The Serum- and Glucocorticoid-Inducible Kinase Sgk-1 Is Involved in Pulmonary Vascular Remodeling
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The Serum- and Glucocorticoid-Inducible Kinase Sgk-1 Is
Involved in Pulmonary Vascular Remodeling
Role in Redox-Sensitive Regulation of Tissue Factor by Thrombin
Rachida S. BelAiba, Talija Djordjevic, Steve Bonello, Ferruh Artunc, Florian Lang,
John Hess, Agnes Görlach
Abstract—The stress-responsive serum- and glucocorticoid-inducible kinase Sgk-1 is involved in osmoregulation and cell
survival and may contribute to fibrosis and hypertension. However, the function of Sgk-1 in vascular remodeling and
thrombosis, 2 major determinants of pulmonary hypertension (PH), has not been elucidated. We investigated the role
of Sgk-1 in thrombin signaling and tissue factor (TF) expression and activity in pulmonary artery smooth muscle cells
(PASMC). Thrombin increased Sgk-1 activity and mRNA and protein expression. H2O2 similarly induced Sgk-1
expression. Antioxidants, dominant-negative Rac, and depletion of the NADPH oxidase subunit p22phox diminished
thrombin-induced Sgk-1 expression. Inhibition of p38 mitogen-activated protein kinase, phosphatidylinositol 3-kinase,
and phosphoinositide-dependent kinase-1 prevented thrombin-induced Sgk-1 expression. Thrombin or Sgk-1 overex-
pression enhanced TF expression and procoagulant activity, whereas TF upregulation by thrombin was diminished by
kinase-deficient Sgk-1 and was not detectable in fibroblasts from mice deficient in sgk-1 (sgk1⫺/⫺). Similarly,
dexamethasone treatment failed to induce TF expression and activity in lung tissue from sgk1⫺/⫺ mice. Transcriptional
induction of TF by Sgk-1 was mediated through nuclear factor B. Finally, Sgk-1 and TF proteins were detected in the
media of remodeled pulmonary vessels associated with PH. These data show that thrombin potently induces Sgk-1
involving NADPH oxidases, phosphatidylinositol 3-kinase, p38 mitogen-activated protein kinase, and phosphoinosi-
tide-dependent kinase-1, and that activation of nuclear factor B by Sgk-1 mediates TF expression and activity by
thrombin. Because enhanced procoagulant activity can promote pulmonary vascular remodeling, and Sgk-1 and TF were
present in the media of remodeled pulmonary vessels, this pathway may play a critical role in vascular remodeling in PH.
(Circ Res. 2006;98:828-836.)
Key Words: sgk-1 䡲 thrombin 䡲 tissue factor 䡲 NADPH oxidase 䡲 coagulant activity
䡲 pulmonary vascular remodeling 䡲 pulmonary artery smooth muscle cells 䡲 nuclear factor B
P ulmonary hypertension (PH) is a complex progressive
disorder characterized by remodeling of the lung vascu-
lature with hypertrophy of the media.1 Moreover, a prothrom-
ing vascular cells by involving mitogen-activated protein
kinases (MAPKs), the phosphatidylinositol 3-kinase (PI3K)
pathway, and reactive oxygen species (ROS).4,5
botic state is frequently associated with PH, which may be The serum- and glucocorticoid-inducible kinase-1, Sgk-1,
propagated by enhanced thrombogenicity of the injured is a known downstream effector of the PI3K cascade. Sgk-1
pulmonary vascular wall, increased thrombin activity, and an belongs to the “AGC” family of serine-threonine kinases and
imbalance between prothrombotic and antithrombotic factors. shares approximately 45% to 55% homology with Akt in its
The observation that antithrombotic treatment can improve catalytic domain.6 In contrast to Akt, Sgk-1 is also regulated
the survival rate in patients with PH points toward the at the transcriptional level in response to various hormones,
importance of thrombosis in the pathogenesis of this growth factors, and extracellular stresses in a cell type–
disorder.1,2 dependent manner, allowing Sgk-1 to be available for its
Thrombin is generated by a series of proteolytic events that targets only when needed.7,8
are initiated when tissue factor (TF) binds to plasma factor Sgk-1 was originally cloned from murine mammary tumor
VIIa.3 Interestingly, thrombin has been described to regulate cells as a glucocorticoid-responsive gene.6 Human Sgk-1 was
its own generation by promoting TF transcription and TF- subsequently cloned as a cell volume-sensitive gene upregu-
dependent procoagulant activity in several cell types includ- lated by hypertonic cell shrinkage.9 Increasing evidence
Original received March 3, 2005; resubmission received October 25, 2005; revised resubmission received January 4, 2006; accepted February 3, 2006.
From the Experimental Pediatric Cardiology (R.S.B., T.D., S.B., J.H., A.G.), Department of Pediatric Cardiology and Congenital Heart Disease,
German Heart Center Munich at the Technical University Munich; and Institute for Physiology I (F.A., F.L.), University of Tuebingen, Germany.
