Sphingosylphosphorylcholine Is a Novel Messenger for Rho-Kinase-Mediated Ca2ⴙ Sensitization in the Bovine Cerebral Artery
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Sphingosylphosphorylcholine Is a Novel Messenger for
Rho-Kinase–Mediated Ca2ⴙ Sensitization in the Bovine
Cerebral Artery
Unimportant Role for Protein Kinase C
Satoshi Shirao, Shiro Kashiwagi, Masafumi Sato, Saori Miwa, Fumiaki Nakao, Tetsu Kurokawa,
Natsuko Todoroki-Ikeda, Kimiko Mogami, Yoichi Mizukami, Shinichi Kuriyama, Kyousuke Haze,
Michiyasu Suzuki, Sei Kobayashi
Abstract—Although recent investigations have suggested that a Rho-kinase–mediated Ca2⫹ sensitization of vascular
smooth muscle contraction plays a critical role in the pathogenesis of cerebral and coronary vasospasm, the upstream
of this signal transduction has not been elucidated. In addition, the involvement of protein kinase C (PKC) may also be
related to cerebral vasospasm. We recently reported that sphingosylphosphorylcholine (SPC), a sphingolipid, induces
Rho-kinase–mediated Ca2⫹ sensitization in pig coronary arteries. The purpose of this present study was to examine the
possible mediation of SPC in Ca2⫹ sensitization of the bovine middle cerebral artery (MCA) and the relation to signal
transduction pathways mediated by Rho-kinase and PKC. In intact MCA, SPC induced a concentration-dependent
(EC50⫽3.0 mol/L) contraction, without [Ca2⫹]i elevation. In membrane-permeabilized MCA, SPC induced Ca2⫹
sensitization even in the absence of added GTP, which is required for activation of G-proteins coupled to membrane
receptors. The SPC-induced Ca2⫹ sensitization was blocked by a Rho-kinase inhibitor (Y-27632) and a dominant-
negative Rho-kinase, but not by a pseudosubstrate peptide for conventional PKC, which abolished the Ca2⫹-independent
contraction induced by phorbol ester. In contrast, phorbol ester–induced Ca2⫹ sensitization was resistant to a Rho-kinase
inhibitor and a dominant-negative Rho-kinase. In primary cultured vascular smooth muscle cells, SPC induced the
translocation of cytosolic Rho-kinase to the cell membrane. We propose that SPC is a novel messenger for
Rho-kinase–mediated Ca2⫹ sensitization of cerebral arterial smooth muscle and, therefore, may play a pivotal role in the
pathogenesis of abnormal contraction of the cerebral artery such as vasospasm. The SPC/Rho-kinase pathway functions
independently of the PKC pathway. (Circ Res. 2002;91:112-119.)
Key Words: vasospasm 䡲 sphingolipid 䡲 protein kinase C 䡲 Rho-kinase 䡲 membrane permeabilization
C erebral vasospasm, a sustained abnormal contraction of
the cerebral artery, is one of the major causes of lethal
complications in patients with subarachnoid hemorrhage
an abnormally enhanced contraction of vascular smooth
muscle (VSM).
Because pharmacological interventions with Ca2⫹ channel
(SAH).1 The mechanisms responsible have been under in- blockers in experimental models4,5 and clinical trials6,7 have
tense investigation but remain poorly understood. The pri- showed no significant reduction in the incidence of cerebral
mary determinant of smooth muscle contraction is phosphor- vasospasm, the Ca2⫹ sensitization mechanism, which is resis-
ylation of 20-kDa myosin light chain (MLC),2 which is tant to Ca2⫹ channel blockers, is currently considered to play
regulated by not only the Ca2⫹/calmodulin (CaM)-dependent a more important role in the pathogenesis of cerebral vaso-
MLC kinase (MLCK)–mediated pathway but also a Ca2⫹- spasm. In addition, multiple second messengers/signaling
independent mechanism (⫽Ca2⫹ sensitization).3 Excitatory pathways, including the Rho A/Rho-kinase,8 –10 protein ki-
agonists, including spasmogenic agonists, initially increase nase C (PKC),11,12 and arachidonic acid pathways,13 have
intracellular Ca2⫹ levels leading to activation of MLCK. This been linked to Ca2⫹ sensitization mechanisms.
action is followed immediately by a decrease in MLC Sato et al14 reported that activation of Rho-kinase was
phosphatase activity and an increase in MLC phosphoryla- observed during the vasospasm induced by SAH. It was also
tion, which induces Ca2⫹ sensitization,3 the result of which is reported that HA1077 (fasudil), an inhibitor of Rho-
Original received August 20, 2001; resubmission received April 15, 2002; revised resubmission received May 29, 2002; accepted June 4, 2002.
From the Departments of Molecular Physiology (S.S., M. Sato, S.M., F.N., T.K., N.T.-I., K.M., Y.M., S. Kobayashi) and Neurosurgery (S.S., S.
Kashiwagi, T.K., M. Suzuki), Yamaguchi University School of Medicine, Ube, Japan; and the Research Center of Mochida Pharmaceutical Co, LTD (S.
Kuriyama, K.H.), Gotemba, Japan.
Correspondence to Sei Kobayashi, MD, PhD, Dept of Molecular Physiology, Yamaguchi University School of Medicine, 1-1-1 Minami-Kogushi, Ube
755-8505, Japan. E-mail seikoba@yamaguchi-u.ac.jp
© 2002 American Heart Association, Inc.
Circulation Research is available at http://www.circresaha.org DOI: 10.1161/01.RES.0000026057.13161.42
112
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015
Shirao et al SPC-Induced Ca2ⴙ Sensitization of Cerebral Artery 113
kinase,10,15 prevents the development of cerebral vasospasm
in two-hemorrhage canine models16 and in humans.17 Taken
together, these observations strongly suggest that a Rho-
kinase pathway plays an important role in the pathogenesis of
cerebral vasospasm.
