Hepatic stellate cell mediates transcription of TNFSF14 in hepatocellular carcinoma cells via H2S/CSE-JNK/JunB signaling pathway - Nature
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

www.nature.com/cddis
ARTICLE OPEN
Hepatic stellate cell mediates transcription of TNFSF14 in
hepatocellular carcinoma cells via H2S/CSE-JNK/JunB signaling
pathway
1,4 1✉
Yanan Ma , Shanshan Wang1,2,4, Yongle Wu1, Bihan Liu1, Lei Li1, Wenjing Wang2, Honglei Weng3 and Huiguo Ding
© The Author(s) 2022
Hepatic stellate cells (HSC) and hydrogen sulfide (H2S) both play important roles in the development of hepatocellar carcinoma
(HCC). Whereas, in the microenvironment of HCC, whether HSC participate in regulating the biological process of HCC cells by
releasing H2S remains elusive. In vitro, Flow cytometry (FCM), CCK-8, RNA-sequencing, Western blotting, RT-qPCR,
immunofluorescence and ChIP assays were carried out in the HCC cells to investigate the effect of H2S on biological functions and
JNK/JunB-TNFSF14 signaling pathway. Specimens from HCC patients were analyzed by RT-qPCR and Western blotting assays for
evaluating the expression of TNFSF14 and CSE. Statistical analysis was used to analyze the correlation between TNFSF14 expression
and clinical data of HCC patients. Based on the FCM and CCK-8 results, we found the LX-2 cells were able to induce HCC cells
1234567890();,:
apoptosis through releasing H2S. RNA-sequencing, RT-qPCR, and Western blotting results showed that TNFSF14 gene was
upregulated in both LX-2 and NaHS group. NaHS treated in HCC cells led to JNK/JunB signaling pathway activating and greater
binding of p-JunB to its responsive elements on TNFSF14 promoter. Impairment of TNFSF14 induction alleviated LX-2 and NaHS
induced apoptosis of HepG2 and PLC/PRF/5 cells. Furthermore, TNFSF14 expression in HCC tissues was lower than the adjacent
tissue. HCC patients with low expression of TNFSF14 had higher malignant degree and poor prognosis. In summary, demonstration
of the involvement of HSC-derived H2S in JNK/JunB mediated expression of TNFSF14 gene strongly indicates H2S palys an
important role in the regulation of HCC apoptosis.
Cell Death and Disease (2022)13:238 ; https://doi.org/10.1038/s41419-022-04678-z
INTRODUCTION and 3-mercapto-pyruvate sulfotransferase (MPST) [11]. Increasing
Nowadays hepatocellular carcinoma (HCC) is the second most lethal evidence shows that H2S affect HCC through the regulation of cell
cancer with persistently increasing mortality in the worldwide due to proliferation [12], migration and invasion [13], angiogenesis and
its highly complex and heterogeneous genetic aberrations [1, 2]. immune responses [14]. Intriguingly, our previous study found that
More and more researches suggest that tumor microenvironment exogenous H2S inhibit HCC progression through the PI3K/Akt/mTOR
(TME) plays an important role in the processes of hepatocarcinogen- signaling pathway in vitro [15]. Our previous data found that
esis, epithelial-mesenchymal transition (EMT), invasion, and metas- activated HSC can produce H2S mainly regulated by CSE. Nowadays,
tasis [3]. TME comprises various cells, including hepatic stellate cells the mechanisms involved in the capacity of HSC relased H2S to affect
(HSC), liver stromal endothelial cells (LSEC), immune cells and the HCC have not been worked out, all evidence up to this point
cytokines like extracellular matrix (ECM) [4]. suggests these mechanisms have great potential importance.
HSC are usually considered to foster the tumor microenvironment In this study, we investigate how activated HSC influence HCC
formation and contribute to a malignant tumor phenotype. Activated fate. We found that activated HSC released H2S to induce HCC cell
HSC contribute to hepatocarcinogenesis through the secretion of apoptosis. H2S induced cancer cell apoptosis through activating
growth factors, and producing ECM and matrix metalloproteinase JNK/JunB signaling, which upregulated tumor necrosis factor
(MMPs) [5–7]. But, there are more and more studies implicated that superfamily member 14 (TNFSF14) expression.
HSC could play an anti-tumor role in HCC progression [8, 9].
Hydrogen sulfide (H2S) is the most recently-discovered gas signal
molecular, participated in regulating cell proliferation, apoptosis, MATERIALS AND METHODS
oxidative stress, inflammation and angiogenesis [10]. H2S is mainly Patients
produced and cleared in the liver tissues by three enzymes: A total of 45 patients with HCC who received hepatectomy were
cystathionine-gamma-lyase (CSE), cystathionine-beta-synthase (CBS), enrolled from Beijing You’an Hospital, affiliated to Capital Medical
1
Department of Gastroenterology and Hepatology, Beijing You’an Hospital affiliated with Capital Medical University, Beijing 100069, China. 2Beijing Institute of Hepatology,
Beijing You’ An Hospital, Capital Medical University, Beijing 100069, China. 3Department of Medicine II, University Medical Center Mannheim, Medical Faculty Mannheim,
Heidelberg University, Mannheim 68167, Germany. 4These authors contributed equally: Yanan Ma, Shanshan Wang. ✉email: dinghuiguo@ccmu.edu.cn
Edited by Professor Boris Zhivotovsky
Received: 13 October 2021 Revised: 14 February 2022 Accepted: 18 February 2022
Official journal of CDDpress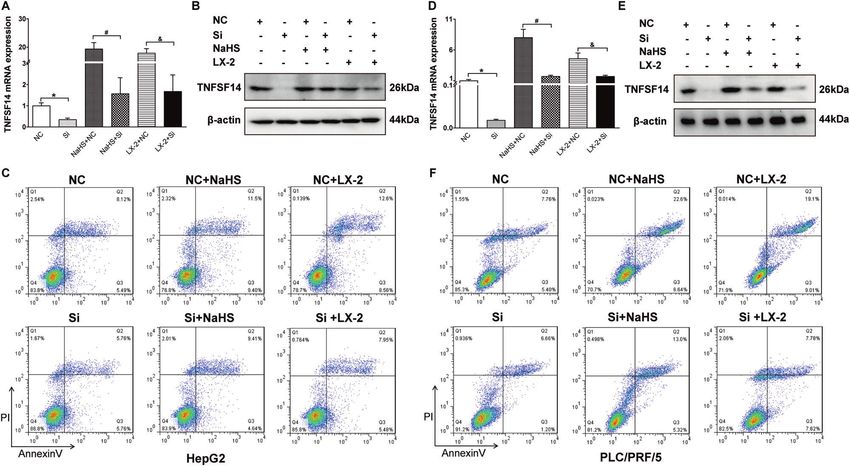
Y. Ma et al.
2
Table 1. Sequences of oligonucleotides used for RT-qPCR, ChIP-qPCR, Table 2. Correlation between TNFSF14 expression with
and RNA interference assays. clinicopathological characteristics of HCC.