Correspondence to Agnes Görlach, MD, Experimental Pediatric Cardiology, Department of Pediatric Cardiology and Congenital Heart Disease,
German Heart Center Munich at the Technical University Munich, Lazarettstr. 36, D-80636 München, Germany. E-mail goerlach@dhm.mhn.de
© 2006 American Heart Association, Inc.
Circulation Research is available at http://circres.ahajournals.org DOI: 10.1161/01.RES.0000210539.54861.27
828
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015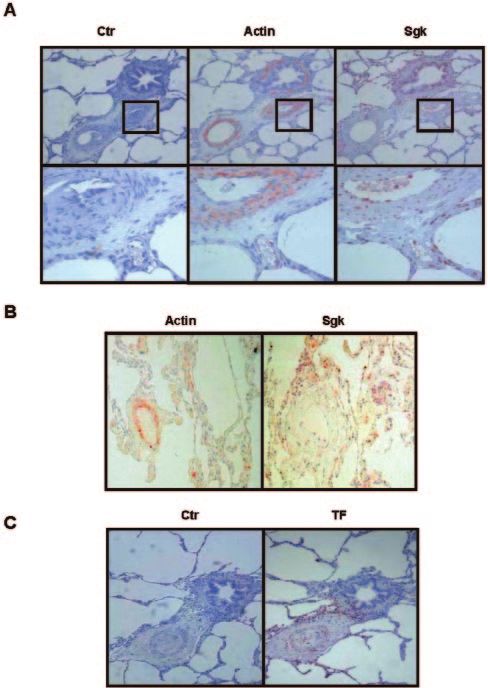
BelAiba et al Sgk-1 Regulates Tissue Factor 829
suggests that expression, enzymatic activity, and cellular Procoagulant Activity Assay
localization of Sgk-1 are regulated in response to various The surface procoagulant activity was evaluated in PASMC trans-
stimuli controlling not only cell volume and epithelial trans- fected with Sgk-1-WT and Sgk-1-DK expression vectors or in
extracted lung protein as described.19,20
port, but also cardiac action potential and cell proliferation,
survival, and apoptosis.7,8 Excessive transcription of Sgk-1 Immunoprecipitation and Sgk-1 Activity Assay
has been shown to parallel diabetic nephropathy,10 glomeru- PASMC were transfected with an Sgk-1 expression vector and
lonephritis,11 hepatic cirrhosis,12 pulmonary fibrosis,13 and stimulated for different time points with 3 U/mL thrombin. Immu-
polymorphisms of the Sgk-1 gene correlated with hyperten- noprecipitation was performed using a GFP antibody as described.21
sion.14 Despite the wide tissue distribution of Sgk-1 and its Sgk-1 activity assay (Stressgen) was performed according to the
instructions of the manufacturer.
sensitivity to various stimuli, the role of Sgk-1 in the
cardiovascular and pulmonary system remained ill defined. Immunofluorescence
Because heparin, an inhibitor of thrombin formation, has Immunofluorescence was performed as described.22 The p50 anti-
been shown to decrease Sgk-1 mRNA in aortic smooth body was from Santa Cruz Biotechnology, the GFP antibody and the
muscle cells,15 we hypothesized that Sgk-1 may play a role in secondary antibodies coupled to Alexa 488 or 594 were
from Mobitech.
thrombin signaling in human pulmonary artery smooth mus-
cle cells (PASMC), the main cell type involved in PH. We Immunohistochemistry
found that Sgk-1 is activated and induced by thrombin, that it Archival lung tissue was obtained from patients with secondary PH
regulates TF expression and activity in PASMC, and that it is with and without signs of pulmonary vascular remodeling or from
present in remodeled pulmonary vessels with media hyper- lung tumor resections lacking apparent pulmonary vascular remod-
eling. Tissue was formalin fixed under vacuum and paraffin embed-
trophy associated with PH.
ded. Immunohistochemistry was performed using the LSAB2 HRP
Systems (Dako) and antibodies against Sgk-1 (Upstate), TF (Amer-
Materials and Methods ican Diagnostica), and actin (Dako). Counterstaining was performed
using Hemalum.
Reagents
Deoxycytidine 5⬘-␣32P-triphosphate (3000 Ci/mmoL) was from Am- Northern Blot Analysis
ersham. Human ␣-thrombin was from Hemochrom Diagnostika. All Total RNA was isolated as described.17 RNA (10 g) was subjected
other chemicals were from Sigma. to Northern blot analysis, and hybridizations were performed with
32
P-labeled cDNA fragments for human Sgk-1 (1338 bp), human TF
Animal Experimentation (1820 bp),17,19 or 18S.