The upstream mediator of the Rho-kinase pathway causing
Ca2⫹ sensitization of cerebral arteries has remained to be
determined. Because translocation and membrane binding is
required for activation of Rho-kinase,18 we hypothesized that
a component of cell membrane or its metabolites may be
present upstream of the Rho-kinase pathway. After extensive
screening of molecules, we recently found that sphingo-
sylphosphorylcholine (SPC) induces a sustained and
concentration-dependent contraction in coronary arteries and
that this contraction was abolished by a Rho-kinase inhibitor
(Y-27632). 19 SPC is a sphingolipid generated by
N-deacylation of sphingomyelin, one of the most abundant
lipids in the cell membrane, and was found to be a critical
molecule in apoptosis, cell proliferation, and endothelial NO
production.20 –22 The slowly developing and sustained nature
of the SPC-induced contraction mediated by Rho-kinase19
suggests a possible role for this mechanism in the abnormal,
prolonged vasoconstriction which occurs in cerebral vaso-
spasm. However, the role of this novel signal transducing
molecule in cerebral arterial contractions has not been
identified.
In the present study, we tested the hypothesis that SPC is a
potential signal mediator for Ca2⫹ sensitization of cerebral
Figure 1. A, Representative recording of simultaneous measure-
arterial smooth muscle contraction as an upstream initiator of ment of [Ca2⫹]i and force, showing the effect of 30 mol/L SPC
the Rho-kinase pathway. Because PKCs may play a role in and 2 mol/L Y-27632 on [Ca2⫹]i and force of the fura-2–loaded
Ca2⫹ sensitization,23 interrelationships between a PKC- MCA strips. Force and fluorescence ratios were expressed as a
mediated pathway and a newly discovered pathway mediated percentage, assuming the values in normal (5.9 mmol/L K⫹) and
118 mmol/L K⫹-PSS to be 0% and 100%, respectively. (B), A
by Rho-kinase and SPC were also examined. concentration-response curve of the SPC-induced contraction.
For these purposes, we directly evaluated Ca2⫹ sensitiza- Several concentrations of SPC were applied in the absence (䡲;
tion of middle cerebral artery (MCA) contraction and its control) and presence (▫) of 2 mol/L Y-27632. Levels of force
were normalized by the response to high K⫹-depolarization
signal transduction mechanisms, using simultaneous mea-
(100%). Data are a summary of 12 independent experiments
surement of [Ca2⫹]i and force and the membrane permeabili- and are expressed as the mean⫾SD. *P⬍0.01.
zation in combination with cytosolic application of a
pseudosubstrate peptide for PKCs and a dominant-negative reached a maximum plateau level (105.8⫾33.6%, n⫽4)
form of Rho-kinase. within 30 minutes, with no change in the [Ca2⫹]i (0.0⫾0.0%,
n⫽4; Figure 1A). This Ca2⫹-independent contraction contin-
Materials and Methods ued for at least 2 hours and did not decrease even after
Ca2⫹ sensitization of the bovine MCA, which we obtained from a
local slaughterhouse24 (Ube Shokuniku Center, Ube, Japan), was replacing the buffer with normal PSS (data not shown).
examined using simultaneous measurement of [Ca2⫹]i and force in The contraction induced by SPC (30 mol/L) was reduced
intact MCA loaded with fura-2,19 and membrane permeabilization from 95.3⫾14.4% (n⫽12) to 0.8⫾1.6% by Y-27632 (2
with ␣-toxin25 and -escin.26 The roles of Rho-kinase and PKCs mol/L) (n⫽4; P⬍0.01; Figure 1), thereby indicating com-
were assessed using a dominant-negative form of Rho-kinase,
GST-RB/PH(TT)27 and a PKC19-31 pseudosubstrate inhibitor,28 plete inhibition of the SPC-induced contraction by Y-27632.
respectively, which were added to the cytosol of -escin–permeab- In contrast, Y-27632 (2 mol/L) had no apparent effect on
ilized MCAs. The expression and translocation of Rho-kinase were the 118 mmol/L K⫹-PSS–induced [Ca2⫹]i elevation and con-
investigated using Western blots in bovine MCA and rat aorta and traction, which is widely regarded as a typical Ca2⫹-
confocal immunofluorescence image processing analysis in vascular
smooth muscle cells (VSMCs) in primary culture, respectively.
dependent contraction (n⫽8; P⬎0.05; Figure 1A). This
An expanded Materials and Methods section can be found in the finding, taken together with the contractile response to a
online data supplement available at http://www.circresaha.org. quick release during SPC-induced contraction, indicates that
the irreversible contraction is physiological (see online data
Results supplement for details).
2ⴙ The SPC-induced contraction was concentration-
SPC-Induced Ca -Independent Contraction in
Intact VSM dependent with an EC50 of 3.0 mol/L (Figure 1B). The
In the simultaneous measurement of [Ca2⫹]i and force, SPC maximum response was obtained at 30 mol/L
(30 mol/L) induced a gradual increase in tension, which (95.3⫾14.4%, n⫽12; Figure 1B).
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015114 Circulation Research July 26, 2002
These results indicate involvement of the G protein–indepen-
dent pathway in the Ca2⫹ sensitization induced by SPC,
although direct measurements of nucleotides and their kinet-
ics in the permeabilized MCA are required for confirmation.
Effects of PKC␣ 19-31, Y-27632, and GST-RB/PH
(TT) on SPC-Induced Ca2ⴙ Sensitization in
-Escin–Permeabilized MCA
To investigate the signal transduction mechanism(s) involved
in the SPC-induced Ca2⫹ sensitization, we used MCA strips
Figure 2. Differential requirement of GTP for the contractile permeabilized with -escin. Because permeabilization with
response induced by SPC and U-46619 at constant Ca2⫹ (pCa -escin allows higher molecular mass compounds (up to 150
6.3, buffered with 10 mmol/L EGTA) in the ␣-toxin–permeabil- kDa) to penetrate the cell membrane, large molecules such as
ized bovine MCA. Sequential experiments were done using the
same vascular strip. U-46619 (30 nmol/L) plus GTP (10 mol/L) recombinant proteins can be introduced into the cytosol of the
was applied in the presence of GDPS (1 mmol/L), and subse- -escin–permeabilized smooth muscle.26,29 At constant
quently, GTP (10 mol/L) was applied in the presence of [Ca2⫹]i (pCa 6.3), 30 mol/L SPC elicited a significant
U-46619 (30 nmol/L). Then, SPC (30 mol/L) was applied in the
absence of GTP to the same strip of VSM. GTP alone (without
additional 64.4⫾7.3% increase in the force (⫽Ca2⫹ sensiti-
an agonist for a receptor) had no apparent contractile effect on zation; n⫽8; P⬍0.01; Figures 3A and 5B) even in the
the force. absence of GTP, which is required for activation of G
proteins coupled to membrane receptors. This 30 mol/L
SPC-Induced Ca2ⴙ Sensitization in SPC-induced Ca2⫹-independent contraction was abolished by
Membrane-Permeabilized MCA With No a Rho-kinase inhibitor (2 mol/L Y-27632) (P⬍0.05; Figures
Requirement for G Protein Activation 3C and 5B) and by a dominant-negative form of Rho-kinase
Using ␣-toxin–permeabilized MCA, the SPC-induced Ca2⫹ (0.5 g/mL GST-RB/PH (TT)) (P⬍0.05; Figures 3D and
sensitization and the involvement of G protein were exam- 5B), whereas a PKC pseudosubstrate, PKC␣ 19-31, had no
ined (Figure 2). Because molecules with a molecular mass of effect on SPC-induced Ca2⫹ sensitization (Figures 3B and
less than 1000 can penetrate the membrane in ␣-toxin–per- 5B). In contrast, the force at pCa 6.3 (⫽Ca2⫹-induced
meabilized strips,25 [Ca2⫹]i can be controlled at a constant contraction) was not affected by 2 mol/L Y-27632 or by 0.5
value with a highly buffering effect of 10 mmol/L EGTA. In g/mL GST-RB/PH (TT) (Figure 5A). These results strongly
constant [Ca2⫹]i (pCa 6.3), a G protein– coupled receptor suggest the requirement of Rho-kinase, but not of G proteins
agonist, U-46619 (30 nmol/L), required the presence of GTP and PKCs, for SPC-induced Ca2⫹ sensitization.