Primers for RT-qPCR Sequences of oligonucleotides Feature Low expression of High expression of P-value
TNFSF14 F: 5′-CGTGAGACCTTCGCTCTTGTAT-3′ TNFSF14 (n = 22) TNFSF14 (n = 23)
R: 5′-CCCTCAGTGTTTGTGGTGGAT-3′ Gender
CSE F: 5′-AAGACGCCTCCTCACAAGGT-3′ Male 20 18 0.4140
R: 5′-ATATTCAAAACCCGAGTGCTGG-3′ Female 2 5
GAPDH F: 5′-TGAAGGTCGGAGTCAACGGA-3′ Age (years)
R: 5′-CCTGGAAGATGGTGATGGGAT-3′ ≤50 14 10 0.2362
Primers for ChIP-qPCR >50 8 13
TNFSF14 F: 5′-TTGTTCATTGCTGCATCCCC-3′ HBsAg
R: 5′-CTCCTCTTCTTCCGGTACCC-3′ Positive 20 18 0.4140
RNA interference assay Negative 2 5
TNFSF14-siRNA426 F: 5′-GACAGACCGACAUCCCAUUCATT-3′ Tumor size (cm)
R: 5′-UGAAUGGGAUGUCGGUCUGUCTT-3′ ≤3 7 10 0.5420
Scrambled siRNA F: 5′-UUCUCCGAACGUGUCACGUTT-3′ >3 15 13
R: 5′-ACGUGACACGUUCGGAGAATT-3′ Tumor number
Single 17 20 0.4591
University from July 2010 to January 2013. HCC was diagnosed and Multiple 5 3
graded according to the criteria of the American Association for the Portal invasion
Study of Liver Diseases [36]. Liver samples were collected from
cancerous and adjacent non-cancerous liver tissues and stored in liquid No 10 20 0.0045
nitrogen for further experimental use. Clinic pathologic characteristics Yes 12 3
of these patients are shown in Table S1. The study protocol was Serum AFP (ug/L)
approved by the Ethics Committee of Beijing You’an Hospital affiliated
to Capital Medical University. The informed consent was obtained from ≤20 10 11 >0.9999
all enrolled patients. >20 12 12
BCLC HCC stage
Cells A 3 11 0.0011a
PLC/PRF/5 (ATCC® Cat. No. CRL-8024TM), HepG2 (ATCC® Cat. No. HB-
8065TM) and LX-2 cells were cultured in DMEM medium supplemented B 5 9 0.0119a
with 10% FBS in a 37 °C incubator with 5% CO2. The Transwell insert C 14 3
system was used for co-culture experiments to analyze the effect of LX-2 a
Compared with BCLC stage C. AFP α-fetoprotein, HBsAg Hepatitis B surface
cells on HCC cells as previously described [37].
antigen, BCLC Barcelona Clinic Liver Cancer, HCC Hepatocellular carcinoma.
CCK-8
Cells proliferation was examined by a Cell Count KIT-8 assay (CCK8, Beijing, China). All the raw data were uploaded on the Gene Expression
AbMole, Houston, TX, America) according to the protocol. 2 × 103 cells with Omnibus (GSE193660).
different treatments were pipetted into 96-well microplates for measure-
ments. The absorbance values were measured under a wavelength of
450 nm on a Universal Microplate Reader (ELx800; Bio-Tek Instruments Inc, Quantitative real-time reverse transcription PCR (RT-qPCR)
Winooski, VT, USA). The percentage of cell viability was calculated as: Cell Total RNA was isolated by TRIzol reagent and subsequently subjected to
proliferation % = [(A450sample-background)/ (A450control-backgroud)] × reverse transcription (4368814, High-Capacity cDNA Reverse Transcrip-
100%. tion Kits, Thermo Fisher Scientific, Waltham, MA, USA). RT-qPCR was
performed using the TB Green (RR420A, Takara) on an ABI ViiA7 (GX-XVI
R2, Applied Biosystems Life Tech, USA). The relative mRNA expression
Flow cytometry levels were calculated with 2−ΔΔCt method and presented as mean fold-
Cell apoptosis was measured by FCM analysis using Annexin V-FITC/PI change of samples to control. The primers used in the study were listed
Apoptosis Detection Kit (40302ES20, Yeasen, Shanghai, China). Briefly, cells in the Table 1.
