. Dev Neurobiol
hair cell regeneration in the neonatal mouse 2017; 77: 1385-1400.
cochlea in vivo. Development 2014; 141: 816- [77] Riccomagno MM, Takada S and Epstein DJ.
829. Wnt-dependent regulation of inner ear mor-
[65] Shi F, Hu L and Edge AS. Generation of hair phogenesis is balanced by the opposing and
cells in neonatal mice by beta-catenin overex- supporting roles of Shh. Genes Dev 2005; 19:
pression in Lgr5-positive cochlear progenitors. 1612-1623.
Proc Natl Acad Sci U S A 2013; 110: 13851- [78] Brown AS and Epstein DJ. Otic ablation of
13856. smoothened reveals direct and indirect re-
[66] Kuo BR, Baldwin EM, Layman WS, Taketo MM quirements for Hedgehog signaling in inner
and Zuo J. In vivo cochlear hair cell generation ear development. Development 2011; 138:
and survival by coactivation of beta-catenin 3967-3976.
33 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
[79] Lu N, Chen Y, Wang Z, Chen G, Lin Q, Chen ZY ear sensory epithelia. Dev Biol 2001; 238:
and Li H. Sonic hedgehog initiates cochlear 247-259.
hair cell regeneration through downregulation [92] Jiang L, Romero-Carvajal A, Haug JS, Seidel
of retinoblastoma protein. Biochem Biophys CW and Piotrowski T. Gene-expression analysis
Res Commun 2013; 430: 700-705. of hair cell regeneration in the zebrafish lateral
[80] Chen Y, Lu X, Guo L, Ni W, Zhang Y, Zhao L, Wu line. Proc Natl Acad Sci U S A 2014; 111:
L, Sun S, Zhang S, Tang M, Li W, Chai R and Li E1383-1392.
H. Hedgehog signaling promotes the prolifera- [93] Pirvola U, Cao Y, Oellig C, Suoqiang Z, Petters-
tion and subsequent hair cell formation of pro- son RF and Ylikoski J. The site of action of neu-
genitor cells in the neonatal mouse cochlea. ronal acidic fibroblast growth factor is the or-
Front Mol Neurosci 2017; 10: 426. gan of Corti of the rat cochlea. Proc Natl Acad
[81] Defourny J, Mateo Sanchez S, Schoonaert L, Sci U S A 1995; 92: 9269-9273.
Robberecht W, Davy A, Nguyen L and Mal- [94] Gerlach LM, Hutson MR, Germiller JA, Nguyen-
grange B. Cochlear supporting cell transdiffer- Luu D, Victor JC and Barald KF. Addition of the
entiation and integration into hair cell layers by BMP4 antagonist, noggin, disrupts avian inner
inhibition of ephrin-B2 signalling. Nat Commun ear development. Development 2000; 127:
2015; 6: 7017. 45-54.
[82] Lanford PJ, Lan Y, Jiang R, Lindsell C, Weinmas- [95] Blauwkamp MN, Beyer LA, Kabara L, Takemu-
ter G, Gridley T and Kelley MW. Notch signalling ra K, Buck T, King WM, Dolan DF, Barald KF,
pathway mediates hair cell development in Raphael Y and Koenig RJ. The role of bone
mammalian cochlea. Nat Genet 1999; 21: morphogenetic protein 4 in inner ear develop-
289-292. ment and function. Hear Res 2007; 225: 71-
[83] D’Amato G, Luxan G and de la Pompa JL. Notch 79.
signalling in ventricular chamber development [96] Vervoort R, Ceulemans H, Van Aerschot L,
and cardiomyopathy. FEBS J 2016; 283: 4223- D’Hooge R and David G. Genetic modification
4237. of the inner ear lateral semicircular canal phe-
[84] Kawamoto K, Izumikawa M, Beyer LA, Atkin GM notype of the Bmp4 haplo-insufficient mouse.
and Raphael Y. Spontaneous hair cell regen- Biochem Biophys Res Commun 2010; 394:
eration in the mouse utricle following gentami- 780-785.
cin ototoxicity. Hear Res 2009; 247: 17-26. [97] Liu W, Oh SH, Kang Yk Y, Li G, Doan TM, Little
[85] Pirvola U, Spencer-Dene B, Xing-Qun L, Ket- M, Li L, Ahn K, Crenshaw EB 3rd and Frenz DA.
tunen P, Thesleff I, Fritzsch B, Dickson C and Bone morphogenetic protein 4 (BMP4): a regu-
Ylikoski J. FGF/FGFR-2(IIIb) signaling is essen- lator of capsule chondrogenesis in the devel-
tial for inner ear morphogenesis. J Neurosci oping mouse inner ear. Dev Dyn 2003; 226:
2000; 20: 6125-6134. 427-438.
