The Evolutionary Origins of Programmed Cell Death Signaling
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
The Evolutionary Origins of Programmed
Cell Death Signaling
Kay Hofmann
Institute for Genetics, University of Cologne, Cologne D-50674, Germany
Correspondence: kay.hofmann@uni-koeln.de
Programmed cell death (PCD) pathways are found in many phyla, ranging from developmen-
tally programmed apoptosis in animals to cell-autonomous programmed necrosis pathways
that limit the spread of biotrophic pathogens in multicellular assemblies. Prominent examples
for the latter include animal necroptosis and pyroptosis, plant hypersensitive response (HR),
and fungal heterokaryon incompatibility (HI) pathways. PCD pathways in the different king-
doms show fundamental differences in execution mechanism, morphology of the dying cells,
and in the biological sequelae. Nevertheless, recent studies have revealed remarkable evo-
lutionary parallels, including a striking sequence relationship between the “HeLo” domains
found in the pore-forming components of necroptosis and some types of plant HR and fungal
HI pathways. Other PCD execution components show cross-kingdom conservation as well,
or are derived from prokaryotic ancestors. The currently available data suggest a model,
wherein the primordial eukaryotic PCD pathway used proteins similar to present-day plant
R-proteins and caused necrotic cell death by direct action of Toll and IL-1 receptor (TIR) and
HeLo-like domains.
P rogrammed cell death (PCD), either initiated
cell autonomously in response to pathogens
or stimulated from the outside through signaling
nisms are beginning to be unraveled. At first
glance, PCD pathways in different kingdoms
appear to work by fundamentally different
molecules, is of crucial importance for the suc- rules, although a number of recent studies have
cess of multicellular organisms. Most of current revealed numerous mechanistic parallels and in-
cell death research is focused on animals and stances of clear evolutionary interkingdom rela-
plants, with mammals taking center stage, tionships of cell death mediators. These findings
whereas substantial work is also devoted to mod- suggest that at least a core pathway for PCD has
el metazoans such as Drosophila melanogaster existed in the common ancestor of metazoans,
and Caenorhabditis elegans, or to Arabidopsis fungi, and plants. The following paragraphs will
thaliana as the main model for plant cell death provide a synopsis of major cell death pathways
pathways. However, cell death pathways have in different kingdoms and will highlight the evo-
also been described in other organisms such as lutionary processes leading to the diversification
filamentous fungi, and their molecular mecha- of PCD pathways observed today.
Copyright © 2020 Cold Spring Harbor Laboratory Press; all rights reserved
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
1
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
K. Hofmann
PROGRAMMED CELL DEATH SYSTEMS IN sis,” because the dying cells look morphological-
DIFFERENT KINGDOMS ly different and the signaling cascades and death
effectors appear—at least at first glance—unre-
PCD modalities can be classified as “necrotic” lated to their metazoan counterparts. In the
(i.e., accompanied by membrane rupture and absence of circulating phagocytic cells, a perma-
release of intracellular material), and “non- nent containment characteristic of metazoan
necrotic” without such leakage (Ashkenazi apoptosis is unlikely; whether this containment
and Salvesen 2014). Because metazoan apopto- is important in the absence of an inflammatory
sis is the major—if not the only—example for system is not clear. There is at least one class
the latter type, non-necrotic cell death is usually of PCD pathway in plants called the “hypersen-
referred to as “apoptotic.” As compared with sitive response” (HR), which has been shown
apoptosis, necrotic cell death is less intricate to be associated with DAMP release, therein
and far more widespread. Because cell death resembling metazoan programmed necrosis
caused by major mechanical, chemical, or bio- pathways (Morel and Dangl 1997; Balint-Kurti
logical insults is typically associated with cell 2019). The HR is a part of the so-called “effector
rupture, necrosis was initially considered to triggered immunity” (ETI) system, which gets
be a hallmark of non-PCD and it took a long activated on the detection of pathogen-derived
time until “programmed necrosis” was accepted proteins within the host cell. The HR cell death
as a reality (Edinger and Thompson 2004). is called “hypersensitive” because it exceeds
In animals, several different types of pro- the damage directly inflicted by the pathogen;
grammed necrosis exist, among which necro- its function is to limit the spread of biotrophic
ptosis and pyroptosis are the best understood pathogens. The production and release of
and probably most important examples (Cook- DAMPs—among them the small molecule sali-
son and Brennan 2001; Bergsbaken et al. 2009; cylic acid (SA)—serves the purpose to alert other
Vandenabeele et al. 2010). Cells dying by parts of the plants of the ongoing infection (Ba-
programmed necrosis release intracellular lint-Kurti 2019).
contents, several components of which are in- Outside of animals and plants, PCD path-
terpreted as “damage-associated molecular ways exist (Ameisen 2002), but only a few of
patterns” (DAMPs) by the immune system, re- them have been characterized in molecular
sulting in inflammation (Schaefer 2014; Roh detail. Filamentous fungi belonging to the Asco-
and Sohn 2018). Apoptosis, in contrast, is a mycetes possess a number of functionally
more complicated process because it has to re- analogous, but molecularly diverse cell death
liably kill the cell while at the same time pre- pathways required for a process called “hetero-
venting the leakage of intracellular material karyon incompatibility” (HI) (Saupe 2000; Das-
and DAMPs. This is no easy task, considering kalov et al. 2017). Filamentous ascomycetes grow
that cellular compartments such as the lyso- an extensive network of hyphae, which can both
some and the mitochondrion contain enzymes branch off and merge back—provided that the
and oxidants with the potential to damage cell merging hyphae are genetically identical. Multi-
membranes. During apoptosis, the cell is bro- ple HI systems prevent the successful fusion of
ken into a number of smaller, membrane- hyphae emanating from genetically different in-
enclosed vesicles called “apoptotic bodies,” dividuals, thereby safeguarding against the
which are subsequently removed by phagocytic spread of pathogens (Daskalov et al. 2017). For
processes (Elmore 2007; Nagata 2018). Apopto- a successful hyphal fusion, each of the available
sis is therefore ideally suited for develop- HI systems has to be “disarmed” separately—
mentally scheduled cell death, a physiological usually by the two fusion partners being homo-
process not supposed to alert the immune sys- zygous at a polymorphic sensor locus. Triggering
tem (Fuchs and Steller 2011). only one of the HI systems is sufficient to cause
Nonmetazoan forms of PCD are not easily localized cell death near the point of fusion. The
classified as “apoptosis” or “programmed necro- formation of intrahyphal septa prevents the
2 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
Evolution of Cell Death
spreading of the cell death over the entire hyphal imal caspase-9 molecules can cleave and thereby
system. For several fungal species, in particular activate each other. There are several pathways
for the HI model organism Podospora anserina, leading to apoptosome activation, either cell au-
multiple HI systems and their sensor proteins tonomously or responding to external stimuli
have been described (Saupe 2000; Daskalov via death receptors (Elmore 2007). These up-
et al. 2017; Gonçalves et al. 2017). stream pathways use other caspases (caspase-8,
caspase-10) and other hetero-oligomerization
domains, such as the “death domain” (DD)
MOLECULAR FEATURES OF PROGRAMMED
and the “death effector domain” (DED), which
DEATH PATHWAYS
connect the apoptotic signaling components
This section provides a synopsis of major PCD and can lead to the activation of caspases via
pathways, which are mechanistically under- induced oligomerization and cleavage.