were collected and incubated with 5 µL Annexin V-FITC and 10 µL PI for
15 min at room temperature. Adjusted the volume to 500 µL with binding
buffer. The apoptosis of HCC cells was analyzed with a FACScan-420 flow Western blotting
cytometer (BD Biosciences, Franklin Lakes, NJ, USA). Cells or tissues were lysed in ice-cold lysis buffer. Total protein from cells or
tissues was extracted using RIPA lysis buffer (R0010, Solarbio, Beijing,
China). Nuclear and cytoplasmic fractions from cells were obtained by a
ELISA Nuclear Protein Extraction Kit (R0050, Solarbio, Beijing, China). Forty μg of
The concentrations of H2S were examined by a Hydrogen sulfide Assay Kit each protein were utilized for Western blotting. Membranes were blocked
(JEB-11780, Jin Yibai Biological Technology, Nanjing, China) according to by TBST with 5% nonfat milk (w/v) for 1 h at room temperature. Incubated
the instruction. the membranes with primary antibodies at 4 °C for overnight. Next day, the
membranes were incubated with horseradish peroxidase-conjugated
secondary antibodies for 1 h following washing with TBST. The immuno-
RNA sequencing
complexes were incubated with the enhanced chemiluminescent system
RNA sequencing analysis was conducted by Novogene (Beijing, China). In
(34580 Thermo Scientific, Waltham, MA, USA) and were subsequently
brief, total RNA was isolated by TRIzol reagent (15596026, Thermo Fisher
exposed by a Biorad Chemidoc XRS + imaging system (USA). Primary
Scientific, Waltham, MA, USA). The RNA quantification and qualification
antibodies against JNK(#9252), phospho-JNK (#4668), JunB (#3753),
were assessed using the RNA Nano 6000 Assay Kit of the Bioanalyzer
phospho-JunB (#8053), GAPDH (#2118), β-actin (#4967), and HDAC1
2100 system (Agilent Technologies, CA, USA). Sequencing of the libraries
(#34589) were purchased from Cell Signaling Technology (CST, USA).
was carried out using the Illumina NovaSeq 6000 platform (Novogene,
Cell Death and Disease (2022)13:238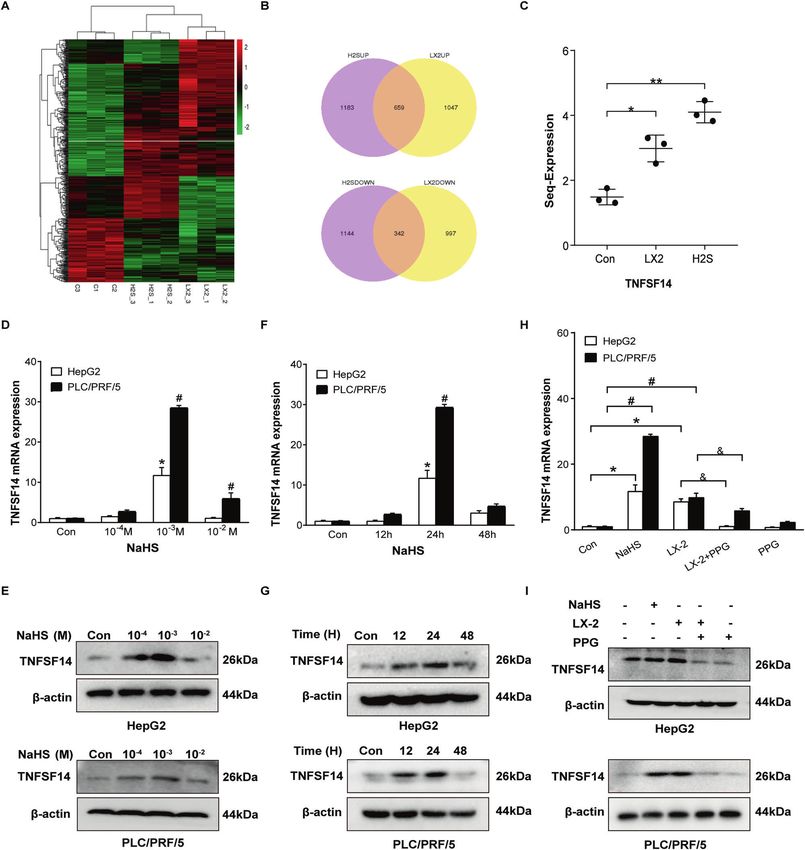
Y. Ma et al.
3
Fig. 1 Activated hepatic stellate cells influence HCC cells biological functions. A HepG2 and PLC/PRF/5 cells were collected after co-
cultured with LX-2 for 24 h, 48 h, 72 h, and 96 h respectively. CCK-8 assay was used to determine the viability of HepG2 and C PLC/PRF/5 cells.
B Flow cytometry was carried out to analyze the apoptotic rates of HepG2 and D PLC/PRF/5 cells. All Bars, ± SD; n = 3; *P < 0.05 compared with
control.
Fig. 2 LX-2 cells induce apoptosis of HCC cells through releasing H2S. The amount of H2S released by cultured LX-2 cells (A) and 10–3 M
NaHS (B) were measured by ELISA. C, D Flow cytometry was used to analyze the effects of exogenous H2S (NaHS) and endogenous H2S (LX-2
cells co-cultured or/and added PPG treated) on HCC cells apoptosis. All Bars, ± SD; n = 3; *P < 0.05 compared with D1 group in LX-2 cells,
#
P < 0.05 compared with D1 group in HepG2 cells.
TNFSF14 antibody (#AF0329) was purchased from Affinity Biosciences targeting the human TNFSF14 promoter containing JunB binding sites.
(USA). Horseradish peroxidase-conjugated goat anti-mouse (#7076) and The primers used for ChIP-PCR are listed in Table 1.
anti-rabbit (#7074) IgG secondary antibodies were purchased from CST.
Full and uncropped western blots were uploaded as ‘Supplemental
Material’. Immunofluorescence staining
Cells were fixed in ice-cold 4% paraformaldehyde solution. Then incubated
the cells with primary antibody (p-JunB) for overnight at 4 °C in the wet
Chromatin immunoprecipitation assay box. Next day, the cells were washed and subsequently incubated with the
The Chromatin immunoprecipitations (ChIP) assay was carried out secondary antibody conjugated with Alexa Fluor 488 for 1 h at room
according to a ChIP assay kit (#56383 CST). Cells were lysed with TNE temperature. Cell nuclei were counterstained with DAPI. Cells were viewed
buffer. Lysates were collected by centrifugation (15,000 g × 10 min) at 4 °C. and captured with a fluorescence microscope (Nikon Eclipse 80i; Nikon,
Aliquots were incubated with Protein G–Sepharose beads (GE Healthcare) Tokyo, Japan).
