HMOF Histone Acetyltransferase Is Required for Histone H4 Lysine 16 Acetylation in Mammalian Cells
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
MOLECULAR AND CELLULAR BIOLOGY, Aug. 2005, p. 6798–6810 Vol. 25, No. 15
0270-7306/05/$08.00⫹0 doi:10.1128/MCB.25.15.6798–6810.2005
Copyright © 2005, American Society for Microbiology. All Rights Reserved.
hMOF Histone Acetyltransferase Is Required for Histone H4 Lysine
16 Acetylation in Mammalian Cells
Mikko Taipale,1 Stephen Rea,1 Karsten Richter,2 Ana Vilar,3,4 Peter Lichter,2 Axel Imhof,3
and Asifa Akhtar1*
European Molecular Biology Laboratory, Gene Expression Programme, Meyerhofstrasse 1, 69117 Heidelberg, Germany1;
Division of Molecular Genetics (B060), Deutsches Krebsforschungszentrum, INF 280, 69120 Heidelberg, Germany2;
Adolf-Butenandt-Institut, Schillerstrasse 44, 80336 München, Germany3; and Cancer Epigenetics Laboratory,
Molecular Pathology Programme, Spanish National Cancer Centre,
Melchor Fernández Almagro 3, 28029 Madrid, Spain4
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
Received 18 March 2005/Returned for modification 11 April 2005/Accepted 5 May 2005
Reversible histone acetylation plays an important role in regulation of chromatin structure and function.
Here, we report that the human orthologue of Drosophila melanogaster MOF, hMOF, is a histone H4 lysine
K16-specific acetyltransferase. hMOF is also required for this modification in mammalian cells. Knockdown
of hMOF in HeLa and HepG2 cells causes a dramatic reduction of histone H4K16 acetylation as detected by
Western blot analysis and mass spectrometric analysis of endogenous histones. We also provide evidence that,
similar to the Drosophila dosage compensation system, hMOF and hMSL3 form a complex in mammalian cells.
hMOF and hMSL3 small interfering RNA-treated cells also show dramatic nuclear morphological deforma-
tions, depicted by a polylobulated nuclear phenotype. Reduction of hMOF protein levels by RNA interference
in HeLa cells also leads to accumulation of cells in the G2 and M phases of the cell cycle. Treatment with
specific inhibitors of the DNA damage response pathway reverts the cell cycle arrest caused by a reduction in
hMOF protein levels. Furthermore, hMOF-depleted cells show an increased number of phospho-ATM and
␥H2AX foci and have an impaired repair response to ionizing radiation. Taken together, our data show that
hMOF is required for histone H4 lysine 16 acetylation in mammalian cells and suggest that hMOF has a role
in DNA damage response during cell cycle progression.
Nucleosomes composed of DNA and histones define the as well as a C2HC-type zinc finger. Recently, the crystal struc-
fundamental structural unit of chromatin, which acts as a scaf- ture of Esa1, an essential yeast HAT, showed that even though
fold for nuclear processes such as transcription and replication. the MYST and GNAT family share sequence homology only in
By modifying the nucleosomal structure in several ways, the motif A, there is high degree of structural conservation in the
chromatinized DNA can be made either more or less accessi- central core region between the two families (59, 60). The
ble. Alterations to chromatin structure are usually brought MYST family can be further divided into subgroups based on
about in three different ways: by ATP-dependent remodeling additional domains present in these proteins. The first sub-
of nucleosomes, by replacement of standard histones with hi- group contains proteins with PhD fingers (such as MOZ and
stone variants, and by covalently modifying the N-terminal tails MORF), the second subgroup contains proteins with a chro-
of histones. Histone modifications include acetylation, phos- modomain (such as Esa1, dMOF, and Tip60) (41, 56), and a
phorylation, methylation, ubiquitination, sumoylation, and third one (including HBO1) has other known domains such as
poly(ADP-ribosyl)ation (for a review, see reference 54). His- zinc fingers.
tone acetylation is the best-characterized modification and is
One of the chromodomain-containing members of the
controlled by histone acetyltransferases (HATs) and histone
MYST family, dMOF, is an integral player in the Drosophila
deacetylases.
melanogaster dosage compensation process. Dosage compen-
Sequence analysis of HAT proteins reveal that they can be
sation ensures that males and females, despite unequal num-
classified in distinct families, with each family having a char-
bers of X chromosomes, express the same amount of X-linked
acteristic substrate specificity (12). GNAT (GCN5-related N-
acetyltransferases) family members mainly acetylate lysines on gene products. In Drosophila, this is thought to occur by an
the histone H3 tail. The founding members of the other family, approximately twofold transcriptional upregulation of most
MYST, include Saccharomyces cerevisiae Ybf2p/Sas3p and male X-linked genes. The male X chromosome is coated by
Sas2p and human MOZ and Tip60 (56). The MYST homology dosage compensation complex (DCC), comprised of at least
domain is exceptionally well conserved among all family mem- five proteins (dMOF, dMSL1, dMSL2, dMSL3, and dMLE)
bers. This region includes the acetyl coenzyme A binding do- and two noncoding RNAs (roX1, roX2). Transcriptional up-
main similar to the one found in GNAT acetyltransferases (41) regulation correlates with specific acetylation of histone H4 at
lysine 16 (H4K16) on the male X chromosome by dMOF (3, 8,
51). A point mutation in a conserved glycine residue of dMOF
* Corresponding author. Mailing address: European Molecular Bi-
that renders the protein enzymatically inactive leads to male-
ology Laboratory, Gene Expression Programme, Meyerhofstrasse 1,
69117 Heidelberg, Germany. Phone: 49 6221 3878550. Fax: 49 6221 specific lethality (19). Biochemical characterization of the
3878518. E-mail: akhtar@embl.de. dMOF has shown that it is an RNA-binding protein that acety-
6798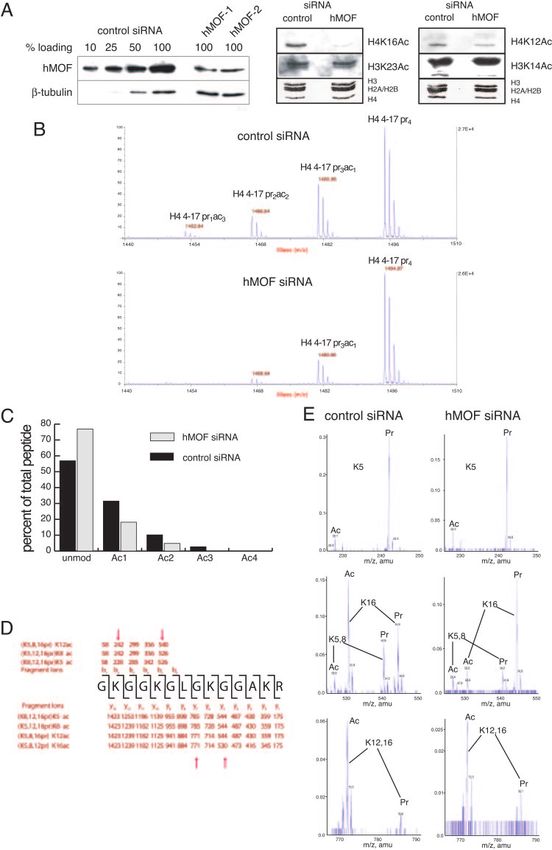
VOL. 25, 2005 hMOF AND H4K16 ACETYLATION IN MAMMALS 6799
lates not only histone H4 lysine 16 but also other members of KWAPPKHKQVKLSKK at Eurogentec, Belgium. Antibody was affinity purified
the DCC, namely dMSL1 and dMSL3 (4, 11, 31). with the peptide KWAPPKHKQVKLSKK. Antibody against RNA helicase A
was obtained from C.-G. Lee (University of Medicine and Dentistry, New Jer-
It is remarkable that all the proteins of the Drosophila DCC sey), MRG15 was obtained from O. Pereira-Smith (University of Texas, San
have been well conserved during evolution (26, 27, 45), even Antonio), RCC1 was obtained from I. Mattaj (EMBL, Heidelberg), lamin A/C
though dosage compensation is brought about by different and lamin B1 were obtained from H. Herrmann (DKFZ, Heidelberg), and
means in other animal phyla. Based on current evidence, it is H4K12Ac was obtained from Bryan Turner (University of Birmingham). -Tu-
bulin and FLAG (Sigma), ATM pS1981 (Rockland Immunochemicals),
reasonable to assume that dosage compensation has evolved
H4K16Ac (Chemicon), H3K14Ac and H3K23Ac (Abcam), and ␥H2AX (Up-
independently several times, illustrating an interesting case of state) were purchased as indicated.