stood to some degree. In particular, the molec-
ular architecture of the key components is sum-
Necroptosis
marized, as this information is necessary to
appreciate the evolutionary ancestry of PCD A simplified version of the core necroptosis
pathways. pathway is shown in Figure 1B, and more de-
tailed descriptions can be found in Newton and
Manning (2016), Weinlich et al. (2017), and
Apoptosis
Petrie et al. (2019). The key step committing a
A simplified version of the apoptosis pathway is cell to necroptosis is the oligomerization and
shown in Figure 1A, and detailed reviews can be activation of the protein kinase RIPK3. RIPK3
found in Elmore (2007) and Nagata (2018). The possesses a central RHIM (RIP homotypic
crucial step in apoptotic cell death induction is interaction motif ), which is important for re-
the activation of caspase-3, a cysteine protease cruitment of RIPK3 to other RHIM-containing
specifically cleaving a number of different sub- proteins, in particular to the related protein
strates, which in combination bring about the kinase RIPK1. Recently, the RIPK1–RIPK3 “ne-
apoptotic phenotype. Apoptosis has, quite ap- crosome” complex was shown to form a RHIM-
propriately, been called “death by a thousand based amyloid fibril with alternating strands
cuts” (Martin and Green 1995). The activation of RIPK1 and RIPK3 (Mompeán et al. 2018).
of caspase-3 happens by proteolytic processing Once part of the oligomeric necrosome, RIPK3
of an inactive precursor; the activating enzyme recruits and phosphorylates a protein called
caspase-9 belongs to the same protease class as MLKL (mixed lineage kinase domain-like).
caspase-3 and several other proteases involved MLKL is an inactive pseudo-kinase with an
in cell death signaling (see below). Caspase-9, in additional amino-terminal four-helix bundle
turn, is activated by formation of a multiprotein (4HB) domain required for cell death execution.
complex called the “apoptosome.” The central On phosphorylation of MLKL, the protein oli-
component of the apoptosome is APAF1, an gomerizes, associates with the cell membrane,
ATPase of the STAND class (Danot et al. causes ion influx, and eventually cells rupture.
2009), which is able to sense the presence of How exactly these events are timed and inter-
cytochrome c released from mitochondria. On connected remains a matter of debate. It is now-
binding to cytochrome c, APAF1 undergoes a adays assumed that membrane-associated
major conformational change, leading to the ex- MLKL oligomers form membrane pores, either
posure of its amino-terminal oligomerization on their own or with the help of other cellular
domain. This so-called CARD (caspase activa- factors (Petrie et al. 2019). Many aspects of nec-
tion and recruitment domain) will then hetero- roptotic cell death can be mimicked by ectopic
oligomerize with another CARD domain found expression of the isolated MLKL amino-termi-
at the amino terminus of the inactive precursor nal domain, thereby obviating the need for up-
of caspase-9. In this oligomeric state, two prox- stream signaling and RIPK3 activity.
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442 3
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
A Ligand
B
Ligand
Apoptosis Necroptosis
Membrane Membrane
DD Membrane DD
Death receptor pore TNFR1
DED DD FADD DD TRADD
DED DED Caspase CASP8 Kinase RH DD RIPK1
Signal Bim Kinase RH RIPK3
Mito
HeLo Kinase MLKL
CytC
CARD NB-ARC APAF1 C Pyroptosis
Membrane
CARD Caspase CASP9 Membrane
pore
Caspase DAMPs
CASP3
PYD NB-ARC NLRP3
Substrates PYD CARD ASC LPS
CARD Caspase CASP11
CARD Caspase CASP1
D Fungal Het-E incompatibility GSD-NT GSD-CT GSDMD
Membrane
Membrane E Hypersensitive response
pore Membrane
Membrane
? pore
Het-C
HET NACHT Het-E AvrAC
? PBL2
RKS1
Fungal Het-S incompatibility CC NB-ARC ZAR1
Membrane
Membrane TIR NB-ARC Wrky
pore RRS1
TIR NB-ARC RPS4
HeLo PFD PFD Het-s AvrRps4
EDS1
HeLo PFD PFD Het-S SAG101 Cell death and
other responses
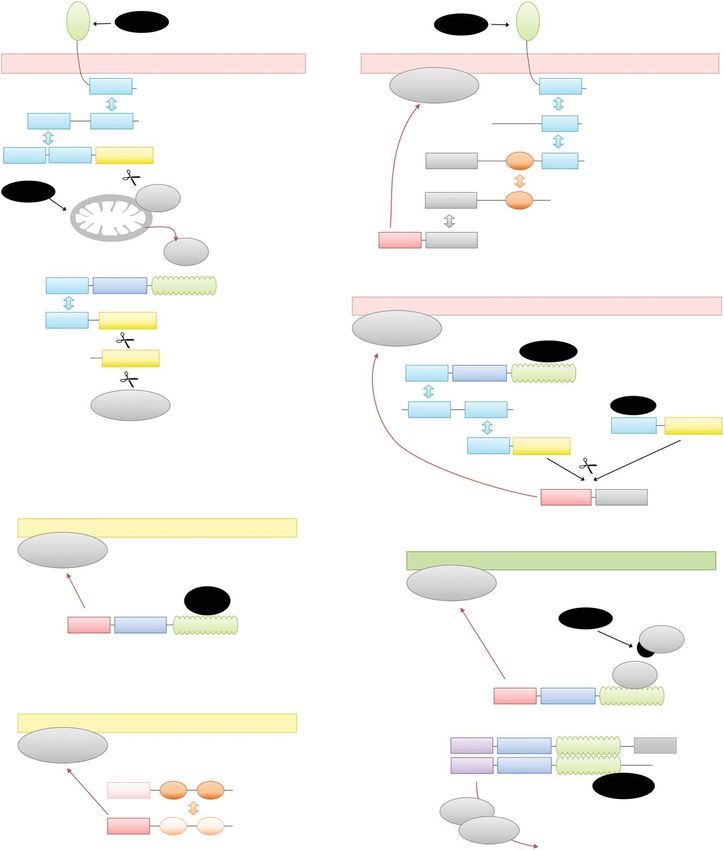
Figure 1. Signaling domains and their functions in programmed cell death (PCD) pathways. This figure shows
simplified versions of major PCD pathways, focusing on the interaction properties of the domains. The arrange-
ment of the proteins in homo-oligomeric complexes is not shown. Coloring: six-helix death-fold domains (death
domain [DD], death effector domain [DED], caspase activation and recruitment domain [CARD], pyrin domain
[PYD]) are shown in cyan, RIP homotypic interaction motifs (RHIMs) in orange, and Toll and IL-1 receptor
(TIR) domains in purple. The central STAND ATPase domains (both NB-ARC and NACHT type) are blue and
repetitive sensor domains (leucine-rich repeat [LRR], WD40) are green. Caspase domains are yellow and sup-
posedly pore-forming domains (gasdermin amino-terminal domain, HET domain, and HeLo/coiled-coil [CC])
are shown in red. All other domain types are shown in gray. Homotypic oligomerization is indicated by double
arrows colored by domain type. Proteolytic cleavage is indicated by a scissors symbol, whereas translocation
events are shown as red arrows. Cell death stimuli are shown on a black background. (A) Apoptosis. Both the
extrinsic pathway, triggered by ligand binding to a death receptor, and the intrinsic pathway initiated by a
mitochondrial signal are shown. (B) Necroptosis. Only the main components of the canonical pathway, triggered
by TNF-receptor type 1 (TNFR1) ligation, is shown here. (C) Pyroptosis. Both major pathways are shown:
caspase-11 (human: caspase-4/5) triggering by intracellular lipopolysaccharide (LPS), and caspase-1 activation
by signalosome signaling. (D) Fungal heterokaryon incompatibility (HI). Two HI systems are shown: Het-E (from
one fusion partner) being triggered by Het-C (contributed by the other partner), and Het-S (from one fusion
partner) being recruited to an amyloid formed by Het-s (from the other fusion partner). (E) Plant hypersensitive
response. One example for each class of R-protein is shown: The CC-NB-LRR (CNL)-based ZAR1 resistosome is
triggered by the ZAR1/RKS1 complex recognizing PBL2, which has been previously modified by the pathogen
effector AvrAC. The dimer of the two TIR-NB-LRR (TNL)-based STAND proteins RPS4 and RRS1 is triggered by
binding to the pathogen effector AvrRps4 and signals cell death via the EDS1/SAG101 complex.
