Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak Dead Wood: An Observation Aided by Squirrels
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

Article
Invertebrate Assemblages on Biscogniauxia Sporocarps on Oak
Dead Wood: An Observation Aided by Squirrels
Yu Fukasawa
Graduate School of Agricultural Science, Tohoku University, 232-3 Yomogida, Naruko, Osaki,
Miyagi 989-6711, Japan; fukasawayuu@gmail.com; Tel.: +81-229-847-397; Fax: +81-229-846-490
Abstract: Dead wood is an important habitat for both fungi and insects, two enormously diverse
groups that contribute to forest biodiversity. Unlike the myriad of studies on fungus–insect rela-
tionships, insect communities on ascomycete sporocarps are less explored, particularly for those
in hidden habitats such as underneath bark. Here, I present my observations of insect community
dynamics on Biscogniauxia spp. on oak dead wood from the early anamorphic stage to matured
teleomorph stage, aided by the debarking behaviour of squirrels probably targeting on these fungi.
In total, 38 insect taxa were observed on Biscogniauxia spp. from March to November. The com-
munity composition was significantly correlated with the presence/absence of Biscogniauxia spp.
Additionally, Librodor (Glischrochilus) ipsoides, Laemophloeus submonilis, and Neuroctenus castaneus were
frequently recorded and closely associated with Biscogniauxia spp. along its change from anamorph
to teleomorph. L. submonilis was positively associated with both the anamorph and teleomorph
stages. L. ipsoides and N. castaneus were positively associated with only the teleomorph but not with
the anamorph stage. N. castaneus reproduced and was found on Biscogniauxia spp. from June to
November. These results suggest that sporocarps of Biscogniauxia spp. are important to these insect
taxa, depending on their developmental stage.
Citation: Fukasawa, Y. Invertebrate
Keywords: fungivory; insect–fungus association; Sciurus lis; Quercus serrata; xylariaceous ascomycetes
Assemblages on Biscogniauxia
Sporocarps on Oak Dead Wood: An
Observation Aided by Squirrels.
Forests 2021, 12, 1124. https://
doi.org/10.3390/f12081124 1. Introduction
Dead wood is an essential component of biodiversity in forest ecosystems [1,2]. Fungi,
Academic Editor: Simon Curling in particular, is a major group of saproxylic communities; they have a large impact on
saproxylic communities due to their unique wood decay abilities [3], and their fruit bodies
Received: 24 July 2021 and spores are important to the diet of a variety of organisms, including protozoa [4],
Accepted: 20 August 2021 invertebrates [5], and vertebrates [6,7]. A better understanding of the relationships between
Published: 22 August 2021
fungi and saproxylic communities is critical to clarifying the mechanisms that maintain
biodiversity in forest ecosystems.
Publisher’s Note: MDPI stays neutral In terms of their diversity and function, insects are another major group present within
with regard to jurisdictional claims in
saproxylic communities [1]. Insects have intimate relationships with fungi as fungivores,
published maps and institutional affil-
vectors of fungal propagules, and foragers of wood decomposed by fungi [8]. Numer-
iations.
ous studies have investigated the insect communities present on fungal fruit bodies, the
majority of which are basidiomycetes [5,9,10]. Host specificity [11–14], the evolution of
host use [15], and spore dispersal [16–20] have been intensively studied for a variety of
fungus–insect relationships. Furthermore, topics in general ecology (e.g., coexisting pat-
Copyright: © 2021 by the author. terns on patchy resources [21,22]) and applied ecology (e.g., effects of forest management
Licensee MDPI, Basel, Switzerland. on ecological communities [23,24]) have also been investigated using the insect–basidiocarp
This article is an open access article
system. However, studies on the relationships between insect communities on ascomycete
distributed under the terms and
fruit bodies, a sister taxon of basidiomycetes which also produces macroscopic fruit bodies,
conditions of the Creative Commons
are quite limited, with examples of symbiotic associations in ambrosia beetles [25], wood-
Attribution (CC BY) license (https://
wasps [26], and fungus-growing termites [27] and insect pathogens, such as in the genera
creativecommons.org/licenses/by/
Beauveria [28], Metarhizium [29], and Ophiocordyceps [30].
4.0/).
Forests 2021, 12, 1124. https://doi.org/10.3390/f12081124 https://www.mdpi.com/journal/forests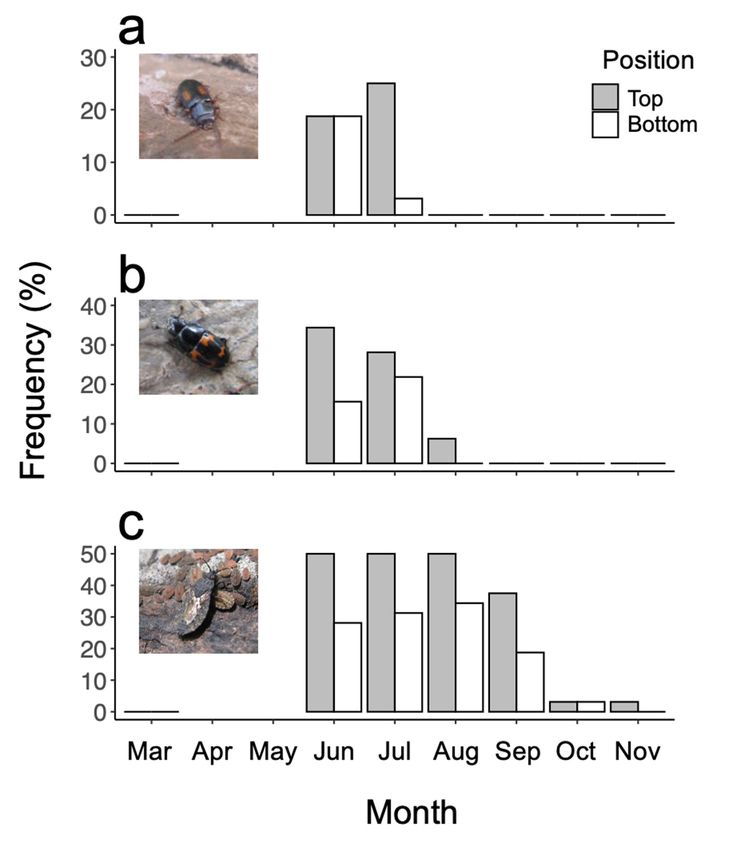
Forests 2021, 12, 1124 2 of 10
Quercus serrata is a deciduous oak that dominates low-elevation rural forests through-
out Japan. In recent decades, a serious dieback of Q. serrata (oak wilt disease) resulted
in huge amount of dead wood [31,32]. Therefore, evaluations of the biotic communities
associated with Q. serrata dead wood is necessary to understand and predict biodiversity
in areas affected by oak wilt disease. The symbiotic association between a pathogenic
fungus, Raffaerea quercivora, and ambrosia beetles, Platypus querucivorus, which carry the
propagules of the fungus in their mycangia, is well studied [33,34]. In addition, since
Q. serrata logs are traditionally used for cultivating shiitake mushrooms (Lentinula edodes),
several ascomycete species that negatively affect the yield of shiitake mushrooms are
known to occur on Q. serrata bed-logs [35]. The most famous ascomycete species is in the
genus Trichoderma (including species formerly denoted as Hypocrea), which is an antago-
nistic and/or mycoparasitic taxon and causes serious damage to shiitake cultivation [36].