convergent evolution (28). How, then, have different dosage Coimmunoprecipitation and GST pull-down assays. HeLa nuclear extract was
compensation mechanisms evolved? Recent data suggest that prepared as previously described (16). Approximately 100 g nuclear protein in
animals have co-opted evolutionary ancient chromatin-modi- HEMG buffer (25 mM HEPES, pH 7.6, 12.5 mM MgCl2, 0.5 mM EDTA, 1 mM
dithiothreitol, 0.2 mM phenylmethylsulfonyl fluoride, 10% glycerol) with 100
fying complexes for a new function in dosage compensation. mM KCl was used in immunoprecipitation. Immunoprecipitates were washed
Caenorhabditis elegans dosage compensation is regulated by three times with HEMG buffer with 100 mM KCl at room temperature, and
condensin-like proteins, which are normally involved in chro- bound proteins were eluted with sodium dodecyl sulfate-polyacrylamide gel
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
mosome compaction during mitosis (18). Likewise, polycomb electrophoresis (SDS-PAGE) loading buffer.
In vitro GST pull-down assays were performed in HEMG buffer with 200 mM
proteins that have been implicated in X chromosome inacti-
KCl. Briefly, 300 ng of FLAG-hMSL3 or RCC1 was incubated with 1 g of
vation in mammals have an evolutionary older function in recombinant hMOF constructs bound to glutathione beads for 1 h at room
repression of homeotic genes during development (35). With the temperature. After incubation, beads were washed three times, for 5 min each
exception of the mammalian dMLE orthologue, the transcrip- time, with HEMG buffer with 200 mM KCl. Bound proteins were eluted with
tional coactivator RNA helicase A, the function of the Drosophila SDS loading buffer.
Histone acetyltransferase assays. Histone acetyltransferase assays were per-
DCC gene orthologues in vertebrates remains unclear. formed as described earlier (2). Protein (100 ng or indicated amounts) was
Recently, a putative human orthologue of Drosophila MOF, incubated for 30 min at 30°C in HAT buffer (20 mM Tris-HCl, pH 8.8, 1.5 mM
hMOF/MYST1, was isolated (34). Like the Drosophila protein, MgCl2, 10 mM NaCl, supplemented with 125 nCi [3H]acetyl-coenzyme A) with
it contains a chromodomain and a MYST family HAT domain. 1 g of recombinant histone octamer or 1 g of nucleosomal histones assembled
by salt exchange. Reactions were either blotted on a hydrophobic p81 paper and
A C-terminal fragment of this protein was shown to possess
scintillation counted or run on 15% SDS-PAGE and Coomassie stained. The
histone acetyltransferase activity toward histones H3, H2A, signal on SDS-PAGE gels was intensified with Amplify solution (Amersham).
and H4 in vitro (34). A human gene hMSL3/MSL3L1 has also Mass spectrometry. For the in vivo analysis of modified histones, histone
been isolated and characterized previously as a candidate gene bands were modified in gel using propionic anhydride or D6 acetic anhydride and
for several developmental disorders (15, 39). It encodes a pro- digested with trypsin (6, 7, 38, 49). Matrix-assisted laser desorption ionization
(MALDI) spectra were recorded on a Voyager STR instrument (PE-Sciex). For
tein with significant homology to the Drosophila MSL3 in three mass spectrometry (MS)/MS analysis, collision-induced decay spectra were re-
distinct regions, including the two chromo-like domains (27). corded on a Q-STAR XL instrument (PE-Sciex) with manually adjusted collision
Intrigued by this evolutionary conservation, we have studied energies. Fragment spectra were interpreted manually.
the role of hMOF and hMSL3 proteins in mammalian cells. Correlation of confocal laser scanning and electron microscopy. Ultrastruc-
tural investigation by electron microscopy was correlated to observations by laser
We report that human MOF possesses acetyltransferase activ-
scanning microscopy as described in reference 40. Briefly, cells grown on gridded
ity on histones and nucleosomes. Interestingly, depletion of coverslips (Cellocate; Eppendorf AG, Germany) were fixed for 1 h on ice in a
hMOF in HeLa cells leads to a dramatic decrease in histone mixture of 4% freshly prepared formaldehyde, 1% glutaraldehyde (electron
H4 lysine 16 acetylation, while other acetylation sites appear to microscopy grade; Sigma), 1 mM MgCl2, and 100 mM sodium phosphate buffer,
be unaffected. In addition, the cells show altered nuclear mor- pH 6.8, rinsed in buffer, and embedded in Vectashield fluorescence mounting
medium (Vector Laboratories) for observation by confocal laser scanning mi-
phology with polylobular nuclei. This striking phenotype can croscopy (LSM). Following LSM investigation, the coverslips were rinsed in
be rescued by treating the affected cells with the histone buffer, postfixed in 1% buffered OsO4 for 30 min at room temperature, dehy-
deacetylase inhibitor trichostatin A (TSA). HeLa cells trans- drated in aqueous ethanol, and embedded in epoxy resin (Epon 812; Sigma). The
fected with hMOF small interfering RNA (siRNA) show pro- coverslips were subsequently removed under liquid nitrogen depicting the neg-
ative imprint of the Cellocate grid. Ultrathin sections at a nominal thickness of
liferation defects and accumulate in the G2/M phase of the cell
70 nm were prepared from the grid region of interest, poststained in 2% uranyl
cycle. We show that the observed G2/M arrest is at least par- acetate and aqueous lead citrate, and observed in a Philips 410 transmission
tially caused by activation of the DNA damage response path- electron microscope.
way, illustrated by an increased number of ATM pS1981 and Cell culture and transfection. HeLa and HepG2 cells were grown in Dulbec-
␥H2AX foci in hMOF-depleted cells. co’s modified Eagle’s medium supplemented with 10% fetal calf serum, penicil-
lin, streptomycin (Invitrogen), and L-glutamine (Invitrogen). Synthetic siRNAs
(hMOF-1, GUGAUCCAGUCUCGAGUGA; hMOF-2, AAAGACCAUAAGA
MATERIALS AND METHODS UUUAUU; hMOF-3, CAAGAUCACUCGCAACCAA; hMSL3-1, CGGUUA
Protein expression and antibodies. Full-length hMOF was produced in Esch- GUGAAACUUCCAU; hMSL3-2, AAAGGUGACUUCGUCUAAA; control,
erichia coli strain BL21(DE3) as a glutathione S-transferase (GST) fusion in CACGTACGCGGAATACTTCG, sense strand) were purchased from MWG
pET41b(⫹) vector (Merck Biosciences). Cells were induced with 0.3 mM iso- Biotech. Approximately 1.5 ⫻ 105 cells were transfected with Oligofectamine
propyl--D-thiogalactopyranoside (IPTG) for 3 h at 18°C. FLAG-hMSL3 was (Invitrogen) with a 60 nM final concentration of siRNA complexes. TSA was
expressed in ER2566 strain as an intein-CBD fusion, with induction with 0.3 mM added to a final concentration of 25 ng/ml for 72 h, and 2 mM caffeine was added
IPTG for 3 h at 30°C. Following the cleavage of intein-CBD tag, the eluted for 24 h where indicated.
protein was immunoprecipitated with anti-FLAG beads (Sigma) and the bound
full-length hMSL3 protein was eluted with FLAG peptide. Cloning details are
available upon request. RESULTS
Antibodies against hMOF-GST and FLAG-hMSL3 fusion proteins were pro-
duced in rats and rabbits, respectively. Both antibodies were further affinity
Recombinant hMOF is an active histone acetyltransferase.