4 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
Evolution of Cell Death
Pyroptosis Heterokaryon Incompatibility
The two main pyroptosis pathways are shown in A simplified depiction of two fungal HI systems
Figure 1C, more detailed descriptions can be is shown in Figure 1D, and more comprehensive
found in Kovacs and Miao (2017), Man et al. descriptions can be found in Saupe (2000), Das-
(2017), and Shi et al. (2017). The key event in kalov et al. (2017), and Gonçalves et al. (2017). A
executing pyroptotic cell death is the proteolytic number of different HI pathways have been de-
cleavage of gasdermin D (GSDMD) at a central scribed in model fungi. Some of these pathways
position, separating the cell-killing amino-termi- are “allelic,” meaning that they are triggered if
nal domain from the inhibitory carboxy-termi- the fused hyphae are heterozygous for a partic-
nal domain. Several (nonapoptotic) caspases are ular polymorphic sensor gene. Other systems
able to cleave GSDMD, depending on the initial are triggered by the interaction of two different
trigger and the upstream signaling pathway. One gene products, each of them contributed by one
subpathway responds to intracellular lipopoly- of the fused cells. The best understood pathway
saccharide (LPS) and cleaves GSDMD by cas- is probably the allelic Het-S system in P.
pase-11 in the mouse and caspase-4/5 in humans. anserina (Seuring et al. 2012; Riek and Saupe
Another pathway depends on inflammasome ac- 2016). Het-s and Het-S are two alleles of a
tivation and uses the major proinflammatory cas- gene encoding a potentially toxic two-domain
pase-1. Once activated, these caspases will not protein. The carboxy-terminal domain can ini-
only cleave GSDMD but also other proteins tiate formation of an amyloid structure, which is
with accessible cleavage sites, most importantly not toxic by itself, but can cluster multiple copies
the proform of interleukin (IL)-1β, thereby form- of the amino-terminal domain. On clustering,
ing the active mature form of this proinflamma- the amino-terminal domain can oligomerize,
tory cytokine. The liberated amino-terminal do- insert into the plasma membrane, and cause
main of GSDMD is thought to undergo a major the loss of membrane integrity. The protein ver-
conformational change, leading to its oligomeri- sion encoded by the Het-s allele is able to initiate
zation and formation of a large membrane pore, an amyloid structure by its carboxy-terminal
which allows the release of the processed IL-1β. “prion-forming domains” (PFDs); however,
Besides GSDMD, other members of the gasder- the amino-terminal “HeLo” domain of the
min family have cleavable amino-terminal do- Het-s protein is not able to permeabilize the
mains that support pore formation (Feng et al. membrane. In contrast, the protein encoded by
2018). Recently, the structure of the gasdermin the Het-S allele has a functional amino-terminal
A3 pore has been solved by cryo-electron micros- domain, but is not able to initiate amyloid for-
copy (EM) and was shown to form a 108-strand- mation owing to a mutation in the PFD region.
ed β-barrel, consisting of 27 gasdermin units When both Het-S and Het-s encoded proteins
(Ruan et al. 2018). Other proteases have also encounter each other during fusion of incom-
been reported to cause gasdermin-dependent py- patible cells, the Het-s protein will initiate am-
roptotic cell death (Xia et al. 2019). In cells infect- yloid formation, whereas the Het-S protein can
ed by the bacterial pathogen Yersinia pestis, the extend these amyloids, thereby triggering the
apoptotic caspase-8 can be activated through a membrane pore formed by the Het-S amino-ter-
multiprotein complex called “RIPoptosome,” minal domain. A fundamentally different incom-
which in turn leads to GSDMD and GSDME patibility mechanism, which is also relevant for
processing by the activated caspase, resulting in the discussion of cell death evolution, is found in
pyroptosis (Orning et al. 2018; Sarhan et al. the non-allelic Het-E/Het-C system and its rela-
2018). The neutrophil-specific elastase ELANE, tives. Here, the central component is the Het-E
a serine-protease unrelated to caspases, has been protein, a STAND-type ATPase with a similar
shown to cleave GSDMD at an alternative site, architecture as the apoptosome component
which also results in the generation of a cytotoxic APAF1 and the NLR components of the inflam-
amino-terminal fragment (Kambara et al. 2018). masome. The carboxy-terminal WD40-repeat
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442 5
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
K. Hofmann
region of Het-E can sense the presence of partic- type of cell death (Paoletti and Clavé 2007). Many
ular alleles of the (unrelated) Het-C gene product. other fungal HI systems exist, but only a few of
On binding to Het-C, the STAND ATPase un- them have been studied for their cell-killing
dergoes a conformational change leading to the mechanism (Fig. 2).
exposure of the Het-E amino-terminal region,
usually referred to as the “HET domain.” Unlike
Hypersensitive Response
the situation in apoptosis and inflammasome ac-
tivation, these HET domains are not thought to Two examples of plant defense signaling leading
be recruitment domains, but rather to directly to cell death are shown in Figure 1E. More de-
disintegrate the membrane, leading to a necrotic tailed descriptions can be found in Coll et al.
Mammals
TIR
Insects
DD TIR
RH PGRP
Kinase RH DD
RH DD
ZBP1 RH RH RH
CARD NB-ARC CARD CARD RIG-I
+ PYD
DD Kinase HeLo Kinase
PYD CARD
+ RIG-I
CARD NACHT
+ ZBP1
PYD NACHT
CARD NB-ARC
+ DD
+ DED Apoptosis
Fungi Death receptors
HeLo NB-ARC
+ CARD + NF-κB
HeLo NACHT
Caspases Plants
RH NACHT Kinase
HeLo RH RH
+ HET HeLo NB-ARC
HET NACHT
+ CC CC NB-ARC
HeLo Kinase TIR NB-ARC
+ RHIM HeLo Kinase
EDS1-
+ signaling
Ancestral
HeLo NB-ARC
TIR NB-ARC
X NACHT
HeLo Kinase
Figure 2. Evolutionary origins of cell death protein architectures. This figure shows generic protein architectures
found in metazoans, fungi, and plants—and what is predicted to have existed in an ancestral (early eukaryote)
organism. Domain coloring identical to Figure 1. The ancestral system is predicted to mainly have used STAND
ATPases, most likely of the NB-ARC type, with amino-terminal Toll and IL-1 receptor (TIR) and HeLo-like
domains. However, NACHT-ATPases and a mixed-lineage kinase domain-like (MLKL) HeLo-domain kinase
might also have been present. In the plant lineage, components of EDS1 and probably many other relevant
proteins have been acquired. The coiled-coil [CC]-domain found in present plant CC-NB-LRR (CNL) proteins
was either acquired or, more likely, evolved from the HeLo domain. RIP homotypic interaction motifs (RHIMs)
are first seen in the opisthokont lineage, whereas the six-helix death-fold domains are specific for the metazoan
lineage. A more detailed description of early cell death evolution is given in the main text.