Trichoderma spp. has an intimate relationship with gall midge belonging to the genus
Camptomyia on shiitake bed-logs [37]. Despite recent advances in the studies of microscopic
and pathogenic ascomycetes associated with insects, relationships between insects and
other taxa in macroscopic ascomycetes in genera such as Biscogniauxia, Daldinia, Diatrype,
Graphostroma, and Hypoxylon, which fruit on Q. serrata dead wood, are largely unknown.
To investigate successive changes of fungal and insect communities and their inter-
actions during decomposition of Q. serrata dead wood, I started a multi-year survey of
32 experimentally cut logs of Q. serrata in August 2015. In March 2016, I observed that a
squirrel or squirrels frequently visited and intensively debarked the logs, where colonies of
anamorphic ascomycetes were found to appear on the sapwood. Subsequently, a variety
of insects were found on the ascomycetes throughout their changes from the anamorph
to the teleomorph stage. In the present study, I describe the insect assemblages that were
observed on the Q. serrata logs and their relationships with the different sexual stages of
ascomycetes during a growing season.
2. Materials and Methods
2.1. Experimental Setup
The present study was conducted in a secondary forest dominated by Q. serrata and
Pinus densiflora in Kami town, Miyagi, Japan (38◦ 37.2 N, 140◦ 48.6 E). In 2016, the mean an-
nual temperature at the nearest meteorological station at Kawatabi (38◦ 44.60 N, 140◦ 45.6 E)
was 11.6 ◦ C (3.2 ◦ C in February to 28.7 ◦ C in August), and the annual precipitation was
1537.5 mm. Snow covers the ground from November to March (Japan Meteorological
Agency, available online: https://www.jma.go.jp/jma/indexe.html (accessed on 21 Au-
gust 2021)). In August 2015, two Q. serrata trees were felled and cut into 32 logs with a
length of 1 m each (diameter 3.4–24.5 cm). The logs were laid on the ground approximately
1 m from each other. I began observations of the logs’ surfaces (top and bottom) in March
2016, when the ground was still covered with snow, but the tops of the logs were visible.
On 10 March 2016, at 6:30 a.m., I observed that a squirrel (Sciurus lis) approached
the logs and tore off the bark using its teeth. I found Geniculosporium type anamorph of
ascomycete on the sapwood surface where the bark had been removed by the squirrel
(Figure 1a). The surface of the anamorph was scratched overall (Figure 1a,b). I found
76 portions of debarking by the squirrel on 15 out of the 32 logs (in 1–10 portions per
log). The frequency of the presence of anamorphs on the debarked portion was 100%. The
observations continued once per week until the end of November 2016, except for April
and May. The frequency of squirrel debarking increased to over 50% (19/32 logs) in July
2016. From March to July, the anamorph layers gradually peeled off and teleomorphs
appeared (Figure 1c,d), which were identified as Biscogniauxia maritima and Biscogniauxia
plana by Dr. Shuhei Takemoto from the University of Tokyo.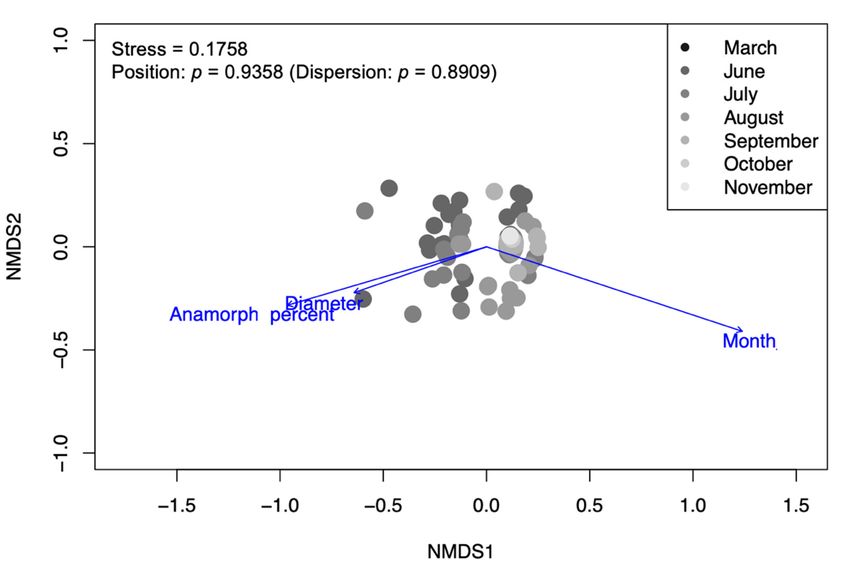
Forests
Forests2021,
2021,12,
12,x1124
FOR PEER REVIEW 3 3ofof10
10
Figure 1. (a) Surface of a Quercus serrata log recently debarked by a squirrel. Powdery ascomycete
Figure 1. (a) Surface of a Quercus serrata log recently debarked by a squirrel. Powdery ascomycete
anamorphappears
anamorph appearsatatthethecentre
centre
of of
thethe debarked
debarked portion
portion where
where numerous
numerous scratch
scratch scarsscars
can becanob-
be
observed,
served, MarchMarch
2016.2016. (b) Zoomed
(b) Zoomed in picture
in picture of the scratched
of the scratched anamorph anamorph surface,
surface, March March
2016. 2016.