for 1 h. After centrifugation (1000 g × 5 min), the supernatants were
incubated with phospho-JunB (#8053 CST) and IgG(#2729 CST) antibodies Transient transfection
overnight at 4 °C. Next day, added Protein G–Sepharose beads for 8 h The small interfering RNAs (siRNAs) technique and TNFSF14-expressing
incubation. Then eluted the precipitate with the sample buffer. Finally, the plasmid were used to evaluate the effect of TNFSF14 on H2S-induced HCC cell
immunoprecipitated chromatin was analyzed by PCR using primers apoptosis. The siRNAs sequences were listed in the Table 1. Briefly, HCC cells
Cell Death and Disease (2022)13:238
Y. Ma et al.
4
Fig. 3 H2S upregulates pro-apoptotic factor TNFSF14 expression in HCC cells. A The heatmap. B Venn diagram of gene predictions based
on the control group and HepG2 cells treated with NaHS and co-cultured with LX-2 cells. C The expression levels of TNFSF14 mRNA changed
in HepG2 cells treated with NaHS and co-cultured with LX-2 cells. D RT-qPCR and (E) Western blotting were performed to examine the effects
of various concentrations of NaHS on expression of TNFSF14 gene in HepG2 and PLC/PRF/5 cells. F RT-qPCR and (G) Western blotting analysis
of the expression of TNFSF14 under 10−3 M NaHS treatment with different time in HepG2 and PLC/PRF/5 cells. H RT-qPCR and (I) Western
blotting were used to detect the effects of PPG treated on TNFSF14 expression in HCC cells. All Bars, ± SD; n = 3; *P < 0.05 compared with
control in HepG2 cells, #P < 0.05 compared with control in PLC/PRF/5 cells, &P < 0.05 compared with the co-culture group in HCC cells.
were seeded into the 6-well plates and cultured to the 80% confluence. information in the platform, the probes were converted into the gene symbols.
SiRNAs and the plasmids were transfected to HCC cells according the We divided the examined HCC patients into TNFSF14 high and low groups
instruction of LipofectamineTM 3000 transfection reagent (L3000-015, Thermo based on the median of the gene expression.
Fisher Scientific, Waltham, MA, USA). After 24 h of transfection, the expression
of TNFSF14 was evaluated by RT-qPCR and Western blotting.
Statistical analysis
Each experiment was repeated at least three times independently. All data
Data analysis of public databases were presented as mean ± S.D. performed using the SPSS software (version
The GSE14520 gene expression dataset was downloaded from the GEO 23.0). Differences between unpaired groups were compared by the
database (http://www.ncbi.nlm.nih.gov/geo). According the annotation Student’s t-test or Mann–Whitney U test. For paired groups, the Wilcoxon
Cell Death and Disease (2022)13:238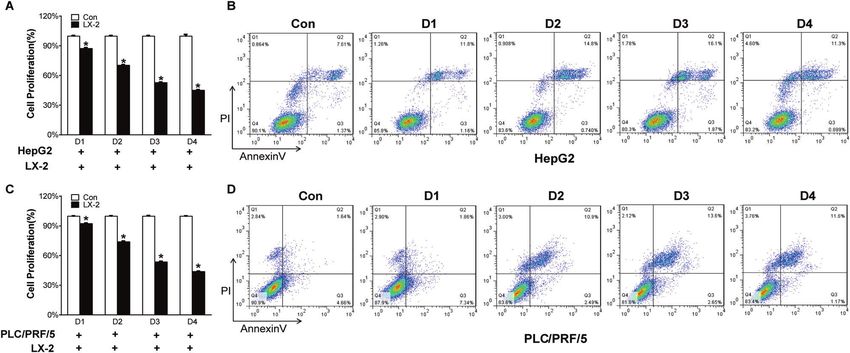
Y. Ma et al.
5
Fig. 4 TNFSF14 is indispensable for H2S-induced HCC cell apoptosis. TNFSF14 mRNA and protein expressions levels were evaluated by
(A, D) RT-qPCR and (B, E) Western blotting assays in HepG2 and PLC/PRF/5 cells under different treatments. Flow cytometry was carried out to
analyze the effects of different TNFSF14 expression levels on HepG2 (C) and PLC/PRF/5 (F) cells apoptosis. All Bars, ± SD; n = 3; *P < 0.05
compared with NC group, #P < 0.05 compared with NC + NaHS group, &P < 0.05 compared with NC + LX-2 group.
signed-rank test was used. Parametric or nonparametric multiple cells. Flow cytometry analyses showed that NaHS resulted in HCC
comparisons were performed using ANOVA-test or Kruskal–Wallis test. cell apoptosis (Fig. 2C, D). LX-2-dependent cancer cell apoptosis
Pearson’s χ2 test or Fisher’s exact test was used to analyze categorical data. was significantly reduced by the administration of PPG (Fig. 2C, D).
Correlations were assessed by the Spearman correlation. Survival analyses PPG alone did not affect HCC apoptosis (Fig. 2C, D). These results
were performed by the Kaplan–Meier and the log rank test. P < 0.05 was suggest that activated HSC result in HCC cell apoptosis through
considered significant.
releasing H2S.
H2S upregulates pro-apoptotic factor TNFSF14 expression in
RESULTS HepG2 cells
Activated hepatic stellate cells induce apoptosis of cancer To clarify how H2S results in HCC cell apoptosis, we performed
cells RNA-seq in HepG2 cells, which were co-cultured with LX-2 cells or
To examine how hepatic stellate cells (HSC) impact HCC cell treated with NaHS. A heat-map shows different transcriptome
behavior, we performed co-culture with activated HSC cell line LX- cluster in cells with different administrations (Fig. 3A). LX-2 co-
2 cells and HCC cell lines, HepG2 or PLC/PRF/5 cells. CCK-8 assays culture and NaHS treatment respectively led to 1047 and 1183
showed that cell viability of HCC cells was significantly reduced gene upregulation as well 997 and 1144 gene downregulation in
over time when the cells were co-cultured with LX-2 cells (Fig. 1A, HepG2 cells. Among these altered genes, 659 up-regulated genes
C). Flow cytometry analyses further demonstrated that co-culture and 342 down-regulated genes were presented in HepG2 cells
with LX-2 cells induced apoptosis of HepG2 and PLC/PRF/5 cells. treated with either LX-2 co-culutre or NaHS (Fig. 3B). Based on the
The maximal apoptosis of these cancer cells was observed at 72 h fold change, 30 genes with maximal fold changes were selected
following co-culture (Fig. 1B, D). These results suggest that to further analyze (Fig. S2). Among these 30 genes, we identified a
activated HSC not only inhibit HCC cell proliferation, but also pro-apoptotic gene TNFSF14 (Fig. 3C). We examined TNFSF14
induce apoptosis of cancer cells. mRNA and protein in HepG2 and PLC/PRF/5 cells treated with
different concentrations of NaHS. NaHS dose-dependently
LX-2 cells induce apoptosis of HCC cells through releasing H2S increased expression of TNFSF14 and reached a maximum at
How do activated HSC induce apoptosis of HCC cells? We 10–3 M treated 24 h in both HCC cells (Fig. 3D–G). Furthermore,
hypothesized that H2S is a crucial mediator in the process given we observed that TNFSF14 expression was up-regulated when
that activated HSC release H2S [16] and H2S is capable of inducing HCC cells were co-cultured with LX-2 cells (Fig. 3H, I). The effect
cancer cell apoptosis [15]. ELISA assay confirmed that LX-2 cells was inhibited by PPG administration (Fig. 3H, I). These results
increased H2S release over time (Fig. 2A). LX-2 cells released the imply that TNFSF14 might be the key factor mediating
same amount of H2S at 72 h as 10–3 M NaHS did at 24 h (Fig. 2B). H2S-induced HCC apoptosis.