to induce contraction and this was abolished by GDPS
(1 mmol/L), thus indicating an essential role of G protein Effects of PKC␣ 19-31, Y-27632, and GST-RB/PH
activation in the U-46619 –induced contraction. In contrast, in (TT) on Phorbol 12,13-Dibutyrate
the same strip, 30 mol/L SPC induced Ca2⫹ sensitization at (PDBu)–Induced Ca2ⴙ Sensitization in
pCa 6.3, even in the absence of added GTP (Figure 2). GTP -Escin–Permeabilized MCA
was also not required for the SPC-induced Ca2⫹ sensitization In -escin–permeabilized smooth muscle and at constant
in -escin–permeabilized MCA (Figure 3). Neither GDPS [Ca2⫹]i (pCa 6.3), 0.03 mol/L PDBu elicited a significant
nor GTP modified the Ca2⫹ sensitization induced by SPC. additional 84.8⫾3.4% increase in the force (⫽Ca2⫹ sensiti-
Figure 3. Effects of PKC␣ 19-31, Y-27632, and
GST-RB/PH (TT) on the SPC-induced Ca2⫹ sensi-
tization of -escin–permeabilized bovine MCA at
constant Ca2⫹ (pCa 6.3, buffered with 10 mmol/L
EGTA). SPC (30 mol/L)-induced Ca2⫹ sensitiza-
tion was observed in the absence of GTP, which
is required for activation of G proteins. Y-27632 (2
mol/L) was applied before (C) and after (A and B)
the application of 30 mol/L SPC. In B and D,
SPC was applied in the presence of 100 mol/L
PKC␣ 19-31 (B) and 0.5 g/mL GST-RB/PH (TT),
a dominant-negative Rho-kinase (D). At the end of
each experiment, the maximal contraction induced
by pCa 4.5 was observed for normalization in the
data analysis (see Figure 5).
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015Shirao et al SPC-Induced Ca2ⴙ Sensitization of Cerebral Artery 115
Figure 4. Effects of PKC␣ 19-31, Y-27632, and GST-RB/PH (TT)
on PDBu-induced Ca2⫹ sensitization of -escin–permeabilized
bovine MCA at constant Ca2⫹ (pCa 6.3, buffered with 10 mmol/L
EGTA). PDBu (0.03 mol/L) was applied in the absence (A) and
presence of several blockers (B through D): 50 mol/L PKC␣
19-31 (B), 2 mol/L Y-27632 (C), and 0.5 g/mL GST-RB/PH
(TT) (D). At the end of each experiment, a maximal contraction
induced by pCa 4.5 was observed for normalization in the data
analysis (see Figure 5).
zation; n⫽3; P⬍0.01; Figures 4A and 5C). A pseudosubstrate
peptide for conventional and novel PKCs (50 mol/L PKC␣
19-31) abolished the Ca2⫹ sensitization induced by 0.03
mol/L PDBu (n⫽4; P⬍0.05; Figures 4B and 5C) without
affecting the force at pCa 6.3 (⫽Ca2⫹-induced contraction)
(P⬎0.05, n⫽4; Figure 5A). In contrast, the 0.03 mol/L
PDBu-induced Ca2⫹ sensitization of force was not affected by
preincubation with 2 mol/L Y-27632 and 0.5 g/mL GST-
RB/PH (TT) (Figures 4C, 4D, and 5C), both of which
abolished the SPC-induced Ca2⫹ sensitization presumably
mediated by Rho-kinase. These results strongly suggest that Figure 5. Summary of experiments shown in Figures 3 and 4. A,
Rho-kinase is not required for the Ca2⫹ sensitization mediated Effects of PKC␣ 19-31 (50 mol/L), Y-27632 (2 mol/L), and
GST-RB/PH (TT) (0.5 g/mL) on the force level at pCa 6.3 (Con-
by phorbol ester–sensitive PKCs, which are selectively inhib- trol) (n⫽4 to 9). B, Effects of PKC␣ 19-31 (100 mol/L), Y-27632
ited by their pseudosubstrate peptide, PKC␣ 19-31. (2 mol/L), and GST-RB/PH (TT) (0.5 g/mL) on the Ca2⫹-
independent contraction induced by 30 mol/L SPC. Control
Immunoblot Analysis and Subcellular Localization indicates the force level at pCa 6.3 (n⫽5 to 7). C, Effects of
PKC␣ 19-31 (50 mol/L), Y-27632 (2 mol/L), and GST-RB/PH
of Rho-Kinase (TT) (0.5 g/mL) on the Ca2⫹-independent contraction induced
Because translocation of Rho-kinase from the cytosol to the by 0.03 mol/L PDBu. Control indicates the force level at pCa
surface membrane plays an important role in activation of the 6.3 (n⫽3 to 4). Tension levels were expressed as a percentage,
assigning the values in normal relaxing (pCaⰇ8, nominally zero
enzyme,18 we asked if SPC would also induce translocation of
Ca2⫹ with 10 mmol/L EGTA) and activating (pCa 4.5, buffered
Rho-kinase in VSMCs. As shown in Figure 6A, the antibody with 10 mmol/L EGTA) solutions to be 0% and 100%, respec-
against Rho-kinase (ROK␣) recognized a single protein with tively. Data are expressed as the mean⫾SD. *P⬍0.05.
