Asexual Female Gametogenesis Involves Contact with a Sexually Fated Megaspore in Apomictic Hieracium1 OPEN
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Asexual Female Gametogenesis Involves Contact with a
Sexually Fated Megaspore in Apomictic Hieracium1[OPEN]
Martina Juranić,a Matthew R. Tucker,b Carolyn J. Schultz,b Neil J. Shirley,b,c Jennifer M. Taylor,d
Andrew Spriggs,d Susan D. Johnson,a Vincent Bulone,b,c and Anna M. Koltunowa,2
a
Commonwealth Scientific and Industrial Research Organisation Agriculture and Food, Urrbrae, South Australia
5064, Australia
b
School of Agriculture, Food, and Wine, University of Adelaide, Urrbrae, South Australia 5064, Australia
c
Australian Research Council Centre of Excellence in Plant Cell Walls, Wine Innovation Central, University of
Adelaide, Urrbrae, South Australia 5064, Australia
d
Commonwealth Scientific and Industrial Research Organisation Agriculture and Food, Black Mountain
Laboratories, Black Mountain, Australian Capital Territory 2601, Australia
ORCID IDs: 0000‑0001‑6113‑7623 (M.J.); 0000‑0003‑4661‑6700 (M.R.T.); 0000‑0003‑2026‑9122 (C.J.S.); 0000‑0001‑8114‑9891 (N.J.S.);
0000‑0001‑6111‑8588 (A.S.); 0000‑0003‑2809‑4160 (V.B.); 0000‑0001‑5016‑7683 (A.M.K.)
Apomixis results in asexual seed formation where progeny are identical to the maternal plant. In ovules of apomictic species of
the Hieracium subgenus Pilosella, meiosis of the megaspore mother cell generates four megaspores. Aposporous initial (AI) cells
form during meiosis in most ovules. The sexual pathway terminates during functional megaspore (FM) differentiation, when an
enlarged AI undergoes mitosis to form an aposporous female gametophyte. Then, the mitotically programmed FM dies along
with the three other megaspores by unknown mechanisms. Transcriptomes of laser-dissected AIs, ovule cells, and ovaries from
apomicts and AI-deficient mutants were analyzed to understand the pathways involved. The steps leading to AI mitosis and
sexual pathway termination were determined using antibodies against arabinogalactan protein epitopes found to mark both
sexual and aposporous female gametophyte lineages at inception. At most, four AIs differentiated near developing megaspores.
The first expanding AI cell to contact the FM formed a functional AI that underwent mitosis soon after megaspore degeneration.
Transcriptome analyses indicated that the enlarged, laser-captured AIs were arrested in the S/G2 phase of the cell cycle and
were metabolically active. Further comparisons with AI-deficient mutants showed that AIs were enriched in transcripts encod-
ing homologs of genes involved in, and potentially antagonistic to, known FM specification pathways. We propose that AI and
FM cell contact provides cues required for AI mitosis and megaspore degeneration. Specific candidates to further interrogate
AI-FM interactions were identified here and include Hieracium arabinogalactan protein family genes.
The polyploid Hieracium subgenus Pilosella II (As- meiosis, followed by double fertilization within the
teraceae) contains species such as H. pilosella that form female gametophyte to initiate seed formation. In con-
seeds and genetically variable progeny sexually. This trast, some subgenus Pilosella species, such as H. praeal-
requires male and female gametophyte formation via tum, H. piloselloides, and H. caespitosum, can reproduce
by an asexual pathway called apomixis where progeny
are genetically identical to the mother plant (Fig. 1;
1
This work was supported by a CSIRO Scientific and Industry Bicknell and Koltunow, 2004; Hand and Koltunow,
Endowment Fund grant (RP01-006 to A.M.K.), a CSIRO Office of 2014).
the Chief Executive Postdoctoral Fellowship Grant (to M.J.), and an
In all of the aforementioned sexual and apomictic spe-
Australian Research Council Future Fellowship Grant (to M.R.T.).
2
Address correspondence to anna.koltunow@csiro.au. cies, sexual reproduction initiates with the formation of
The author responsible for distribution of materials integral to a MMC in the ovule (Fig. 1). The MMC undergoes mei-
the findings presented in this article in accordance with the policy osis, giving rise to a tetrad of haploid megaspores. Three
described in the Instructions for Authors (www.plantphysiol.org) is: of these die during megaspore selection, while the me-
Anna M. Koltunow (anna.koltunow@csiro.au). gaspore closest to the chalazal end enlarges and matures
A.M.K. conceived the idea for the project; A.M.K., M.J., M.R.T., into the FM (Fig. 1B). In sexual Hieracium species, the FM
C.J.S., and J.M.T. designed the experiments; M.J. performed the is the progenitor of the sexual female gametophyte, and
majority of the experiments and identified and classified the arab- it undergoes three rounds of mitosis. Cellularization re-
inogalactan protein gene family with C.J.S. and A.S.; M.R.T. and sults in a mature sexual female gametophyte (Koltunow
N.J.S. carried out the transcriptome analysis of carbohydrate-related
et al., 2011b; Hand and Koltunow, 2014). Conversely, in
genes; J.M.T. and A.S. conducted the computational analyses; M.J.,
M.R.T., C.J.S., N.J.S., S.D.J., J.M.T., V.B., and A.M.K. analyzed the apomictic Hieracium, a somatic AI cell is the progenitor
data; M.J., M.R.T., C.J.S., J.M.T., V.B., and A.M.K. wrote the article of the aposporous female gametophyte. AI cells enlarge
with contributions from all authors. in the ovule near cells undergoing meiosis. Typically,
[OPEN]
Articles can be viewed without a subscription. a single AI cell undergoes mitotic divisions, forming a
www.plantphysiol.org/cgi/doi/10.1104/pp.18.00342 mature aposporous female gametophyte, while all four
®
Plant Physiology , July 2018, Vol. 177, pp. 1027–1049, www.plantphysiol.org © 2018 American Society of Plant Biologists. All Rights Reserved.1027
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.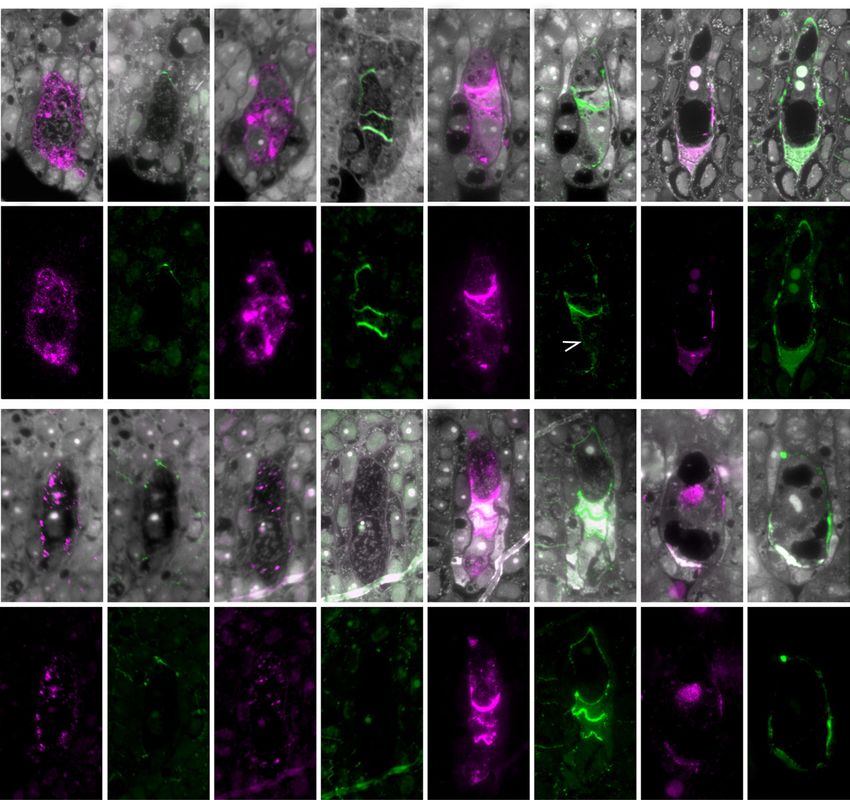
Juranić et al.