6 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
Evolution of Cell Death
(2011), Jones et al. (2016), and Balint-Kurti ing a common evolutionary history becomes
(2019). Although plant defense mechanisms apparent.
against pathogens have been studied intensively,
and protective cell death by the HR is one im-
STAND ATPases in Cell Death and Immunity
portant branch of this defense, the exact cell
death mechanism and the death-specific sig- One of the most obvious recurring features in
naling components are much less clear. The cell death signaling is the use of STAND-type
central orchestrators of intracellular defense ATPases as a signaling hub (Leipe et al. 2004;
pathways are the “R-proteins” (resistance pro- Danot et al. 2009). The name STAND, originally
teins). Most R-proteins belong to the class of an acronym for “signal transducing ATPases
STAND-type ATPases with a leucine-rich re- with numerous domains,” encompasses two
peat (LRR) domain at the carboxyl terminus major subfamilies of large ATPases of similar
and an effector/signaling domain at the amino core architecture, which are both important
terminus. Like all other STAND-type ATPases, for cell death signaling. One class has a central
R-proteins are thought to undergo a structural “NB-ARC” ATPase domain (named after the
rearrangement on binding to its cognate ligand, proteins APAF1/R-Proteins/Ced-4), the other
which may be bacterial effectors secreted into class has a central “NACHT” ATPase domain
the host cell or any other molecule indicative (named after NAIP/CIIA/Het-E/TEP1). Both
of biotic stress. Depending on the nature of the classes contain carboxy-terminal sensor re-
amino-terminal domain, different downstream gions, typically consisting of repeat-forming do-
signaling pathways can be engaged, eventually mains, such as LRRs, WD40 repeats, or tetra-
leading to the induction of defense genes, the tricopeptide repeats (TPRs). At the amino
generation and release of the long-range signal- terminus, different effector domains can be
ing molecule SA, and/or to the induction of hy- found. Both NB-ARC and NACHT ATPases
persensitive cell death. The factors required for work similarly. In the resting state, the proteins
cell death execution are not known and depend are monomeric and show a “closed” conforma-
on the R-protein, which might belong to one of tion, which shields the amino-terminal effector
several subtypes. The TNL proteins carry an domain. On binding of the carboxy-terminal
amino-terminal Toll and IL-1 receptor (TIR) sensor domain to its cognate stimulus, the cen-
domain and signal cell death via the EDS1- tral ATPase domain undergoes a conformation-
SAG101-NRG1 pathway (Lapin et al. 2019) al- al change, causing an oligomerization of the
though the details of that mechanism are not ATPase and leading to the exposure of the ami-
known. A second class of R-proteins are the no-terminal effector domains. This process is
“CC-NB-LRR” (CNL) proteins, which carry an best understood for the NB-ARC protein
amino-terminal domain that was initially con- APAF1, the central component of the apopto-
sidered to form a coiled-coil (CC) structure, al- some (Cheng et al. 2016; Dorstyn et al. 2018).
though this is not necessarily true. At least some Here, the active conformation has the shape of
members of the CNL-type R-proteins can in- a heptameric “wheel” formed by seven ATPase
duce cell death dependent on the amino-termi- domains; the amino-terminal CARD domains
nal domain (Collier et al. 2011), although the of the seven APAF1 units are found clustered
exact mechanism remains unresolved. above the plane of the ATPase wheel. Ced-4,
the APAF1 homolog from the nematode
C. elegans, forms an octameric ring with a sim-
RECURRING MOTIFS AND MECHANISMS IN
ilar domain arrangement (Qi et al. 2010; Huang
CELL DEATH SIGNALING
et al. 2013).
When comparing the mechanisms of the cell The vast majority of eukaryotic NB-ARC
death pathways described above, in particular and NACHT proteins are known or suspected
the domain architecture of the key proteins in- to be involved in innate immunity and cell death
volved, a number of recurring features suggest- signaling. As mentioned before, the (sole) hu-
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442 7
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
K. Hofmann
man NB-ARC protein APAF1 and its nema- type R-proteins. When comparing the domain
tode homolog Ced-4 use their amino-terminal architectures of APAF1 (CARD, NB-ARC,
CARD domain for apoptosome formation. WD40) with that of ZAR1 (CC, NB-ARC,
Mammals possess several NACHT proteins LRR) or HET-E (HET, NACHT, WD40), it be-
with amino-terminal CARD or pyrin domains comes obvious that these proteins have the same
(PYDs), which are the key components of “in- overall architecture but did not evolve by diver-
flammasomes,” protein complexes similar to the gent evolution from a common ancestor. It can
apoptosome but activating caspase-1 rather be assumed that early eukaryotes already con-
than caspase-9 and thereby leading to pyropto- tained STAND-ATPases because most extant
sis (Broz and Dixit 2016). bacteria encode several such ATPases. However,
The relationship between the mammalian the classical bacterial STAND-ATPases do not
STAND ATPases and the plant R-proteins is belong to the NB-ARC or NACHT subtype and
readily visible and has stimulated a number of have no connection to cell death signaling
analyses comparing animal and plant cell- (Leipe et al. 2004). A careful phylogenetic anal-
autonomous immunity (Maekawa et al. 2011; ysis of the STAND family concluded that plant
Jones et al. 2016; Urbach and Ausubel 2017). and animal STAND proteins evolved indepen-
Recently, it was shown by cryo-EM that plant dently from bacterial precursors, in at least two
R-proteins can also form wheel-like oligomers. separate events (Urbach and Ausubel 2017).
In the example of ZAR1, an R-protein of the However, STAND ATPases and other innate
CNL class, the wheel has a pentameric structure immunity proteins are clearly subject to evolu-
called the “ZAR1 resistosome” to emphasize the tion by “domain swapping,” and when analyz-
analogy to the metazoan apoptosome and in- ing eukaryotic proteomes available nowadays,
flammasome complexes (Wang et al. 2019). In virtually all combinations of effector domains,
the activated ZAR1 resistosome, the five individ- ATPase subtype, and carboxy-terminal sensor
ual “CC-type” effector domains undergo a con- domains can be observed. Therefore, alternative
formational rearrangement and form a pore-like evolutionary hypotheses should not be dis-
α-barrel with an additional funnel-like structure counted, for example, a pervasive “mixing and
formed by the first α-helices of each CC domain. matching” of domains in early eukaryotic evo-
This funnel appears to be crucial for cell death lution, with a subsequent “fixing” of select do-
induction by activated ZAR1 (Wang et al. 2019). main architectures that proved most beneficial
Although there is no formal proof that the CC for the particular eukaryotic lineage.
domain of ZAR1 forms a membrane pore suffi-
cient for ion influx or even cell rupture, it
Evolution of Effector and Oligomerization
appears that the plant R-proteins use a more
Domains
direct way to cell death than their metazoan
counterparts, which rely on further downstream The formation of transient multiprotein com-
signaling. plexes, leading to the recruitment of adaptor
Several of the fungal HI systems also make and effector proteins to activated receptors, is
use of STAND ATPases. The Het-E protein a recurring feature of apoptosis, necroptosis,
mentioned above, but also Het-D and Het-R and pyroptosis signaling. In metazoan systems,
all contain a central NACHT ATPase domain, the prevalent oligomerization domain types are
followed by a carboxy-terminal sensor domain the DD, the DED, the CARD, and the PYD. All
consisting of WD40-repeats. The amino-termi- four domain types are distantly related to each
nal effector domains (HET-domains) are only other (Hofmann 1999; Park et al. 2007a; Kersse
found in filamentous fungi and are thought to et al. 2011) and share a common structural fold
directly form membrane pores on oligomeriza- consisting of six α-helices, often referred to as
tion (Paoletti and Clavé 2007). This mode of the “death fold” or “six-helix bundle fold.” The
action would be analogous to what has been principal function of all four domain classes is
suggested for ZAR1 and possibly other CC- to recruit another domain of the same class.