an approximate molecular mass of 180 kDa, as determined by
immunoblotting bovine MCA and rat aorta, and thereby
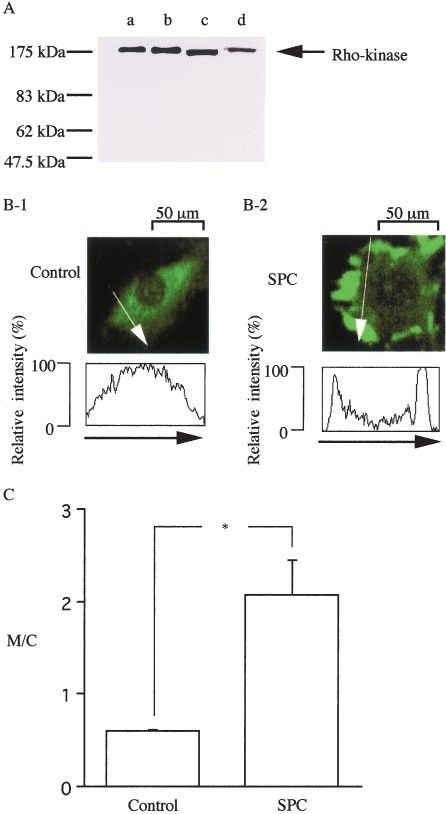
indicating specificity of the antibody for Rho-kinase. Using 6B and 6C). The M/C increased gradually with a time course
the same antibody to immunolocalize ROK␣, the ROK␣ label compatible with slow Ca2⫹ sensitization induced by SPC:
intensities in the cytosol (C) and surface membrane (M) were 1.45⫾0.28 (n⫽12) at 20 minutes and 2.08⫾0.41 (n⫽9) at 30
digitized, and the ratio of the membrane signal to the cytosol minutes. Gong et al30 also noted translocation of Rho-kinase
signal (M/C) was calculated (Figures 6B and 6C). In the to the membrane in portal vein smooth muscle corresponding
control cells (before SPC stimulation), homogeneous distri- to Ca2⫹ sensitization. The nucleus was negatively stained. No
bution of Rho-kinase in the cytosol, but not in the nucleus, specific fluorescence was observed when only the secondary
was observed. Treatment of rat aortic VSMCs in primary antibody (without an anti-ROK␣ antibody) was used for
culture with SPC (30 mol/L, 30 minutes) led to transloca- immunostaining. In addition, increase in the M/C ratio
tion of cytosolic ROK␣ to the free edge of the cells and induced by SPC was reduced by preincubation (30 minutes)
thereby induced a marked increase in M/C (P⬍0.01; Figures with Y-27632.
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015116 Circulation Research July 26, 2002
Figure 7. Effects of PKC inhibitors on contractions induced by
PDBu (0.03 mol/L) or SPC (30 mol/L) in the intact bovine
MCA. A, Control contraction induced by 30 mol/L SPC in the
absence of the inhibitors. B and C, 10 mol/L chelerythrine
chloride (B) and 1 mol/L bisindolylmaleimide 1 (C) were applied
15 minutes before the subsequent application of SPC. D, Sum-
mary of the experiments investigating the effects of PKC inhibi-
Figure 6. Immunoblot detection of Rho-kinase and translocation tors on the PDBu-induced contraction (n⫽5, left) and on the
of Rho-kinase in VSMCs in primary cultures of the rat aorta. A, contractions induced by SPC or 118 mmol/L K⫹ depolarization
Lane a, Positive control (RSV-3T3 lysate, prepared from a (n⫽4 to 12, right). Force was expressed as a percentage,
mouse fibroblast cell line). Lanes b through d, Homogenates of assuming the values in normal (5.9 mmol/L K⫹) and 118 mmol/L
the bovine MCA smooth muscles (10 g of protein) (b), bovine K⫹-PSS to be 0% and 100%, respectively. Data are expressed
brain (9 g of protein) (c), and VSMCs in primary culture of the as the mean⫾SD. *P⬍0.01; **P⬍0.05.
rat aorta (10 g of protein) (d), respectively. Note that Rho-
kinase was recognized to be a single protein with an approxi-
mate molecular mass of 180 kDa. B, Rat VSMCs (Control; B-1) 1 mol/L, and the maximum response was obtained at 0.03
were stimulated with SPC at 30 mol/L for 30 minutes at 37°C mol/L (150.8⫾21.3%, n⫽10, data not shown). Cheleryth-
(B-2). Cells were fixed with paraformaldehyde and stained with
anti-ROK␣ antibody. In B-2, the SPC-induced contraction of
rine chloride (10 mol/L) antagonized the PDBu-induced
cultured VSMCs results in the cell shape change and irregular contraction, with a rightward shift of the concentration-
outline of the cells. C, Changes in the ratio of the membrane response curve (data not shown), and the maximal tension
signal to the cytosol signal (M/C). ROK␣ label intensities were was inhibited from 150.8⫾21.3% (n⫽10) to 35.6⫾12.1%
digitized along the white line (a direction was indicated by an
arrow). The ratio of the intensities of the surface membrane (M) (n⫽5; P⬍0.01; Figure 7D). Bisindolylmaleimide 1 (1
to the intensities of the cytosol (C), M/C, was calculated (n⫽9 to mol/L) also inhibited the PDBu-induced contraction more
15). Data are expressed as the mean⫾SD. *P⬍0.01. potently than did chelerythrine chloride, and the maximal
tension was inhibited from 150.8⫾21.3% (n⫽10) to
Effects of PKC Inhibitors on the SPC-Induced 5.5⫾4.0% (n⫽5; P⬍0.01; Figure 7D).