Subsequently, we observed the effects of NaHS, a donor of H2S,
and DL-Propargylglycine (PPG), a CSE inhibitor, on LX-2 cell- TNFSF14 is indispensable for H2S-induced HCC cell apoptosis
dependent HCC cell apoptosis. As shown in Fig. S1, NaHS To confirm the role of TNFSF14 in H2S-induced HCC cell apoptosis,
increased H2S release, whereas PPG dose-dependently inhibited we knocked down TNFSF14 expression in both HepG2 and PLC/
mRNA and protein expression of CSE as well as H2S release in LX-2 PRF/5. RT-qPCR and Western blotting analyses confirmed that
Cell Death and Disease (2022)13:238
Y. Ma et al.
6
NaHS and LX-2 co-culture-induced TNFSF14 mRNA and protein expression was knocked down (Fig. 4C, F). On the other hand, the
expression was inhibited by siRNA-TNFSF14 in both HepG2 effect of TNFSF14 was also confirmed through transfection of the
(Fig. 4A, B) and PLC/PRF/5 cells (Fig. 4D, E). Flow cytometric tnfsf14 gene into HCC cells, which showed that transfection with
analyses further showed that NaHS and LX-2 co-culture-induced pcDNA3.1-tnfsf14 results in apparent increase of apoptosis in
apoptosis of cancer cells was significantly inhibited when TNFSF14 HepG2 and PLC/PRF/5 cells with different treatments (Fig. S3).
Cell Death and Disease (2022)13:238Y. Ma et al.
7
Fig. 5 H2S regulates TNFSF14 transcription through JNK/JunB signal. A Schematic overview of the predicted transcription factors of
TNFSF14 gene by PROMO dataset (upper) and JunB binding sites on TNFSF14 promoter predicted with JASPAR website (lower). B ChIP assays
for JunB binding onto TNFSF14 promoter were carried out in HepG2 (upper) and PLC/PRF/5 (lower) cells. The right panel showed the statistical
analysis for the ChIP assay. C NaHS was used to treat HepG2 (left) and PLC/PRF/5 (right) cells for 0, 1, 2, 8, and 24 h. Expression of p-JunB in
nuclear of HCC cells was detecting using immunofluorescence staining, and Nuclei and p-JunB were stained with DAPI (blue) and FITC (green),
respectively. The image is representative of three independent experiments. D The effects of different time of NaHS treatment on the
expression of p-JunB in the nuclear and cytoplasm were analyzed with Western blotting in the HepG2 cells and PLC/PRF/5 cells. E The
expression levels of p-JunB protein were detected by Western blotting in the HCC cells under exogenous and endogenous H2S treatment. F
Western blotting for the effects of PPG (CSE inhibitor) and SP600125 (p-JNK inhibitor) in the expressions of JNK/p-JNK, JunB/p-JunB, and
TNFSF14 proteins in HepG2 and PLC/PRF/5 cells. All Bars, ± SD; n = 3; *P < 0.05 compared with control group in HepG2 cells, #P < 0.05
compared with control group in PLC/PRF/5 cells.
These results suggest that TNFSF14 expression is required for hepatocyte growth factor (HGF), platelet-derived growth factor
H2S-induced HCC cell apoptosis. (PDGF) and connective tissue growth factor (CTGF), altering
components of the ECM, regulating angiogenesis and immune
H2S regulates TNFSF14 transcription through JNK/JunB signal surveillance [17–21]. In stark contrast, HSC are a source of
To clarify how H2S regulates the expression of TNFSF14, we transforming growth factor beta (TGF-β), which exert tumor-
analyzed PROMO dataset and adopted JASPAR software to predict suppressing activity by inducing cytostasis and apoptosis of
potential transcript factors binding to the TNFSF14 promoter. JunB hepatocytes in early tumor phases [8]. Recently, HSC were
got the highest scored among all the analyzed factors (Fig. 5A reported to be negative regulators of HCC progression through
upper panel) and possessed multiple binding sites on the TNFSF14 upregulating endosialin which is capable of inhibiting tumor-
promoter (Fig.5A lower panel). ChIP assays confirmed the binding promoting cytokines, including IGF2, RBP4, DKK1, and CCL5 [9].
of p-JunB to the TNFSF14 promoter in both HepG2 and PLC/PRF/5 The findings of our study shed further insight into the increasing
cells (Fig. 5B). The binding was increased by NaHS treatment (Fig. appreciation that HSC do not just act as pro-tumorigenic role, but
5B). Immunofluorescence staining and Western blotting showed may inhibit tumor progression. In our study, we found that HSC
that administration of NaHS remarkably induced p-JunB expres- cells co-cultured with HepG2 and PLC/PRF/5 cells could promote
sion and nuclear translocation in both HCC cell lines (Fig. 5C–D). cell apoptosis which was time-dependently manner. Further
Besides NaHS, LX-2 cell co-culture also increased p-JunB expres- research found that HSC cells through releasing H2S induced
sion and nuclear translocation (Fig. 5E). JunB phosphorylation and HCC cells apoptosis, which was relieved by adding PPG (CSE
nuclear translocation required upstream p-JNK. Administration of inhibitor) to reduce the amount of H2S.
