Aldosterone Induces Vascular Insulin Resistance by Increasing Insulin-Like Growth Factor-1 Receptor and Hybrid Receptor
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Aldosterone Induces Vascular Insulin Resistance by
Increasing Insulin-Like Growth Factor-1 Receptor and
Hybrid Receptor
Shamshad J. Sherajee, Yoshiko Fujita, Kazi Rafiq, Daisuke Nakano, Hirohito Mori, Tsutomu Masaki,
Taiga Hara, Masakazu Kohno, Akira Nishiyama, Hirofumi Hitomi
Objective—We previously showed that aldosterone induces insulin resistance in rat vascular smooth muscle cells (VSMCs).
Because insulin-like growth factor-1 receptor (IGF1R) affects insulin signaling, we hypothesized that aldosterone induces
vascular insulin resistance and remodeling via upregulation of IGF1R and its hybrid insulin/insulin-like growth
factor-1 receptor.
Methods and Results—Hybrid receptor expression was measured by immunoprecipitation. Hypertrophy of VSMCs was
evaluated by 3H-labeled leucine incorporation. Aldosterone (10 nmol/L) significantly increased protein and mRNA
expression of IGF1R and hybrid receptor in VSMCs but did not affect insulin receptor expression. Mineralocorticoid
receptor blockade with eplerenone inhibited aldosterone-induced increases in IGF1R and hybrid receptor. Aldosterone
augmented insulin (100 nmol/L)-induced extracellular signal-regulated kinase 1/2 phosphorylation. Insulin-induced
leucine incorporation and ␣-smooth muscle actin expression were also augmented by aldosterone in VSMCs. These
aldosterone-induced changes were significantly attenuated by eplerenone or picropodophyllin, an IGF1R inhibitor.
Chronic infusion of aldosterone (0.75 g/hour) increased blood pressure and aggravated glucose metabolism in rats.
Expression of hybrid receptor, azan-positive area, and oxidative stress in aorta was increased in aldosterone-infused rats.
Spironolactone and tempol prevented these aldosterone-induced changes.
Conclusion—Aldosterone induces vascular remodeling through IGF1R- and hybrid receptor– dependent vascular
insulin resistance. Mineralocorticoid receptor blockade may attenuate angiopathy in hypertensive patients with
hyperinsulinemia. (Arterioscler Thromb Vasc Biol. 2012;32:257-263.)
Key Words: insulin resistance 䡲 reactive oxygen species 䡲 receptors 䡲 signal transduction 䡲 aldosterone
I nsulin resistance is a major attribute of type 2 diabetes
mellitus and metabolic syndrome.1 Cardiovascular compli-
cations are often seen in these patients, and vascular insulin
in VSMCs. However, the precise molecular mechanisms respon-
sible for aldosterone-induced VSMC insulin resistance and
proatherogenic changes have not been identified.
resistance is considered to be involved in proatherogenic Insulin induces various actions, such as glucose metabolism
changes and subsequent cardiovascular events.2 There is grow- and normal cell physiology, by binding to the insulin receptor
ing interest in the role of aldosterone and its receptor, mineralo- (IR).8 The VSMCs express not only IR but also insulin-like
corticoid receptor (MR), in the pathogenesis of insulin resis- growth factor-1 receptor (IGF1R).8 Compared with IR, IGF1R is
tance.3 For instance, clinical studies have shown that patients more abundant in VSMCs, and expression of IGF1R is increased
with primary aldosteronism exhibit impaired glucose tolerance.4 in aortas of diabetic animals.9 Furthermore, subunits of IR and
Some possible mechanisms of insulin resistance induced by IGF1R easily form hybrid receptors, depending on the ratio of
aldosterone have been considered, such as a low serum potas- these receptors’ expressions.10 These hybrid receptors function
sium concentration, and direct effects of aldosterone on insulin more like IGF1R in that they have higher affinity for insulin-like
signaling.5,6 Previously, we have demonstrated that aldosterone growth factor-1 (IGF1) than to insulin.11,12 IGF1 induces hyper-
induces insulin resistance in rat vascular smooth muscle cells trophic changes and insulin resistance via IGF1R in vasculature.
(VSMCs) via the downregulation of insulin receptor substrate-1, Although the affinity of IGF1R for insulin is very low compared
a key molecule of insulin signaling pathway.7 Our data also with IGF1, high concentrations of insulin, which are often seen
clearly showed that aldosterone attenuates glucose metabolism in patients with insulin resistance, may affect intracellular
Received on: April 1, 2011; final version accepted on: December 2, 2011.
From the Departments of Pharmacology (S.J.S., K.R., D.N., A.N., H.H.), Cardiorenal and Cerebrovascular Medicine (Y.F., T.H., M.K.), and
Gastroenterology and Neurology (H.M., T.M.), Faculty of Medicine, Kagawa University, Kagawa, Japan.
Dr Sherajee and Dr Fujita contributed equally to this work.