(c) Grad-
(c) peeling
ual Gradualofpeeling of the anamorph
the anamorph surfaceportion)
surface (brown (brown portion) and appearance
and appearance of the teleomorph
of the teleomorph surface
(grey portion),
surface June 2016.
(grey portion), (d)2016.
June Zoomed in picture
(d) Zoomed inof teleomorph,
picture which was
of teleomorph, identified
which as Biscog-
was identified as
niauxia maritima
Biscogniauxia and Biscogniauxia
maritima plana by
and Biscogniauxia Dr. by
plana Shuhei Takemoto
Dr. Shuhei from the
Takemoto University
from of Tokyo,
the University July
of Tokyo,
2016. Scale Scale
July 2016. bars: 5bars:
mm.5 mm.
2.2.Data
2.2. DataCollection
Collection
Debarked,anamorph,
Debarked, anamorph,and andteleomorph
teleomorphareas areaswere
weremeasured
measuredevery
everymonth
month(except
(exceptfor for
Apriland
April andMay)
May)by byplacing
placing11cmcmgrid
gridsquares
squareson onthethelog
logsurface
surfaceandandcounting
countingthethenumber
number
ofgrid
of gridsquares
squaresthat
thathad
haddebarkation,
debarkation,anamorphs,
anamorphs,and andteleomorphs.
teleomorphs.
Insects that were observed on the log surface
Insects that were observed on the log surface were recordedwere recordedevery
everyweek
week(except
(exceptfor for
April and May) as binary data (presence/absence on each log). Insects
April and May) as binary data (presence/absence on each log). Insects were identified with were identified
with reference
reference to thetokeys
the keys
and and nomenclature
nomenclature of Kurosawa
of Kurosawa et al.
et al. [38],
[38], Uenoetetal.al.[39],
Ueno [39],and
and
Hayashi et al. [40] for Coleoptera; Ishikawa et al. [41] for Hemiptera; and Terayama et[42]
Hayashi et al. [40] for Coleoptera; Ishikawa et al. [41] for Hemiptera; and Terayama et al. al.
for Hymenoptera (ants). Identification to the species level was difficult, so
[42] for Hymenoptera (ants). Identification to the species level was difficult, so several several species
were identified
species at genus,
were identified family,family,
at genus, and orderand level.
order Taxa
level.that
Taxaoccurred on ≥20
that occurred onlogs≥20 were
logs
recorded as dominant taxa.
were recorded as dominant taxa.
2.3. Data Analysis
2.3. Data Analysis
All statistical analyses were conducted using R ver. 4.0.5 [43]. A generalised linear
All statistical analyses were conducted using R ver. 4.0.5 [43]. A generalised linear
model (GLM) was applied to explain the species richness of invertebrates. The diameter of
model (GLM) was applied to explain the species richness of invertebrates. The diameter of
the logs, position (top/bottom) on the logs, and anamorph and teleomorph areas on the
the logs, position (top/bottom) on the logs, and anamorph and teleomorph areas on the log
log surface were set as explanatory variables. A binomial distribution error was assumed,
surface werelink
and a logit set function
as explanatory variables.
was used. A binomial distribution
The log-transformed error
surface area ofwas assumed,
the logs was setandas
aan
logit link function
offset term. was used. The log-transformed surface area of the logs was set as an
offset The
term.relationship between invertebrate community composition and environmental
The
factors wasrelationship
visualisedbetween invertebrate
using non-metric community composition
multi-dimensional scaling (NMDS)and environmental
with the vegan
factors
package was visualised
[44]. using
Similarities non-metric
of the multi-dimensional
invertebrate scaling
communities across the(NMDS)
logs werewith the ve-
calculated
gan package [44]. Similarities of the invertebrate communities across the
using the Raup–Crick similarity index (vegdist command), and this matrix was used logs were calcu-
to
lated using the Raup–Crick similarity index (vegdist command), and this matrix
develop the NMDS ordination plot (metaMDS command). The significance of the difference was used
to
indevelop
community the NMDS ordination
composition plotthe
between (metaMDS
top and command). The significance
bottom positions of the logs ofwasthedeter-
dif-
ference in community composition between the top and bottom positions of
mined using permutation multivariate analysis of variance [45] with 10,000 permutations the logs was
determined using permutation
(adonis command). In addition,multivariate
communityanalysis
varianceofbetween
variancesamples
[45] with 10,000 permu-
(calculated using
tations (adonis command)
the betadisper command). wasIn addition,
compared community
between thevariance between
top and bottomsamples
positions(calculated
using anwere set as environmental variables.
A set of GLMs were applied to explain the occurrence o
brate species. The environmental variables that were detect
Forests 2021, 12, 1124
vfit (diameter, anamorph percentage, and teleomorph percen
4 of 10
variables. Binomial distribution errors were assumed. Logit
analysis of variance (anova command). Finally, the significance of the effects of the en-
vironmental variables on the invertebrate communities was determined using the envfit
3. Results
command; sampling month, diameter, anamorph percentage, and teleomorph percentage
were set as environmental variables.