SP600125, a p-JNK inhibitor, completely inhibited NaHS- or LX-2 As a gas signaling molecule, the role of H2S in HCC is still
cell co-culture-induced p-JunB expression and nuclear transloca- unclear. The previous reports shown that in the HepG2 and PLC/
tion in both cancer cells (Fig. 5F). The effects of NaHS- or LX-2 cell PRF/5 cells, endogenous H2S/CSE pathway regulated cell growth
co-culture-induced p-JunB expression and nuclear translocation mainly through EGFR pathway [12]. Another similar study found
was also inhibited by PPG (Fig. 5F). These results suggest that JNK/ that in the HepG2 cells, H2S/CSE pathway was activated after
JunB signaling mediates H2S to regulate TNFSF14 transcription. irradiated, which led to the long-term cell invasion and tumor
metastasis [22]. The function of exogenous H2S has also been
TNFSF14 expression is associated with the clinical outcome of studied in HCC cells. In the PLC/PRF/5 cells, 500 μM NaHS could
HCC induce cell proliferation, migration, and anti-apoptotic via activat-
Subsequently, we observed the association between TNFSF14 ing NF-κB pathway [13]. Other researchers found that exogenous
expression and clinical outcome of 45 HCC patients. TNFSF14 and H2S effectively restricted the tumor development in H22 HCC-
CSE mRNA expression were significantly downregulated in tumor bearing mice and HCC cell lines proliferation by blocking STAT3
tissues compared with surrounding non-tumor tissues (Fig. 6A). and NF-κB pathways [14, 23]. Similar with our pervious study,
Western blotting analyses performed in 6 patients confirmed the 10−3 M NaHS treated HepG2 and HLE cells could inhibit cells
results (Fig. 6B). CSE mRNA expressions in tumor tissues were proliferation, migration and pro-apoptotic via mTOR signaling
positively correlated with that of TNFSF14 (r = 0.438, P < 0.05, Fig. pathway [15]. In this study, our results showed that both
6C). We further investigated the association between TFNSF14 endogenous and exogenous H2S promoted HCC cells apoptosis,
expression and tumor characteristics. Patients with high TNFSF14 and the apoptosis was reduced when endogenous H2S was
expression in HCC showed less portal invasion, lower Barcelona inhibited by adding PPG.
Clinic Liver Cancer (BCLC) HCC stages and longer survival and How do H2S regulate HCC cells apoptosis? Based on the RNA-
disease-free time than those with low TNFSF14 expression (Table 2 sequencing results, we found that both endogenous and
and Fig. 6D, E). These results were similar as the data in GSE 14520: exogenous H2S could up-regulate TNFSF14 in HepG2 cell.
TNFSF14 and CSE gene expression were decreased in tumor TNFSF14, mainly expressed on activated T cells, is activated in
tissues compared with non-tumor tissues (Fig. S4). CSE gene Natural Killer (NK) cells, and immature dendritic cells (DC) [24].
expressions in HCC tissues were positively associated with Previous studies reported that TNFSF14 act as an immunomodu-
TNFSF14 expression (r = 0.168, PY. Ma et al.
8
Fig. 6 TNFSF14 expression is associated with the clinical outcome of HCC. A TNFSF14 mRNA and CSE mRNA expression levels in tumorous
tissues (HCC; N = 45) and nontumorous tissues (NT; N = 45). B Western blotting analysis for the expression of CSE and TNFSF14 in the
tumorous and nontumorous tissues in 6 patients with HCC. C Correlation between CSE mRNA expression levels and TNFSF14 mRNA
expression levels in tumorous tissues (N = 45). D, E Overall survival and disease-free survival curves showed that patients with high expression
levels of TNFSF14 have better prognosis. All Bars, ± SD; *P < 0.05, **P < 0.01, ***P < 0.001, ****P < 0.0001.
regulatory region of TNFSF14 gene was analyzed on the JASPAR DATA AVAILABILITY
website. We found transcription factor JunB binds to the TNFSF14 The datasets generated and/or analyzed during the current study are available from
gene. The transcriptional activity of JunB is enhanced through the corresponding author on reasonable request.
phosphorylation-JNK [32]. In HCC cells, JNK/JunB exerts anti-tumor
effects by promoting tumor cell apoptosis and inhibiting cell
growth [33]. In this study, we found that H2S regulated TNFSF14 REFERENCES
transcription mainly through activating JNK/JunB singal in both 1. Kocarnik JM, Compton K, Dean FE, Fu W, Gaw BL, Harvey JD, et al. Global
Burden of Disease 2019 Cancer Collaboration. Cancer incidence, mortality,
HepG2 and PLC/PRF/5 cells.
years of life lost, years lived with disability, and disability-adjusted life years for
All these in vitro results demonstrated that H2S, released by LX- 29 cancer groups from 2010 to 2019: A systematic analysis for the global
2 cells, participated in the JNK/JunB-TNFSF14 pathway to promote burden of disease study 2019. JAMA Oncol. 2021;e216987. https://doi.org/
HCC cell apoptosis. Further validation was performed in the GEO 10.1001/jamaoncol.2021.6987.E1-E24.
dataset and our cohort, the results showed that patients with high 2. Fujiwara N, Friedman SL, Goossens N, Hoshida Y. Risk factors and prevention of
CSE and TNFSF14 expression in HCC showed less portal invasion, hepatocellular carcinoma in the era of precision medicine. J Hepatol.
lower BCLC stages and longer survival and disease-free time 2018;68:526–49.
compared with lower TNFSF14 expression patients. Similar with 3. Sevic I, Spinelli FM, Cantero MJ, Reszegi A, Kovalszky I, García MG, et al. The role of
our results was found in the primary colorectal cancer patients, the tumor microenvironment in the development and progression of hepato-
cellular carcinoma. In: Tirnitz-Parker JEE, editor. Hepatocellular Carcinoma
TNFSF14 expression was decreased on tumor infiltrating lympho-
[Internet]. Codon Publications: Brisbane, AU, 2019, Chapter 2.