purified. To generate hMOF peptide antibodies, rabbits were injected with To analyze the enzymatic activity of hMOF protein, we pro-
keyhole limpet hemocyanin-conjugated peptides PERKITRNQKRKHDE and duced recombinant full-length hMOF protein in bacterial cells.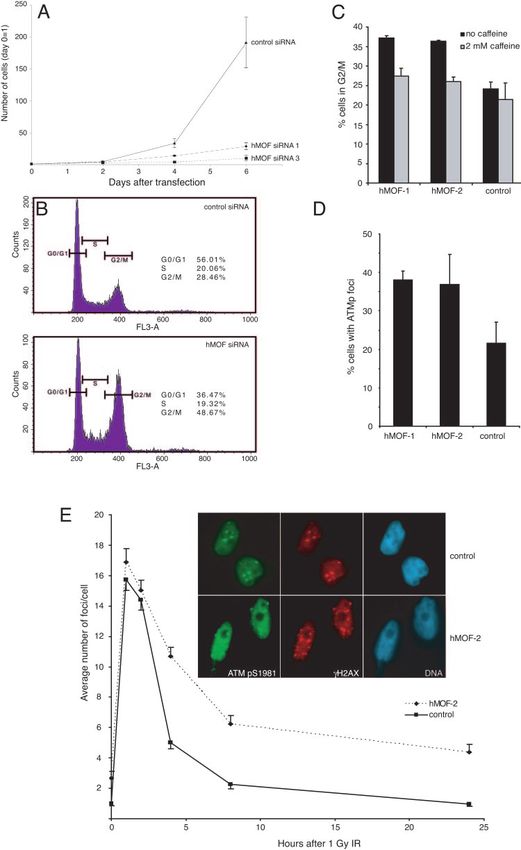
6800 TAIPALE ET AL. MOL. CELL. BIOL.
To study the functional status of the recombinant enzyme, modified in gel using propionic anhydride or D6 acetic anhy-
histone acetyltransferase assays were performed with recom- dride (Fig. 2B and 2C, respectively), and digested with trypsin
binant histones, mononucleosomes, oligonucleosomes, and de- (6). Comparing the MALDI-time of flight spectra of the di-
rivatives of hMOF lacking N or C termini (Fig. 1A and B). As gested peptides from control and hMOF-depleted cells showed
expected, the MYST homology domain is sufficient for hMOF a 20% increase in the unacetylated peptide, a 20% drop of the
acetyltransferase activity, and the C2HC zinc finger region is mono- and diacetylated forms, and a complete absence of tri-
required for activity (Fig. 1C, lanes 3 and 4). As shown in Fig. and tetra-acetylated H4 in cells treated with an siRNA against
1C and D, hMOF is a robust histone acetyltransferase with hMOF.
substrate preference for histone H4, but it also acetylates his- As the Western blot results with acetylation-specific antibod-
tone H3 and histone H2A on free histones as well as on ies suggested a strong effect of hMOF ablation on the acety-
nucleosomes (Fig. 1D, right panel, compare lanes 1 to 3 with 4 lation of H4K16 (Fig. 2A), we performed an MS/MS analysis
to 9). In contrast to recombinant dMOF, which exclusively on the monoacetylated/propionylated H4 peptide that carries
acetylates histone H4 in a nucleosomal context (2), hMOF amino acids 4 to 17 of the H4 tail. By comparing the relative
significantly acetylates both histone H3 and histone H4 (Fig. abundance of the expected fragment ions for the nonacetylated
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
1D, compare lanes 10 and 11, and data not shown). (propionylated) and the acetylated peptides (Fig. 2D), we con-
We were next interested in testing whether this difference in cluded that in HeLa cells, the major histone H4 acetylation
substrate specificity observed between hMOF and dMOF sites are K16 and K12 (85%). There is, however, a marked
would also be observed with hMOF immunoprecipitated from difference in the ratio of K12/K16 acetylation between cells
cells. For this purpose, we immunoprecipitated hMOF from a depleted for hMOF and control cells. Fragment ions contain-
stable cell line expressing hemagglutinin (HA)-2⫻ FLAG- ing only K16 (m/z of 530.35 for the acetylated form and m/z of
tagged hMOF with an anti-FLAG antibody. Nuclear extracts 544.35 for the unacetylated, propionylated form) show an ap-
prepared from untagged HeLa cells were used as controls. proximately sevenfold-lower appearance of the acetylated
Interestingly, we observed that in contrast to recombinant form in hMOF knockdown cells compared to control cells (Fig.
hMOF, immunoprecipitated hMOF acetylates only histone H4 2E, second panel). This strong decrease in acetylation is partly
in mononucleosomes (Fig. 1E, left panel), suggesting that rescued by an increase in the acetylation of lysines 5, 8, and 12
hMOF is more specific in vivo. Indeed, Western blot analysis of (compare the peak height of for acetylated versus nonacety-
acetylated histones revealed that immunoprecipitated hMOF lated in the top, middle, and bottom panels). This shift also
acetylates H4K16, while H4K12 remained unacetylated (Fig. explains the moderate effect of hMOF depletion on overall
1E, right panel). It therefore appears that while Drosophila monoacetylation levels despite its strong impact on K16 acet-
MOF is capable of specific H4K16 acetylation in vitro, hMOF ylation.
specificity for histone H4 is regulated either by additional com- hMOF and hMSL3 interact directly in vitro and in vivo. It
plex components or posttranslational modifications (see be- has previously been shown in Drosophila that dMOF and
low). dMSL3 can interact in vitro and that this interaction leads to
Loss of hMOF abolishes H4 lysine 16 acetylation in vivo. To acetylation of dMSL3 in vivo and in vitro (11). These obser-
study the function of the hMOF protein in vivo, we used RNA vations led us to test whether the interaction is conserved
interference in HeLa and HepG2 cells. Using three indepen- between hMOF and hMSL3 in mammalian cells. We per-
dent synthetic siRNAs against the hMOF coding region, we formed coimmunoprecipitation experiments with hMOF anti-
were able to specifically reduce the levels of hMOF protein body or corresponding preimmune serum, as a negative con-
down to 10% of its original level in HeLa cells (Fig. 2A, upper trol, from nuclear extracts prepared from HeLa cells. The
panel, and data not shown). To determine which lysine resi- coimmunoprecipitated samples were analyzed by SDS-PAGE
dues would be targets of acetylation by hMOF in vivo, we followed by Western blot analysis. The results of these exper-
isolated endogenous histones by acid extraction from cells iments show that hMSL3 coimmunoprecipitates with hMOF
treated with either control siRNA or hMOF-specific siRNA. (Fig. 3A, compare lanes 1 and 2). Interaction was also detected
The histones were separated by SDS-PAGE, and subsequently, when coimmunoprecipitation experiments were performed
Western blot analysis was performed with antibodies against with anti-hMSL3 or using nuclear extracts from HEK293 cells
acetylated histones. Ponceau S staining of the membranes re- (data not shown). It is important to note that we could observe
vealed equal loading of histones. Interestingly, the level of two isoforms of hMSL3 (hMSL3a and hMSL3c) coimmuno-
histone H4 lysine 16 acetylation (H4K16) in hMOF-depleted precipitating with hMOF. Based on bioinformatics analysis
cells was severely reduced in comparison to the control cells and GenBank, these isoforms most likely represent two splice
(Fig. 2A, right and middle panels). In contrast, there was no variants of the hMSL3 gene.