The mean percentage of debarked area per log was 10%
A set of GLMs were applied to explain the occurrence of the three dominant inverte-
brate species. The environmental variables that were detected to be significant using envfit
from June to July, and remained constant thereafter (Figure
(diameter, anamorph percentage, and teleomorph percentage) were set as explanatory
variables. Binomial distribution errors were assumed. Logit link functions were used.
debarked more than bottoms of the logs. The anamorph are
the3. debarked
Results
area in March but decreased to zero by the en
The mean percentage of debarked area per log was 10% in March, increased slightly
trast,
from the
June toteleomorph area was
July, and remained constant seldom
thereafter (Figure 2a).observed until
The tops of the logs were June
debarked more than bottoms of the logs. The anamorph area occupied almost all parts
and occupied
of the debarked area almost alldecreased
in March but parts of theby debarked
to zero area 2b).
the end of July (Figure until
In Nov
contrast, the teleomorph area was seldom observed until June but greatly increased in July
and occupied almost all parts of the debarked area until November (Figure 2c).
Figure 2. Percentage area of debarked (a), anamorph (b), and teleomorph (c) of Bisogniauxia spp. on
Figure 2. Percentage area of debarked (a), anamorph (b), and teleom
Quercus serrata logs. Grey and white bars show the top and bottom sides of the logs, respectively.
Quercus serrata
In total, 38 taxalogs. Grey
of insects were and white
recorded bars
(Table S1). show was
Coleoptera thethetop and
largest bottom
group
and consisted of 28 taxa, followed by 4 taxa of Hemiptera and 3 taxa of Lepidoptera.
Archaeognatha, Hymenoptera, and Psocodea included one taxon each. The GLM indicated
In total, 38 taxa of insects were recorded (Table S1). Cole
and consisted of 28 taxa, followed by 4 taxa of Hemiptera a
chaeognatha, Hymenoptera, and Psocodea included one taxForests 2021, 12, 1124 5 of 10
that insect diversity was negatively correlated with log diameter and positively correlated
with teleomorph area (Table 1). The bottom sides of the logs had lower insect richness than
Forests 2021, 12, x FOR PEER REVIEWthe top sides. 5 of 10
The observed insect communities were significantly correlated with sampling month,
log diameter, and anamorph area (Figure 3); however, the position on the logs (top/bottom)
was not correlated with the insect communities. Among the recorded insect taxa, Librodor
(Glischrochilus) ipsoides, Anamorph
Laemophloeus submonilis, and Neuroctenus castaneus– were observed
d.f. (null) 444
on ≥20 logs and were thus assigned as dominant taxa; these species were not recorded
Null deviance
in March but were frequently 814.12
recorded in June and July, particularly on the top sides of
the logs (Figure 4). d.f. (residual)
Although 441 by August and
L. submonilis and L. ipsoides disappeared
September, respectively, N. deviance
Residual castaneus remained until November. Additionally,
770.35 N. castaneus
reproduced on the surfaceAIC of Biscogniauxia teleomorphs (Figure 4c). The GLM based on the
1517.1
data from July suggested that the occurrence of L. submonilis was positively associated with
*the
pForests 2021, 12, x FOR PEER REVIEW 6 of 10
Forests 2021, 12, 1124 6 of 10
Figure 4. Frequencies of Laemophloeus submonilis (a), Librodor ipsoides (b), and Neuroctenus castaneus (c)
Figure 4. Frequencies of Laemophloeus submonilis (a), Librodor ipsoides (b), and Neuroctenus castaneus
on Quercus serrata logs. Grey and white bars show the top and bottom sides of the logs, respectively.
(c) on Quercus serrata logs. Grey and white bars show the top and bottom sides of the logs, respec-
tively.
Table 2. GLM results showing estimated coefficients between occurrences of the three dominant
insect 2.
Table species
GLM and log showing
results variables.estimated coefficients between occurrences of the three dominant
insect species and log variables.
Variable Laemophloeus Librodor Neuroctenus
Variable
Diameter Laemophloeus
0.18 Librodor
0.32 * Neuroctenus
0.20 **
Anamorph
Diameter area 0.97
0.18* 0.321.17* 0.20–**
Teleomorph
Anamorph area area 0.08
0.97** 1.17 *
0.24 0.51
– **
d.f. (null) 63 63 63
Teleomorph
Null deviance area 0.08
51.98* 0.24
71.98* 0.51 **
86.46
d.f. (null)
d.f. (residual) 63
60 6360 6361
Residual deviance
Null deviance 24.47
51.98 22.51
71.98 41.33
86.46
AIC
d.f. (residual) 32.47
60 30.51
60 47.33
61
* p < 0.05; ** p < 0.01.
Residual deviance 24.47 22.51 41.33
AIC 32.47 30.51 47.33
4. Discussion
* p < 0.05; ** p < 0.01.
In the present study, the debarking behaviour of squirrels improved the visibility of in-
4.sect communities on Biscogniauxia sporocarps, as their initial anamorphic stages are usually
Discussion
hidden under the bark. Fungivorous saproxylic insects with flat morphology often inhabit
In the present study, the debarking behaviour of squirrels improved the visibility of
invisible microhabitats, such as the space between bark and wood, which reduces their
insect communities on Biscogniauxia sporocarps, as their initial anamorphic stages are usu-
visibility considerably [1,46]. Debarking by squirrels targeting Biscogniauxia sporocarps
ally hidden under the bark. Fungivorous saproxylic insects with flat morphology often
enabled the author to observe the sporocarps starting from the very early stages of their
inhabit invisible microhabitats, such as the space between bark and wood, which reduces
development. Squirrels very accurately found Biscogniauxia sporocarps hidden under the
their visibility considerably [1,46]. Debarking by squirrels targeting Biscogniauxia sporo-
bark. It is well-known that squirrels can detect hypogeous sporocarps by smell [47,48]. Xy-
carps enabled the author to observe the sporocarps starting from the very early stages of
lariaceous ascomycetes effuse a variety of volatiles, depending on the developmental stage
their development.
of their sporocarps Squirrels very
[49], which may accurately found
be detected Biscogniauxia
by squirrels. sporocarps
McKeever hidden
[47] also un-
reported
der the bark. It is well-known that squirrels can detect hypogeous sporocarps
that fungi constitute the largest portion of squirrel diets, particularly in the summer and by smell
[47,48]. Xylariaceous
early autumn, ascomycetes
and represent effuse alargest
the second varietyportion
of volatiles, depending
of their on the
diets in the develop-
early spring;
mental
however, the majority of the fungi eaten by squirrels may be basidiomycetes [48]. I [47]
stage of their sporocarps [49], which may be detected by squirrels. McKeever also
also reported
observed that fungi
a squirrel constitute
eating the largest
old (fruiting portion
from the of squirrel
previous diets, particularly
year) Sarcomyxa in the
serotina (basid-Forests 2021, 12, 1124 7 of 10
iomycetes) sporocarps on 18 April 2021 in Japan (the personal observation of Yu Fukasawa).