8 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
Evolution of Cell Death
Despite the relationship between DD, DED, TRAM, and TRIF. Typical downstream events
CARD, and PYD, interactions across the class of mammalian TIR signaling include the up-
boundaries are rare. It is interesting to note that regulation of defense genes via transcription fac-
RHIM motifs and TIR domains fulfil analogous tors of the NF-κB (nuclear factor κ light chain
roles of “recruitment by oligomerization,” but enhancer) or IRF (interferon regulatory factor)
are not related to the death-fold superfamily families, but also necroptosis induced via the
and do not share the six-helix bundle structure RHIM motif of TRIF. Despite a completely un-
(Nanson et al. 2019). related structure, the role of TIR domains is re-
Despite exhaustive bioinformatical searches markably similar to members of the six-helix
(Hofmann 1999; unpubl. data), no members of death-fold family. This analogy is further em-
the death-fold superfamily could be detected phasized by the ability of TIR domains to form
outside of metazoans, with the exception of vi- filaments akin to those of the death-fold
ruses and other pathogens, which have probably domains (Ve et al. 2017; Nanson et al. 2019).
acquired these domains from their metazoan Because there clearly is no evolutionary relation-
hosts (Thome et al. 1997; Lamkanfi and Dixit ship between these domain classes, oligomeri-
2010). It thus appears that the six-helix death zation through TIR and death-fold domains is
fold arose during metazoan evolution. It is also another example of convergent evolution.
remarkable that all characterized members of In contrast to animals, plants do not use TIR
the death-fold superfamily reside in proteins in- domains in their surface receptors, but rather as
volved in cell death and innate immunity signal- the amino-terminal effector domains of one
ing; other pathways requiring oligomerization class of intracellular R-proteins (TIR-NB-LRR
appear to use other domains for this purpose. or TNL type). Surprisingly, plants lack TIR-con-
A possible reason for this pathway specificity taining adaptor proteins and the TNL proteins
might lie in the availability of multiple interac- signal via a complex of EDS1 (enhanced disease
tion surfaces of the fold, which can be used si- susceptibility 1) and SAG101 (senescence-asso-
multaneously and thus support the formation of ciated gene 101) through an as-yet uncharacter-
higher-order oligomeric structures called “fila- ized mechanism (Lapin et al. 2019). However,
ments” (Park et al. 2007b; Kersse et al. 2011; plant TIR domains share with their metazoan
Hauenstein et al. 2015). Such filaments can be counterparts the capacity to mediate dimeriza-
nucleated by a di- or trimerization and then tion between different R-proteins (Bernoux et al.
grow by using the remaining interaction sur- 2011; Williams et al. 2014; Zhang et al. 2017).
faces—analogous to an amyloid but without An interesting twist to the enigma of TIR sig-
the β-stack structure typical of real amyloids. naling was introduced recently by detecting an
These cooperatively formed higher-order struc- NAD-cleaving activity for the TIR domain of
tures are instrumental for inflammasome for- the human SARM1 (sterile α and TIR motif )
mation (Hauenstein et al. 2015) and support protein (Essuman et al. 2017), as well as in sev-
the “all-or-nothing” characteristics required eral bacterial TIR proteins (Essuman et al.
for life/death decisions. 2018). SARM1 has a role in mediating neuro-
degeneration and the NAD depletion by its TIR
domain was suggested to be a main factor in this
Evolution of TIR Domains
process (Essuman et al. 2017). It is unlikely that
TIR domains have a particularly interesting evo- other human TIR domains share this enzymatic
lutionary history. In mammals, TIR domains are activity, because they either lack the active site
found at the cytoplasmic end of Toll-like recep- glutamate residue or have a structure that places
tors (TLRs) and receptors for IL-1 cytokines— this residue outside the catalytic cleft. However,
all of them receptors that alert a cell to a danger it remains possible that in the presence of suit-
situation from the outside. Their function is the able binding partners or posttranslational mod-
recruitment of other TIR domains found in ifications, the structure may be converted into
“adaptor proteins” such as MYD88, TIRAP, an active form. Plant TIR domains, in contrast,
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442 9
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
K. Hofmann
tend to show conservation in the active site re- the fungal PFDs form an amyloid of similar
gion, and the few available structures appear to structure (Wasmer et al. 2008; Riek and Saupe
support catalysis. It is therefore an intriguing 2016), supporting a common evolutionary ori-
possibility that either NAD depletion, or the gin of these two oligomerization systems. This
generation of (cyclic) ADP-ribose as the NAD evolutionary parallel is further underscored by
degradation product, play a role in cell death the finding that the “HeLo domain,” the cell-
signaling by R-proteins of the TIR-NB-LRR killing moiety of the Het-S system, is related to
architecture. Given that TIR proteins are abun- the functionally analogous amino-terminal do-
dant in extant bacteria and appear to be gener- main of MLKL (Daskalov et al. 2016). It is thus
ally catalytically active (Essuman et al. 2018), it very likely that the metazoan necroptosis system
is highly likely that both animal and plant TIR evolved from a simpler precursor similar to Het-
domains evolved from a prokaryotic ancestor S, with amyloid-forming domain and killing
and later gained the capacity for oligomeriza- domain within the same polypeptide. During
tion, although (mostly) losing their catalytic ac- vertebrate evolution, the two functionalities
tivity in the process. were probably split into separate proteins. It
is difficult to decide from the available data
whether the common precursor was using a
Evolutionary History of Necroptosis Execution
one-component amyloid (as in Het-S) or a
The execution phase of necroptosis is character- two-component version (as in RIPK1/RIPK3).
ized by MLKL phosphorylation and oligomeri- Because even the one-component amyloid uses
zation, initiated by formation of a mixed amy- two alternating strands—PFD1 and PFD2 of
loid structure formed by the RHIM motifs of Het-S—it appears more likely that the extant
RIPK1 and RIPK3 (Mompeán et al. 2018; Petrie Het-S system is a degenerate version of a former
et al. 2019), or possibly by similar structures two-component systems, now perfectly adapted
formed by RIPK3 with other RHIM proteins to the task of detecting heterozygosity.
such as TRIF or ZBP1/DAI. Despite its funda-
mentally different structure, the role of RHIMs
Evolutionary History of Pyroptosis and
in necroptosis signaling is analogous to those of
Caspases
six-helix death-fold domains or TIR domains.
All of these domains form homotypic oligomers The decisive step in pyroptosis execution is the
and have the tendency to create higher-order cleavage of GSDMD by caspase-1 or other cas-
superstructures. In the case of RHIM, this pases with similar cleavage specificities. Cas-
superstructure is a “real” amyloid (Li et al. pase-1 itself is activated by inflammasomes,
2012; Mompeán et al. 2018). In evolution, which can be induced by many proinflammatory
RHIM-based oligomerization is far more wide- stimuli. The fact that caspase-1 also activates IL-
spread than the few examples known in mam- 1β makes pyroptosis the major modality for re-
mals. A bioinformatical analysis showed a large leasing processed IL-1β and thus causing the in-
array of RHIM-containing proteins in non- flammatory phenotype (Man et al. 2017; Green
mammalian metazoans. In many of these cases, 2019). The other GSDMD-cleaving caspases 4, 5,
the RHIM appears to substitute for TIR or six- and 11 are directly activated by intracellular LPS.
helix death-fold domains, which are found in The details of this activation are not fully under-
the mammalian version of these proteins (Ka- stood. All of the GSDMD-cleaving caspases are
java et al. 2014). This finding suggests that thought to oligomerize via their amino-terminal
RHIM motifs work similar to TIRs and death- CARD domains, but the involvement of fila-
fold domains and actually can replace them. ments of higher-order structures has not been
Interestingly, the same bioinformatical anal- reported.
yses found an evolutionary relationship between Neither gasdermins nor “proper” caspases
RHIM motifs and the PFDs used in the Het-S are found outside of metazoans; the IL-1 cyto-
system of HI (Kajava et al. 2014). Like RHIMs, kine family is even restricted to vertebrates.