Figure 1. Early events of sexual and aposporous female gametophyte development in Hieracium spp. A, H. piloselloides (D36)
plant showing floral capitula, florets, and ovules. B, Comparison of megasporogenesis in sexual H. pilosella (P36) and in apo-
sporous H. piloselloides (D36). Mutants m115 and m134 are defective in AI formation. They undergo meiosis and functional
megaspore formation as found in sexual P36, except that they form a linear tetrad of megaspores. Both mutants form meiotically
reduced embryo sacs. The numbers of aposporous initial (AI) cells formed in apomicts, their locations, and steps leading to
a functional AI cell were examined in this study. DM, Degenerating megaspores; eFM, enlarging functional megaspore; FAI,
functional aposporous initial; FM, functional megaspore; LOA, LOSS OF APOSPORY locus required for AI cell specification and
growth; MMC, megaspore mother cell; MS, megaspores; NE, nucellar epidermis.
megaspores degenerate by unknown mechanisms and formation in R35 and D36, while AutE also controls
sexual development ceases (Fig. 1B). Formation of the autonomous endosperm formation in D36 (Catanach
embryo and endosperm within the aposporous female et al., 2006; Koltunow et al., 2011b; Ogawa et al.,
gametophyte is fertilization independent (i.e. autono- 2013). Deletion of either LOA or LOP by γ-irradiation
mous), which is rare in apomicts (Koltunow et al., 2011b; in apomict R35 leads to apomixis mutants showing
Hand and Koltunow, 2014). partial reversion to sexual reproduction. Sexual fe-
Apomixis in subgenus Pilosella species is controlled male gametophyte formation occurs if LOA is deleted,
by dominant loci, but the causal genes are unknown. and fertilization is required for seed formation if LOP
Aposporous female gametophyte formation and sex- is deleted. Deletion of both loci leads to full rever-
ual female gametophyte termination are controlled
sion to sexual reproduction (Koltunow et al., 2011b).
by the LOA locus in H. praealtum (isolate R35) and H.
piloselloides (D36). Signals arising during the initiation These observations indicate that LOA and LOP loci
of meiosis in ovules of the D36 apomict are required suppress sexual reproduction and that the sexual
for AI cell differentiation, indicating that early cross talk pathway is the default reproductive state (Catanach
occurs between sexual and apomictic pathways at apo- et al., 2006; Koltunow et al., 2011b). This also is con-
mixis initiation (Koltunow et al., 2011b). Fertilization- sistent with the facultative nature of apomixis in sub-
independent seed formation is controlled by two genus Pilosella apomicts, because a small percentage
known loci in different species. The LOSS OF PARTHE- of seeds are consistently derived via the sexual path-
NOGENESIS (LOP) locus controls both fertilization- way (Bicknell and Koltunow, 2004; Koltunow et al.,
independent embryo and autonomous endosperm 2011a).
1028 Plant Physiol. Vol. 177, 2018
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.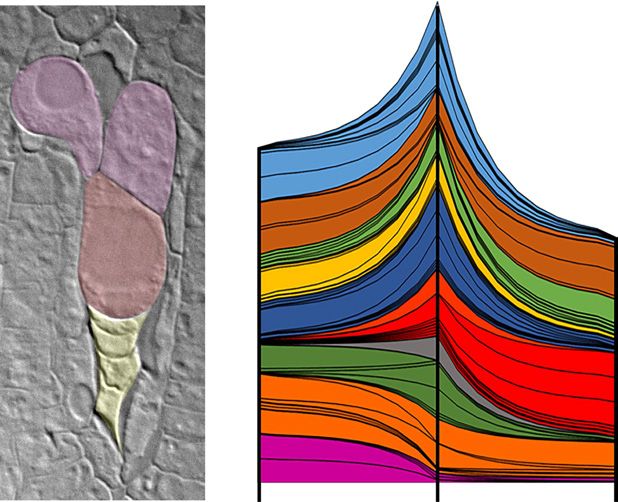
Cell Contact Promotes Apomixis in Hieracium
Processes favoring AI cell growth and leading to de- AI cell enlargement remain elusive. Callose distribu-
generation of the four megaspores may hypothetically tion was examined previously in whole-mount ovary
share similar mechanisms to those observed during squashes in apomictic and sexual species using Aniline
sexual FM selection and nonselected megaspore death. Blue staining to determine if gross alterations in callose
Although mechanistic information concerning FM patterning or deficiencies during meiosis correlated
specification, FM selection, and megaspore death in with sexual demise in the apomict (Tucker et al., 2001).
the sexual pathway remains sparse, nonselected mega- Callose was detected in the MMC, megaspores, and de-
spore death is thought to involve aspartic protease ac- generating megaspores in both species but not in AI cell
tivity in rice (Oryza sativa; Chen et al., 2008) and calcium walls (Tucker et al., 2001; Bicknell and Koltunow, 2004).
ion concentration in lettuce (Lactuca sativa; Qiu et al., Molecular signatures of AI cells also have been chal-
2008). Callose deposition in cell walls is thought to pro- lenging to define. Laser-capture microdissection, in
mote megaspore demise by cell isolation. Callose marks conjunction with 454 pyrosequencing, was used previ-
sexual gametogenic cells in ovules during MMC for- ously to examine transcripts in enlarging AI cells, early
mation, MMC meiosis, and FM selection in Hieracium, aposporous embryo (EAE) sacs, and somatic ovule (SO)
but it is selectively removed from the cell wall of the ex- cells in apomictic H. praealtum (R35; Fig. 2A; Okada et al.,
panding FM as it transitions toward the mitotic events 2013). These analyses showed that the AI cell transcrip-
of sexual female gametophyte formation (Tucker tome was most similar to the EAE sac transcriptome. It
et al., 2001; Bicknell and Koltunow, 2004; Tucker and was hypothesized that the captured, enlarging AI cells
Koltunow, 2014). had bypassed meiosis and transitioned to an asexual fe-
In relation to FM differentiation, studies with Arabi- male gametophyte program (Okada et al., 2013). How-
dopsis (Arabidopsis thaliana) indicate that a sporophytic ever, in silico assembly of cell type-specific transcripts
cytokinin signal is important for FM specification, generated by 454 pyrosequencing and their analyses
being prominent in the chalazal region of the ovule were limited due to the lack of sequencing depth, pref-
where the FM enlarges (Cheng et al., 2013). Arabinoga- erential enrichment of 3′ end sequences in amplified
lactan Protein18 (AGP18) is required for the selection RNA, the absence of a Hieracium survey genome, and
and viability of the FM in Arabidopsis. Overexpres- suitable assembled tissue transcriptomes to effectively
sion of AGP18 can promote the survival of megaspores generate, examine, and annotate gene models.
that would otherwise degenerate (Demesa-Arévalo Here, an analysis of differential gene expression in
and Vielle-Calzada, 2013). Arabinogalactan proteins AI and SO cells and EAE sacs using the Illumina se-
are highly glycosylated (up to 99% glycan) and form quencing platform was conducted, leveraging the
a large, diverse family within the Hyp-rich glycopro- recently established Hieracium survey genome and
tein superfamily. Arabinogalactan proteins have been transcriptome resource (Rabiger et al., 2016). In ad-
implicated in plant growth, development, cell death, dition, transcriptional signatures of enlarged AI cells
biotic and abiotic responses, and cell-to-cell commu- were identified by comparing laser-captured AI cell
nication (Ellis et al., 2010; Su and Higashiyama, 2018). transcriptomes with apomictic ovaries at specific de-
Arabinogalactan proteins also are cellular markers for velopmental stages and with deletion mutants that
female gametogenesis. In Arabidopsis ovules, JIM8 have lost the capacity to form AI cells. The in silico
and JIM13 antibodies detect arabinogalactan protein approaches allowed the identification of AI-enriched
epitopes in the cell wall of the FM, in cells of the sexual genes, including genes potentially involved in AI cell
female gametophyte, in the filiform apparatus, and in expansion and asexual female gametophyte growth.