Correspondence to Hirofumi Hitomi, MD, PhD, Department of Pharmacology, Faculty of Medicine, Kagawa University, 1750-1 Ikenobe, Miki-cho,
Kita-gun, Kagawa 761-0793, Japan. E-mail hitomi@kms.ac.jp
© 2011 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol is available at http://atvb.ahajournals.org DOI: 10.1161/ATVBAHA.111.240697
257258 Arterioscler Thromb Vasc Biol February 2012
Figure 1. Effect of aldosterone on insulin-like growth factor-1 receptor (IGF1R) expression in vascular smooth muscle cells (VSMCs). A,
VSMCs were incubated with the indicated concentrations of aldosterone for 48 hours. B and C, VSMCs were incubated with 10 nmol/L
aldosterone for the indicated times. A and B, Western blotting was performed with anti-IGF1R-subunit or anti--actin antibody. C,
IGF1R-subunit mRNA was measured by reverse transcription–polymerase chain reaction. A to C, Data represent mean⫾SE (n⫽4),
expressed as fold change compared with unstimulated cells. *P⬍0.05 vs control VSMCs.
signaling pathway via IGF1R or hybrid receptors.13 However, overnight incubation with anti-IR-subunit antibody, followed by
the roles of IGF1R and hybrid receptors in insulin resistance are immunoblot analysis with an antibody against IGF1R-subunit.8,9
not well understood.
Real-Time Reverse Transcription-Polymerase
In the present study, we hypothesized that aldosterone
Chain Reaction
induces insulin resistance via upregulation of IGF1R and its
The mRNA expressions of glyceraldehyde-3-phosphate dehydrogenase
hybrid receptors. To test this hypothesis, VSMCs were used (GAPDH), type I and type III collagen, serum and glucocorticoid-
to examine the effects of aldosterone on expression of these regulated kinase 1 (Sgk1), and IGF1R were analyzed by real-time
receptors. We also examined the hypertrophic effect of polymerase chain reaction, as described previously.16 –18 Primer se-
insulin in aldosterone-treated VSMCs. To confirm these in quences for GAPDH, Sgk1, type I and type III collagen, and IGF1R
were described previously.16,17 All data were normalized against
vitro findings in in vivo experimental models, insulin signal-
GAPDH mRNA expression.
ing pathways and vascular remodeling were also evaluated in
aortas of aldosterone-infused rats. [3H]Leucine Incorporation and ␣-Smooth Muscle
Actin Expression
Methods Protein synthesis was evaluated by measurement of [3H]leucine
For a detailed description of the procedures, see the expanded incorporation.19 Expression of ␣-smooth muscle actin was deter-
Methods section in the Supplemental Data, available online at mined using immunofluorescence, as previously reported.16
http://atvb.ahajournals.org.
Oral Glucose Tolerance Test
Cell Culture Glucose metabolism was evaluated by oral glucose tolerance test, as
The VSMCs were isolated from thoracic aortas of 4-week-old male previously reported.20
Sprague-Dawley rats (CLEA Japan, Shizuoka, Japan) by enzymatic
digestion as previously described.14 The VSMCs were used from the Histological Examination
third to eighth passages. Aortic fibrosis was evaluated by the percentage of azan-positive area
in 5 randomly selected microscope fields (⫻200) in each section.
Animal Preparation Medial thickness and lumen ratio were measured using a previously
All experimental procedures were performed according to the described method.21
guidelines for the care and use of animals established by Kagawa
University. Sprague-Dawley rats (male, 4 weeks old) were randomly Statistical Analysis
treated with 1 of the following combinations for 6 weeks: group 1, Results are expressed as mean⫾SE. Statistical significance was
tap water (control; n⫽6); group 2, 1% NaCl plus 1% KCl in drinking assessed using ANOVA and the Bonferroni post hoc test. Student t
water (salt; n⫽6); group 3, aldosterone (Aldo, 0.75 g/hour, SC; tests were performed to compare the means when the experimental
n⫽6); group 4, 1% NaCl plus 1% KCl plus aldosterone (Aldo⫹salt; design was comprised of 2 individual groups. A value of P⬍0.05
n⫽10); group 5, 1% NaCl plus 0.2% KCl plus aldosterone plus was considered statistically significant.
spironolactone, an MR antagonist (200 mg/kg per day, PO; n⫽8);
group 6, 1% NaCl plus 1% KCl plus aldosterone plus tempol, a Results
superoxide dismutase mimetic (Aldo⫹salt⫹Temp, 3 mmol/L in drink-
ing water; n⫽8); group 7, 1% NaCl plus 1% KCl plus aldosterone plus In Vitro Study
hydralazine, a nonspecific vasodilator (Aldo⫹salt⫹hydralazine, 25 Effects of Aldosterone on IGF1R, IR, and Hybrid
mg/kg per day by gavage; n⫽8).
Receptor Expressions in VSMCs
Aldosterone increased IGF1R protein expression in a dose-
Western Blot Analysis and Immunoprecipitation dependent manner, with a significant increase at 10 nmol/L
Protein samples from VSMCs and aorta were extracted, as previ-
ously described.14,15 Hybrid receptor expression was determined by (Figure 1A), whereas IR protein expression was not affected
coimmunoprecipitation and Western blotting. Protein samples from (Supplemental Figure IA and IB). Aldosterone (10 nmol/L)
VSMCs or aorta (600 g of protein) were immunoprecipitated by induced a time-dependent increase in IGF1R protein expression,Sherajee et al Aldosterone Induces Vascular Insulin Resistance 259
Figure 3. Effect of aldosterone on hypertrophic changes
induced by insulin in vascular smooth muscle cells (VSMCs).