significant difference in acetylation of H3K14, H3K23, or Since the interaction between hMOF and hMSL3 was con-
H4K12, although these lysines were targets of recombinant served, we wanted to test whether the proteins interact with
hMOF in vitro (Fig. 2A and data not shown). Although we did another orthologue of the Drosophila DCC proteins. However,
not analyze H4K5 or H4K8 acetylation status by Western blot- the orthologue of maleless (MLE) protein, RNA helicase A,
ting, mass spectrometric analysis of endogenous histones did did not coimmunoprecipitate with hMOF or hMSL3, even in
not reveal significant changes (see below). low stringency conditions, suggesting that it is not part of the
To further analyze the acetylation status of the endogenous mammalian MSL complex (Fig. 3A and data not shown). Fur-
histones, mass spectrometric analysis was performed on the thermore, in contrast to a previous report (36), we could not
isolated histones. To this end, histones extracted from control detect an interaction between MRG15 and hMOF in vivo
and hMOF-depleted cells were separated by SDS-PAGE, under our experimental conditions (Fig. 3A).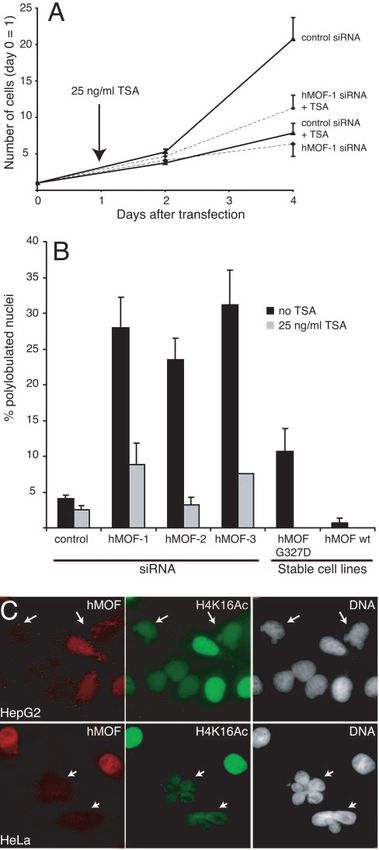
VOL. 25, 2005 hMOF AND H4K16 ACETYLATION IN MAMMALS 6801
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
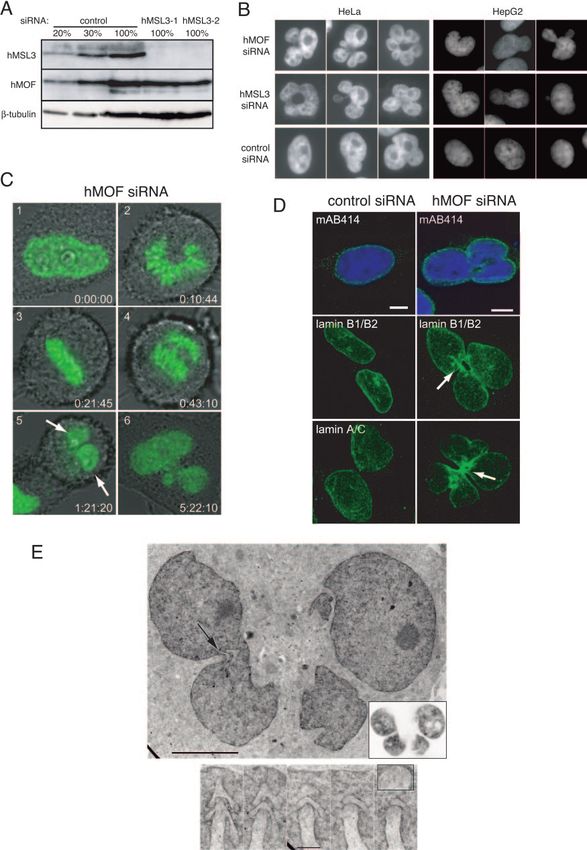
FIG. 1. hMOF is a histone acetyltransferase. (A) Schematic representation of the GST-hMOF fusion constructs used in histone acetyltrans-
ferase assays. (B) Constructs of GST-hMOF and FLAG-hMSL3 expressed in E. coli stained with Coomassie. (C) Filter binding HAT assay with
different hMOF constructs using a recombinant histone octamer as a substrate for acetylation. Bars 1 to 4 correspond to lanes 1 to 4 in panel B;
bar 5, GST alone; bar 6, histones alone. ⫹, with; ⫺, without. (D) Separation of histones from HAT assay by SDS-PAGE. Left panel: top and
middle, autoradiographs (short exposure, overnight; long exposure, 2 weeks) of the gel. Bottom, Coomassie staining of the same gel. Right panel:
comparison of hMOF (lane 10) and dMOF (lane 11) HAT activity on nucleosomes. mononucl., mononucleosomes. (E) Nucleosomal HAT activity
of anti-FLAG immunoprecipitates from HA–2⫻ FLAG-tagged hMOF or from HeLa cells. Left panel: autoradiograph (top) and corresponding
Coomassie gel (bottom). Right panel: detection of modified residues with acetylation-specific antibodies.Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
FIG. 2. Analysis of the histone modification state in cells treated with siRNA against hMOF. (A, right and middle panels) Acetylation of histone
H4 lysine 16 is specifically reduced in hMOF-depleted cells. Cells were transfected with either control or hMOF-1 siRNA. Western blot analysis
of the acid-soluble fraction was performed with acetylation-specific antibodies as indicated. The amount of histones loaded was controlled by
staining the membrane with Ponceau S. (Left panel) Western blot analysis to show reduction of proteins levels by MOF siRNA 1 and 2 in
comparison to control siRNA. (B) MALDI-time of flight analysis of H4 molecules purified via SDS-PAGE and propionylated and digested as
described previously (6). Only the peptide containing amino acids 4 to 17 of H4 is shown. Note that the peptide with the highest m/z carries four
propionyl groups and corresponds to the unmodified form in vivo. (C) Quantitation of acetylation levels seen in vivo. For quantitation, H4
molecules have been modified using D6 acetic anhydride rather than propionic anhydride to ensure similar ionization behavior of the in vitro- and
in vivo-modified peptides. (D) Schematic display of the fragment ions generated by collision-induced decay from a monoacetylated peptide. To
analyze a specific effect of hMOF knockdown on H4K16 acetylation, we have analyzed the monoacetylated peptide by MS/MS. (E) Enlarged
regions of the MS/MS spectra generated from the monoacetylated peptide from control (left panels) or hMOF knockdown (right panels) cells.
Note that the fragment ions containing lysines 5, 8, or 12 show a similar ratio of the acetylated (Ac) form to the nonacetylated (propionylated [Pr])
form, whereas the peptide containing K16 (middle panel) is strongly hypoacetylated in cells treated with siRNA against hMOF.
6802VOL. 25, 2005 hMOF AND H4K16 ACETYLATION IN MAMMALS 6803
directly in vitro (Fig. 3B, top panel, lane 1). An interaction
could still be detected with a deletion derivate of hMOF con-
taining the MYST domain (lane 3 and 4), but no interaction
was detected with the derivative containing only the chromo-
domain of hMOF (lane 2). The C2HC-type zinc finger that is
embedded in the catalytic domain of hMOF was not required
for interaction (Fig. 3B, lane 4). hMSL3 interaction with the C
terminus of hMOF was specific, since another chromatin-as-
sociated protein, RCC1, did not interact with hMOF in the
same experimental conditions (Fig. 3B, bottom panel). In sum-
mary, we can demonstrate a direct interaction between hMOF
and hMSL3. We also found that the C terminus of hMOF
mediates this interaction.
hMOF- and hMSL3-depleted cells show nuclear morphol-
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
ogy defects. Apart from severe reduction of lysine 16 acetyla-
tion, another striking feature of hMOF knockdown cells was
their abnormal nuclear structure. About 36 h after hMOF
siRNA transfection, cells started to undergo dramatic changes
in nuclear morphology and to form multiple lobes (Fig. 4B).
The phenotype ranged from nuclei showing one extra lobe to
cells with 6 to 8 lobes in the nucleus. This phenotype was
observed with three independent hMOF siRNAs tested, and
on average, 25 to 30% of the cells treated with hMOF siRNA
showed severe morphological defects (see below). Interest-
ingly, hMSL3-depleted cells also showed similar defects, albeit
with a lower frequency, suggesting that the two proteins func-
tion in the same pathway (Fig. 4B and see below). Knockdown
of hMSL3 in HeLa cells was effective with two independent
siRNAs tested, as shown in Fig. 4A. Similar results were ob-
tained when hMOF and hMSL3 were depleted in HepG2 cells,
excluding a cell type-specific effect (Fig. 4B and data not
shown). The polylobular phenotype was also observed in HeLa
cells stably expressing enzymatically inactive epitope-tagged
hMOF (HA-2⫻ FLAG-hMOF G327D), whereas a cell line
stably transfected with a wild-type hMOF construct (HA-2⫻
FLAG-hMOF) had a normal morphological appearance (Fig.