Although it is unclear whether squirrel fungivore of xylariaceous ascomycetes is common,
I found the same type of debarking on freshly felled oak logs approximately 8 km away
from the study site in 2018 and again at the study site in February 2021 (Figure S1, the
personal observation of Yu Fukasawa). Currah et al. [6] investigated the stomach contents
of squirrels and flying squirrels in North America and found fragments of Xylariaceae
and Diatripaceae ascocarps. However, again, basidiomycetes sporcarps constituted the
majority of their gut contents. Xylariaceous ascomycetes may constitute a supplemental
diet during the early spring when other fresh food sources are unavailable. Fungal spores
contain a high percentage of nitrogen, but their digestibility is very low in squirrels [50].
Among the recorded insect taxa, almost all taxa except for Niponius osorioceps and
Pheidole fervida belonged to families that include species reported to be fungivores [1,51,52].
More specifically, species belonging to Aradidae, Cucujidae, Nitidulidae, Anthribidae,
Monotomidae, Silvanidae, Corylophidae, and Biphyllidae are known as ascomycete
eaters [51,53]. I even observed Lepidopteran larvae of unknown identity grazing the
surface of Biscogniauxia stromata. Tineoidea moths are known to have fungivorous habits,
although they feed on basidiomycotan wood decay fungi [22,54]. Powell [55] reported that
Lepidopteran species belonging to Pyralidae feed on the stromata of Hypoxylon occidentale
(Xylariaceae, Ascomycota). As shown through NMDS (Figure 3), the presence of Biscogni-
auxia spp. may strongly attract fungivorous insects and affect their community structure.
Lee et al. [56] reported that stem canker caused by Annulohypoxylon truncatum (Xylariaceae)
on oak stem significantly increases invertebrate diversity in Korea, irrespective of the
presence or absence of sap flow.
In the present study, I identified three dominant insect species: Librodor ipsoides (Ni-
tidulidae, Coleoptera); L. submonilis (Cucujidae, Coleoptera); and N. castaneus (Aradidae,
Hemiptera)—they each had significant relationships with the occurrence of Bisocogni-
auxia spp. (Table 2). Although the gut contents of these species were not surveyed in
the present study, it is highly likely that these three species were all fungivores and had
intimate associations with Biscogniauxia spp. during their life cycle. Nitidulidae is a well-
known fungivorous Coleoptera occurring in not only fungal fruit bodies but also in the sap
flows of damaged trees, fermenting fruits, and pathological plant tissues—-such species
probably have yeasts or pathogenic fungi as a normal and essential part of their diet [51].
Specifically, species in the genus Librodor are known as ‘sap beetles’ that forage and breed
in fermented sap flow [57](referred as a genus Glischrochilus). L. ipsoides has also been
found in the sap on Quercus acutissima trees in Japan, although its abundance is quite low
compared with that of other sap beetles [58]. On the other hand, L. ipsoides has been found
in the fruit bodies of a basidiomycete Cryptoporus volvatus on Pinus densiflora in Korea [59],
but it was not found in C. volvatus in Japan [22]. In the present study, I newly found
L. ipsoides on Biscogniauxia spp. fruit bodies at a relatively high frequency (>30%; Figure 4),
indicating that the stromata and/or conidia of Biscogniauxia spp. are an important habitat
of this species. Lawrence [60] reported observations of Prometopia sexmaculata (Nitidulidae)
breeding in the stromata of Hypoxylon (Xylariaceae) on oak. Cucujidae are also known as
Coleopteran ascomycete eaters. Species in the genus Laemophloeus have been frequently
reported in association with ascomata, such as Daldinia, Tubercularia, Hypoxylon, and Biscog-
niauxia (=Nummularia) [51]. Therefore, it is not surprising that I found L. submonilis on
Biscogniauxia spp. (B. maritima and B. plana) in the present study, even though it is a newly
found association in Japanese species.
In Aradidae (Hemiptera), most species are fungivores [53,61] that feed on fungal
hyphae, using their piercing-sucking mouthparts to suck the cell contents, and have
adapted gut systems [62], but little is known about the fungal host association of most
species, particularly for ascomycetes [53]. Most of the Aradidae species occur preferentially
on dead wood during the early stages of decay, probably due to the presence of their dietary
fungal species [53]. As such, outbreaks of some Aradidae species are closely associated with
large dieback events, such as forest fires and pests. Aradus lugubris can appear immediatelyForests 2021, 12, 1124 8 of 10
after forest fires, when they feed on Daldinia loculata (Lév.) Sacc. (Xylariaceae), a known
fire-related ascomycete in boreal forests [63]. Similarly, large-scale wind throws and bark
beetle outbreaks in a Norway spruce forest in the Bavarian Forest in Germany, where
dead spruce snags were intensively colonised by a basidiomycete Fomitopsis pinicola [64],
provided a suitable habitat for Aradus obtectus [53]. In Japan, Q. serrata trees have recently
suffered from oak wilt disease [65]. However, relationships between oak wilt disease and
the occurrence of Biscogniauxia spp. are unclear. Fukasawa et al. [66] compared latent
fungal communities within Q. serrata trunks in stands with or without the prevalence of
oak wilt disease and did not detect Biscogniauxia spp.