Contraction in Intact VSM The SPC (30 mol/L)-induced contraction was markedly
The effect of chelerythrine chloride, a nonselective PKC inhibited by a nonselective PKC inhibitor, chelerythrine
inhibitor, and bisindolylmaleimide 1, which is a relatively chloride (10 mol/L) (Figure 7), and was blocked only
selective inhibitor of conventional PKC (cPKC) and novel slightly by a relatively selective blocker of cPKC and nPKC,
PKC (nPKC) isoforms, on the PDBu-induced contraction bisindolylmaleimide 1 (1 mol/L) (Figure 7). Bisindolylma-
were examined using intact VSM strips. PDBu developed a leimide 1 (1 mol/L) also inhibited the 118 mmol/L K⫹-PSS–
concentration-dependent contraction, ranging from 0.0001 to induced contraction from 100% to 85.85⫾6.66% (n⫽6;
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015Shirao et al SPC-Induced Ca2ⴙ Sensitization of Cerebral Artery 117
Figure 7D), a finding compatible with the reported nonspe- (up to 150 kDa) to enter the cytosol of VSM. SPC (30
cific effect of these PKC inhibitors on the Ca2⫹-induced mol/L)-induced Ca2⫹ sensitization was abolished by GST-
contraction.31 This nonspecific effect may be due to the RB/PH (TT) (0.5 g/mL), thus providing direct evidence that
inhibition of a kinase downstream of Rho-kinase.32 SPC-induced Ca2⫹ sensitization is mediated by Rho-kinase
(Figures 3D and 5B). Such specific effects of GST-RB/PH
Discussion (TT) on Rho-kinase–mediated Ca2⫹ sensitization were sup-
The major findings of this study are as follows: (1) SPC, a ported by findings that GST-RB/PH (TT) (up to 1 g/mL)
novel mediator of the Rho-kinase–induced Ca2⫹ sensitization had no effect on Ca2⫹ (pCa 6.3)-induced contraction (Figure
of the cerebral artery, induces a sustained and long-lasting 3D) and on the Ca2⫹ sensitization induced by the PKC
Ca2⫹-independent contraction (with no [Ca2⫹]i elevation), activator, phorbol ester (Figures 4D and 5C), in membrane-
which is characteristic of vasospasm in the cerebral artery; (2) permeabilized vascular strips.
SPC may not require activation of G proteins to induce Ca2⫹ It was reported that sphingolipids, including SPC, are
sensitization and, therefore, may act intracellularly, but not ligands for G protein– coupled membrane receptors, AGR
extracellularly, as agonists for G protein– coupled receptors; 1633 and ovarian cancer G protein– coupled receptor 1
(3) phorbol ester–sensitive PKCs play only a minor role in (OGR1).34 Activation of these sphingolipid receptors induces
SPC-induced Rho-kinase–mediated Ca2⫹ sensitization, and Ca2⫹ mobilization.33,34 However, the present study showed
Rho-kinase apparently does not contribute to the Ca2⫹ sensi- that SPC did induce contraction of bovine MCA with no
tization induced by phorbol ester–sensitive PKCs (cPKC and change in the [Ca2⫹]i (Figure 1). In addition, we also found
nPKC); and (4) translocation of Rho-kinase from the cytosol that SPC-induced Ca2⫹ sensitization was not affected by GTP
to the cell membrane may play a role in SPC-induced Ca2⫹ and GDPS, which respectively activates and inactivates G
sensitization mechanisms. proteins coupled to membrane receptors (including AGR16
In the present study, Ca2⫹ sensitization of the MCA and OGR1). Furthermore, the potency of SPC for OGR1
contraction was evaluated directly using 2 techniques: (1) (Kd⫽33.3 nmol/L) is far different from that noted in the
simultaneous measurement of [Ca2⫹]i and force in intact present study (EC50⫽3.0 mol/L). Taken together, all of
MCA loaded with fura-2 and (2) membrane permeabilization these findings strongly support the notion of intracellular
with -escin or ␣-toxin. In intact MCA, SPC induced a large actions of SPC as an intracellular messenger. Indeed, exog-
and sustained contraction without affecting the [Ca2⫹]i level enously applied SPC is rapidly taken up by intact cells35;
(Figure 1), thereby indicating that the SPC-induced contrac- therefore, the contractile effect of SPC on intact muscle may
tion does not require activation of the Ca2⫹/CaM-dependent not necessarily be mediated through the action of SPC on
MLCK pathway. This Ca2⫹ independence was confirmed by these membrane receptors. It seems highly likely that SPC
findings that 30 mol/L SPC elicited a significant additional may act intracellularly, not extracellularly (as agonist for
increase in force at constant [Ca2⫹]i (pCa 6.3, buffered with these G protein– coupled membrane receptors). We propose
10 mmol/L EGTA) in MCA permeabilized with -escin or that SPC may be a novel mediator for Ca2⫹ sensitization of
␣-toxin (Figures 2, 3A, and 5B). Using a Rho-kinase inhibitor cerebral VSM contraction, with a possible signal transduction
(Y-27632) and a dominant-negative Rho-kinase, we found by Rho-kinase. However, it is also possible that the contrac-
that the SPC-induced contraction of the cerebral artery in the tile effect of SPC may be mediated through the action of SPC
absence of a change in [Ca2⫹]i is mediated by Rho-kinase on yet to be identified membrane receptors not coupled to G
(Figures 1 and 3). This also supports the Ca2⫹-sensitizing protein.