cytes in resected colorectal cancer liver metastases patients and 4. Tahmasebi Birgani M, Carloni V. Tumor microenvironment, a paradigm in hepa-
significantly associated with overall survival and recurrence-free tocellular carcinoma progression and therapy. Int J Mol Sci. 2017;18:405.
survival [34]. But in the nonalcoholic steatohepatitis (NASH)- 5. Rhee H, Kim HY, Choi JH, Woo HG, Yoo JE, Nahm JH, et al. Keratin 19 expression in
related HCC patients, NKT cells secreted TNFSF14 to promote hepatocellular carcinoma is regulated by fibroblast-derived HGF via a MET-ERK1/
NASH-to-HCC transition [35]. These discrepant researches sug- 2-AP1 and SP1 axis. Cancer Res. 2018;78:1619–31.
gested that TNFSF14 may play different roles in different liver 6. Yan C, Yang Q, Gong Z. Activation of hepatic stellate cells during liver carcino-
diseases. genesis requires Fibrinogen/Integrin αvβ5 in zebrafish. Neoplasia.
It is presently unclear how significant the impact of H2S 2018;20:533–42.
7. Scheau C, Badarau IA, Costache R, Caruntu C, Mihai GL, Didilescu AC, et al. The
mediated TNFSF14 expression through JNK/JunB pathway on the
role of matrix metalloproteinases in the epithelial-mesenchymal transition of
occurrence and high mortality of HCC. However, taking into hepatocellular carcinoma. Anal Cell Pathol. 2019;2019:9423907.
consideration the effects of H2S in the HCC, it is reasonable to 8. Dewidar B, Meyer C, Dooley S, Meindl-Beinker AN. TGF-β in hepatic stellate cell
speculate that H2S has great potential significance and impor- activation and liver fibrogenesis-Updated 2019. Cells. 2019;8:1419.
tance in HCC apoptosis. Taken our results together, HSC released 9. Mogler C, König C, Wieland M, Runge A, Besemfelder E, Komljenovic D, et al.
H2S could upregulate the TNFSF14 expression through JNK/JunB Hepatic stellate cells limit hepatocellular carcinoma progression through the
axis to induce HCC cells apoptosis, and TNFSF14 was significantly orphan receptor endosialin. Embo Mol Med. 2017;9:741–9.
associated with prognosis of HCC patients. Thus, enhance the 10. Szabo C, Papapetropoulos A. International union of basic and clinical pharma-
circulating H2S to promote cells apoptosis may be a potential cology. CII: pharmacological modulation of H2S levels: H2S donors and H2S bio-
synthesis inhibitors. Pharm Rev. 2017;69:497–564.
strategy for HCC. Nonetheless, further clinical study of this
11. Mani S, Cao W, Wu L, Wang R. Hydrogen sulfide and the liver. Nitric Oxide.
approach will be required. 2014;41:62–71.
Cell Death and Disease (2022)13:238Y. Ma et al.
9
12. Pan Y, Ye S, Yuan D, Zhang J, Bai Y, Shao C. Hydrogen sulfide(H2S)/cystathionine 35. Wolf MJ, Adili A, Piotrowitz K, Abdullah Z, Boege Y, Stemmer K, et al. Metabolic
γ-lyase (CSE) pathway contributes to the proliferation of hepatoma cells. Mutat activation of intrahepatic CD8+ T cells and NKT cells causes nonalcoholic stea-
Res. 2014;763–764:10–8. tohepatitis and liver cancer via cross-talk with hepatocytes. Cancer Cell.
13. Zhen Y, Pan W, Hu F, Wu H, Feng J, Zhang Y, et al. Exogenous hydrogensulfide 2014;26:549–64.
exerts proliferation/anti-apoptosis/angiogenesis/migration effects via amplifying 36. Marrero JA, Kulik LM, Sirlin CB, Zhu AX, Finn RS, Abecassis MM, et al. Diagnosis,
the activation of NF-κB pathway in PLC/PRF/5 hepatoma cells. Int J Oncol. staging, and management of hepatocellular carcinoma: 2018 practice guidance
2015;46:2194–204. by the American association for the study of liver diseases. Hepatology.
14. Yang D, Li T, Li Y, Zhang S, Li W, Liang H, et al. H2S suppresses indoleamine 2, 3- 2018;68:723–50.
dioxygenase 1 and exhibits immunotherapeutic efficacy in murine hepatocellular 37. Li ZQ, Wu WR, Zhao C, Zhao C, Zhang XL, Yang Z, et al. CCN1/Cyr61 enhances the
carcinoma. J Exp Clin Cancer Res. 2019;38:88. function of hepatic stellate cells in promoting the progression of hepatocellular
15. Wang SS, Chen YH, Chen N, Wang LJ, Chen DX, Weng HL, et al. Hydrogen sulfide carcinoma. Int J Mol Med. 2018;41:1518–28.
promotes autophagy of hepatocellular carcinoma cells through the PI3K/Akt/
mTOR signaling pathway. Cell Death Dis. 2017;8:e2688.
16. Damba T, Zhang M, Buist-Homan M, van Goor H, Faber KN, Moshage H. Hydrogen ACKNOWLEDGEMENTS
sulfide stimulates activation of hepatic stellate cells through increased cellular This work was supported by the National Natural Science Foundation (81672725,
bio-energetics. Nitric Oxide. 2019;92:26–33. 81970525), Beijing Natural Science Foundation(7212052), Beijing Hospitals Authority
17. Cho Y, Park MJ, Kim K, Park JY, Kim J, Kim W, et al. Tumor-stroma crosstalk Youth Programme (QML20211701), Sino-German Cooperation Group (GZ1517), and
enhances REG3A expressions that drive the progression of hepatocellular carci- Sino-German Mobility program (M-0200). The authors express their gratitude to the
noma. Int J Mol Sci. 2020;21:472. National Natural Science Foundation (81672725, 81970525), Beijing Natural Science
18. Makino Y, Hikita H, Kodama T, Shigekawa M, Yamada R, Sakamori R, et al. CTGF Foundation(7212052), Beijing Hospitals Authority Youth Programme (QML20211701),
mediates tumor-stroma interactions between hepatoma cells and hepatic stellate Sino-German Cooperation Group (GZ1517), and Sino-German Mobility program(M-
cells to accelerate HCC progression. Cancer Res. 2018;78:4902–14. 0200) for financial support.