Parental care has been observed in Aradidae, during which the male safeguards the
egg mass for several weeks, and parental care may be extended to the nymphal stage [53].
In the present study, I also observed that adults appeared to guard the mass of nymphs
(Figure 4). In addition, I observed many nymphs spraying (probably some chemicals)
simultaneously from their tail when they were shaded by my hands.
To summarise, I observed the insect communities on saproxylic Biscogniauxia spp. on
oak dead wood during the transition from the anamorphic to teleomorphic stages; this was
aided by debarking by squirrels. The presence of Biscogniauxia spp. significantly affected
the insect communities. More specifically, the population dynamics of two Coleoptera and
one Hemiptera dominant species revealed that Biscogniauxia spp. represents an important
habitat in their life cycles.
Supplementary Materials: The following are available online at https://www.mdpi.com/article/
10.3390/f12081124/s1. Figure S1: Another evidence of debarking of Quercus serrata log by squirrel
targeting on ascomycete anamorph, Table S1: Insect taxa recorded in the present study.
Funding: This research received no external funding.
Acknowledgments: The author is grateful to Shuhei Takemoto for identification of Biscogniauxia spp.
I also thank Chisato Kobayashi for letting me use our garden for my experiment.
Conflicts of Interest: The author declares that they have no conflict of interest.
References
1. Stokland, J.N.; Siitonen, J.; Jonsson, B.G. Biodiversity in Dead Wood; Cambridge University Press: Cambridge, UK, 2012.
2. Seibold, S.; Bässler, C.; Brandl, R.; Gossner, M.M.; Thorn, S.; Ulyshen, M.D.; Müller, J. Experimental studies of dead-wood
biodiversity—A review identifying global gaps in knowledge. Biol. Conserv. 2015, 191, 139–149. [CrossRef]
3. Rayner, A.D.M.; Boddy, L. Fungal Decomposition of Wood: Its Biology and Ecology; John Wiley & Sons: Chichester, UK, 1988.
4. Fukasawa, Y.; Hyodo, F.; Kawakami, S. Foraging association between myxomycetes and fungal communities on coarse woody
debris. Soil Biol. Biochem. 2018, 121, 95–102. [CrossRef]
5. Hanski, I. Fungivory: Fungi, insects and Ecology. In Insect–Fungus Interactions; Wilding, N., Collins, N.M., Hammond, P.M.,
Webber, J.F., Eds.; Academic Press: London, UK, 1989.
6. Currah, R.S.; Smreciu, E.A.; Lehesvirta, T.; Niemi, M.; Larsen, K.W. Fungi in the winter diets of northern flying squirrels and red
squirrels in the boreal mixedwood forest of northern Alberta. Can. J. Bot. 2000, 78, 1514–1520.
7. Elliott, T.F.; Elliott, K. Vertebrate consumption and dispersal of the Nothofagaceae associated ascomycete Cyttaria. Austral. Ecol.
2019, 44, 747–751. [CrossRef]
8. Schigel, D.S. Fungivory and host associations of Coleoptera: A bibliography and review of research approaches. Mycology 2012, 3,
258–272. [CrossRef]
9. Buxton, C.M.G. British Diptera associated with fungi. III. Flies of all families reared from about 150 species of fungi. Entomol.
Mon. Mag. 1960, 96, 61–94.
10. Elton, C.S. The Pattern of Animal Communities; Methuen and Co.: Slingsby, UK, 1966.
11. Jonsell, M.; Nordlander, G. Host selection patterns in insects breeding in bracket fungi. Ecol. Entomol. 2004, 29, 697–705. [CrossRef]
12. Yamashita, S.; Hijii, N. The role of fungal taxa and developmental stage of mushrooms in determining the composition of the
mycophagous insect community in a Japanese forest. Eur. J. Entomol. 2007, 104, 225–233. [CrossRef]
13. Nakamori, T.; Suzuki, A. Surface properties of the mushroom Strobilurus ohshimae result in food differentiation by collembolan
species. Eur. J. Soil Biol 2008, 44, 478–482. [CrossRef]
14. Yamashita, S.; Ando, K.; Hoshina, H.; Ito, N.; Katayama, Y.; Kawanabe, M.; Maruyama, M.; Itioka, T. Food web structure of the
fungivorous insect community on bracket fungi in a Bornean tropical rain forest. Ecol. Entomol. 2015, 40, 390–400. [CrossRef]
15. Kobayashi, T.; Sota, T. Evolution of host use in fungivorous ciid beetles (Coleoptera: Ciidae): Molecular phylogeny focusing on
Japanese taxa. Mol. Phylogen Evol. 2021, 162, 107197. [CrossRef] [PubMed]Forests 2021, 12, 1124 9 of 10
16. Tuno, N. Insect feeding on spores of a bracked fungus, Elfvingia applanata (Pers.) Karst. (Ganodermataceae, Aphyllophorales).