5B).
Next, we were interested in determining the stage of the cell
cycle at which hMOF-depleted cells acquire polylobular nuclei.
We therefore produced a stable HeLa cell line expressing
FIG. 3. hMOF and hMSL3 interact in vivo and in vitro. (A) Im-
C-terminally GFP-tagged histone H2B (H2B-GFP) and used
munoprecipitation (IP) of hMOF (lane 1) with preimmune serum these cells in live imaging. hMOF-depleted and control cells
(C) (lane 2) as a control. The amount of input is indicated (lanes 3 and were imaged for 48 h to ensure that we see at least one mitotic
4). (B) In vitro pull-down assay of hMSL3 (top panel) and RCC1 event during the imaging process. We observed that, in com-
(bottom panel) with hMOF-GST constructs (lanes 1 to 4). Input cor- parison to the control cells that had normal nuclei before and
responds to 10% of the starting material (lanes 5 to 8).
after mitosis, hMOF siRNA-treated cells that had normal
round-shaped nuclei before mitosis showed a polylobular ap-
pearance in late telophase after nuclear envelope reassembly
The experiment above demonstrates that interaction be- (Fig. 4C).
tween hMOF and hMSL3 is conserved in mammalian cells. It It has previously been reported that depletion of nuclear
does not address, however, whether this interaction is direct or pore complex component RanBP2 leads to changes in nuclear
mediated by another unknown component. To address this morphology (42). Since the nuclei appear polylobulated after
issue, we performed in vitro GST pull-down assays with GST- completion of mitosis, we wanted to address whether this de-
hMOF fusion derivatives and full-length hMSL3. Another nu- fect appears due to abnormal nuclear envelope reassembly.
clear protein, RCC1, was used as a control. Following binding, For this purpose, we immunostained hMOF siRNA-treated
the beads were washed and the bound fraction was analyzed by and control cells with mAB414 monoclonal antibody that rec-
Western blotting. The blots were first probed with hMSL3 or ognizes several nucleoporins. However, we observed no major
RCC1 antisera and subsequently with a GST antibody to con- defects in the nuclear pore distribution (Fig. 4D). Similarly, the
trol for equal loading of hMOF-GST constructs. Consistent cytoskeleton of hMOF knockdown cells appeared normal, as
with our observations in vivo, hMOF and hMSL3 interacted judged by phalloidin (F-actin) and -tubulin staining (data not6804 TAIPALE ET AL. MOL. CELL. BIOL.
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
FIG. 4. Nuclear morphology defects in hMOF-depleted cells. (A) Knockdown of hMSL3 by RNA interference in HeLa cells 4 days after
transfection of two independent hMSL3 siRNAs. The Western blot was probed with antibodies against hMSL3 (top panel), hMOF (middle panel),
and -tubulin (bottom panel). (B) Spectrum of nuclear defects in HeLa and HepG2 cells. Panels show hMOF-depleted cells, hMSL3-depleted
cells, and control cells 4 days after transfection of siRNA. Nuclei were stained with Hoechst 33342. (C) Still images from a movie of a dividing
hMOF-depleted cell with chromatin labeled with H2B-GFP from early prophase (top left) to G1 (bottom right). Arrows indicate lobes appearingVOL. 25, 2005 hMOF AND H4K16 ACETYLATION IN MAMMALS 6805
shown). We have also tested differences in distribution of nu- tion of the nuclear morphology in these cells. hMOF was
cleoli (fibrillarin), heterochromatin (HP1␣, H3K9 methylation, efficiently knocked down in untreated and TSA-treated cells
H3K27 methylation), and centromeres (CREST autoimmune (data not shown). Interestingly, whereas control cells prolifer-
serum) by immunofluorescence but have not been able to find ated significantly more slowly in the presence of TSA, hMOF
any differences between hMOF knockdown and control cells knockdown cells grew slightly better in the presence than in the
(data not shown). absence of TSA (Fig. 5A). TSA did not, however, fully restore
Another component influencing nuclear architecture is the normal growth of hMOF knockdown cells. More importantly,
nuclear lamina. Loss or overexpression of lamins, which con- nuclear morphology was restored in TSA-treated hMOF
stitute the nuclear lamina, has been reported to cause defects knockdown cells. Similar results were obtained with all hMOF
in nuclear structure (9, 20). We therefore tested whether the siRNAs tested (Fig. 5B). We further confirmed that in both
distribution or levels of nuclear lamina proteins, namely lamin HeLa and HepG2 cells, polylobulation correlates with loss of
A/C and lamin B1, were affected in hMOF siRNA-treated hMOF, which can also be tracked with an H4K16Ac-specific
cells. Remarkably, we could observe a modest but consistent antibody (Fig. 5C).
enrichment of lamin B1 and lamin A/C, typically in the con- However, TSA treatment did not lead to an increase in
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
striction between the lobes (Fig. 4D; see below). The differ- H4K16 acetylation in hMOF-depleted cells (data not shown),
ence between hMOF-depleted and control cells was mainly in suggesting that this modification is not directly linked to nu-
the distribution of lamins, as there was no significant change in clear shape changes and that hMOF has other cellular targets
lamin B1 or lamin A/C protein levels upon hMOF depletion (see Discussion).
(data not shown). Cell cycle checkpoint activation in hMOF-depleted HeLa
Ultrastructural investigation by electron microscopy re- cells. We also observed that hMOF siRNA-treated HeLa cells
vealed that the nucleoli were normal in shape and size and showed proliferation defects in comparison to the control
showed their characteristic stacked composition in fibrillar siRNA-treated cells (Fig. 6A). hMOF-depleted cells were neg-
center, fibrillar component, and granular component, indicat- ative for Trypan blue staining, indicating that the primary
ing normal physiological activity in hMOF-depleted versus cause of growth arrest was not due to apoptosis or necrosis
control cells. Nuclear pores appeared as in normal control (data not shown). Next, we analyzed the control siRNA and
cells. In contrast, the lamina underneath the nuclear envelope hMOF siRNA-treated populations with flow cytometry to ob-
was more discrete and regular in structure than in control cells. serve changes in cell cycle profiles. We found that hMOF
This is consistent with observations made above with enriched siRNA-treated cells accumulate in the G2/M phase of the cell
lamin staining. cycle in comparison to the control cells (Fig. 6B). Furthermore,
Major structural reorganization was, however, seen at the fluorescence-activated cell sorter (FACS) analysis also re-
cytoplasmic side of the folds which separate the nuclear lob- vealed that the cells treated with hMOF siRNA were approx-
ules. The folding process appears to be more complicated than imately 10% larger than the control cells in all phases of the
a simple indentation of the nuclear envelope from the periph- cell cycle (data not shown).
ery to the center. At the front of the fold, the substrate-facing Eukaryotic cells have developed a number of checkpoints
side (bottom) of the nuclear envelope forms stacks of wrinkles that delay cell cycle progression in response to environmental
in an oblique direction to the main fold (Fig. 4E). This front stress. There is an evolutionarily conserved checkpoint at the
region of folds is densely packed with vesicles and filaments, G2/M boundary of the cell cycle, and a wide variety of agents
suggesting that folds are formed also by an active engagement can trigger this G2/M checkpoint (for a review, see reference
of the cytoplasm. In summary, we did not observe gross 43). Regulation of the checkpoint is a complex process with
changes in nuclear organization in hMOF-depleted cells. redundancy in response to different genotoxic and nongeno-
However, it remains possible that subtle changes in chroma- toxic stresses. Recent evidence suggests that there are two
tin acetylation levels in cells or changes in acetylation of an major signaling cascades regulating this G2 delay. One pathway
unknown substrate may contribute to the observed nuclear is caffeine insensitive and involves the p38 kinases activated in
deformations. We reasoned that if this was the case, changing the cytoplasm (10), while the other, which is caffeine sensitive,
the balance of acetylation in the cell might rescue the poly- is driven by the nuclear ATM/ATR/DNA-PK pathway (43).
lobular phenotype. To address this issue, we first transfected We reasoned that activation of either of these checkpoints
HeLa cells with a control siRNA or hMOF siRNA, and 24 h could explain the accumulation of hMOF-depleted cells in
after transfection, we added trichostatin A (25 ng/ml), a potent G2/M. First, we tested activation of p38␣ in hMOF-depleted
histone deacetylase inhibitor, to the growth medium for an- and control cells but detected no difference in p38␣ phosphor-
other 72 h. This was followed by Western blot analysis to assess ylation (data not shown).
the knockdown efficiency, assay for cell growth, and observa- Next, we tested whether the arrest in hMOF knockdown
in the late telophase. (D) Confocal images of hMOF-depleted and control cells. Top left, confocal slice of mAB414-stained nuclei. Bar, 5 m.