integument cells lining the micropyle (Coimbra et al., Counterparts of some of the AI-enriched genes ap-
2007). JIM13 and JIM15 antibodies detect epitopes pres- peared to be involved in FM selection in the sexual
ent in cell walls of multicellular archesporial cells and pathway. The use of specific antibodies confirmed that
cells designated as MMCs and female gametophytes in arabinogalactan protein epitopes are markers of AI cell
the facultatively apomictic strawberry (Fragaria spp.; specification and demonstrated that physical contact
Dziadczyk et al., 2011; Leszczuk and Szczuka, 2018). between an AI cell and the enlarging, sexually derived
Arabinogalactan proteins also are detected in maturing FM preempted the formation of a functional AI cell ca-
asexual female gametophytes of apomictic Taraxacum pable of mitosis. Furthermore, dynamic changes in cal-
spp., which develop by mitotic diplospory (Gawecki lose cell wall patterning between the functional AI and
et al., 2017). Despite several studies involving arabino- secondary AI cells suggest that communication also
galactan proteins, the underlying mechanisms of their occurs between the functional AI and AI cells.
function remain unclear, and a range of models have
been proposed (Ellis et al., 2010; Lamport and Várnai,
2013; Lamport et al., 2018). RESULTS
Morphological markers defining AI cell identity prior Analysis of Laser-Captured Cell Transcriptomes Identifies
to their enlargement have not been identified in apo- Transcriptional Signatures in AI Cells
sporous apomicts. Thus, in Hieracium spp., the tempo-
ral and spatial specification of AI cells and their likely Illumina-derived transcriptome assemblies were
numbers within ovules relative to the sexual process established from amplified RNA isolated from laser-
remain unclear. Similarly, the mechanisms governing dissected uninucleate AI cells (Fig. 2A), EAE sacs with
Plant Physiol. Vol. 177, 2018 1029
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.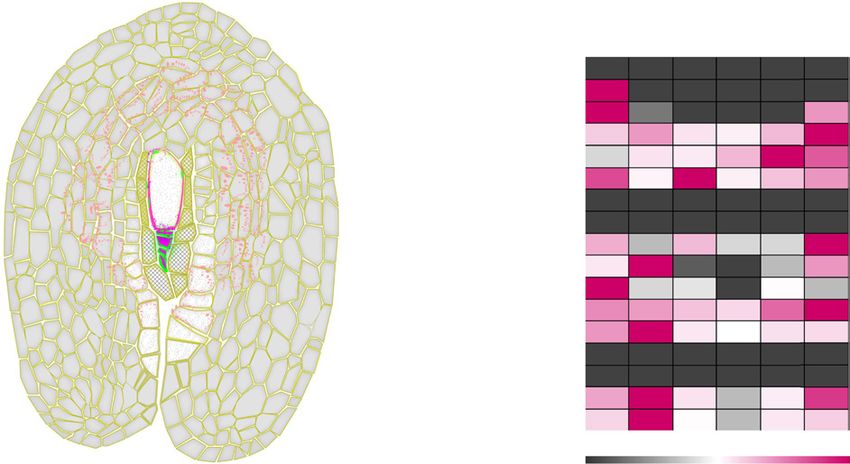
Juranić et al.
two to four nuclei, and randomly selected SO cells
from apomictic H. praealtum (R35). Our analysis indi-
cated that the Illumina-derived assemblies resulted in
a significant increase in gene models with read cover-
age over previous data generated with 454 sequenc-
ing by Okada et al. (2013). For example, the number
of gene models detected as expressed in AI cells har-
vested was 9 times higher than that found previously
using 454 sequencing (Okada et al., 2013; Rabiger et al.,
2016; Supplemental Table S1).
Differential expression analyses of gene models
between the three laser-captured cell types revealed
10,039 and 15,843 differentially expressed gene mod-
els in the AI cell transcriptome comparisons with EAE
sacs and SO cell transcriptomes, respectively (2-fold or
greater, P ≤ 0.01 corrected for multiple testing; Supple-
mental Table S2). Comparisons between EAE sacs and
SO cells showed 19,292 differentially expressed gene
models (Supplemental Table S2). These genes were
annotated to Arabidopsis and other databases with a
success rate ranging from 33% to 43% (Supplemental
Table S2), which is consistent with the annotation rate
observed by Rabiger et al. (2016) comparing whole tis-
sue samples. Full lists are provided in Supplemental
Tables S3 to S5.
These lists were further subdivided into increased
or decreased fold changes and analyzed independent-
ly for functional themes through the identification of
enriched Gene Ontogeny (GO) terms (Supplemental
Tables S6–S9). Both enriched and depleted functional
themes within the AI transcriptome relative to those
found in EAE sacs and SO cell transcriptomes are
summarized in Figure 2B. Shifts in GO terms across
the three cell types were explored for large changes in
both the diversity and significance of terms identified
under the parental GO term categories. These data
showed that the AI cell is highly metabolically active,
with increases in significance and diversity of enriched
GO terms relative to both SO cells and EAE sacs. Indi-
vidual enriched themes in the AI cell typically showed
varying degrees of enrichment in either SO cells or EAE
sacs. The AI cell shared enrichment of terms associated
Figure 2. Gene expression in laser-captured cell types from apomict with ribosome biogenesis, translation, protein modifi-
H. praealtum (R35) and identification of transcripts enriched in AI cells
cation, intracellular transport, substrate-specific trans-
following comparisons with mutants. A, Enlarged, uninucleate AI cells
were laser dissected from ovules isolated from stage 3/4 capitula (be- porters, ion channel activity, kinase activity, and DNA
tween stages 3 and 4; Koltunow et al., 1998; Okada et al., 2013). The
majority of stage 3/4 ovules contain multiple AI cells near cells under-
going FM selection. The remaining ovules contain four megaspores un- ATP, GTP binding; E, intracellular transport, substrate-specific trans-
dergoing degeneration (DM) and a functional AI cell (FAI), as shown. porters, ion channel activity; F, response to biotic and abiotic stimuli;
AI cells most distal to sexual events in stage 3/4 ovules were identified G, macromolecular complex; H, kinase activity, DNA replication; I,
in serial sections (dashed line) and captured to avoid contamination cell growth, differentiation, and morphogenesis; and J, regulation of
with either the FM or the functional AI cell displacing degenerating cell size and chromatin modification. C, Schematic of comparisons
megaspores. B, GO terms enriched in AI cells versus SO cells and AI for differentially expressed genes showing total numbers of gene mod-
cells versus EAE sacs from analyses of differential gene models. Bands els tested and the resulting numbers of significantly differential gene
represent individual GO terms grouped into parental terms by color. candidates (2-fold change, P ≤ 0.01). Comparisons between cell type
The increased width of each band shows the increased significance transcriptomes and the AI cell transcriptome are shown in red, com-
of enrichment, whereas the growth in the numbers of bands shows parisons between whole ovary samples of the R35 apomict wild type
increasing diversity of terms across the transition from SO cells to the and R35 sexual mutants (m115 and m134) are shown in blue, and the
AI cell and, consequently, to EAE sacs. GO terms are as follows: A, intersection of these sets of comparisons yields candidate gene lists
translation, protein modification, ribosome biogenesis; B, carboxylic of 50 and 14 genes. Note that the 14 genes form part of the set of 50
acid and hydrolase activity; C, carbohydrate biosynthetic process; D, genes.