Ecol. Res. 1999, 14, 97–103. [CrossRef]
17. Kadowaki, K.; Keschen, R.A.B.; Beggs, J.R. No evidence for a Ganoderma spore dispersal mutualism in an obligate spore-feeding
beetle Zearagytodes maculifer. Fung Biol. 2011, 115, 768–774. [CrossRef] [PubMed]
18. Persson, Y.; Ihrmark, K.; Stenlid, J. Do bark beetles facilitate the establishment of rot fungi in Norway spruce? Fung. Ecol. 2011, 4,
262–269. [CrossRef]
19. Kitabayashi, K.; Tuno, N. Soil burrowing Muscina angustifrons (Diptera: Muscidae) larvae excrete spores capable of forming
mycorrhizae underground. Mycoscience 2018, 59, 252–258. [CrossRef]
20. Seibold, S.; Müller, J.; Baldrian, P.; Cadotte, M.W.; Stursová, M.; Biedermann, P.H.W.; Krah, F.S.; Bässler, C. Fungi associated with
beetles dispersing from dead wood—Let’s take the beetle bus! Fung. Ecol. 2019, 39, 100–108. [CrossRef]
21. Takahashi, K.H.; Tuno, N.; Kagaya, T. The relative importance of spatial aggregation and resource partitioning on the coexistence
of mycophagous insects. Oikos 2005, 109, 125–134. [CrossRef]
22. Kadowaki, K. Species coexistence patterns in a mycophagous insect community inhabiting the wood-decaying bracket fungus
Cryptoporus volvatus. Eur. J. Entomol. 2010, 107, 89–99. [CrossRef]
23. Komonen, A.; Penttila, R.; Lindgren, M.; Hanski, I. Forest fragmentation truncates a food chain based on an old-growth forest
bracket fungus. Oikos 2000, 90, 119–126. [CrossRef]
24. Komonen, A.; Kouki, J. Occurrence and abundance of fungus-dwelling beetles (Ciidae) in boreal forests and clearcuts: Habitat
associations at two spatial scales. Anim. Biodiv. Conserv. 2005, 28, 137–147.
25. Joseph, R.; Keyhani, N.O. Fungal mutualisms and pathosystems: Life and death in the ambrosia beetle mycangia. Appl. Microbiol.
Biotechnol. 2021, 105, 3393–3410. [CrossRef]
26. Pažoutová, S.; Srutka, P.; Holusa, J.; Chudickova, M.; Kolarik, M. Diversity of xylariaceous symbionts in Xiphydria woodwasps:
Role of vector and a host tree. Fung. Ecol. 2010, 3, 392–401. [CrossRef]
27. Visser, A.A.; Ros, V.I.D.; de Beer, Z.W.; Debets, A.J.M.; Hartog, E.; Kuyper, T.W.; Læssøe, T.; Slippers, B.; Aanen, D.K. Level of
specificity of Xylaria species associated with fungus-growing termites: A phylogenetic approach. Mol. Ecol. 2009, 18, 553–567.
[CrossRef]
28. Xiao, G.; Ying, S.H.; Zheng, P.; Wang, Z.L.; Zhang, S.; Xie, X.Q.; Shang, Y.; St Leger, R.J.; Zhao, G.P.; Wang, C.; et al. Genomic
perspectives on the evolution of fungal entomopathogenicity in Beauveria bassiana. Sci. Rep. 2012, 2, 483. [CrossRef] [PubMed]
29. Stone, L.B.L.; Bidochka, M.J. The multifunctional lifestyles of Metarhizium: Evolution and applications. Appl. Microbiol. Biotechnol.
2020, 104, 9935–9945. [CrossRef] [PubMed]
30. Nikoh, N.; Fukatsu, T. Interkingdom host jumping underground: Phylogenetic analysis of entomoparasitic fungi of the genus
Cordyceps. Mol. Biol. Evol. 2000, 17, 629–638. [CrossRef] [PubMed]
31. Watanabe, N.; Okada, T.; Tomaru, N.; Nishimura, N.; Nakagawa, M. Forest dynamics of stands damaged by Japanese oak wilt in
the Kaisho forest, Aichi Prefecture. J. Jap. For. Soc. 2016, 98, 273–278. [CrossRef]
32. Nishikawa, S.; Kubo, M.; Ozaki, Y. Mass mortality due to the spread of oak wilt disease in a 19-ha secondary Quercus serrata
forest within Sambe Forest at Shimane University. Bull Fac. Life Env. Sci. Shimane Univ. 2018, 23, 21–26.
33. Endoh, R.; Suzuki, M.; Okada, G.; Takeuchi, Y.; Futai, K. Fungus symbionts colonizing the galleries of the ambrosia beetle Platypus
quercivorus. Microb. Ecol. 2011, 62, 106–120. [CrossRef]
34. Masuya, H.; Manabe, R.; Ohkuma, M.; Endoh, R. Draft genome sequence of Raffaelea quercivora JCM 11526, a Japanese oak wilt
pathogen associated with the Platypodid beetle, Platypus quecivorus. Genome Announc. 2016, 4, e00755-16. [CrossRef]
35. Kobayashi, T.; Oguro, M.; Akiba, M.; Taki, H.; Kitajima, H.; Ishihara, H. Mushroom yield of cultivated shiitake (Lentinula edodes)
and fungal communities in logs. J. Res. 2020, 25, 269–275. [CrossRef]
36. Tokimoto, K.; Komatsu, M.; Fukumasa-Nakai, Y. Establishing the selection method for the Lentinula edodes strains resistant to
Trichoderma spp. Proc. Jpn. Acad 1994, 70, 112–116. [CrossRef]
37. Kim, J.Y.; Kwon, H.W.; Yun, Y.H.; Kim, S.H. Identification and characterization of Trichoderma species damaging Shiitake
mushroom bed-logs infested by Camptomyia pest. J. Microbiol Biotechnol 2016, 26, 909–917. [CrossRef] [PubMed]
38. Kurosawa, Y.; Hisamatsu, S.; Sasaji, H. The coleoptera of Japan in color Vol.III; Hoikusha: Tokyo, Japan, 1998. (In Japanese)
39. Ueno, S.; Kurosawa, Y.; Sato, M. The coleoptera of Japan in color Vol.II; Hoikusha: Tokyo, Japan, 1999. (In Japanese)
40. Hayashi, M.; Morimoto, K.; Kimoto, S. The coleoptera of Japan in color Vol.IV; Hoikusha: Tokyo, Japan, 2002. (In Japanese)
41. Ishikawa, T.; Takai, M.; Yasunaga, T. A Field Guide to Japanese Bugs: Terrestrial Heteropterans; Zenkoku Noson Kyoiku Kyokai:
Tokyo, Japan, 2012; Volume 3. (In Japanese)
42. Terayama, M.; Kubota, S.; Eguchi, K. Encyclopedia of Japanese Ants; Asakura Shoten: Tokyo, Japan, 2014. (In Japanese)
43. R Core Team. R: A Language and Environment for Statistical Computing; The R Foundation for Statistical Computing: Vienna,
Austria, 2021.