VSMCs were pretreated with eplerenone (10 mol/L) for 30 min-
utes and then incubated with aldosterone (10 nmol/L) or vehicle
for 48 hours. Subsequently, VSMCs were treated with vehicle or
insulin (2–100 nmol/L) for 24 hours. A, Protein synthesis was
measured by [3H]leucine incorporation. B, Expression of
Figure 2. Effect of aldosterone on hybrid receptor expression in ␣-smooth muscle actin (␣-SMA) was determined using immuno-
vascular smooth muscle cells (VSMCs). A, VSMCs were incu- fluorescence. A and B, Data represent mean⫾SE (n⫽4),
bated with the indicated concentrations of aldosterone for 48 expressed as fold change compared with unstimulated cells.
hours. B, VSMCs were pretreated with eplerenone (10 mol/L) *P⬍0.05 vs control VSMCs, #P⬍0.05 vs aldosterone⫹insulin.
for 30 minutes and then exposed to 10 nmol/L aldosterone or
vehicle for 48 hours. A and B, VSMCs were lysed and immuno-
precipitated (IP) with anti-insulin receptor (IR) -subunit anti- completely inhibited by pretreatment with eplerenone, sug-
body. Western blot analysis was performed using insulin-like gesting that these responses are mediated by MR.
growth factor-1 receptor (IGF1R) -subunit or IR-subunit anti-
body. Data represent mean⫾SE (n⫽4), expressed as fold Effect of Aldosterone on Hypertrophic Changes Induced
change compared with unstimulated cells. *P⬍0.05 vs control by Insulin in VSMCs
VSMCs, #P⬍0.05 vs aldosterone alone. VSMCs pretreated with aldosterone showed the concentration-
dependent effects of insulin (100 nmol/L, 5 minutes) on extra-
with a significant effect occurring at 48 hours (Figure 1B). To cellular signal–regulated kinase 1/2 phosphorylation (Supple-
clarify the genomic effect of aldosterone, we measured the effect mental Figure IIIA), indicating that aldosterone augmented
of aldosterone on IGF1R mRNA expression. Aldosterone treat- insulin-induced mitogen-activated protein kinase phosphoryla-
ment at 10 nmol/L resulted in a significant increase in IGF1R tion. Aldosterone treatment attenuated Akt phosphorylation
mRNA expression after 3 hours, and expression continued to induced by insulin (Supplemental Figure IIIB).
increase for up to 72 hours (Figure 1C). Next, we measured leucine incorporation as an index of
Next, we examined the effect of aldosterone on hybrid protein synthesis in VSMCs. Insulin at different concentrations
receptor expression using immunoprecipitation in VSMCs. (0 –100 nmol/L) for 24 hours showed no effect on leucine
Aldosterone treatment increased hybrid receptor expression, incorporation (Figure 3A). However, pretreatment with aldoste-
with maximal effect at 10 nmol/L and 48 hours (Figure 2A). rone significantly augmented leucine incorporation induced by
The following experiments were therefore performed using insulin (100 nmol/L). These aldosterone-induced increases in
10 nmol/L aldosterone for 48 hours. leucine incorporation were completely inhibited by preincuba-
Effects of Eplerenone on Aldosterone-Induced Receptor tion with eplerenone, suggesting that insulin-induced protein
Expressions in VSMCs synthesis is augmented by aldosterone via MR. We also mea-
To investigate the role of MR on aldosterone-induced recep- sured ␣-smooth muscle actin expression, using immunofluores-
tor expressions, VSMCs were pretreated with eplerenone cence staining to confirm that aldosterone induces hypertrophic
(10 mol/L), a selective MR antagonist, for 30 minutes. changes in VSMC. Aldosterone increased insulin-induced
Aldosterone-induced IGF1R (Supplemental Figure IIA and ␣-smooth muscle actin expression, which had been diminished
IIB) and hybrid receptor (Figure 2B) expressions were by pretreatment with eplerenone (Figure 3B), suggesting that260 Arterioscler Thromb Vasc Biol February 2012
␣-smooth muscle actin expression induced by insulin is aug-
mented by pretreatment with aldosterone via MR.
Finally, we evaluated the effect of IGF1R on insulin-
induced protein synthesis in aldosterone-treated VSMCs. The
increased leucine incorporation was completely inhibited by
picropodophyllin (1 mmol/L), a selective IGF1R inhibitor
(Supplemental Figure IIIC); this suggests that aldosterone
augments protein synthesis induced by insulin via IGF1R. On
the other hand, IGF1 alone (0.1–100 ng/mL) increased
leucine incorporation (Supplemental Figure IIID).
In Vivo Study
Systolic Blood Pressure in Rats
The systolic blood pressure profile over time is shown in
Supplemental Figure IV. There were no significant differ-
ences in basal systolic blood pressure levels among the
groups. The systolic blood pressure of Aldo⫹salt-treated rats
was significantly higher than that in controls (213⫾6.0 versus
115⫾0.4 mm Hg). Treatment with spironolactone or tempol
normalized systolic blood pressure in these animals (117⫾0.8
and 122⫾0.9 mm Hg, respectively).
Glucose Metabolism in Rats
The oral glucose tolerance test was performed to analyze
whole-body insulin sensitivity, in which both blood glucose and
insulin concentration were measured and their respective areas
under the curve were calculated. Compared with controls, the
Aldo⫹salt rats showed higher glucose (Figure 4A) and insulin
(Supplemental Figure VA) levels after administration of oral
glucose. Spironolactone and tempol treatment significantly at-
Figure 4. Glucose metabolism. A, the oral glucose tolerance
tenuated the increase in glucose and insulin levels seen in the test was performed to analyze whole-body insulin sensitivity.