1030 Plant Physiol. Vol. 177, 2018
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.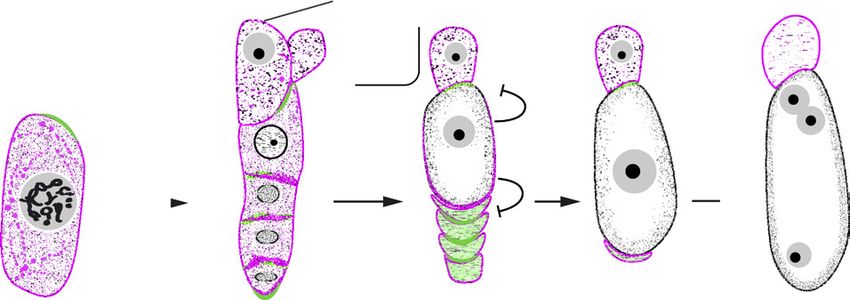
Cell Contact Promotes Apomixis in Hieracium
replication with SO cells. The AI cell demonstrated transcriptomes were compared with differential gene
enrichment in terms associated with response to biotic lists generated previously from comparisons between
and abiotic stimuli compared with EAE sacs. In con- whole ovary transcriptomes of the apomict R35 and
trast, the AI cell showed depletion in terms associated two independent apospory-deficient mutants, m115
with regulation of cell size and chromatin modification and m134 (Fig. 2C). These mutants cannot form AI cells
relative to SO cells, depletion in carbohydrate biosyn- and are unable to initiate apospory but competent-
thetic processes relative to EAE sacs, and depletion in ly form sexual female gametophytes (Rabiger et al.,
terms associated with cell membrane (lipid) and cell 2016). Therefore, comparisons of m115 and m134 ovary
wall (xylan and arabinan) catabolic processes relative to samples at the FM stage of female gametophyte devel-
both SO cells and EAE sacs. Substantial shifts also were opment (stage 3/4) with the cell type transcriptomes
observed in the specific GO terms contributing to the from apomict R35 ovules at the same developmental
parental GO category of cell growth, differentiation, and stage aimed to identify genes associated with LOA
morphogenesis. This was attributable to terms associ- function and increased expression in the enlarged AI
ated with chromosome organization and segregation, cells (Fig. 2C). Apospory-deficient mutants analyzed at
regulation of gene expression, pollen tube growth, and the FM stage shared 285 Hieracium D18 gene models
embryo sac development (Fig. 2B; Supplemental Table that displayed significantly decreased expression rel-
S9). Analysis of differential gene lists for protein secre- ative to R35. A total of 50 unique gene models (17.5%)
tion signals and protein subcellular localization did not were identified with higher differential expression in
reveal any distinct patterns (see “Materials and Meth- the AI cell relative to at least one of the other cell types
ods”; Supplemental Table S10). Genes related to meiosis in both mutants (Fig. 2C). In summary, 14 gene models
were not a feature of the AI cell transcriptome, support- had higher expression in the AI versus both SO cells
ing our prior findings (Okada et al., 2013). and EAE sacs, 13 had higher expression in the AI ver-
Transcripts of three genes shown previously to be sus EAE sacs but not SO cells, and 23 had higher ex-
up-regulated and detected in AIs and EAE sacs by pression in the AI versus SO cells but not EAE sacs in
in situ hybridization, which were not evident in 454- both mutants (Fig. 2C; Supplemental Table S11).
derived transcriptomes (Okada et al., 2013), were Only 16 of the 50 unique gene models identified were
present in the Illumina-sequenced set, supporting functionally annotated. Ribosomal protein elongation
factors, DNA/RNA polymerases, and UDP-glycosyl-
the improved depth of the data set. These genes are
transferase (UGT)-like genes were higher in expression
members of the response to biotic and abiotic stress
in the AI cell versus both EAE sacs and SO cells (Sup-
GO category and have homology to genes involved in
plemental Table S11). UGT enzymes transfer monosac-
drought stress and pathogen responses. They include
charides to elongating glycan chains and are required
an abscisic acid-induced RESPONSIVE TO DESIC-
for cell growth and the biosynthesis of plant cell wall
CATION22 (HpRD22)-like gene, a COILED COIL NU-
polysaccharides (Oikawa et al., 2013). Enzymes from
CLEOTIDE BINDING SITE LEUCINE RICH REPEAT this family also can transfer monosaccharides to pep-
(HpCC-NBS-LRR)-like disease resistance gene, and a tides, lipids, and hormones such as cytokinins. In the
putative LIPOXYGENASE (HpLOX)-like gene (Fig. 2B; latter case, glycosylation typically reduces the cytoki-
Supplemental Tables S3 and S4). We previously spec- nin activity (Wang et al., 2011; Šmehilová et al., 2016).
ulated that the aforementioned genes may function in The annotated genes with increased expression in
cell death or its amelioration, hypothetically playing AI cells versus EAE sacs and decreased expression in
a role in sexual pathway demise (Okada et al., 2013). both mutants included putative counterparts of mito-
Given the paucity of Hieracium megaspore-specific chondrial ATP synthases, ankyrin domain-containing
promoters, we tested if the infiltration of these genes proteins, and a sterol-acyl desaturase. A phenazine
into Nicotiana benthamiana leaves under the control of biosynthesis-like gene from Arabidopsis involved in
the cauliflower mosaic virus 35S promoter, either in- d-amino acid production also was identified in this
dividually or in mixed combinations, resulted in cell comparison (Supplemental Table S11). d-Amino ac-
death. Evidence for cell death induction or ameliora- ids can influence calcium ion influx via Glu receptor
tion in leaves was not observed, in contrast to the clear channels, which are required for plant reproduction
cell death response resulting from the infiltration of a and chemotaxis. Plus, high calcium concentrations
control rice CC-NBS-LRR gene, RGA4 (Supplemental are thought to promote FM selection (Qiu et al., 2008;
Fig. S1; Supplemental Materials and Methods; Supple- Michard et al., 2011; Strauch et al., 2015; Genchi, 2017;
mental Literature S1). The roles of these three genes in Ortiz-Ramírez et al., 2017). Expressed genes enriched
aposporous female gametophyte formation are still to in the AI cell versus SO cells and decreased in the apo-
be determined. spory-deficient mutants corresponded to a putative
acyl-CoA N-acetyltransferase, H+-transporting ATPase,
NAD(P)-linked oxidoreductase, and a reticulon-like
Cell Type and Apospory-Deficient Mutant Comparisons
gene with potential roles in plasmodesmata function
Identify Candidate LOA-Associated Pathways
and cell communication (Knox et al., 2015).
Genes with a significantly increased fold change Transcripts from an additional four candidate genes
in the AI cell relative to EAE sacs and SO cell type with possible linkage to known pathways involved in
Plant Physiol. Vol. 177, 2018 1031
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Juranić et al.
FM specification, selection, and megaspore death were whether the laser-captured AI cells were arrested at
found to be enriched in AI cells. The first, annotated either of these cell cycle phase transitions. Cell cycle-
as a purple acid phosphatase-like gene, is involved in related genes are highly conserved among angiosperms.
the degradation of xyloglucan and oligosaccharides Therefore, a reference set of core cell cycle genes from
as well as the increased synthesis of β-glucans such as Arabidopsis was used to identify a set of 97 putative Hi-
callose in transgenic tobacco (Nicotiana tabacum) cells eracium cell cycle homologs from the Hieracium resource
(Kaida et al., 2009, 2010). Another was an aspartic with various potential functions during cell cycle tran-
protease-like gene whose homologs have roles in the sitions. In total, 44 putative cyclins, 19 cyclin-dependent
promotion and inhibition of cell death in Arabidop- kinases (CDKs), nine CDK inhibitors (KRPs; interactor/
sis and in megaspore cell death in rice (Oryza sativa; inhibitor of cyclin-dependent kinase/Kip-related pro-
Ge et al., 2005; Chen et al., 2008). The remaining two teins), and 25 additional cell cycle regulators were iden-
were cytokinin oxidase-like transcripts found in the AI tified. Gene expression was analyzed in silico in SO
cell-versus-SO cell comparison and a type A response cells, AI cells, and laser-captured cell types from the
regulator found in the AI cell-versus-EAE sac compar- EAE sac, with a focus on the G1-to-S and the G2-to-M
ison, both with roles in cytokinin signal transduction transition phases of the cell cycle (Vandepoele et al.,
(Supplemental Table S11). Sporophytic cytokinin sig- 2002; Menges et al., 2005; Fig. 3; Supplemental Tables
naling is involved in FM specification in Arabidopsis, S12–S15; Supplemental Fig. S3).