effect of SPC, because we earlier found that cytosolic In the present study, the PKC␣ 19-31 pseudosubstrate had
application of constitutively active Rho-kinase induces max- no effect on the Ca2⫹ (pCa 6.3)-induced contraction of
imal contraction even with complete removal of cytosolic -escin–permeabilized MCA (Figure 5A). In this study, the
Ca2⫹ with 10 mmol/L EGTA in membrane-permeabilized SPC (30 mol/L)-induced Ca2⫹ sensitization was not blocked
VSM,9 which clearly indicates that activation of Rho-kinase by PKC␣ 19-31 (100 mol/L), whereas the PDBu (0.03
induces a Ca2⫹-independent contraction of VSM. We earlier mol/L)-induced Ca2⫹ sensitization was abolished by PKC␣
reported the involvement of Rho-kinase in the SPC-induced 19-31 (50 mol/L) (Figures 3, 4, and 5), thereby indicating a
Ca2⫹ sensitization in the pig coronary artery, although we highly specific effect of PKC␣ 19-31 on PDBu-induced Ca2⫹
used only a Rho-kinase inhibitor and not a dominant-negative sensitization. In contrast, the SPC (30 mol/L)-induced Ca2⫹
Rho-kinase, and PKC involvement was not investigated in sensitization was blocked by preincubation with either
that study.19 Y-27632 (2 mol/L) or GST-RB/PH (TT) (0.5 g/mL),
Although Y-27632 is widely used as a relatively specific whereas the PDBu-induced Ca2⫹ sensitization was not af-
inhibitor of Rho-kinase, the possibility that the observed fected (Figures 3, 4, and 5). These results suggest that the
inhibitory effect of Y-27632 on the SPC-induced Ca2⫹ sen- Ca2⫹ sensitization mediated by phorbol ester–sensitive PKCs
sitization (Figures 1 and 3) may be due to inhibition of other plays only a minor role in SPC-induced Rho-kinase–mediated
unknown protein kinases sensitive to Y-27632, but not of Ca2⫹ sensitization (Figure 8). The notion that PKCs may play
Rho-kinase, needs to be ruled out. Therefore, in the present a role in Ca2⫹ sensitization was based mainly on the finding
study, in order to specifically inhibit Rho-kinase, we exam- that excitatory agonists producing diacyl glycerol (DAG) and
ined the effect of a dominant-negative Rho-kinase, GST- exogenous phorbol esters, both of which activate PKCs, also
RB/PH (TT), on the SPC-induced contraction in the MCA cause Ca2⫹ sensitization.12,36 However, the physiological
permeabilized with -escin, which allows for large molecules roles of PKCs in Ca2⫹ sensitization have recently been
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015118 Circulation Research July 26, 2002
contribute to the Ca2⫹ sensitization induced by SPC and
Rho-kinase. However, our results do not rule out the possible
involvement of aPKC, which is much less sensitive to
bisindolylmaleimide 1 than is chelerythrine chloride, in Ca2⫹
sensitization mediated by a SPC/Rho-kinase pathway. This
notion is also supported by data that aPKC, but not cPKC and
nPKC, may play an important role in the Ca2⫹ sensitization
induced by agonists.23
SPC is produced by N-deacylation of sphingomyelin, one
of the most abundant lipids in cell membrane. In addition, it
is likely, but not proven, that membrane lipids, including
SPC, are released into the perivascular subarachnoid space in
the process of degradation of RBC, WBC, and platelets, as
induced by SAH.40 If SAH induces increasing amounts of
Figure 8. Putative Ca2⫹ sensitization pathways in the cerebral SPC into the cerebrospinal fluid, long term narrowing of the
VSM. SPC induces translocation to the cell membrane and acti- cerebral artery can occur through the vasospasm induced by
vation of Rho-kinase, without affecting the [Ca2⫹]i, which in turn
induces Ca2⫹ sensitization. Y-27632 and GST-RB/PH (TT) inhibit an SPC/Rho-kinase pathway. We propose that SPC is a novel
the activation of Rho-kinase and thereby abolish the Ca2⫹ sensi- mediator for the pathological Ca2⫹-independent contraction
tization induced by SPC. Phorbol 12,13-dibutyrate (PDBu) acti- of the cerebral artery (⫽cerebral vasospasm) as mediated by
vates cPKC and/or nPKC, which in turn induces Ca2⫹ sensitiza-
tion. PKC␣ 19-31 inhibits the activation of cPKC and/or nPKC Rho-kinase, but not by conventional and novel PKCs. A very
and thereby abolishes the Ca2⫹ sensitization induced by PDBu. recent in vivo study demonstrated that Rho-kinase, but not
PKC␣ 19-31 has no apparent effect on the SPC-induced Ca2⫹ PKC, activity contributes to cerebral vascular tone.41 There-
sensitization, and Y-27632 and GST-RB/PH (TT) do not affect
the PDBu-induced Ca2⫹ sensitization. Taken together, our
fore, taken together with this finding, our results in the
results indicate complete separation of the phorbol ester–sensi- present study support the new concept that the Ca2⫹ sensiti-
tive PKC (cPKC and nPKC) pathway and the SPC/Rho-kinase zation induced by Rho-kinase, but not by PKC, may play a
pathway in the bovine cerebral artery VSM. major role also in the maintenance of cerebral artery tone
under normal conditions. To examine the physiological roles
reexamined in more detailed studies of isoforms of PKCs. of SPC, measurements of SPC concentrations in human
cPKC and nPKC, which are both sensitive to phorbol ester, cerebrospinal fluid are currently under investigation.
play only minor roles in the agonists- and G protein–induced Supporting data and expanded text can be found in the online
Ca2⫹ sensitization mediated by Rho-kinase,11,37–39 although data supplement available at http://www.circresaha.org.
atypical (aPKC) does seem to have a role in Ca2⫹ sensitiza-
tion.23 All these studies strongly suggest that phorbol ester– Acknowledgments
sensitive PKCs (cPKC and nPKC) play only minor roles in Y-27632 was a generous gift from Welfide Corporation (Osaka,
Rho-kinase–mediated Ca2⫹ sensitization, as induced by either Japan). This study was supported in part by Grants-in-Aid for
SPC or G proteins. Scientific Research from the Ministry of Education, Science, Tech-
nology, Sports, and Culture, Japan. M. Ohara (Fukuoka) provided
Complete separation of the Ca2⫹-sensitizing pathways me-
language assistance.
diated by SPC/Rho-kinase and phorbol ester–sensitive PKCs
was clearly demonstrated by the cytosolic application of a References
pseudosubstrate peptide of PKC and a dominant-negative 1. Kassell NF, Drake CG. Timing of aneurysm surgery. Neurosurgery.
Rho-kinase to the membrane-permeabilized MCA (Figures 3 1982;10:514 –519.
2. Hartshorne DJ. Biochemistry of the contractile process in smooth muscle.
through 5 and 8). Although membrane permeabilized tech-
In: Johnson LR, ed. Physiology of the Gastrointestinal Tract. New York,
niques are useful tools for investigations of Ca2⫹ sensitization NY; Raven Press; 1987:423-482.
and its signal transduction, membrane permeabilization is not 3. Somlyo AP, Somlyo AV. Signal transduction and regulation in smooth
a physiological condition. Therefore, we also examined this muscle. Nature. 1994;372:231–236.
4. Krueger C, Weir B, Nosko M, Cook D, Norris S. Nimodipine and chronic
point using intact muscles, although use of less specific vasospasm in monkeys, part 2: pharmacological studies of vessels in
pharmacological agents was needed. In the present study, a spasm. Neurosurgery. 1985;16:137–140.
relatively selective inhibitor of cPKC and nPKC isoforms, 5. Nosko M, Weir B, Krueger C, Cook D, Norris S, Overton T, Boisvert D.