Lamin B1 and lamin A/C (middle left and bottom left, respectively) are stacks of multiple confocal slices. (E) Ultrastructure of a lobulated nucleus
of an hMOF-depleted cell. Upper panel: ultrathin section from the substrate side of the nucleus. The inset shows the corresponding LSM optical
section of the GFP signal (inverted grayscale) from the same nucleus. The arrow points to the fold shown in detail in the lower panel. Note that
the arrowed structure of this fold is also visible in the LSM image. Bar, 5 m. Lower panel, consecutive ultrathin sections of the nuclear fold,
starting with the bottommost section at the left. The inset shows an enlarged view of the front of the fold to show vesicles and filaments in this
region. Bar, 1 m.6806 TAIPALE ET AL. MOL. CELL. BIOL.
cells involved the ATM/ATR pathway by using two inhibitors
of these cascades, caffeine and wortmannin. It is important to
note that both caffeine and wortmannin can inhibit activation
of both ATM and ATR, but the 50% inhibitory concentration
of both drugs for ATR is an order of magnitude higher than for
ATM (46, 47). Therefore, to distinguish between these two
possibilities, we used 2 mM caffeine or 2 M wortmannin,
doses that should primarily inhibit ATM. At 72 h after trans-
fection, hMOF-depleted and control cells were treated with
caffeine or wortmannin for 24 h or left untreated. Subse-
quently, cells were analyzed by FACS to determine cell cycle
profiles. Both caffeine and wortmannin restored the cell cycle
profile of hMOF-depleted cells to almost that of control cells
(Fig. 6C and data not shown).
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
To examine whether hMOF-depleted cells had activated the
ATM-dependent checkpoint, we immunostained hMOF
knockdown and control cells with an antibody against phos-
phorylated ATM (pS1981). In control cells, about 20% of cells
had one or several ATMp foci. However, the number of cells
with ATMp foci after depletion of hMOF was consistently
higher, about 40%, with both siRNAs tested (Fig. 6D). Both
the percentage of the cells with phospho-ATM foci and the
average number of foci was increased upon hMOF knock-
down. These foci colocalized with ␥H2AX foci (data not
shown), suggesting that they represent double-stranded breaks
(DSBs) occurring during the normal cell cycle.
Hypoacetylation of histones could render DNA susceptible
to breakage, thereby activating the checkpoint, or alternatively,
it could impair repair of normally occurring DSBs. In both
cases, the result would be an increased number of ATMp and
␥H2AX foci. To distinguish between these possibilities, we
treated the hMOF-depleted and control cells with ionizing
radiation (1 Gy) and performed a time course study to assess
DNA repair kinetics by counting the presence of ATMp and
␥H2AX foci. hMOF-depleted cells consistently showed a sig-
nificant delay in kinetics of DNA repair as observed by the
presence of more ATMp foci in comparison to control cells
(Fig. 6E). Similar results were obtained with a higher dose of
6 Gy and with HepG2 cells. These results suggest that hMOF-
depleted and thus H4K16-hypoacetylated HeLa cells activate
the G2/M checkpoint due to a delay in the kinetics of the DNA
repair process of hypoacetylated chromatin, as opposed to
hypoacetylation leading to more DNA damage without having
an effect on the repair process itself.
DISCUSSION
FIG. 5. TSA treatment enhances the growth of hMOF-depleted cells. Here we show that the interaction between hMOF and
Cells were transfected with hMOF-1 siRNA or control siRNA and treated hMSL3 is conserved in mammalian cells and is mediated by the
with 25 ng/ml TSA or left untreated as indicated. Error bars indicate
standard errors of the means. (B) Effect of TSA on nuclear morphology in
C-terminal region of the hMOF protein. We also observe that
hMOF-depleted and control HeLa cells. Cells were transfected with hMOF hMOF-depleted and hMSL3-depleted HeLa and HepG2 cells
siRNAs (hMOF-1, -2, or -3) or control siRNA (control). Nuclear morphol- have polylobular nuclei. hMOF knockdown cells have severely
ogy defects were scored in TSA-treated (gray bars) and untreated (black
bars) cells 4 days after transfection. The experiment was performed at least reduced levels of histone H4 lysine 16 acetylation, which may
3 times, counting a minimum of 400 cells per sample each time. Error bars be the primary cause of these defects. This is consistent with
indicate standard errors of the means. Stable cell lines are HeLa cells our observations that nuclear morphology defects can be res-
expressing either wild-type hMOF (wt) or inactive LMOF (G327D).
(C) Loss of hMOF correlates with H4K16 acetylation. HeLa (top) and cued by treating the cells with the histone deacetylase inhibitor
HepG2 (bottom) cells were stained for hMOF (left, red) and acetylated trichostatin A. Furthermore, we find that depletion of hMOF
H4K16 (middle, green). Right panels, DNA stained with Hoechst 33342.
Arrows indicate cells in which hMOF has efficiently been knocked down. causes increased phosphorylation of ATM, which can explain
Note the altered structure of the hMOF-depleted nuclei. the accumulation of cells in G2/M. Consistent with this, inhib-VOL. 25, 2005 hMOF AND H4K16 ACETYLATION IN MAMMALS 6807
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
FIG. 6. (A) Growth of HeLa cells after transfection with hMOF or control siRNA. Error bars indicate standard errors of the means. (B) FACS
profile of hMOF-depleted and control cells 4 days after transfection. The percentage of cells in each phase of the cell cycle is also indicated.
(C) The PIKK inhibitor caffeine partly suppresses the accumulation of hMOF-depleted cells in G2/M. Caffeine-treated (2 mM, 24 h) or untreated
HeLa cells were analyzed by FACS 4 days after transfection of hMOF siRNA or control siRNA. Bars indicate percentages of G2/M cells. Error
bars indicate standard errors of the means. (E) hMOF-depleted cells show an increase in ATM S1981 phosphorylation. (E) Kinetics of DSB repair
in hMOF-depleted and control cells after irradiation (IR) (1 Gy). At each time point, ATM pS1981 foci were counted from 50 randomly selected
cells. Error bars indicate standard errors of the means within the experiment. Inset, example of ATM pS1981 and ␥H2AX foci 4 h after 1-Gy
irradiation in control (top) and hMOF-depleted (bottom) cells.6808 TAIPALE ET AL. MOL. CELL. BIOL.
iting the ATM pathway with caffeine or wortmannin sup- and lamin B1 were redistributed to the concave surfaces of
presses the G2/M arrest caused by depletion of hMOF. invaginations. Consistent with this observation, we could de-
Role of hMOF in histone H4 lysine 16 acetylation in vivo. tect thickened nuclear lamina structures in hMOF-depleted
We detected a significant decrease in H4K16 acetylation in cells by electron microscopy. Previous studies have shown that
hMOF knockdown cells, whereas acetylation of other lysines overexpression of full-length or dominant-negative forms of
(H4K12, H3K14, and H3K23) remained mostly unaffected, lamins leads to nuclear structure aberrations (9, 20). It was
although these lysines are acetylated by recombinant hMOF in recently shown that expression of Chk tyrosine kinase in the
vitro. Endogenous mammalian MSL complex therefore ap- nucleus leads to a strikingly similar polylobular phenotype,
pears to be specific for H4K16, or alternatively, there could be including redistribution of lamin B1 and a high concentration
more redundancy between HATs acetylating H3K14, H3K23, of microtubules around the invaginations (33). However, Chk-
and H4K12 in mammalian cells. Purification of the human dependent nuclear shape change was mitosis independent,
MSL complex should shed more light on this issue in the whereas we could only see morphological changes following
future. mitosis. It is therefore likely that there are multiple mecha-
The dramatic decrease in H4K16 acetylation in hMOF nisms maintaining nuclear shape.