Aldo⫹salt group. The areas under the curve for blood glucose Aldosterone (Aldo)⫹salt-treated rats showed higher glucose lev-
(Figure 4B) and insulin (Supplemental Figure VB) were signif- els after administration of oral glucose. Spironolactone (Spl) and
tempol (Temp) treatment significantly attenuated increases in
icantly larger in Aldo⫹salt rats and were significantly reduced glucose level. B, AUC indicated area under the curve. Data rep-
by the spironolactone and tempol treatment. resent mean⫾SE (n⫽6 –10). *P⬍0.05 vs control rats, #P⬍0.05
Because glucose metabolism mediated by insulin is af- vs Aldo⫹salt–treated rats.
fected by potassium, we measured serum potassium concen-
tration. There was no difference between potassium concen- covered Akt phosphorylation. We also measured extracellular
tration in control (4.8⫾0.1 mmol/L) and Aldo⫹salt-treated signal–regulated kinase phosphorylation under ex vivo condi-
(5.3⫾0.3 mmol/L) rats. Spironolactone did not affect serum tions (Supplemental Figure VIIB). Insulin-induced extracellular
potassium concentrations (5.2⫾0.2 mmol/L). signal–regulated kinase phosphorylation was significantly aug-
mented in the Aldo⫹salt group compared with controls.
Oxidative Stress in Aortas
We previously reported that oxidative stress is involved in Fibrotic Changes and Collagen Synthesis in the Aorta
insulin resistance induced by aldosterone in cultured VSMCs.7 Fibrotic changes in aortas were assessed using azan staining.
We used dihydroethidium fluorescence to evaluate reactive Aldo⫹salt-treated aortas showed more azan-positive area
oxygen species (ROS) formation. Aldo⫹salt rats showed more than did controls that had been normalized with spironolac-
intense dihydroethidium staining than did controls; dihydro- tone and tempol treatment (Figure 5A). Medial thickness-to-
ethidium staining was significantly reduced in aortas of rats lumen diameter ratio, as an index of vascular hypertrophy,
treated with spironolactone or tempol (Supplemental Figure VI), was increased in Aldo⫹salt-treated aortas and was normal-
suggesting an effect by the MR on ROS formation. ized by spironolactone and tempol treatment (Figure 5B).
Next, we found that mRNA expressions of matrix protein
Insulin Signaling in the Aorta type I (Supplemental Figure VIIIA) and type III (Supplemen-
We measured Akt phosphorylation in aortas using insulin tal Figure VIIIB) collagens were significantly upregulated in
incubation under ex vivo conditions. Insulin (100 nmol/L, 30 Aldo⫹salt-treated aortas and were normalized by spironolac-
minutes) increased Akt phosphorylation in the control group tone and tempol treatment. Furthermore, mRNA of the MR
(Supplemental Figure VIIA). Insulin-induced Akt phosphoryla- target gene, Sgk1, was upregulated in Aldo⫹salt-treated
tion was significantly less in the Aldo⫹salt group compared aortas, and was normalized by spironolactone and tempol
with controls, indicating that insulin signaling was attenuated by treatment (Supplemental Figure VIIIC), suggesting that aldo-
Aldo⫹salt treatment. Spironolactone and tempol treatment re- sterone induces a hypertrophic effect via MR.Sherajee et al Aldosterone Induces Vascular Insulin Resistance 261
Involvement of Blood Pressure in
Aldosterone-Treated Rats
We examined the effect of blood pressure on metabolic
changes in aldosterone treatment. Hydralazine, a nonspecific
vasodilator, normalized blood pressure in Aldo⫹salt-treated
rats (Supplemental Figure XA). Lowering blood pressure by
hydralazine did not improve glucose metabolism in
Aldo⫹salt-treated rats (Supplemental Figure XB). Further-
more, hydralazine had no effect on IGF1 and hybrid receptor
expression (Supplemental Figure XC and XD).
Discussion
Aldosterone is the final mediator of the renin-angiotensin-aldo-
sterone system; it mediates blood pressure and electrolytic
balance. Increasing evidence has indicated that aldosterone is
also directly involved in tissue damage, proliferative and hyper-
trophic changes in the vasculature, and vascular insulin resis-
tance.7,22 Several clinical studies have shown that MR inhibition
attenuates cardiovascular complications of diabetes mellitus23,24;
however, the molecular mechanism of these protective effects
has not been elucidated. The present study showed that aldoste-
rone induces insulin resistance by upregulating IGF1R and
hybrid receptor and provides a possible molecular mechanism
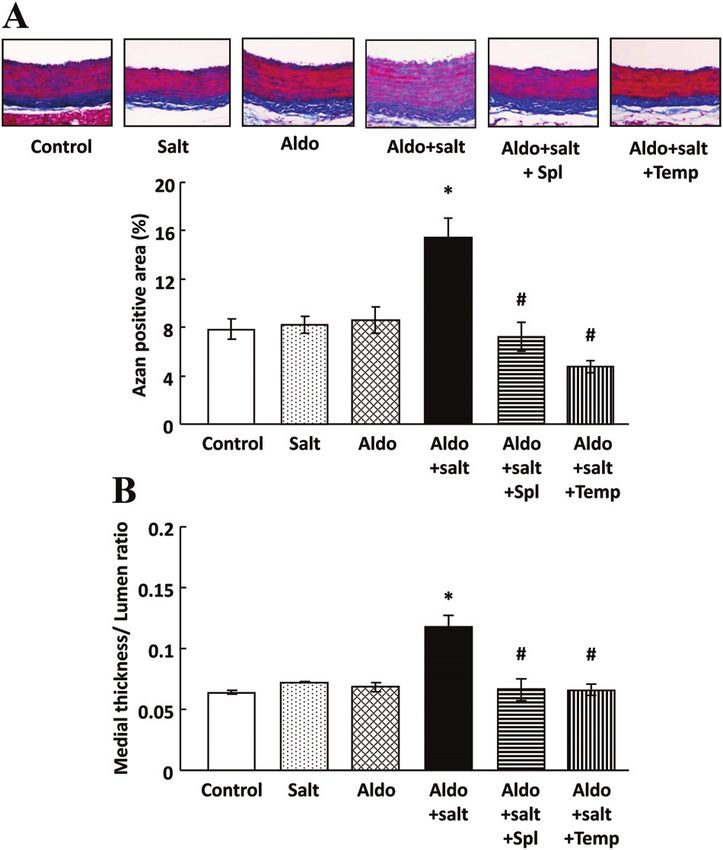
Figure 5. Fibrotic and hypertrophic changes in the aorta. A,
for atherosclerosis induced by insulin in presence of aldosterone.