44. Oksanen, J.; Blanchet, F.G.; Kindt, R.; Legendre, P.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; Stevens, M.H.H.;
Szoecs, E.; et al. Vegan: Community Ecology Package; R Package Version 2.5-7; 2020. Available online: https://cran.r-project.org/
web/packages/vegan/index.html.
45. Anderson, M.J. A new method for non parametric multivariate analysis of variance. Austral Ecol. 2001, 26, 32–46.
46. Ulyshen, M.D. Saproxylic Insects: Diversity, Ecology and Conservation; Springer: Cham, Switzerland, 2019.
47. McKeever, S. The biology of the Golden-mantled ground squirrel, Citellus lateralis. Ecol. Monogr. 1964, 34, 383–401. [CrossRef]Forests 2021, 12, 1124 10 of 10
48. Maser, C.; Maser, Z. Interactions among squirrels, mycorrhizal fungi, and coniferous forests in Oregon. Great Basin Nat. 1988, 48,
358–369.
49. Stadler, M. Importance of secondary metabolites in the Xylariaceae as parameters for assessment of their taxonomy, phylogeny,
and functional biodiversity. Curr. Res. Environ. Appl. Mycol. 2011, 1, 75–133. [CrossRef]
50. Cork, S.J.; Kenagy, G.J. Nutritional value of hypogeous fungus for a forest-dwelling ground squirrel. Ecology 1989, 70, 577–586.
[CrossRef]
51. Crowson, R.A. The associations of Coleoptera with Ascomycetes. In Fungus-Insect Relationships; Wheeler, Q., Blackwell, M., Eds.;
Columbia University Press: New York, NY, USA, 1984; pp. 257–285.
52. Katanoda, I.; Sato, Y.; Morita, S.; Okubo, H. Adult emergence and oviposition preference of Mesosa longipennis for Chinese Quercus
acutissima. Kyushu J. Res. 2005, 58, 105–108. (In Japanese)
53. Gossner, M.M.; Damken, C. Diversity and ecology of saproxylic Hemiptera. In Saproxylic Insects: Diversity, Ecology and Conservation
(Zoological Monographs 1); Ulyshen, M.D., Ed.; Springer: Cham, Switzerland, 2018.
54. Powell, J.A.; Mitter, C.; Farrell, B. Evolution of larval food preferences in Lepidoptera. In Lepidoptera, Moths and Butterflies Volume
1: Evolution, Systematics, and Biogeography; Kristensen, N.P., Ed.; de Gruyter: Berlin, Germany, 1998; pp. 403–422.
55. Powell, J.A. Apomyelois bistriatella: A moth which feeds in an ascomycete fungus (Lepidoptera: Pyralidae). J. N. Y. Entomol. Soc.
1967, 75, 190–194.
56. Lee, Y.-B.; An, S.J.; Park, C.G.; Kim, J.; Han, S.; Kwak, Y.-S. Oak tree canker disease supports arthropod diversity in a natural
ecosystem. Plant Path. J. 2014, 30, 43–50. [CrossRef]
57. Jacobsen, R.M.; Birkemore, T.; Sverdrup-Thygeson, A. Priority effects of early successional insects influence late successional
fungi in dead wood. Ecol. Evol. 2015, 5, 4896–4905. [CrossRef] [PubMed]
58. Yoshimoto, J.; Kakutani, T.; Nishida, T. Influence of resource abundance on the structure of the insect community attracted to
fermented tree sap. Ecol. Res. 2005, 20, 405–414. [CrossRef]
59. Park, M.S.; Fong, J.J.; Lee, H.; Shin, S.; Lee, S.; Lee, N.; Lim, Y.W. Determination of coleopteran insects associated with spore
dispersal of Cryptoporus volvatus. J. Asia-Pac. Entomol 2014, 17, 647–651. [CrossRef]
60. Lawrence, J.F. Coleoptera associated with an Hypoxylon species (Ascomycetes: Xylariaceae) on oak. Coleopt. Bull. 1977, 31,
309–312.
61. Seibold, S.; Bässler, C.; Baldrian, P.; Thorn, S.; Müller, J.; Gossner, M.M. Wood resource and not fungi attract early-successional
saproxylic species of Heteroptera—An experimental approach. Insent. Conserv. Divers. 2014, 7, 533–542. [CrossRef]
62. Nardi, J.B.; Bee, C.M.; Miller, L.A.; Taylor, S.J. Distinctive features of the alimentary canal of a fungus-feeding hemipteran, Mezira
granulata (Heteroptera: Aradidae). Arthropod Struct. Dev. 2009, 38, 206–215. [CrossRef] [PubMed]
63. Wikars, L.-O. The wood-decaying fungus Daldinia loculata (Xylariaceae) as an indicator of fire-dependent insects. Ecol. Bull. 2001,
49, 263–268.
64. Vogel, S.; Alvarez, B.; Bässler, C.; Müller, J.; Thorn, S. The red-belted bracket (Fomitopsis pinicola) colonizes spruce trees early after
bark beetle attack and persists. Fung. Ecol. 2017, 27, 182–188. [CrossRef]
65. FFPRI. Naragare Boujo No Shintenkai; Forestry and Forest Products Research Institute: Tsukuba, Japan, 2015; ISBN 978-4-905304-48-7.
(In Japanese)
66. Fukasawa, Y.; Matsukura, K.; Stephan, J.; Makoto, K.; Suzuki, S.N.; Kominami, Y.; Takagi, M.; Tanaka, N.; Takemoto, S.; Kinuura,
H.; et al. Patterns of community composition and diversity in latent fungi of living Quercus serrata trunks across a range of oak
wilt prevalence and climate variables in Japan. Fung. Ecol. 2021, in press. [CrossRef]You can also read

















