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
knockdown cells suggests that hMOF is the major H4K16 Acetylation appears to be involved in the process, since
acetyltransferase in mammals. To our knowledge, this is the expression of an enzymatically inactive hMOF induced
first report connecting a mammalian histone acetyltransferase changes in nuclear morphology similar to those induced by
to a specific lysine residue in vivo. In mammals, lysine 16 is hMOF knockdown. Consistently, polylobulation of HeLa cells
clearly the most abundant acetylation site on histone H4 (32, was rescued upon treatment with TSA, a potent histone
53, 55). In bulk chromatin preparations, nearly all acetylated deacetylase inhibitor. It is, however, unlikely that H4K16 acet-
histone H4 molecules are acetylated at H4K16, be it mono-, ylation plays a direct role in nuclear morphology changes.
di-, tri-, or tetra-acetylated H4 (32, 53, 55). It will be interesting First, TSA treatment of hMOF-depleted cells did not lead to
to study whether localization of hMOF coincides with H4K16 an increase in H4K16 acetylation. Second, hMSL3 depletion
acetylation patterns on promoters and/or coding regions pre- had no impact on bulk K16 acetylation despite a similar phe-
viously documented in mammalian cells. notype (data not shown). Third, the histone deacetylases
It was originally suggested that histone acetylation is a hier- known to deacetylate H4K16 belong to the TSA-insensitive
archical process, where one modification is required for sub- SIR family (22, 57). It is thus conceivable that hMOF also has
sequent acetylation (55). H4K16, being the most commonly other TSA-sensitive cellular targets that remain to be identi-
acetylated residue, would be placed on top of the cascade. fied.
However, we could detect no reduction in H4K5, H4K8, or hMOF, H4K16 acetylation, and DNA damage response
H4K12 acetylation in hMOF knockdown cells. On the con- pathway. We also observed that the depletion of hMOF af-
trary, hMOF-depleted cells appeared to compensate for the fected cell proliferation in HeLa cells. This effect was not due
loss of K16 acetylation by higher levels of acetylation on other to apoptosis or necrosis, but we found that the cells were
lysine residues on the H4 tail. This result clearly implies that enriched at the G2/M phase of cell cycle. In an effort to un-
K16 acetylation is not a prerequisite for subsequent acetyla- derstand whether a checkpoint cascade is activated in hMOF-
tion. depleted cells, we observed that the G2/M defect could be
Conserved interaction between hMOF and hMSL3. We suppressed by inhibiting checkpoint response with low doses of
found that hMOF and hMSL3 coimmunoprecipitate in vivo in caffeine. Low doses of caffeine were used to rule out the in-
HeLa cells as well as HEK293 cells. Furthermore, we detect a volvement of DNA-PK and the ATR pathway. Consistent with
direct interaction between hMOF and hMSL3 in vitro. We this observation, we found no defects in UV-induced DNA
have also mapped the sites of interaction to the C-terminal damage response in hMOF siRNA-treated cells (data not
MYST domain. A similar nuclear morphology phenotype in shown). In addition to caffeine, we also observed that wort-
hMOF- and hMSL3-depleted cells corroborates the finding mannin had a similar effect on the cell cycle profile (data not
that the two proteins function in the same complex. shown). These results strongly suggest that hMOF-depleted
The human orthologue of dMLE, RNA helicase A, did not cells have activated the DNA damage pathway. Decreasing
interact with hMOF or hMSL3 under the conditions used. cellular hMOF levels led to increased phosphorylation of
dMLE seems to interact with other members of the Drosophila serine 1981 of ATM and ␥H2AX, hallmarks of activation of
DCC only transiently (14), suggesting that it is not a stable this pathway (5). hMOF knockdown cells do not seem to be
component of the complex. It is possible that dMLE is a later more susceptible to DNA damage. However, the kinetics of
addition to the DCC, perhaps reflecting the role of noncoding DNA repair after irradiation is slowed down in these cells.
RNAs as functional units of the Drosophila complex. This Thus, the most likely explanation for the activation of the
would be consistent with the results of this study. checkpoint is that hMOF-depleted cells fail to efficiently repair
hMOF is involved in the maintenance of nuclear structure DSBs that occur during the cell cycle, thus delaying the pro-
in HeLa cells. We observed that the nuclei of both hMOF and gression to mitosis.
hMSL3 knockdown cells appear polylobulated. By using live Previously, acetylation of histone H4 has been linked to cell
cell imaging, we were able to show that, in hMOF-depleted cycle progression through G2/M in yeast (29, 30). Yeast cells
cells, these defects appear during nuclear envelope reassembly with four mutant lysines (K5Q, K8Q, K12Q, and K16Q) in the
in late telophase. We did not observe any lagging chromo- H4 tail accumulate in G2/M in a RAD9-dependent manner
somes in mitosis in these cells, indicating the defects are not (29, 30). RAD9 is a sensor protein that arrests cell cycle pro-
due to missegregation of chromosomes. However, lamin A/C gression if cells have accumulated DNA damage (58). Inter-VOL. 25, 2005 hMOF AND H4K16 ACETYLATION IN MAMMALS 6809
estingly, yeast Esa1p is required for cell cycle progression, and This work partially funded by DFG Transregio 5. S.R. is a recipient
rad9 can also suppress the esa1 phenotype (13, 50). Esa1p is a of EMBO and HFSP postdoctoral fellowships.
MYST family acetyltransferase closely related to hMOF. It is REFERENCES
the catalytic subunit of the NuA4 histone acetyltransferase 1. Akhtar, A. 2003. Dosage compensation: an intertwined world of RNA and
complex that also contains Eaf3p, the yeast ortholog of chromatin remodelling. Curr. Opin. Genet. Dev. 13:161–169.
2. Akhtar, A., and P. B. Becker. 2000. Activation of transcription through
hMSL3. Another striking phenotype of esa1 mutant cells is histone H4 acetylation by MOF, an acetyltransferase essential for dosage
accumulation of visible changes in chromatin structure (13). compensation in Drosophila. Mol. Cell 5:367–375.
3. Akhtar, A., and P. B. Becker. 2001. The histone H4 acetyltransferase MOF
hMOF-depleted cells accumulate similarly in G2/M, in a caf- uses a C2HC zinc finger for substrate recognition. EMBO Rep. 2:113–118.
feine-sensitive manner, and show changes in nuclear shape, 4. Akhtar, A., D. Zink, and P. B. Becker. 2000. Chromodomains are protein-
suggesting that the involvement of histone H4 acetylation in RNA interaction modules. Nature 407:405–409.
5. Bakkenist, C. J., and M. B. Kastan. 2003. DNA damage activates ATM
cell cycle progression and/or DNA repair is evolutionarily con- through intermolecular autophosphorylation and dimer dissociation. Nature
served from yeast to mammals. 421:499–506.
Recently, several studies have shown the involvement of 6. Bonaldi, T., A. Imhof, and J. T. Regula. 2004. A combination of different
mass spectroscopic techniques for the analysis of dynamic changes of histone
histone methylation and acetylation in DSB repair (21, 23, 25, modifications. Proteomics 4:1382–1396.
Downloaded from http://mcb.asm.org/ on April 12, 2021 by guest
37, 44). Intriguingly, acetylation of histones has been shown to 7. Bonaldi, T., J. T. Regula, and A. Imhof. 2004. The use of mass spectrometry
for the analysis of histone modifications. Methods Enzymol. 377:111–130.
increase the activity of DNA-PK in a nucleosomal context in 8. Bone, J. R., J. Lavender, R. Richman, M. J. Palmer, B. M. Turner, and M. I.
vitro (37). Furthermore, it has been shown that within 1 to 2 kb Kuroda. 1994. Acetylated histone H4 on the male X chromosome is associ-
of a defined DSB, very little ␥H2AX can be detected (48) and ated with dosage compensation in Drosophila. Genes Dev. 8:96–104.