Aortic fibrosis was evaluated by azan-positive area. Representa- Our study also showed that aldosterone treatment impaired
tive photographs of azan-stained aorta. B, Medial thickness and insulin signaling in the aorta and suppressed whole-body insulin
lumen ratio as an index of vascular hypertrophy was measured. sensitivity, accompanied by upregulation of IGF1R and its
A and B, Data represent mean⫾SE (n⫽6 –10). *P⬍0.05 vs con-
trol rats, #P⬍0.05 vs aldosterone (Aldo)⫹salt–treated rats. Spl hybrid receptor. This is novel information about the role of
indicates spironolactone; Temp, tempol. IGF1R in vasculature; it supports the idea that insulin resistance
by aldosterone affects the pathogenesis of vascular remodeling
in this model.
IGF1R and Hybrid Receptor in Aorta High concentrations of insulin may activate intracellular
Hybrid receptor expression was significantly increased in signaling pathway via IGF1R or hybrid receptors, although the
Aldo⫹salt-treated aorta and was normalized by spironolac- affinity of IGF1R for insulin is much lower than IGF1.13 In this
tone and tempol treatment (Figure 6A). Expression of IGF1R study, high concentration (100 nmol/L) of insulin did not affect
mRNA and protein were also significantly increased in aortic hypertrophic changes in VSMCs; however, pretreatment with
tissues of Aldo⫹salt-treated rats (Figure 6B and 6C). The aldosterone significantly increased mitogen-activated protein
IGF1R inhibitor picropodophyllin (1 mmol/L) improved Akt kinase activation and leucine incorporation, accompanied by
phosphorylation induced by insulin in Aldo⫹salt-treated rats increased levels of IGF1R and hybrid receptor. Pretreatment
(Supplemental Figure IX). with IGF1R inhibitor attenuated protein synthesis induced by a
Figure 6. Insulin-like growth factor-1 receptor (IGF1R) and hybrid receptor in aorta. A, Aortas were lysed and immunoprecipitated (IP)
with anti-insulin receptor (IR) -subunit antibody. Western blot analysis was performed using IGF1R-subunit or IR-subunit antibody.
B, IGF1R mRNA was measured by reverse transcription–polymerase chain reaction. C, Western blot analysis was performed using
IGF1R-subunit or -actin antibody. A to C, Data represent mean⫾SE (n⫽6 –10). *P⬍0.05 vs control rats, #P⬍0.05 vs aldosterone
(Aldo)⫹salt–treated rats. Spl indicates spironolactone; Temp, tempol.262 Arterioscler Thromb Vasc Biol February 2012
high insulin concentration, whereas a lower insulin concentra- signaling pathway and subsequently result in vascular remodel-
tion (2 nmol/L) had no effect on mitogen-activated protein ing via ROS.
kinase activation and protein synthesis. These results indicate The roles of IGF1R and hybrid receptor in VSMCs are still
that high concentrations of insulin induce hypertrophic actions unclear. Several studies have shown that IGF1 has potential
via upregulation of IGF1R or hybrid receptor. In addition, beneficial effects on vasculopathy and glucose metabolism in
previous reports also showed that plasma insulin levels were cardiovascular tissue,32–34 whereas IGF1 reportedly induces
significantly increased in type 2 diabetes mellitus patients vascular remodeling.35,36 Expression of IGF1R is much higher
compared with control subjects, and insulin levels were corre- than that of IR, and increased IGF1R forms hybrid receptors in
lated with decreased IRs and increased hybrid receptor propor- VSMCs.37 Recently, Cascella et al38 reported that aldosterone
tions.25 These data suggest that decreased IRs due to high plasma accelerates the development of atherosclerosis via IGF1-
insulin levels result in increased hybrid receptors. In the present mediated signaling in VSMCs. In addition, Engberding et al9
reported that aortas of diabetic mice expressed significantly
study, exogenous aldosterone increased expression of IGF1R
more IGF1R in all wall layers than did those of age-matched
mRNA and protein, but not IRs leading to hybrid receptor
controls. However, it is methodologically difficult to distinguish
formation, indicating that aldosterone increases hybrid receptor
the effects of IGF1R and IR, because both receptors partly share
mainly by IGF1R production.