and cytokinin also has a general role in cell prolifer- KRPs can negatively regulate cell division and block
ation (Kieber and Schaller, 2014). Cytokinin oxidase entry into both S and M cell cycle phases (De Veylder
catabolizes cytokinins to form inactive products, and et al., 2001; Zhou et al., 2002). Recently, they were
type A response regulators are activated by cytokinin shown to function in restricting the formation of one
in Arabidopsis; however, they subsequently inhibit cy- MMC and one FM per ovule in Arabidopsis (Zhao et
tokinin signaling (Kieber and Schaller, 2014). Enrich- al., 2017; Cao et al., 2018). KRPs block the G1-to-S tran-
ment of these latter two transcripts suggests that the sition by inhibiting the CDKA-CYCD complex, which
laser-captured AI cell may be desensitized to cytokinin is required by the E2F transcription factor to activate
signaling. the expression of S phase-related genes (Fig. 3). E2F
The identities of the UGT, cytokinin oxidase, gene targets encode proteins involved in DNA replica-
phenazine biosynthesis, aspartic protease, purple tion (e.g. CDC6, CDC45, MCM, PCNA, and RNR; Fig.
acid phosphatase, and type A response regulator-like 3; Vandepoele et al., 2005). The promoters of Hieraci-
genes were verified by RNA isolation from apomict um homologs of the putative E2F target genes identi-
Hieracium R35 ovaries and resequencing. Their gene fied contained E2F-binding sites (WTTSSCSS, where
expression profiles also were examined in ampli- W = A or T and S = C or G; Supplemental Table S15).
fied RNA from the ovule laser-captured cell types Transcripts of HpKRP9, a predicted homolog of Arabi-
and RNA isolated from roots and leaves of the R35 dopsis KRP6/7, were low and very low in abundance
apomict. Although the reactions were not strictly in AI cells and SO cells, respectively, suggesting that
quantitative, the UGT, purple acid phosphatase, and there may not be a block in the G1/S regulatory cas-
type A response regulator-like genes showed enrich- cade (Fig. 3; Supplemental Fig. S3). The expression of
ment in the AI laser-captured samples (Supplemental a Hieracium homolog of RETINOBLASTOMA-RELATED
Fig. S2). With the exception of purple acid phospha- PROTEIN1, which negatively regulates E2F transcription
tase, transcripts from the other verified genes also factor activity (Gutierrez et al., 2002), and a Hieracium
were evident in roots and leaves. Functional exam- homolog of AtCDC6, which is associated with endore-
ination of the roles of these proteins in sexual and duplication, also was low in AI and SO cells (Fig. 3,
asexual female gametophyte development will re- right; Castellano et al., 2001). By contrast, high ex-
quire the utilization of AI and FM cell type-specific pression levels of the predicted HpE2F target genes
promoters. Such promoters remain to be identified in that presumably enable the transition through to the
Hieracium spp. The promoter of the purple acid phos- S phase were observed in the AI cell, including Hp-
phatase-like gene, however, is a potential AI cell- CDC45, HpMCM2, HpPCNA2, and HpEBP1 (Fig. 3;
specific candidate. Supplemental Table S12). These in silico data suggest
that the harvested laser-captured AI cells progressed
beyond the G1/S checkpoint.
Laser-Captured AI Cells Appear to Be Arrested in the
Key regulators of the G2/M cell cycle transition are
S/G2 Phase of the Cell Cycle
shown in Figure 3, left. The expression of the puta-
There is evidence that cytokinin regulates the G1- tive Hieracium homolog of WEE1, which inhibits CY-
to-S phase transition and also the progression from G2 CA/B and CDK complexes required for the activation
to M in the plant cell cycle (Riou-Khamlichi et al., 1999; of three-repeat MYB, was low in all three cell types
Lipavská et al., 2011). The observed enrichment in AI (Fig. 3). MYB3R protein transcription factors can ei-
cells of UGT, cytokinin oxidase, and type A response ther promote or repress the transcription of genes in-
regulator transcripts, which may potentially reduce volved in the G2/M transition phase (Kobayashi et al.,
responses to cytokinin, prompted an examination of 2015). Only one out of the five putative MYB3R genes
1032 Plant Physiol. Vol. 177, 2018
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Cell Contact Promotes Apomixis in Hieracium
Figure 3. Expression of putative Hieracium cell cycle genes in three laser-captured ovule cell types from the aposporous
apomict, R35. A set of 97 putative cell cycle genes was isolated from the Hieracium genome, and the abundance of 45 of these
with possible roles in the transition through the G1-to-S and the G2-to-M cell cycle phase was examined in the three laser-
captured cell types (SO cells, AI cells, and EAE sacs). Yellow/white/black reflect normalized cpm values, with yellow indicating
the highest transcript abundance. The arrow within the cell cycle indicates the region where the laser-captured AI cells are likely
to be arrested. Supplemental Tables S12 and S13 provide expression metrics.
identified in Hieracium showed detectable expression Initial examination of the Hieracium survey genome
in laser-captured cell types, with the highest expression and transcriptome resource for sequences encod-
observed in the EAE sac samples. A number of MYB3R ing putative Carbohydrate Active Enzymes (CAZy;
gene targets are required for mitosis (e.g. KN, CYCB1, Lombard et al., 2014) showed that representatives of
CYCB2, IMK2, NACK1, CDC20.1, PLE, MAP65-1, and all CAZy families are present (Supplemental Fig. S4).
EDE1; Fig. 3, left). These candidates were isolated from Subsequent analyses of transcripts encoding enzymes
Hieracium, and their expression was significantly lower involved in sugar metabolism (Fig. 4B) indicated that
in the AI cells compared with SO cells, even though the expression of sucrose synthase (SuSy), fructoki-
they contained the requisite MSA elements (AACGG) nase (FRK), and sucrose phosphate synthase (SPS)
within their promoter regions (Supplemental Table transcripts was highest in the AI cell and EAE sacs.
S15). Collectively, these in silico analyses suggest that Distinct transcripts encoding SuSy isoforms were the
the majority of the laser-captured AI cells have pro- most prevalent transcript class (63%; Fig. 4C), suggest-
gressed through the G1/S checkpoint and are likely ing that expanding AI cells and EAE sacs may have
to be arrested in S/G2 (Fig. 3; Supplemental Tables a specific need for SuSy and its products. Transcripts
S12–S15). of several SuSy isoforms (HpSuSy3_1, HpSuSy4_2,
and HpSuSy4_4) showed increased abundance, coin-
ciding with the potential transition from SO cells to
Carbohydrate Metabolism Analyses Highlight Key
AI cells and subsequently to EAE sac formation (Fig.
Enzymes Involved in AI Cell and EAE Sac Growth
4B; Supplemental Table S16). The SO cell profile was
Various carbohydrate biosynthetic genes also exhib- distinct from that of the AI and EAE sac cell types, as
ited higher differential expression levels in the AI cell transcripts representing five distinct enzyme classes
relative to the other ovule cell types (Fig. 2B; Supple- involved in Suc metabolism were prevalent. They in-
mental Tables S3 and S4). In general, little is known cluded phosphoglucomutase (PGM) and Glc-6-P isom-
about cell type-specific carbohydrate metabolism erases, which act as interconverting enzymes of the
during female gametophyte development. To address hexose-phosphate pool, together with a sucrose phos-
this, the relative expression of genes encoding en- phate phosphatase (SPP), a homolog of HpSuSy4_3
zymes involved in nucleotide-sugar interconversion, and FRK. PGM was the most prevalent class (Fig. 4, B
Suc, starch, and cell wall polysaccharide metabolism and C). Thus, the isoforms involved in Suc metabolism
(Fig. 4A) was examined in the laser-captured cell types, appear to vary between the three laser-captured ovule
thereby providing insight into pathways involved in cell types, suggesting that the metabolic reactions may
AI cell and EAE sac growth (Fig. 4, B and C). be catalyzed by different cell type-specific enzymes.