9. Broers, J. L., E. A. Peeters, H. J. Kuijpers, J. Endert, C. V. Bouten, C. W.
that H4K16 is generally hypoacetylated in fission yeast (23). Oomens, F. P. Baaijens, and F. C. Ramaekers. 2004. Decreased mechanical
These studies suggest that there is a connection between stiffness in LMNA-/- cells is caused by defective nucleo-cytoskeletal integrity.
H4K16 acetylation, DNA-PK activity, and H2AX phosphory- Implications for the development of laminopathies. Hum. Mol. Genet. 13:
2567–2580.
lation in DSB repair. If this aspect of DSB repair is evolution- 10. Bulavin, D. V., S. A. Amundson, and A. J. Fornace. 2002. p38 and Chk1
arily conserved to mammals, it would have interesting impli- kinases: different conductors for the G(2)/M checkpoint symphony. Curr.
Opin. Genet. Dev. 12:92–97.
cations for the role of hMOF in DSB repair. 11. Buscaino, A., T. Kocher, J. H. Kind, H. Holz, M. Taipale, K. Wagner, M.
H4K16 acetylation and cancer. It is remarkable that lysine Wilm, and A. Akhtar. 2003. MOF-regulated acetylation of MSL-3 in the
16 acetylation by the MOF enzyme has been evolutionarily Drosophila dosage compensation complex. Mol. Cell 11:1265–1277.
12. Carrozza, M. J., R. T. Utley, J. L. Workman, and J. Cote. 2003. The diverse
used for various purposes. In Drosophila, it correlates with functions of histone acetyltransferase complexes. Trends Genet. 19:321–329.
increased X-linked gene transcription (1), while in yeast, 13. Clarke, A. S., J. E. Lowell, S. J. Jacobson, and L. Pillus. 1999. Esa1p is an
H4K16 acetylation by orthologous SAS2 regulates heterochro- essential histone acetyltransferase required for cell cycle progression. Mol.
Cell. Biol. 19:2515–2526.
matin spreading (52) and, perhaps surprisingly, negatively cor- 14. Copps, K., R. Richman, L. M. Lyman, K. A. Chang, J. Rampersad-Ammons,
relates with gene transcription (24). It appears that mammals and M. I. Kuroda. 1998. Complex formation by the Drosophila MSL pro-
teins: role of the MSL2 RING finger in protein complex assembly. EMBO J.
have adopted histone H4 lysine 16 acetylation to be one of the 17:5409–5417.
most abundant histone modifications. 15. Cormier, T. A., S. K. Prakash, D. B. Magner, H. Y. Zoghbi, and I. B. Van den
Recently, it was shown that loss of acetylation at H4K16 is a Veyver. 2001. Analysis of Mid1, Hccs, Arhgap6, and Msl3l1 in X-linked
polydactyly (Xpl) and Patchy-fur (Paf) mutant mice. Mamm. Genome 12:
common hallmark of human cancer (17). A variety of human 796–798.
tumor cell lines and primary tumors show significant hy- 16. Dignam, J. D., R. M. Lebovitz, and R. G. Roeder. 1983. Accurate transcrip-
poacetylation at H4K16 and hypo(tri)methylation at H4K20. tion initiation by RNA polymerase II in a soluble extract from isolated
mammalian nuclei. Nucleic Acids Res. 11:1475–1489.
Remarkably, reduction in H4K16 acetylation correlates with 17. Fraga, M. F., E. Ballestar, A. Villar-Garea, M. Boix-Chornet, J. Espada, G.
tumor progression. It was also shown that hMOF is not asso- Schotta, T. Bonaldi, C. Haydon, S. Ropero, K. Petrie, N. G. Iyer, A. Perez-
Rosado, E. Calvo, J. A. Lopez, A. Cano, M. J. Calasanz, D. Colomer, M. A.
ciated with hypomethylated repetitive D4Z4 sequences in the Piris, N. Ahn, A. Imhof, C. Caldas, T. Jenuwein, and M. Esteller. 2005. Loss
HL60 tumor cell line, in contrast to wild-type lymphocytes, of acetylation at Lys16 and trimethylation at Lys20 of histone H4 is a
where these elements are normally methylated. common hallmark of human cancer. Nat. Genet. 37:391–400.
18. Hagstrom, K. A., and B. J. Meyer. 2003. Condensin and cohesin: more than
We have shown that hMOF is the major H4K16-specific chromosome compactor and glue. Nat. Rev. Genet. 4:520–534.
histone acetyltransferase in human cell lines. If this is the case 19. Hilfiker, A., D. Hilfiker-Kleiner, A. Pannuti, and J. C. Lucchesi. 1997. mof,
for most tissues, it implies a significant role for hMOF activity a putative acetyl transferase gene related to the Tip60 and MOZ human
genes and to the SAS genes of yeast, is required for dosage compensation in
in tumorigenesis. Clearly, future studies need to address the Drosophila. EMBO J. 16:2054–2060.
role of hMOF and H4K16 acetylation in healthy and cancer 20. Hoffmann, K., C. K. Dreger, A. L. Olins, D. E. Olins, L. D. Shultz, B. Lucke,
H. Karl, R. Kaps, D. Muller, A. Vaya, J. Aznar, R. E. Ware, N. Sotelo Cruz,
tissues. T. H. Lindner, H. Herrmann, A. Reis, and K. Sperling. 2002. Mutations in
the gene encoding the lamin B receptor produce an altered nuclear mor-
ACKNOWLEDGMENTS phology in granulocytes (Pelger-Huet anomaly). Nat. Genet. 31:410–414.
21. Huyen, Y., O. Zgheib, R. A. Ditullio, Jr., V. G. Gorgoulis, P. Zacharatos, T. J.
We thank Jan Ellenberg and the members of his laboratory as well Petty, E. A. Sheston, H. S. Mellert, E. S. Stavridi, and T. D. Halazonetis.
as ALMF for help with confocal and live cell imaging. We thank 2004. Methylated lysine 79 of histone H3 targets 53BP1 to DNA double-
Thomas Köcher and Matthias Wilms for mass spectrometric analysis strand breaks. Nature 432:406–411.
of recombinant hMOF and Klaus-Josef Weber for invaluable help with 22. Imai, S., C. M. Armstrong, M. Kaeberlein, and L. Guarente. 2000. Tran-
irradiation experiments. We are also grateful to Iain Mattaj, Matthias scriptional silencing and longevity protein Sir2 is an NAD-dependent histone
Hentze, Elisa Izaurralde, Jan Ellenberg, and Juerg Müller for critical deacetylase. Nature 403:795–800.
reading of the manuscript. In addition, we thank Iain Mattaj, Chee- 23. Jazayeri, A., A. D. McAinsh, and S. P. Jackson. 2004. Saccharomyces cer-
evisiae Sin3p facilitates DNA double-strand break repair. Proc. Natl. Acad.
Gun Lee, Harald Herrmann, Olivia Pereira-Smith, Bryan Turner, and
Sci. USA 101:1644–1649.
Jan Ellenberg for providing antibodies. Tej Pandita and John Lucchesi 24. Kurdistani, S. K., S. Tavazoie, and M. Grunstein. 2004. Mapping global
are acknowledged for communication of results prior to publication. histone acetylation patterns to gene expression. Cell 117:721–733.
We are also indebted to Harald Herrmann and to the members of the 25. Kusch, T., L. Florens, W. H. Macdonald, S. K. Swanson, R. L. Glaser, J. R.
Akhtar laboratory for useful discussions. Yates III, S. M. Abmayr, M. P. Washburn, and J. L. Workman. 2004.You can also read

















