a common downstream signaling pathway. Picropodophyllin, an
Glucose metabolism mediated by insulin is affected by IGF1R inhibitor, completely diminished leucine incorporation
serum potassium concentration. Hypokalemia is often ob- induced by insulin in aldosterone-treated VSMCs and improved
served in patients with primary aldosteronism4; it is a risk Akt phosphorylation in aorta of aldosterone-treated rats in this
factor for insulin resistance and glucose intolerance in these study, whereas previous studies showed that an small interfering
patients. In this study, drinking water for aldosterone-treated RNA gene knockdown of IGF1R increased glucose uptake,9
rats was supplemented with potassium chloride to prevent suggesting that IGF1R is involved in insulin resistance and
potential hypokalemia. In addition, there was no significant hypertrophic changes in vasculature. The present study showed
difference in serum potassium levels. Therefore, we consider that treatment with eplerenone significantly prevented increases
that the effect of potassium concentration on insulin action in IGF1R and hybrid receptor expression. In this regard, we
might be minimal in this study, and insulin resistance was recently showed that mechanical stretch also increased IGF1R
directly induced by aldosterone itself. On the other hand, expression in VSMCs,39 suggesting that high blood pressure
systemic glucose metabolism is predominantly regulated in may affect VSMC insulin resistance via IGF1R. On the other
skeletal muscle, adipose tissue, and liver.26 Therefore, glu- hand, the present study showed that nonspecific vasodilator has
cose metabolism in vasculature might not play a major role in no effect on glucose metabolism and IGF1R expression, indi-
systemic glucose metabolism. Recent studies have indicated cating that both MR activation and blood pressure are involved
that aldosterone impairs insulin and glucose metabolisms via in insulin resistance in vasculature.
downregulation of insulin receptor substrate-1 in adi- In summary, the present findings showed that the hyper-
pocytes.27 Furthermore, other studies also reported that MR trophic effect of insulin is augmented by aldosterone via
blockade improved systemic insulin sensitivity by changing upregulation of IGF1R and hybrid receptor, and the relevant
adiponectin, peroxisome proliferator–activated receptor-␥, signal transduction mechanism in VSMCs. Aldosterone in-
and proinflammatory adipokines in obese rats.28 These results duces cellular hypertrophy by activating the extracellular
suggest that aldosterone attenuated not only vascular insulin signal–regulated kinase 1/2 pathway in VSMCs, indicating
the potential role of aldosterone in the development of the
signaling but also systemic glucose metabolism.
vascular hypertrophy. In aldosterone-infused hypertensive
Previous experiments clarified that ROS affect the insulin
rats, hypertension, insulin resistance, vascular hypertrophy,
signaling pathway.29 We have also reported that aldosterone
and fibrosis are associated with increased vascular IGF1R
induces insulin receptor substrate-1 degradation and attenuates
and hybrid receptor expression. These changes were amelio-
glucose metabolism through the activation of ROS generation by rated by treatment with spironolactone and tempol, suggest-
NADPH oxidase.7 The present study shows that aldosterone ing that aldosterone augments vascular hypertrophic effect of
treatment increases ROS generation in the aorta, and treatment insulin through upregulation of IGF1R via an MR- and
with an antioxidant, tempol, improved glucose intolerance, as oxidative stress-mediated pathway. These data indicate that
evaluated by oral glucose tolerance test in aldosterone-treated MR inhibition may attenuate angiopathy in hypertensive
rats. Taken together, these facts indicate that aldosterone atten- patients with hyperinsulinemia or hypertensive diabetic pa-
uates vascular and systemic glucose metabolism by decreased tients who are treated with insulin. Therefore, IGF1R may be
insulin receptor substrate-1 and increased IGF1R via ROS a novel therapeutic target for insulin resistance and vascular
generation. However, the involvement of aldosterone in arterio- complications in hypertensive patients with diabetes mellitus.
sclerosis in diabetes mellitus is more complicated, because
aldosterone both directly affects proatherogenic changes via Acknowledgments
ROS30 and indirectly augments insulin-induced hypertrophy We are grateful to Pfizer Inc. (New York, NY) for supplying
through IGF1R-mediated hypertrophic signaling pathways, such eplerenone.
as mitogen-activated protein kinase activation. In addition, we
reported that ROS is involved in insulin resistance by shift of
Sources of Funding
This work was supported in part by grants-in-aid for scientific
insulin signaling from glucose metabolism into vascular remod- research from the Ministry of Education, Culture, Sports, Science
eling.31 Aldosterone-augmented IGF1R may affect the insulin and Technology of Japan (20590253 and 22790792).Sherajee et al Aldosterone Induces Vascular Insulin Resistance 263
Disclosures aldosterone and mineralocorticoid receptors in rat mesangial cell prolif-
eration and deformability. Hypertension. 2005;45:710 –716.
None.
20. Nagai Y, Ichihara A, Nakano D, Kimura S, Pelisch N, Fujisawa Y, Hitomi
H, Hosomi N, Kiyomoto H, Kohno M, Ito H, Nishiyama A. Possible con-
References tribution of the non-proteolytic activation of prorenin to the development of
1. Pravenec M, Kurtz TW. Molecular genetics of experimental hypertension insulin resistance in fructose-fed rats. Exp Physiol. 2009;94:1016–1023.
and the metabolic syndrome: from gene pathways to new therapies. 21. Fujii K, Umemoto S, Fujii A, Yonezawa T, Sakumura T, Matsuzaki M.