Plant Physiol. Vol. 177, 2018 1033
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Juranić et al.
Figure 4. Suc, starch, and nucleotide-sugar metabolism pathways in Hieracium ovule cell types. A, Enzymes involved in the
interconversion of sugars and nucleotide sugars are indicated in small, black font. Sugar substrates are shown in larger black
font. Polysaccharides are indicated in italicized red font. Molecules highlighted with black boxes and white type are enriched
in AI cells and EAE sacs. B, Transcriptional dynamics of genes involved in Suc metabolism and nucleotide-sugar interconversion
in three ovule cell types from apomictic Hieracium R35. Putative Suc metabolism, starch metabolism, and nucleotide-sugar
interconversion genes are shown. The heat map with pink, white, and black colors shows relative transcript abundance (RTA),
where normalized cpm values were transformed to proportions of the maximum cpm value observed for each gene. Pink
1034 Plant Physiol. Vol. 177, 2018
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Cell Contact Promotes Apomixis in Hieracium
Transcripts encoding 11 putative starch-related other carbohydrate-related genes showed increased
enzymes were up-regulated in the AI and EAE sac abundance in m115, including gene transcripts encod-
transcriptomes (Fig. 4B). The majority of these en- ing UDP-Glc pyrophosphorylase3_1, HpSPP1_1, HpS-
coded starch synthase (SS) and β-amylase (Fig. 4C). BE2.1_2, HpUGE2_1, and HpGAE1_2 (Supplemental
HpSS1_2 and HpSS3_2 were up-regulated in AI cells, Fig. S5).
while the HpSS4_5 transcript was abundant in both the
AI cell and EAE sac samples relative to the SO cells.
Survey of Cell Wall Carbohydrate Epitopes in Apomict
By contrast, the number of genes involved in starch
Ovule Cells during Sexual Degeneration and the
breakdown was higher in SO cells compared with
Transition to Mitotic Aposporous Female Gametophyte
other starch-related activities. The greatest number of
Development
distinct transcripts was detected for β-amylase (46%),
while transcripts corresponding to SS, starch-branching A total of 17 monoclonal antibodies directed against
enzyme (SBE), and ADP-Glc pyrophosphorylase also cell wall carbohydrate epitopes were used to survey
were detected. Similar to those observed for Suc me- the presence of glycan motifs in reproductive cells in
tabolism, specific transcripts encoding different starch- ovules of two apomicts, R35 and D36, at stage 4 of flo-
related enzymes appear to be functionally recruited ral capitulum development in Hieracium spp. At this
by the different ovule cell types. The enlarged AI cells stage, all four sexual megaspores have typically de-
and EAE sacs appear to be investing energy into the generated and an enlarged functional AI cell is evident
synthesis of starch, possibly in preparation for down- and/or transitioning to mitosis (Fig. 5A). The selected
stream stages of female gametophyte development. antibodies detect epitopes in four major classes of cell
Transcripts encoding at least 31 nucleotide-sugar in- wall glycans, including glycan epitopes of glycopro-
terconverting enzymes were expressed in the different teins such as arabinogalactan proteins and extensins,
cell types: 15 genes were up-regulated in the AI cells hemicelluloses (e.g. xyloglucan and mannan), callose,
and/or EAE sacs and 15 were up-regulated in SO cells and pectins. These polymers are typically evident in
(Fig. 4B). Genes representing five different enzyme primary cell walls of tissues undergoing cell division,
families were enriched in the AI cell and EAE sac sam- expansion, and elongation.
ples, and the highest numbers of distinct transcripts Epitopes detected by the JIM4 and the LM8 to LM11
were detected for the UDP-Glc 6-dehydrogenase antibodies were not observed in ovule cells at this
(33%) and UDP-Glc 4-epimerase (UGE; 20%) classes. stage (Fig. 5). By contrast, pectin epitopes detected by
By contrast, transcripts from three families were en- LM5, LM6, LM19, and LM20 antibodies, and the gly-
riched in the SO cell sample, with the majority encod- coprotein epitopes detected by LM1, LM2, and LM14,
ing isoforms of UDP-Glc 4-epimerase (GAE; 47%) and were found in most ovule cell types (Fig. 5B). LM2-
UDP-Xyl synthase (40%; Fig. 4C). Thus, transcripts for epitope binding was prevalent in the integument cells,
nucleotide-sugar biosynthesis enzymes were preva- particularly in the chalazal region of the ovule (Fig. 5B;
lent in all samples analyzed (i.e. AI cells, EAE sacs, and Supplemental Fig. S6). The anti-xyloglucan antibody
SO cells), ensuring the availability of precursors for the LM15 predominantly labeled epitopes in micropylar
production of new cell wall polymers involved in their cell walls (Fig. 5B; Supplemental Fig. S5). LM19 and
growth and expansion. LM20 antibodies detected pectin epitopes in nucellar
The candidate genes discussed above also were in- epidermal cells, and LM19 epitopes were evident in
vestigated in ovaries from m115 and apomict R35 to the enlarging functional AI cell. Callose deposition,
determine if transcript levels were dependent on LOA detected by the BS-400-2 antibody, was evident in cell
function (Supplemental Fig. S5). Several genes that plates of dividing cells in the ovule, in patches of the
appeared to be up-regulated in AI cells and EAE sacs, functional AI cell wall, and was prevalent in the de-
relative to SO cells, showed decreased abundance in generating megaspores (Fig. 5A).
the apospory-deficient m115, including HpSuSy4_1, JIM8 and JIM13 antibodies detected epitopes in
HpSPS1F_1, HpSS3_2, HpSS4_3, HpGAE4_2, and the enlarging functional AI cell and in the degener-
HpUGE3_1 (Supplemental Fig. S5). Although the ap- ating megaspores, although higher signal intensities
parent down-regulation of these genes may be a result were observed using JIM13 (Fig. 5; Supplemental
of secondary mutations in the m115 background, it also Fig. S6). The epitope detected by JIM8 is unknown
may be due to the absence of AI cells in m115, consis- (Supplemental Table S17; Supplemental Literature S1).
tent with a function during the early stages of AI and JIM13 detects the trisaccharide β-GlcA-(1→3)-α-GalA-
EAE cell development. Curiously, transcripts of several (1→2)-Rha, indicative of arabinogalactans attached to
Figure 4. (Continued.)
indicates high abundance, and black indicates low abundance. The second heat map with green and black colors shows the
rank of each transcript in the cell type-specific LCM data sets, with green indicating high transcript abundance within the full
transcriptome data set for each independent cell type. C, Pie charts showing the proportion of putative transcripts encoding
enzymes involved in Suc metabolism, starch metabolism, and nucleotide-sugar biosynthesis in SO cells and reproductive cell
types. Values indicate percentages of unique transcripts analyzed.
Plant Physiol. Vol. 177, 2018 1035
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Juranić et al.
Figure 5. Cell wall glycans detected in apomictic H. piloselloides (D36) and H. praealtum (R35) ovules at stage 4 of capitulum
development. A, Schematic of a longitudinal section through a Hieracium ovule indicating cell types and the spatial patterning
of selected cell wall epitopes detected using antibodies specified in the color-coded key (inset). Nucellar epidermal cells are
hatched, the endothelial cells are colored white, and the integument cells are pale gray. DM, Four degenerating megaspores;
FAI, functional AI cell; NE, nucellar epidermis. B, Summary of the ovule cell type location and relative signal intensity of epi-
topes detected by the 17 antibodies in apomictic D36 and R35 ovules. A minimum of two sections in the plane shown in A
were examined per antibody tested. Supplemental Table S17 provides details of antibodies used and the epitopes they detect.