[86] Schimmang T. Expression and functions of [98] Waqas M, Sun S, Xuan C, Fang Q, Zhang X, Is-
FGF ligands during early otic development. Int lam IU, Qi J, Zhang S, Gao X, Tang M, Shi H, Li
J Dev Biol 2007; 51: 473-481. H and Chai R. Bone morphogenetic protein 4
[87] Wright TJ and Mansour SL. Fgf3 and Fgf10 are promotes the survival and preserves the struc-
required for mouse otic placode induction. De- ture of flow-sorted Bhlhb5+ cochlear spiral
velopment 2003; 130: 3379-3390. ganglion neurons in vitro. Sci Rep 2017; 7:
[88] Ono K, Kita T, Sato S, O’Neill P, Mak SS, Pas- 3506.
chaki M, Ito M, Gotoh N, Kawakami K, Sasai Y [99] Li H, Corrales CE, Wang Z, Zhao Y, Wang Y, Liu
and Ladher RK. FGFR1-Frs2/3 signalling ma- H and Heller S. BMP4 signaling is involved in
intains sensory progenitors during inner ear the generation of inner ear sensory epithelia.
hair cell formation. PLoS Genet 2014; 10: BMC Dev Biol 2005; 5: 16.
e1004118. [100] Ohyama T, Basch ML, Mishina Y, Lyons KM, Se-
[89] Pirvola U, Ylikoski J, Trokovic R, Hebert JM, Mc- gil N and Groves AK. BMP signaling is neces-
Connell SK and Partanen J. FGFR1 is required sary for patterning the sensory and nonsenso-
for the development of the auditory sensory ry regions of the developing mammalian
epithelium. Neuron 2002; 35: 671-680. cochlea. J Neurosci 2010; 30: 15044-15051.
[90] Ku YC, Renaud NA, Veile RA, Helms C, Voelker [101] Lewis RM, Keller JJ, Wan L and Stone JS. Bone
CC, Warchol ME and Lovett M. The transcrip- morphogenetic protein 4 antagonizes hair cell
tome of utricle hair cell regeneration in the regeneration in the avian auditory epithelium.
avian inner ear. J Neurosci 2014; 34: 3523- Hear Res 2018; 364: 1-11.
3535. [102] Hong AW, Meng Z and Guan KL. The Hippo
[91] Bermingham-McDonogh O, Stone JS, Reh TA pathway in intestinal regeneration and dis-
and Rubel EW. FGFR3 expression during devel- ease. Nat Rev Gastroenterol Hepatol 2016;
opment and regeneration of the chick inner 13: 324-337.
34 Am J Stem Cells 2020;9(3):25-35Hair cell regeneration
[103] Fu V, Plouffe SW and Guan KL. The Hippo path- [107] Loh SL, Teh C, Muller J, Guccione E, Hong W
way in organ development, homeostasis, and and Korzh V. Zebrafish yap1 plays a role in dif-
regeneration. Curr Opin Cell Biol 2017; 49: 99- ferentiation of hair cells in posterior lateral
107. line. Sci Rep 2014; 4: 4289.
[104] Zygulska AL, Krzemieniecki K and Pierzchalski [108] Walters BJ, Coak E, Dearman J, Bailey G, Ya-
P. Hippo pathway - brief overview of its rele- mashita T, Kuo B and Zuo J. In vivo interplay
vance in cancer. J Physiol Pharmacol 2017; between p27(Kip1), GATA3, ATOH1, and
68: 311-335. POU4F3 converts non-sensory cells to hair
[105] Mueller KA, Glajch KE, Huizenga MN, Wilson
cells in adult mice. Cell Rep 2017; 19: 307-
RA, Granucci EJ, Dios AM, Tousley AR, Iuliano
320.
M, Weisman E, LaQuaglia MJ, DiFiglia M, Keg-
[109] Zhang S, Zhang Y, Dong Y, Guo L, Zhang Z,
el-Gleason K, Vakili K and Sadri-Vakili G. hippo
signaling pathway dysregulation in human Shao B, Qi J, Zhou H, Zhu W, Yan X, Hong G,
huntington’s disease brain and neuronal stem Zhang L, Zhang X, Tang M, Zhao C, Gao X and
cells. Sci Rep 2018; 8: 11355. Chai R. Knockdown of Foxg1 in supporting
[106] Poon CL, Mitchell KA, Kondo S, Cheng LY and cells increases the trans-differentiation of sup-
Harvey KF. The hippo pathway regulates neuro- porting cells into hair cells in the neonatal
blasts and brain size in drosophila melanogas- mouse cochlea. Cell Mol Life Sci 2020; 77:
ter. Curr Biol 2016; 26: 1034-1042. 1401-1419.
35 Am J Stem Cells 2020;9(3):25-35You can also read

















