Hypertension. 2007;49:941–952. Angiotensin II type 1 receptor antagonist downregulates nonmuscle myosin
2. Schulman IH, Zhou MS. Vascular insulin resistance: a potential link heavy chains in spontaneously hypertensive rat aorta. Hypertension. 1999;
between cardiovascular and metabolic diseases. Curr Hypertens Rep. 33:975–980.
2009;11:48 –55. 22. Miyata K, Hitomi H, Guo P, Zhang GX, Kimura S, Kiyomoto H, Hosomi
3. Lastra-Lastra G, Sowers JR, Restrepo-Erazo K, Manrique-Acevedo C, N, Kagami S, Kohno M, Nishiyama A. Possible involvement of Rho-
Lastra-Gonzalez G. Role of aldosterone and angiotensin II in insulin kinase in aldosterone-induced vascular smooth muscle cell remodeling.
resistance: an update. Clin Endocrinol (Oxf). 2009;71:1– 6. Hypertens Res. 2008;31:1407–1413.
4. Conn JW. Hypertension, the potassium ion and impaired carbohydrate 23. Davies JI, Band M, Morris A, Struthers AD. Spironolactone impairs
tolerance. N Engl J Med. 1965;273:1135–1143. endothelial function and heart rate variability in patients with type 2
5. Giacchetti G, Ronconi V, Turchi F, Agostinelli L, Mantero F, Rilli S, diabetes. Diabetologia. 2004;47:1687–1694.
Boscaro M. Aldosterone as a key mediator of the cardiometabolic syndrome 24. O’Keefe JH, Abuissa H, Pitt B. Eplerenone improves prognosis in post-
in primary aldosteronism: an observational study. J Hypertens. 2007;25: myocardial infarction diabetic patients with heart failure: results from
177–186. EPHESUS. Diabetes Obes Metab. 2008;10:492– 497.
6. Shimamoto K, Shiiki M, Ise T, Miyazaki Y, Higashiura K, Fukuoka M, 25. Federici M, Porzio O, Zucaro L, Giovannone B, Borboni P, Marini MA,
Hirata A, Masuda A, Nakagawa M, Iimura O. Does insulin resistance Lauro D, Sesti G. Increased abundance of insulin/IGF-I hybrid receptors in
participate in an impaired glucose tolerance in primary aldosteronism? J adipose tissue from NIDDM patients. Mol Cell Endocrinol. 1997;135:41–47.
Hum Hypertens. 1994;8:755–759. 26. Koopmans SJ, Mandarino L, DeFronzo RA. Time course of insulin action
7. Hitomi H, Kiyomoto H, Nishiyama A, Hara T, Moriwaki K, Kaifu K, on tissue-specific intracellular glucose metabolism in normal rats. Am J
Ihara G, Fujita Y, Ugawa T, Kohno M. Aldosterone suppresses insulin Physiol. 1998;274:E642–E650.
signaling via the downregulation of insulin receptor substrate-1 in 27. Wada T, Ohshima S, Fujisawa E, Koya D, Tsuneki H, Sasaoka T. Aldoste-
vascular smooth muscle cells. Hypertension. 2007;50:750 –755. rone inhibits insulin-induced glucose uptake by degradation of insulin
8. Johansson GS, Arnqvist HJ. Insulin and IGF-I action on insulin receptors, receptor substrate (IRS) 1 and IRS2 via a reactive oxygen species-mediated
IGF-I receptors, and hybrid insulin/IGF-I receptors in vascular smooth pathway in 3T3–L1 adipocytes. Endocrinology. 2009;150:1662–1669.
muscle cells. Am J Physiol Endocrinol Metab. 2006;291:E1124 –E1130. 28. Guo C, Ricchiuti V, Lian BQ, Yao TM, Coutinho P, Romero JR, Li J,
9. Engberding N, San Martin A, Martin-Garrido A, Koga M, Pounkova L, Williams GH, Adler GK. Mineralocorticoid receptor blockade reverses
Lyons E, Lassegue B, Griendling KK. Insulin-like growth factor-1 obesity-related changes in expression of adiponectin, peroxisome
receptor expression masks the antiinflammatory and glucose uptake proliferator-activated receptor-␥, and proinflammatory adipokines.
capacity of insulin in vascular smooth muscle cells. Arterioscler Thromb Circulation. 2008;117:2253–2261.
Vasc Biol. 2009;29:408 – 415. 29. Taniyama Y, Hitomi H, Shah A, Alexander RW, Griendling KK. Mech-
10. Moxham CP, Duronio V, Jacobs S. Insulin-like growth factor I receptor anisms of reactive oxygen species-dependent downregulation of insulin
-subunit heterogeneity: evidence for hybrid tetramers composed of receptor substrate-1 by angiotensin II. Arterioscler Thromb Vasc Biol.
insulin-like growth factor I and insulin receptor heterodimers. J Biol 2005;25:1142–1147.
Chem. 1989;264:13238 –13244. 30. Ishizawa K, Izawa Y, Ito H, Miki C, Miyata K, Fujita Y, Kanematsu Y,
11. Frattali AL, Pessin JE. Relationship between ␣ subunit ligand occupancy Tsuchiya K, Tamaki T, Nishiyama A, Yoshizumi M. Aldosterone stim-
and  subunit autophosphorylation in insulin/insulin-like growth factor-1 ulates vascular smooth muscle cell proliferation via big mitogen-activated
hybrid receptors. J Biol Chem. 1993;268:7393–7400. protein kinase 1 activation. Hypertension. 2005;46:1046 –1052.