10 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
Evolution of Cell Death
Thus, pyroptosis, like apoptosis, appears to be a to cause pyroptosis (Wang et al. 2017; Jiang et al.
relatively recent addition to the arsenal of PCD 2019). In contrast, the sixth gasdermin, Pejva-
pathways. On the other hand, the metazoan cas- kin/PJVK, does not appear to be toxic (Feng
pases are related to two other classes of cysteine et al. 2018) but has homologs in invertebrates.
proteases, the metacaspases and paracaspases, The exact role of PJVK is not known; the human
which have a much wider evolutionary distribu- gene is implicated in nonsyndromic hearing loss
tion and are even found in bacteria (Koonin and (Harris et al. 2017) and has been proposed to
Aravind 2002). Initially, the finding of metacas- regulate pexophagy, the autophagic removal of
pases in yeasts, together with the finding that peroxisomes (Defourny et al. 2019). Based on
overexpression of some mammalian apoptosis these considerations, it appears likely that the
proteins killed yeast cells, gave rise to specula- metazoan gasdermin family evolved from a Pej-
tions that caspase-dependent cell death or even vakin-like precursor and possibly acquired the
apoptosis might be conserved in lower eukary- cell-killing activity of the amino-terminal later
otes (Váchová and Palková 2007). By now, it is on. However, it remains possible that the proto-
known that metacaspases, despite their evolu- gasdermin gene encoded a cell-killing protein
tionary relationship to caspases, have a totally and that Pejvakin lost this activity. As a third
different cleavage specificity (cleavage after ar- possibility, Pejvakin might still be able to kill
ginine rather than aspartate) and cannot replace cells, but requires a specific activation mode
the function of proper metazoan caspases awaiting to be discovered. A much older evolu-
(Tsiatsiani et al. 2011). Nevertheless, metacas- tionary history—or possibly a horizontal acqui-
pases might have a more general role in DAMP sition—is suggested by the recently published
processing during necrotic cell death. A recent structure of the mouse gasdermin A3 pore
study showed that plant metacaspases process a (Ruan et al. 2018), which is probably a good
cytoplasmic immunomodulatory plant protein, model for other gasdermin pores as well. Both
the active part of which PEP1 ( plant elicitor the pore structure and the conformational
peptide 1) is secreted from damaged cells and change undergone by the GSDMA3 amino-ter-
acts as a defense signal (Hander et al. 2019). This minal domain on pore formation strongly resem-
situation is comparable to the IL-1β release dur- ble the pores of bacterial cytolysins such as Pneu-
ing pyroptosis but without any sequence or molysin and Perfringolysin O, but also the pores
structural similarity between PEP1 and the of mammalian perforin and the membrane at-
metazoan interleukin. tack complex (MAC) of the complement system
Another question of evolutionary relevance (Ruan et al. 2018). Despite the structural similar-
concerns the origins of the gasdermin family. Of ities of these pores, there is no overt sequence
the six gasdermins known in the human ge- similarity, which makes it difficult to judge
nome, four are relatively closely related to each whether these proteins are truly related or just
other (GSDMA, GSDMB, GSDMC, GSDMD). further examples of convergent evolution.
This subfamily is fast evolving, with some addi-
tional members in rodents and other mammals.
HOW IT ALL BEGAN
Common to all these proteins is the architecture
with an amino-terminal toxic domain, a car- An inventory of present-day cell death com-
boxy-terminal inhibitory domain, and a prote- ponents, their domain architecture, and their
ase cleavage site in the middle. Two more gas- interrelationship should, in principle, allow the
dermins (DFNA5/GSDME and Pejvakin/PJVK) reconstruction of key evolutionary events shap-
form a second subfamily, which is somewhat ing cell death signaling. However, the fast
more distantly related, in particular in the quite evolution of some components, the pervasive
divergent inhibitory region. GSDME, which un- domain shuffling, and the incomplete knowl-
like gasdermin A-D has homologs in fish, be- edge of death signaling outside the classical
haves like a classical gasdermin; it is cleaved by model organisms make this task difficult if not
caspases and its amino-terminal domain is able impossible. The sequence of evolutionary events
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442 11
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
K. Hofmann
proposed here is in accordance with the avail- cell death system, triggered by some pathogen-
able data, but there is no certainty that it reflects or damage-derived molecule and leading to cell
the history correctly. death by NAD depletion. Another early cell
Because STAND-type ATPases are abun- death system could have used a HeLo-like do-
dant in both pro- and eukaryotes, and fulfill main as the effector. Although HeLo domains
similar purposes in multiple kingdoms, it is have not been detected in bacteria, several copies
safe to assume that STAND-based signaling of this domain exist in plants, fungi, animals,
was available in early eukaryotes including the and other eukaryotes. Because the HeLo domain
last common ancestor to animals, fungi, and appears to cause membrane pores autonomous-
plants. At the carboxyl terminus of extant ly, at least in fungi, this domain is an equally
STAND ATPases, many different sensor do- good candidate for an early PCD system. In-
mains are found, typically repeat domains. In terestingly, the two most likely architectures
plants, most of the sensing appears to be per- resemble the two classes of plant R-proteins:
formed by LRR domains, which are also com- TIR-NB-LRR and CC-NB-LRR. However, it is
mon in mammalian STAND ATPases. Howev- well possible that the ancient versions of this
er, when including invertebrates and fungi system used another subtype of STAND
(which are more closely related to animals ATPase, or another type of sensor repeat.
than plants), a much greater sensor diversity is Cell death systems relying on caspase activ-
observed, including WD40, TPR, and ankyrin ity, such as apoptosis and pyroptosis, most likely
repeats. One published study concluded that the did not develop before the advent of metazoans,
NB-ARC subtype (as used in plant R-proteins) although it cannot be excluded that there have
and the NACHT subtype (as used in animal been pyroptosis-like systems using another type
inflammasomes and fungal Het-E) evolved in- of protease. However, there are no indications
dependently from a bacterial ancestor (Urbach for such a system in the presently available data.
and Ausubel 2017). This study assumed bacte- Apoptosis signaling, at least in its present mam-
ria-specific subfamilies of the STAND super- malian form, relies not only on caspases but also
family as the precursor, from which eukaryot- on numerous six-helix death-fold domains, all
ic NACHT and NB-ARC subtypes evolved. of which appear only in metazoans, suggesting
Whereas this is undoubtedly possible, there are that apoptosis is probably the youngest of the
plenty of bacterial NB-ARC and NACHT mem- PCD systems discussed here. In contrast, nec-
bers in present-day genomes. This observation roptosis appears to be ancient, at least the down-
means that either the two independent events stream events. Necroptotic cell death is caused
leading to NACHT and NB-ARC happened be- by the HeLo domain of MLKL, which is clearly
fore the split of eukaryotes, or that the NACHT ancient, whereas RHIM motifs are only found in
and NB-ARC proteins seen in extant bacteria animals and the related PFD of Het-s has not
are late horizontal acquisitions from a eukary- been found outside of fungi. Nevertheless, we
otic source. Most likely, early eukaryotes coded recently identified an MLKL pseudokinase in
for NACHT and NB-ARC-type ATPases with a plants, whose amino-terminal domain forms a
wide range of possible sensor motifs. 4HB similar to that of mammalian MLKL and is
A most relevant question of cell death evo- also able to cause cell death (Mahdi et al. 2019).