Nimodipine and chronic vasospasm in monkeys, part 1: clinical and
bisindolylmaleimide 1 (1 mol/L), abolished the PDBu-
radiological findings. Neurosurgery. 1985;16:129 –136.
induced contraction, whereas at the same concentration, it 6. Haley EC. Jr, Kassell NF, Torner JC. A randomized trial of nicardipine in
had a minimal inhibitory effect on the SPC-induced contrac- subarachnoid hemorrhage: angiographic and transcranial Doppler
tion (Figure 7). In intact bovine MCA, chelerythrine chloride, ultrasound results. A report of the Cooperative Aneurysm Study. J Neu-
rosurg. 1993;78:548 –553.
a nonselective PKC inhibitor, antagonized both PDBu- 7. Haley EC, Kassell NF, Torner JC, Kongable G. Nicardipine ameliorates
induced and SPC-induced contractions (Figure 7). The dif- angiographic vasospasm following subarachnoid hemorrhage (SAH).
ferential effects of bisindolylmaleimide 1 and chelerythrine Neurology. 1991;41:346. Abstract.
8. Kimura K, Ito M, Amano M, Chihara K, Fukata Y, Nakafuku M,
chloride are compatible with our finding that cPKC and
Yamamori B, Feng J, Nakano T, Okawa K, Iwamatsu A, Kaibuchi K.
nPKC, which is blocked by the PKC 19-31 pseudosubstrate Regulation of myosin phosphatase by Rho and Rho-associated kinase
and is more sensitive to bisindolylmaleimide 1, may not (Rho-kinase). Science. 1996;273:245–248.
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015Shirao et al SPC-Induced Ca2ⴙ Sensitization of Cerebral Artery 119
9. Kureishi Y, Kobayashi S, Amano M, Kimura K, Kanaide H, Nakano T, 24. Miyagi Y, Kobayashi S, Nishimura J, Fukui M, Kanaide H. Resting load
Kaibuchi K, Ito M. Rho-associated kinase directly induces smooth muscle regulates cytosolic calcium-force relationship of the contraction of bovine
contraction through myosin light chain phosphorylation. J Biol Chem. cerebrovascular smooth muscle. J Physiol. 1995;484:123–137.
1997;272:12257–12260. 25. Kitazawa T, Kobayashi S, Horiuti K, Somlyo AV, Somlyo AP. Receptor-
10. Uehata M, Ishizaki T, Satoh H, Ono T, Kawahara T, Morishita T, coupled, permeabilized smooth muscle. Role of the phosphatidylinositol
Tamakawa H, Yamagami K, Inui J, Maekawa M, Narumiya S. Calcium cascade, G-proteins, and modulation of the contractile response to Ca2⫹.
sensitization of smooth muscle mediated by a Rho-associated protein J Biol Chem. 1989;264:5339 –5342.
kinase in hypertension. Nature. 1997;389:990 –994. 26. Kobayashi S, Kitazawa T, Somlyo AV, Somlyo AP. Cytosolic heparin
inhibits muscarinic and ␣-adrenergic Ca2⫹ release in smooth muscle:
11. Jensen PE, Gong MC, Somlyo AV, Somlyo AP. Separate upstream and
physiological role of inositol 1,4,5-trisphosphate in pharmacomechanical
convergent downstream pathways of G-protein- and phorbol ester-
coupling. J Biol Chem. 1989;264:17997–18004.
mediated Ca2⫹ sensitization of myosin light chain phosphorylation in
27. Amano M, Chihara K, Nakamura N, Kaneko T, Matsuura Y, Kaibuchi K.
smooth muscle. Biochem J. 1996;318:469 – 475. The COOH terminus of Rho-kinase negatively regulates Rho-kinase
12. Walsh MP, Andrea JE, Allen BG, Clement-Chomienne O, Collins EM, activity. J Biol Chem. 1999;274:32418 –32424.
Morgan KG. Smooth muscle protein kinase C. Can J Physiol Pharmacol. 28. House C, Kemp BE. Protein kinase C contains a pseudosubstrate pro-
1994;72:1392–1399. totope in its regulatory domain. Science. 1987;238:1726 –1728.
13. Gong MC, Fuglsang A, Alessi D, Kobayashi S, Cohen P, Somlyo AV, 29. Iizuka K, Ikebe M, Somlyo AV, Somlyo AP. Introduction of high
Somlyo AP. Arachidonic acid inhibits myosin light chain phosphatase molecular weight (IgG) proteins into receptor coupled, permeabilized
and sensitizes smooth muscle to calcium. J Biol Chem. 1992;267: smooth muscle. Cell Calcium. 1994;16:431– 445.
21492–21498. 30. Gong MC, Gorenne I, Read P, Jia T, Nakamoto RK, Somlyo AV, Somlyo
14. Sato M, Tani E, Fujikawa H, Kaibuchi K. Involvement of Rho- AP. Regulation by GDI of RhoA/Rho-kinase-induced Ca2⫹ sensitization
kinase–mediated phosphorylation of myosin light chain in enhancement of smooth muscle myosin II. Am J Physiol. 2001;281:C257–C269.
of cerebral vasospasm. Circ Res. 2000;87:195–200. 31. Zagorodnyuk V, Santicioli P, Maggi CA. Evidence for the involvement of
15. Nagumo H, Sasaki Y, Ono Y, Okamoto H, Seto M, Takuwa Y. Rho multiple mechanisms in the excitatory action of bradykinin in the circular
kinase inhibitor HA-1077 prevents Rho-mediated myosin phosphatase muscle of guinea-pig colon. Naunyn Schmiedebergs Arch Pharmacol.
inhibition in smooth muscle cells. Am J Physiol Cell Physiol. 2000;278: 1998;357:197–204.
C57–C65. 32. Ichikawa K, Ito M, Hartshorne DJ. Phosphorylation of the large subunit
16. Satoh S, Suzuki Y, Harada T, Ikegaki I, Asano T, Shibuya M, Sugita K, of myosin phosphatase and inhibition of phosphatase activity. J Biol
Chem. 1996;271:4733– 4740.