12. Arnqvist HJ. The role of IGF-system in vascular insulin resistance. Horm 31. Hitomi H, Kaifu K, Fujita Y, Sofue T, Nakano D, Moriwaki K, Hara T,
Metab Res. 2008;40:588 –592. Kiyomoto H, Kohno M, Kobori H, Nishiyama A. Angiotensin II shifts
13. Schumacher R, Mosthaf L, Schlessinger J, Brandenburg D, Ullrich A. Insulin insulin signaling into vascular remodeling from glucose metabolism in
and insulin-like growth factor-1 binding specificity is determined by distinct vascular smooth muscle cells. Am J Hypertens. 2011;24:1149 –1155.
regions of their cognate receptors. J Biol Chem. 1991;266:19288–19295. 32. Abbas A, Grant PJ, Kearney MT. Role of IGF-1 in glucose regulation and
14. Hitomi H, Fukui T, Moriwaki K, Matsubara K, Sun GP, Rahman M, cardiovascular disease. Expert Rev Cardiovasc Ther. 2008;6:1135–1149.
Nishiyama A, Kiyomoto H, Kimura S, Ohmori K, Abe Y, Kohno M. 33. Imrie H, Abbas A, Viswambharan H, Rajwani A, Cubbon RM, Gage M,
Synergistic effect of mechanical stretch and angiotensin II on superoxide Kahn M, Ezzat VA, Duncan ER, Grant PJ, Ajjan R, Wheatcroft SB,
production via NADPH oxidase in vascular smooth muscle cells. Kearney MT. Vascular insulin-like growth factor-I resistance and diet-
J Hypertens. 2006;24:1089 –1095. induced obesity. Endocrinology. 2009;150:4575– 4582.
15. Nishiyama A, Kobori H, Fukui T, Zhang GX, Yao L, Rahman M, Hitomi 34. Ren J, Samson WK, Sowers JR. Insulin-like growth factor I as a cardiac
H, Kiyomoto H, Shokoji T, Kimura S, Kohno M, Abe Y. Role of hormone: physiological and pathophysiological implications in heart
angiotensin II and reactive oxygen species in cyclosporine A-dependent disease. J Mol Cell Cardiol. 1999;31:2049 –2061.
hypertension. Hypertension. 2003;42:754 –760. 35. Bornfeldt KE, Raines EW, Nakano T, Graves LM, Krebs EG, Ross R.
16. Diah S, Zhang GX, Nagai Y, Zhang W, Gang L, Kimura S, Hamid MR, Insulin-like growth factor-I and platelet-derived growth factor-BB induce
Tamiya T, Nishiyama A, Hitomi H. Aldosterone induces myofibroblastic directed migration of human arterial smooth muscle cells via signaling
transdifferentiation and collagen gene expression through the Rho-kinase pathways that are distinct from those of proliferation. J Clin Invest. 1994;
dependent signaling pathway in rat mesangial cells. Exp Cell Res. 2008;314: 93:1266–1274.
3654–3662. 36. Gockerman A, Prevette T, Jones JI, Clemmons DR. Insulin-like growth
17. Du J, Fan YY, Hitomi H, Kiyomoto H, Kimura S, Kong CZ, Noma T, factor (IGF)-binding proteins inhibit the smooth muscle cell migration
Kohno M, Nishiyama A, Nakano D. Mineralocorticoid receptor blockade responses to IGF-I and IGF-II. Endocrinology. 1995;136:4168 – 4173.
and calcium channel blockade have different renoprotective effects on 37. Chisalita SI, Arnqvist HJ. Expression and function of receptors for
glomerular and interstitial injury in rats. Am J Physiol Renal Physiol. insulin-like growth factor-I and insulin in human coronary artery smooth
2009;297:F802–F808. muscle cells. Diabetologia. 2005;48:2155–2161.
18. Nagai Y, Yao L, Kobori H, Miyata K, Ozawa Y, Miyatake A, Yukimura T, 38. Cascella T, Radhakrishnan Y, Maile LA, Busby WH Jr, Gollahon K,
Shokoji T, Kimura S, Kiyomoto H, Kohno M, Abe Y, Nishiyama A. Tem- Colao A, Clemmons DR. Aldosterone enhances IGF-I-mediated signaling
porary angiotensin II blockade at the prediabetic stage attenuates the devel- and biological function in vascular smooth muscle cells. Endocrinology.
opment of renal injury in type 2 diabetic rats. J Am Soc Nephrol. 2005;16: 2010;151:5851–5864.
703–711. 39. Liu G, Hitomi H, Hosomi N, Lei B, Nakano D, Deguchi K, Mori H, Masaki
19. Nishiyama A, Yao L, Fan Y, Kyaw M, Kataoka N, Hashimoto K, Nagai T, Ma H, Griendling KK, Nishiyama A. Mechanical stretch augments
Y, Nakamura E, Yoshizumi M, Shokoji T, Kimura S, Kiyomoto H, insulin-induced vascular smooth muscle cell proliferation by insulin-like
Tsujioka K, Kohno M, Tamaki T, Kajiya F, Abe Y. Involvement of growth factor-1 receptor. Exp Cell Res. 2011;317:2420–2428.You can also read

















