lution is what kind of effector domains were Although plants appear to lack RIPK3 homo-
used by these early signaling hubs. One prime logs and other RHIM-based signaling proteins,
candidate are the TIR domains, they clearly pre- this finding suggests that at least the execution
date the advent of eukaryotes, and they were step of necroptosis predates the split of animals
most likely enzymatically active like the extant and plants.
bacterial versions (Essuman et al. 2018). Actu- When considering the later evolutionary
ally, the NAD-degrading function of TIR do- steps of cell death pathways, there are several
mains in the context of a STAND ATPase might examples of intermediate steps being added,
have formed the basis for a very early eukaryotic probably to allow more regulatory layers, or to
12 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
Evolution of Cell Death
further amplify the reaction—making PCD Bergsbaken T, Fink SL, Cookson BT. 2009. Pyroptosis: host
cell death and inflammation. Nat Rev Microbiol 7: 99–
induction an “all-or-nothing” decision. One ex- 109. doi:10.1038/nrmicro2070
ample is the ( proposed) conversion of TIR do- Bernoux M, Ve T, Williams S, Warren C, Hatters D, Valkov
mains from its original catalytic form, thought E, Zhang X, Ellis JG, Kobe B, Dodds PN. 2011. Structural
to exert a direct killing effect, into a signaling and functional analysis of a plant resistance protein TIR
domain reveals interfaces for self-association, signaling,
domain that recruits further TIR-containing and autoregulation. Cell Host Microbe 9: 200–211. doi:10
proteins in large numbers. A similar change .1016/j.chom.2011.02.009
might have occurred to the HeLo domain, Broz P, Dixit VM. 2016. Inflammasomes: mechanism of
thought to be a direct cell-killing domain in assembly, regulation and signalling. Nat Rev Immunol
16: 407–420. doi:10.1038/nri.2016.58
fungi, into a possible signaling domain in plants.
Cheng TC, Hong C, Akey IV, Yuan S, Akey CW. 2016. A near
On a smaller scale, the addition of regulatory atomic structure of the active human apoptosome. eLife 5:
layers is also visible in apoptosis evolution e17755.
among metazoans. Whereas in nematodes, the Coll NS, Epple P, Dangl JL. 2011. Programmed cell death in
Ced-4 apoptosome directly recruits the effector the plant immune system. Cell Death Differ 18: 1247–
1256. doi:10.1038/cdd.2011.37
caspase Ced-3, the mammalian APAF1 apopto- Collier SM, Hamel LP, Moffett P. 2011. Cell death mediated
some recruits and activates an intermediate cas- by the N-terminal domains of a unique and highly con-
pase (caspase-9), each molecule of which can served class of NB-LRR protein. Mol Plant Microbe Inter-
then activate multiple copies of the mammalian act 24: 918–931. doi:10.1094/MPMI-03-11-0050
Cookson BT, Brennan MA. 2001. Pro-inflammatory pro-
effector caspase-3. It might even be possible that grammed cell death. Trends Microbiol 9: 113–114.
the RHIM motifs and six-helix death-fold do- doi:10.1016/S0966-842X(00)01936-3
mains, in their original form, did have a direct Danot O, Marquenet E, Vidal-Ingigliardi D, Richet E. 2009.
cell-killing effect and only later evolved into sig- Wheel of life, wheel of death: a mechanistic insight into
signaling by STAND proteins. Structure 17: 172–182.
naling and recruitment domains. However, this doi:10.1016/j.str.2009.01.001
remains speculative because there are no data to Daskalov A, Habenstein B, Sabaté R, Berbon M, Martinez D,
support that idea. Taken together, it becomes Chaignepain S, Coulary-Salin B, Hofmann K, Loquet A,
clear that PCD pathways across kingdoms use Saupe SJ. 2016. Identification of a novel cell death-induc-
ing domain reveals that fungal amyloid-controlled pro-
a similar signaling logic and evolved from a grammed cell death is related to necroptosis. Proc Natl
common ancestral pathway, probably much Acad Sci 113: 2720–2725. doi:10.1073/pnas.1522361113
simpler than the intricate multilayer systems ob- Daskalov A, Heller J, Herzog S, Fleißner A, Glass NL. 2017.
served in multicellular organisms. Molecular mechanisms regulating cell fusion and hetero-
karyon formation in filamentous fungi. Microbiol Spectr
5. doi:10.1128/microbiolspec.FUNK-0015-2016
Defourny J, Aghaie A, Perfettini I, Avan P, Delmaghani S,
ACKNOWLEDGMENTS Petit C. 2019. Pejvakin-mediated pexophagy protects au-
ditory hair cells against noise-induced damage. Proc Natl
We thank Shuhua Chen for valuable discussions. Acad Sci 116: 8010–8017. doi:10.1073/pnas.1821844116
This work was funded by a grant from the Dorstyn L, Akey CW, Kumar S. 2018. New insights into
apoptosome structure and function. Cell Death Differ
Deutsche Forschungsgemeinschaft (SFB 670). 25: 1194–1208. doi:10.1038/s41418-017-0025-z
Edinger AL, Thompson CB. 2004. Death by design: apopto-
sis, necrosis and autophagy. Curr Opin Cell Biol 16: 663–
669. doi:10.1016/j.ceb.2004.09.011
REFERENCES
Elmore S. 2007. Apoptosis: a review of programmed cell
Reference is also in this collection. death. Toxicol Pathol 35: 495–516. doi:10.1080/0192
6230701320337
Ameisen JC. 2002. On the origin, evolution, and nature of Essuman K, Summers DW, Sasaki Y, Mao X, DiAntonio A,
programmed cell death: a timeline of four billion years. Milbrandt J. 2017. The SARM1 Toll/interleukin-1 recep-
Cell Death Differ 9: 367–393. doi:10.1038/sj.cdd.4400950 tor domain possesses intrinsic NAD+ cleavage activity
Ashkenazi A, Salvesen G. 2014. Regulated cell death: signal- that promotes pathological axonal degeneration. Neuron
ing and mechanisms. Annu Rev Cell Dev Biol 30: 337– 93: 1334–1343.e5. doi:10.1016/j.neuron.2017.02.022
356. doi:10.1146/annurev-cellbio-100913-013226 Essuman K, Summers DW, Sasaki Y, Mao X, Yim AKY,
Balint-Kurti P. 2019. The plant hypersensitive response: DiAntonio A, Milbrandt J. 2018. TIR domain proteins
concepts, control and consequences. Mol Plant Pathol are an ancient family of NAD+-consuming enzymes.
20: 1163–1178. Curr Biol 28: 421–430.e4. doi:10.1016/j.cub.2017.12.024
Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442 13
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.This is a free sample of content from Cell Survival and Cell Death, Second Edition.
Click here for more information on how to buy the book.