Supplemental Figure S6 provides an example set of micrographs supporting this figure.
some arabinogalactan protein backbones (Yates et the micropylar poles of the MMC (Fig. 6AIV). MMCs
al., 1996; Ellis et al., 2010; Supplemental Table S17; of m115 and both apomicts elongated significantly
Supplemental Literature S1). Antibodies to arabino- and progressed through meiosis at a slower rate than
galactan proteins can display cross reactivity with P36. Callose was prevalent in transverse megaspore
rhamnogalacturonan I (pectins) from a range of plant cell walls (Figs. 6 and 7, stages 3 and 3/4). Sexual P36
species (Pattathil et al., 2010). However, JIM13 specif- predominantly formed a T-shaped megaspore tetrad
ically detected epitopes in the expanding functional that had not been described previously (Figs. 1B and
AI cell in Hieracium ovules at stage 4, unlike the other
6CIV). Callose was evident in the wall of the enlarging
anti-pectin antibodies we examined, which detected
FM, and signal intensity diminished as it progressed
epitopes in cell walls of a range of ovule cell types
(Fig. 5B). Therefore, JIM13 was considered to be rec- through mitotic events of sexual female gametophyte
ognizing epitopes in Hieracium ovules from members formation (Supplemental Fig. S7). The FM featured an
of the large arabinogalactan protein family, including apical nucleus and a basal vacuole as it lost callose and
arabinogalactan (AG) peptides, classical arabinoga- the three nonselected megaspores degenerated. It sub-
lactan proteins, and various chimeric arabinogalactan sequently expanded rapidly, the nucleus then became
proteins (Ellis et al., 2010; Johnson et al., 2017a). We centrally positioned, and apical and basal vacuoles de-
refer to the epitope as an AGP hereafter for simplicity. veloped at cell poles before mitosis initiated.
AGPs detected by the JIM13 antibodies accumulat-
ed in the MMC walls of sexual and apomictic plants
Callose and AGP Patterning in Reproductive Cells and either preceded or coincided with callose accu-
during Sexual and Aposporous Female Gametophyte mulation. Therefore, JIM13 detects epitopes that act as
Development early markers of the sexual lineage in Hieracium (Figs.
6 and 7, stages 2 and 3). AGP epitopes were more prev-
JIM13 and anti-callose antibodies were used to fol-
alent in MMCs of sexual P36 and apomictic D36 than
low the patterning of AGP and callose epitopes in cells in MMCs of apomict R35 or m115. Therefore, high lev-
during female gametophyte development in sexual H. els of proteins containing JIM13-specific epitopes are
pilosella (P36), apomictic H. piloselloides (D36) and H. unlikely to be critical for the initiation and completion
praealtum (R35), and the apomictic mutant m115, which of meiotic events (Figs. 6, A and E, and 7, A and E).
forms sexual female gametophytes (Figs. 6 and 7). Cal- JIM13 epitopes colocalized with callose in transverse
lose deposition during sexual megasporogenesis in megaspore cell walls and also were distributed with-
Hieracium essentially followed previously described in the megaspores in a punctate manner. Colocation of
events, being detected first in the chalazal and then at callose and AGPs was evident in the expanding FM of
1036 Plant Physiol. Vol. 177, 2018
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Cell Contact Promotes Apomixis in Hieracium
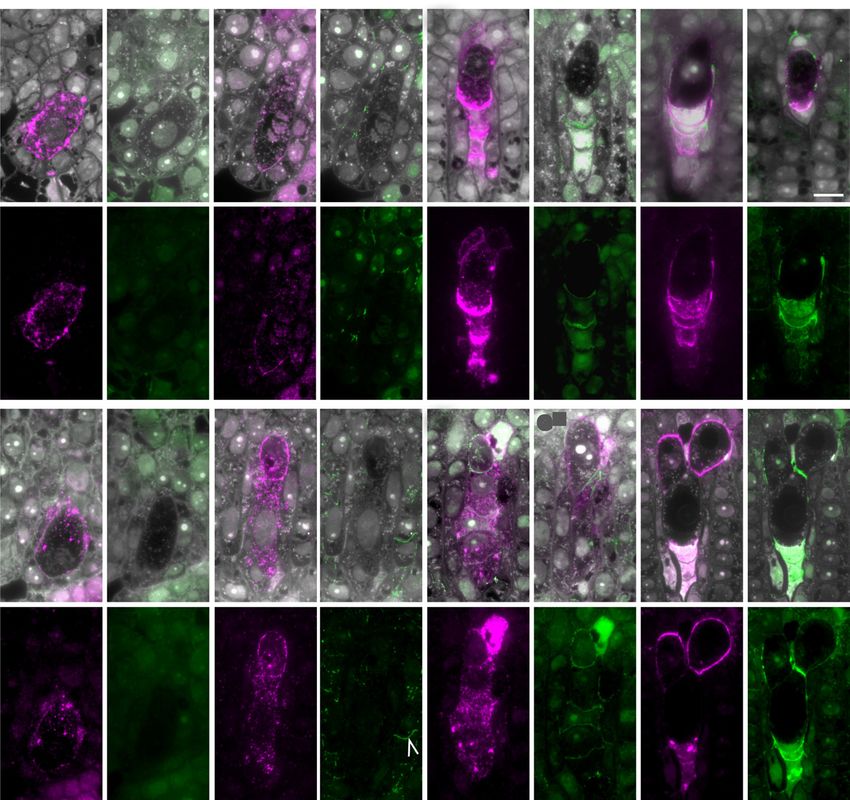
Figure 6. Cellular distribution of glycan epitopes detected by JIM13 and anti-callose (BS-400-2) antibodies during early female
gametophyte development in ovules of sexual P36 (A–D) and in mutant m115 with a sexual phenotype (E–H) at four indicated
stages. Numerals I and II indicate an identical section, with I representing the autofluorescence image merged with the flu-
orescent image detecting AGP epitopes shown in II. Numerals III and IV indicate identical sections, with III representing the
autofluorescence image merged with that detecting callose shown in IV. Sections are oriented with chalazal and micropylar
poles at top and bottom, respectively, in relation to ovule polarity. A, MMC (yellow arrowhead) in sexual P36 at the first meiotic
prophase. JIM13 epitopes are evident throughout the MMC. Callose is deposited in walls at both poles of the MMC; AIV shows
callose at the chalazal pole of the MMC. B, During megasporogenesis in P36, callose is evident primarily in the transverse walls
separating each megapospore and in cell walls at chalazal and micropylar apices of the developing tetrad. C, JIM13 epitopes
in the T-shaped tetrad of P36 accumulate in a polar manner primarily in transverse walls oriented toward the micropyle of the
ovule away from the FM (arrowhead). A T-shaped tetrad outlined in white forms in P36 with two adjacent megaspores evident
in the micropylar pole (CIV; see also Fig. 1B). The white arrowhead indicates a callose wall between two adjacent megaspores.
D, JIM13 epitopes and callose diminish during FM expansion and early mitotic events of megagametogenesis. Residual callose
remains evident at the chalazal apex. E, MMC in m115 with epitopes detected by JIM13 within the cell and a small amount of
callose at the chalazal pole. F, Elongated MMC (arrowhead) at first meiotic prophase with increased levels of epitopes detected
by JIM13. Callose was not detectable above background. Early meiosis is slower in the R35 mutant background relative to P36.