19. Li Q, Wang C, Wang Y, Sun L, Liu Z, Wang L, et al. HSCs-derived COMP drives
hepatocellular carcinoma progression by activating MEK/ERK and PI3K/AKT sig-
naling pathways. J Exp Clin Cancer Res. 2018;37:231.
20. Morse MA, Sun W, Kim R, He AR, Abada PB, Mynderse M, et al. The role of AUTHOR CONTRIBUTIONS
angiogenesis in hepatocellular carcinoma. Clin Cancer Res. 2019;25:912–20. HD designed and supervised the study; YM, SW, YW, BL, LL, and WW conducted
21. Hsieh CC, Hung CH, Chiang M, Tsai YC, He JT. Hepatic stellate cells enhance liver experiments or interpreted the data; HW provided technical support; YM and SW
cancer progression by inducing myeloid-derived suppressor cells through wrote the paper. All authors read and approved the final paper.
interleukin-6 signaling. Int J Mol Sci. 2019;20:5079.
22. Zhang H, Song Y, Zhou C, Bai Y, Yuan D, Pan Y, et al. Blocking endogenous H2S
signaling attenuated radiation-induced long-term metastasis of residual HepG2 COMPETING INTERESTS
cells through inhibition of EMT. Radiat Res. 2018;190:374–84. The authors declare no competing interests.
23. Lu S, Gao Y, Huang X, Wang X. GYY4137, a hydrogen sulfide (H2S) donor, shows
potent anti-hepatocellular carcinoma activity through blocking the STAT3 path-
ETHICS STATEMENT
way. Int J Oncol. 2014;44:1259–67.
The study protocol was approved by the Ethics Committee of Beijing You’an Hospital
24. Skeate JG, Otsmaa ME, Prins R, Fernandez DJ, Da Silva DM, Kast WM. TNFSF14:
affiliated to Capital Medical University. The informed consent was obtained from all
Lighting the way for effective cancer immunotherapy. Front Immunol.
enrolled patients.
2020;11:922.
25. Harrop JA, McDonnell PC, Brigham-Burke M, Lyn SD, Minton J, Tan KB, et al.
Herpesvirus entry mediator ligand (HVEM-L), a novel ligand for HVEM/TR2, sti-
mulates proliferation of T cells and inhibits HT29 cell growth. J Biol Chem.
ADDITIONAL INFORMATION
1998;273:27548–56. Supplementary information The online version contains supplementary material
26. Zou W, Zheng H, He TC, Chang J, Fu YX, Fan W. LIGHT delivery to tumors by available at https://doi.org/10.1038/s41419-022-04678-z.
mesenchymal stem cells mobilizes an effective antitumor immune response.
Cancer Res. 2012;72:2980–9. Correspondence and requests for materials should be addressed to Huiguo Ding.
27. Qiao G, Qin J, Kunda N, Calata JF, Mahmud DL, Gann P, et al. LIGHT elevation
enhances immune eradication of colon cancer metastases. Cancer Res. Reprints and permission information is available at http://www.nature.com/
2017;77:1880–91. reprints
28. Han B, Wu LQ, Ma X, Wang ZH, Li JP, Bi CY, et al. Synergistic effect of IFN-γ gene
on LIGHT-induced apoptosis in HepG2 cells via down regulation of Bcl-2. Artif Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims
Cells Blood Substit Immobil Biotechnol. 2011;39:228–38. in published maps and institutional affiliations.
29. Zhu X, Su D, Xuan S, Ma G, Dai Z, Liu T, et al. Gene therapy of gastric cancer using
LIGHT-secreting human umbilical cord blood-derived mesenchymal stem cells.
Gastric Cancer. 2013;16:155–66.
30. Hu X, Zimmerman MA, Bardhan K, Yang D, Waller JL, Liles GB, et al. Lymphotoxin
beta receptor mediates caspase-dependent tumor cell apoptosis in vitro and Open Access This article is licensed under a Creative Commons
tumor suppression in vivo despite induction of NF-kappaB activation. Carcino- Attribution 4.0 International License, which permits use, sharing,
genesis. 2013;34:1105–14. adaptation, distribution and reproduction in any medium or format, as long as you give
31. Kim YS, Nedospasov SA, Liu ZG. TRAF2 plays a key, nonredundant role in LIGHT- appropriate credit to the original author(s) and the source, provide a link to the Creative
lymphotoxin beta receptor signaling. Mol Cell Biol. 2005;25:2130–7. Commons license, and indicate if changes were made. The images or other third party
32. Cargnello M, Roux PP. Activation and function of the MAPKs and their substrates, material in this article are included in the article’s Creative Commons license, unless
the MAPK-activated protein kinases. Microbiol Mol Biol Rev. 2011;75:50–83. indicated otherwise in a credit line to the material. If material is not included in the
33. Guo C, Liu Q, Zhang L, Yang X, Song T, Yao Y. Double lethal effects of fusion gene article’s Creative Commons license and your intended use is not permitted by statutory
of wild-type p53 and JunB on hepatocellular carcinoma cells. J Huazhong Univ regulation or exceeds the permitted use, you will need to obtain permission directly
Sci Technol Med Sci. 2012;32:663–8. from the copyright holder. To view a copy of this license, visit http://creativecommons.
34. Maker AV, Ito H, Mo Q, Weisenberg E, Qin LX, Turcotte S, et al. Genetic evidence org/licenses/by/4.0/.
that intratumoral T-cell proliferation and activation are associated with recur-
rence and survival in patients with resected colorectal liver metastases. Cancer
Immunol Res. 2015;3:380–8. © The Author(s) 2022
Cell Death and Disease (2022)13:238You can also read

















