Hidaka H. Possible prophylactic potential of HA1077, a Ca2⫹ channel
33. Gonda K, Okamoto H, Takuwa N, Yatomi Y, Okazaki H, Sakurai T,
antagonist and vasodilator, on chronic cerebral vasospasm. Eur
Kimura S, Sillard R, Harii K, Takuwa Y. The novel sphingosine
J Pharmacol. 1992;220:243–248.
1-phosphate receptor AGR16 is coupled via pertussis toxin-sensitive and
17. Shibuya M, Suzuki Y, Sugita K, Saito I, Sasaki T, Takakura K, Nagata I, -insensitive G-proteins to multiple signalling pathways. Biochem J. 1999;
Kikuchi H, Takemae T, Hidaka H, Nakashima M. Effect of AT877 on 337:67–75.
cerebral vasospasm after aneurysmal subarachnoid hemorrhage. Results 34. Xu Y, Zhu K, Hong G, Wu W, Baudhuin LM, Xiao Y, Damron DS.
of a prospective placebo-controlled double-blind trial. J Neurosurg. 1992; Sphingosylphosphorylcholine is a ligand for ovarian cancer
76:571–577. G-protein– coupled receptor 1. Nat Cell Biol. 2000;2:261–267.
18. Sin WC, Chen XQ, Leung T, Lim L. RhoA-binding kinase ␣ translo- 35. Desai NN, Carlson RO, Mattie ME, Olivera A, Buckley NE, Seki T, Brooker
cation is facilitated by the collapse of the vimentin intermediate filament G, Spiegel S. Signaling pathways for sphingosylphosphorylcholine-mediated
network. Mol Cell Biol. 1998;18:6325– 6339. mitogenesis in Swiss 3T3 fibroblasts. J Cell Biol. 1993;121:1385–1395.
19. Todoroki-Ikeda N, Mizukami Y, Mogami K, Kusuda T, Yamamoto K, 36. Lee MW, Severson DL. Signal transduction in vascular smooth muscle:
Miyake T, Sato M, Suzuki S, Yamagata H, Hokazono Y, Kobayashi S. diacylglycerol second messengers and PKC action. Am J Physiol. 1994;
Sphingosylphosphorylcholine induces Ca2⫹ sensitization of vascular 267:C659 –C678.
smooth muscle contraction: possible involvement of Rho-kinase. FEBS 37. Hori M, Sato K, Miyamoto S, Ozaki H, Karaki H. Different pathways of
Lett. 2000;482:85–90. calcium sensitization activated by receptor agonists and phorbol esters in
20. Spiegel S, Milstien S. Sphingolipid metabolites: members of a new class vascular smooth muscle. Br J Pharmacol. 1993;110:1527–1531.
of lipid second messengers. J Membr Biol. 1995;146:225–237. 38. Itoh T, Suzuki A, Watanabe Y. Effect of a peptide inhibitor of protein
21. Meyer zu Heringdorf D, van Koppen CJ, Windorfer B, Himmel HM, kinase C on G-protein-mediated increase in myofilament Ca2⫹-sensitivity
in rabbit arterial skinned muscle. Br J Pharmacol. 1994;111:311–317.
Jakobs KH. Calcium signalling by G protein-coupled sphingolipid
39. Fujita A, Takeuchi T, Nakajima H, Nishio H, Hata F. Involvement of
receptors in bovine aortic endothelial cells. Naunyn Schmiedebergs Arch
heterotrimeric GTP-binding protein and rho protein, but not protein
Pharmacol. 1996;354:397– 403. kinase C, in agonist-induced Ca2⫹ sensitization of skinned muscle of
22. Mogami K, Mizukami Y, Todoroki-Ikeda N, Ohmura M, Yoshida K, guinea pig vas deferens. J Pharmacol Exp Ther. 1995;274:555–561.
Miwa S, Matsuzaki M, Matsuda M, Kobayashi S. Sphingosylphospho- 40. Suzuki H, Kanamaru K, Kuroki M, Sun H, Waga S, Miyazawa T. Effects
rylcholine induces cytosolic Ca2⫹ elevation in endothelial cells in situ and of tirilazad mesylate on vasospasm and phospholipid hydroperoxides in a
causes endothelium-dependent relaxation through nitric oxide production primate model of subarachnoid hemorrhage. Stroke. 1999;30:450 – 455;
in bovine coronary artery. FEBS Lett. 1999;457:375–380. discussion 455– 456.
23. Gailly P, Gong MC, Somlyo AV, Somlyo AP. Possible role of atypical 41. Chrissobolis S, Sobey CG. Evidence that Rho-kinase activity contributes
protein kinase C activated by arachidonic acid in Ca2⫹ sensitization of to cerebral vascular tone in vivo and is enhanced during chronic hyper-
rabbit smooth muscle. J Physiol. 1997;500:95–109. tension. Circ Res. 2001;88:774 –779.
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015Sphingosylphosphorylcholine Is a Novel Messenger for Rho-Kinase−Mediated Ca2+
Sensitization in the Bovine Cerebral Artery: Unimportant Role for Protein Kinase C
Satoshi Shirao, Shiro Kashiwagi, Masafumi Sato, Saori Miwa, Fumiaki Nakao, Tetsu Kurokawa,
Natsuko Todoroki-Ikeda, Kimiko Mogami, Yoichi Mizukami, Shinichi Kuriyama, Kyousuke
Haze, Michiyasu Suzuki and Sei Kobayashi
Circ Res. 2002;91:112-119; originally published online June 13, 2002;
doi: 10.1161/01.RES.0000026057.13161.42
Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231
Copyright © 2002 American Heart Association, Inc. All rights reserved.
Print ISSN: 0009-7330. Online ISSN: 1524-4571
The online version of this article, along with updated information and services, is located on the
World Wide Web at:
http://circres.ahajournals.org/content/91/2/112
Data Supplement (unedited) at:
http://circres.ahajournals.org/content/suppl/2002/07/19/91.2.112.DC1.html
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published
in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the
Editorial Office. Once the online version of the published article for which permission is being requested is
located, click Request Permissions in the middle column of the Web page under Services. Further information
about this process is available in the Permissions and Rights Question and Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Circulation Research is online at:
http://circres.ahajournals.org//subscriptions/
Downloaded from http://circres.ahajournals.org/ by guest on May 20, 2015You can also read

















