G, Linear tetrad in m115 showing the characteristic polar distribution of AGP epitopes detected by JIM13 in transverse mega-
spore cell walls oriented toward the micropyle. Callose is evident in the cell wall of the FM. H, Levels of both JIM13-detected
Plant Physiol. Vol. 177, 2018 1037
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Juranić et al.
all plants, and AGP levels diminished with callose as the other three megaspores and the formation of a
the FM underwent mitosis (Figs. 6 and 7). functional AI cell that subsequently underwent mito-
Importantly, the JIM13 antibody detected AGP epi- sis (Fig. 7, G and H). In some cases, supernumerary
topes in AI cells of both apomicts prior to their enlarge- AI cells in contact with the functional AI cell (Fig.
ment, representing the earliest known marker for AI 7GIII) exhibited collapsed cell walls and degenerat-
cell specification. AI cells developed close to sexual ed nuclei. In other cases, contact points between su-
cells and were positioned toward the chalazal region pernumerary AIs and an expanded functional AI cell
of the ovule. The timing of AI cell appearance was exhibited significant callose deposition (Fig. 7, HIII
earlier in R35, as AI cells formed near the MMC as it and HIV). Callose deposition between the functional
entered meiotic prophase (Fig. 7F). AGP-marked AI AI and supernumerary AI cells may function to limit
cells appeared later in D36 during megaspore tetrad the growth and numbers of aposporous female game-
formation. Callose was not detected in AI cells early tophytes formed from supernumerary AI cells. This
in their specification (Fig. 7, CIV and FIV), but it was is supported by the observation that, although up to
detected in AI cells at FM selection (Fig. 7GIV) and in four AI cells may form per ovule, a single mature apo-
the AI cells that remained undivided after megaspore sporous female gametophyte predominantly forms in
degeneration (Fig. 7HIV). AI cells marked with AGPs R35 and D36 ovules, while undivided AGP-marked AI
were not evident in the apomictic deletion mutant cells remain evident in the chalazal region. Two ma-
m115, confirming its sexual phenotype (Fig. 6, E–H). ture aposporous female gametophytes were observed
Collectively, these data show that AGPs are early in R35 and D36 apomicts in 13% and 10% of ovules,
markers of both the sexual and apomictic cell lineages respectively, prior to autonomous seed formation
in Hieracium spp., even though AI cells are incapable (n = 120 ovules per apomict examined at capitulum
of undergoing meiosis. Furthermore, meiosis initiation stage 10).
precedes AI cell specification in apomicts R35 and D36. AGP epitopes were not detected by JIM13 antibodies
in the walls of mitotically dividing sexual and apospo-
AI and FM Cell Contact Precedes Megaspore Demise and rous female gametophytes. However, AGPs became
Mitotic Formation of an Aposporous Female Gametophyte prevalent in cell walls of both gametophyte types
during cellularization and maturation. The micropy-
The distribution of callose and AGPs in cell walls of lar cells of the ovule where the pollen tube enters also
the sexual and aposporous female gametophyte lineag- were rich in AGPs (Supplemental Fig. S7). In summary,
es aided in establishing the sequence of events leading although the mitotic events of sexual and aposporous
to sexual female gametophyte mitosis and sexual cell female gametophyte formation begin with different
demise in apomicts D36 and R35. The number of AI progenitor cells in Hieracium spp., one meiotically de-
cells labeled with anti-AGP antibodies varied from one rived and the other a somatic cell, respectively, AGPs
to four in both apomicts, and they were located in the similarly mark the resultant mature gametophytes.
chalazal zone of the developing FM. In apomict D36, This supports the likely conservation of differentiation
an AGP-marked AI cell with no detectable cell wall cal- processes during sexual and asexual female gameto-
lose made direct contact with the FM (Fig. 7, CII and phyte maturation suggested from recent observations
CIV). The AI expanded rapidly, forming a functional AI in mature female gametophytes of apomictic Tarax-
cell resembling a meiotically derived FM cell in struc- acum spp. and strawberry using antibodies to detect
ture, and the FM degenerated (Fig. 7DI). AGP epitopes AGP epitopes (Gawecki et al., 2017; Leszczuk and
were lost in the functional AI cell as it enlarged rapidly, Szczuka, 2018) and our previous observations compar-
the nucleus became centrally located, and it initiated ing reporter gene expression in maturing sexual and
the mitotic events of aposporous female gametophyte aposporous female gametophytes in Hieracium (Tucker
development (Supplemental Fig. S7). Contact between et al., 2003).
the AI cell and the FM in apomict D36 also was ex-
amined and confirmed by confocal microscopy (Sup-
Diverse AGP Family Gene Types Are Expressed in
plemental Fig. S8; Supplemental Movie S1). In these
Early Sexual and Aposporous Female Gametophyte
analyses, the FM appeared to be elongating toward the
Development
AI cell to facilitate contact, implying a mutual attrac-
tion between the two cell types (Supplemental Fig. S8). The JIM13 antibody detects the oligosaccharide
However, it is also possible that this may be associated β-GlcA-(1→3)-α-GalA-(1→2)-Rha-GlcA-(1→3)-α-
with early FM cell degeneration events. GalA-(1→2)-Rha (Yates et al., 1996) that may occur
In the R35 apomict, direct contact of an AI with the in a range of protein backbones in AGP family mem-
FM also resulted in the demise of the FM along with bers early in sexual and aposporous Hieracium female
Figure 6. (Continued.)
AGP epitopes and callose diminish as the FM elongates and differentiates prior to initiating the mitotic events of megasporogen-
esis. eMMC, Elongated megaspore mother cell; MS, megaspores; TET, tetrad. A minimum of two biological replicates for each
species and all developmental stages were serially sectioned in these analyses. Bars = 10 μm.
1038 Plant Physiol. Vol. 177, 2018
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Cell Contact Promotes Apomixis in Hieracium
Figure 7. Cellular distribution of sugar epitopes detected by JIM13 and anti-callose (BS-400-2) antibodies during early female
gametophyte formation in ovules of apomicts D36 (A–D) and R35 (E–H) at four indicated stages. Designations for numerals
I to IV and section orientations are as for Figure 6. Exceptions are DIII and DIV and GIII and GIV, where III and IV are different
serial sections in the same ovule to highlight cells in a different plane. Yellow and red arrowheads indicate sexually derived
and aposporous cell types, respectively. A, Early MMC in apomict D36, where AGP epitopes are evident throughout the cell
and in the cell wall. Callose was not detected in these sections. B, An elongated MMC (yellow arrowhead) in meiotic prophase.
Early meiosis is slower in both apomicts relative to sexual P36 (Fig. 6B). JIM13 epitopes were detected, while callose was not
detected in the sections shown. C, Linear sexual megaspore tetrad in apomict D36. JIM13-detected epitopes are evident in
horizontal megaspore walls oriented toward the micropyle and within the megaspores. Two AI cells (red arrowheads) indis-
tinguishable in size from other somatic cells are marked with JIM13 epitopes. The AI at left is in contact with the FM. Callose
is evident in transverse megaspore walls. D, An elongated functional AI cell (red arrowhead) above four crushed megaspores
(yellow bracket) with a central nucleus and vacuoles at micropylar and chalazal poles resembling a sexual uninucleate me-
gagametophyte prior to first mitosis. In section DIII, a crescent-shaped AI cell is located at the apex of the FAI and marked with
callose (red arrowhead). The AI cell and the FAI are separated partially by a callose wall. E, Early MMC of apomict R35 showing
the pattern of JIM13-detected AGP epitopes; callose was not detected. F, Elongated MMC (yellow arrowhead) with AI cell (red
arrowhead). Both contain AGP epitopes. Significant callose was not observed in both cell types even though it was evident
in cell plates of dividing cells in the endothelium (white arrowhead in FIV). G, Elongated AI cell (red arrowhead) positioned at
the top of a megaspore tetrad (yellow bracket; GIV) and in contact with the enlarging FM (yellow arrowhead). JIM13-detected
Plant Physiol. Vol. 177, 2018 1039
Downloaded on March 26, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