K. Hofmann
Feng S, Fox D, Man SM. 2018. Mechanisms of gasdermin Lamkanfi M, Dixit VM. 2010. Manipulation of host cell
family members in inflammasome signaling and cell death pathways during microbial infections. Cell Host Mi-
death. J Mol Biol 430: 3068–3080. doi:10.1016/j.jmb crobe 8: 44–54. doi:10.1016/j.chom.2010.06.007
.2018.07.002 Lapin D, Kovacova V, Sun X, Dongus JA, Bhandari DD, von
Fuchs Y, Steller H. 2011. Programmed cell death in animal Born P, Bautor J, Guarneri N, Rzemieniewski J, Stutt-
development and disease. Cell 147: 742–758. doi:10.1016/ mann J, et al. 2019. A coevolved EDS1-SAG101-NRG1
j.cell.2011.10.033 module mediates cell death signaling by TIR-domain im-
Gonçalves AP, Heller J, Daskalov A, Videira A, Glass NL. mune receptors. Plant Cell. doi:10.1105/tpc.19.00118
2017. Regulated forms of cell death in fungi. Front Micro- Leipe DD, Koonin EV, Aravind L. 2004. STAND, a class of P-
biol 8: 1837. doi:10.3389/fmicb.2017.01837 loop NTPases including animal and plant regulators of
Green DR. 2019. The coming decade of cell death research: programmed cell death: multiple, complex domain archi-
five riddles. Cell 177: 1094–1107. doi:10.1016/j.cell.2019 tectures, unusual phyletic patterns, and evolution by hor-
.04.024 izontal gene transfer. J Mol Biol 343: 1–28. doi:10.1016/j
Hander T, Fernández-Fernández AD, Kumpf RP, Willems P, .jmb.2004.08.023
Schatowitz H, Rombaut D, Staes A, Nolf J, Pottie R, Yao P, Li J, McQuade T, Siemer AB, Napetschnig J, Moriwaki K,
et al. 2019. Damage on plants activates Ca2+-dependent Hsiao YS, Damko E, Moquin D, Walz T, McDermott A, et
metacaspases for release of immunomodulatory peptides. al. 2012. The RIP1/RIP3 necrosome forms a functional
Science 363: eaar7486. doi:10.1126/science.aar7486 amyloid signaling complex required for programmed ne-
Harris SL, Kazmierczak M, Pangršič T, Shah P, Chuchvara crosis. Cell 150: 339–350. doi:10.1016/j.cell.2012.06.019
N, Barrantes-Freer A, Moser T, Schwander M. 2017. Con- Maekawa T, Kufer TA, Schulze-Lefert P. 2011. NLR func-
ditional deletion of pejvakin in adult outer hair cells tions in plant and animal immune systems: so far and yet
causes progressive hearing loss in mice. Neuroscience so close. Nat Immunol 12: 817–826. doi:10.1038/ni.2083
344: 380–393. doi:10.1016/j.neuroscience.2016.12.055 Mahdi L, Huang M, Zhang X, Nakano RT, Kopp LB, Saur
Hauenstein AV, Zhang L, Wu H. 2015. The hierarchical IML, Jacob F, Kovacova V, Lapin D, Parker JE, et al. 2019.
structural architecture of inflammasomes, supramolecu- Plant mixed lineage kinase domain-like proteins limit
lar inflammatory machines. Curr Opin Struct Biol 31: 75– biotrophic pathogen growth. bioRxiv 681015. doi:10
83. doi:10.1016/j.sbi.2015.03.014 .1101/681015
Hofmann K. 1999. The modular nature of apoptotic signal- Man SM, Karki R, Kanneganti TD. 2017. Molecular mech-
ing proteins. Cell Mol Life Sci 55: 1113–1128. doi:10.1007/ anisms and functions of pyroptosis, inflammatory cas-
s000180050361 pases and inflammasomes in infectious diseases. Immu-
Huang W, Jiang T, Choi W, Qi S, Pang Y, Hu Q, Xu Y, Gong nol Rev 277: 61–75. doi:10.1111/imr.12534
X, Jeffrey PD, Wang J, et al. 2013. Mechanistic insights Martin SJ, Green DR. 1995. Protease activation during apo-
into CED-4-mediated activation of CED-3. Genes Dev 27: ptosis: death by a thousand cuts? Cell 82: 349–352. doi:10
2039–2048. doi:10.1101/gad.224428.113 .1016/0092-8674(95)90422-0
Jiang S, Gu H, Zhao Y, Sun L. 2019. Teleost gasdermin E is Mompeán M, Li W, Li J, Laage S, Siemer AB, Bozkurt G, Wu
cleaved by caspase 1, 3, and 7 and induces pyroptosis. H, McDermott AE. 2018. The structure of the necrosome
J Immunol 203: 1369–1382. doi:10.4049/jimmunol.190 RIPK1-RIPK3 core, a human hetero-amyloid signaling
0383 complex. Cell 173: 1244–1253.e10. doi:10.1016/j.cell
Jones JD, Vance RE, Dangl JL. 2016. Intracellular innate .2018.03.032
immune surveillance devices in plants and animals. Sci- Morel JB, Dangl JL. 1997. The hypersensitive response and
ence 354: aaf6395. doi:10.1126/science.354.6316.1174-b the induction of cell death in plants. Cell Death Differ 4:
Kajava AV, Klopffleisch K, Chen S, Hofmann K. 2014. Evo- 671–683. doi:10.1038/sj.cdd.4400309
lutionary link between metazoan RHIM motif and prion- Nagata S. 2018. Apoptosis and clearance of apoptotic cells.
forming domain of fungal heterokaryon incompatibility Annu Rev Immunol 36: 489–517. doi:10.1146/annurev-
factor HET-s/HET-s. Sci Rep 4: 7436. doi:10.1038/srep immunol-042617-053010
07436 Nanson JD, Kobe B, Ve T. 2019. Death, TIR, and RHIM: self-
Kambara H, Liu F, Zhang X, Liu P, Bajrami B, Teng Y, Zhao assembling domains involved in innate immunity and
L, Zhou S, Yu H, Zhou W, et al. 2018. Gasdermin D exerts cell-death signaling. J Leukoc Biol 105: 363–375. doi:10
anti-inflammatory effects by promoting neutrophil death. .1002/JLB.MR0318-123R
Cell Rep 22: 2924–2936. doi:10.1016/j.celrep.2018.02.067 Newton K, Manning G. 2016. Necroptosis and inflamma-
Kersse K, Verspurten J, Vanden Berghe T, Vandenabeele P. tion. Annu Rev Biochem 85: 743–763. doi:10.1146/an
2011. The death-fold superfamily of homotypic interac- nurev-biochem-060815-014830
tion motifs. Trends Biochem Sci 36: 541–552. doi:10.1016/ Orning P, Weng D, Starheim K, Ratner D, Best Z, Lee B,
j.tibs.2011.06.006 Brooks A, Xia S, Wu H, Kelliher MA, et al. 2018. Pathogen
Koonin EV, Aravind L. 2002. Origin and evolution of eu- blockade of TAK1 triggers caspase-8-dependent cleavage
karyotic apoptosis: the bacterial connection. Cell Death of gasdermin D and cell death. Science 362: 1064–1069.
Differ 9: 394–404. doi:10.1038/sj.cdd.4400991 doi:10.1126/science.aau2818
Kovacs SB, Miao EA. 2017. Gasdermins: effectors of pyrop- Paoletti M, Clavé C. 2007. The fungus-specific HET domain
tosis. Trends Cell Biol 27: 673–684. doi:10.1016/j.tcb.2017 mediates programmed cell death in Podospora anserina.
.05.005 Eukaryot Cell 6: 2001–2008. doi:10.1128/EC.00129-07
14 Cite this article as Cold Spring Harb Perspect Biol doi: 10.1101/cshperspect.a036442
© 2020 by Cold Spring Harbor Laboratory Press. All rights reserved.You can also read

















