Mice deficient in Sgk-1 (sgk1⫺/⫺) were generated as described.16
Wild-type (sgk1⫹/⫹) and sgk1⫺/⫺ (6 weeks old) mice were treated Western Blot Analysis
with dexamethasone (10 g/g body weight) for 4 days or vehicle Western blot analysis was performed as described.17 The antibody
(daily SC injection) and euthanized. Lungs were dissected and snap against human TF (3D1) was provided by Dr T. Zioncheck (Genen-
frozen in liquid nitrogen. tech). The antibody against mouse TF was from American Diagnos-
tica. The antibody against pSer422-Sgk-1 was provided by Sir P.
Cohen (Dundee University). The antibody against pThr256-Sgk-1
Cell Culture was from Santa Cruz Biotechnology. The antibody against p22phox
Human PASMC were from Cambrex and cultured in the medium was provided by Dr D. Roos (University of Amsterdam, Sanquin
provided as recommended. PASMC were serum deprived for 24 Research, and Landsteiner Laboratory, Academic Medical Center,
hours before stimulation with thrombin (3 U/mL).17 Because Amsterdam, The Netherlands). The antibodies against actin or the
PASMC do not efficiently express luciferase constructs, A7r5 rat c-myc epitope were from Sigma.
smooth muscle cells (rSMC) were used for reporter gene assays, as
described.17 Fibroblasts harvested from the tail of sgk1⫹/⫹ and Statistical Analysis
sgk1⫺/⫺ mice were identified by positive staining for fibronectin and Values are presented as means⫾SD. Results were compared by
used in experiments as described.16 ANOVA for repeated measurements followed by Student–Newman–
Keuls t test. P⬍0.05 was considered statistically significant.
Plasmids and Transfections
Expression vectors encoding wild-type Sgk-1 (Sgk-1-WT) or kinase- Results
deficient Sgk-1 (Sgk-1-DK) fused to green fluorescent protein (GFP)
have been described.12 Expression vectors encoding mitogen-acti- Thrombin Stimulates Expression and Activity
vated protein kinase kinase 3 (MKK3), kinase-deficient MKK3 of Sgk-1
(MKK3KD), myc-tagged phosphoinositide-dependent kinase-1 To investigate whether Sgk-1 is regulated by thrombin,
(PDK1), kinase-deficient (K111A/D223A) PDK1 (PDKKD), myc- PASMC were stimulated with thrombin and Sgk-1 mRNA
tagged RacT17N or RacG12V, and the TF promoter constructs and protein levels were determined. Sgk-1 mRNA levels were
pTF636 and pTF111 have been described.17,18 PCR-directed mu-
tagenesis was performed on pTF636 to generate pTF636AEm with
peaking after 0.5 hour of exposure (Figure 1A and 1B) and
mutations in all AP-1 and EGR-1 sites and on pTF636NFm with a Sgk-1 protein levels peaked after 2 hour stimulation (Figure
mutated nuclear factor B (NFB) site. NFB-Luc was from 1A and 1B). Pretreatment with cycloheximide or actinomycin
Clontech and I␣B-dn was from Becton Dickinson. The siSTRIKE D decreased Sgk-1 protein induction indicating that thrombin
U6 Hairpin Cloning System (Promega) was used to create vectors regulates Sgk-1 at the transcriptional level and via de novo
encoding for small interference RNA (siRNA) targeting p22phox or
synthesis (Figure 1C). To evaluate whether thrombin is also
for unspecific, nonsilencing siRNA. Transfection of PASMC or
rSMC and luciferase assays were performed as described.17 The able to activate Sgk-1, PASMC were transfected with a vector
transfection efficiency was controlled by fluorescence microscopy encoding Sgk-1 and stimulated with thrombin for increasing
and Western blot analysis revealing on average 40%. time periods. Sgk-1 was immunoprecipitated and Western
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015
830 Circulation Research March 31, 2006
blot analyses were performed to detect phosphorylation of
Sgk-1 at serine 422 and threonine 256, which are required for
full Sgk-1 activity. In both cases, thrombin rapidly stimulated
the phosphorylation of Sgk-1 after only 1 minute of exposure
(Figure 1D). Similarly, thrombin enhanced Sgk-1 enzyme
activity within 1 minute (data not shown).
ROS Are Involved in Thrombin-Stimulated
Sgk-1 Expression
ROS have been described as important signaling molecules in
the response to thrombin.5 To determine whether Sgk-1
expression is mediated by ROS, PASMC were exposed to
H2O2. Sgk-1 protein levels were maximally increased at a
dose of 50 mol/L (Figure 2A) and an exposure time of 2
hours (Figure 2B). Pretreatment with the antioxidants vitamin
C and N-acetyl cysteine (NAC) or the nonspecific NADPH
oxidase inhibitor diphenyleneiodonium (DPI) prevented in-
duction of Sgk-1 by thrombin (Figure 2C). To further
delineate the role of NADPH oxidases in this response,
constitutively active (RacG12V) or dominant-negative
(RacT17N) mutants of Rac, which is required for ROS
production by NADPH oxidases, were expressed in PASMC.
Active Rac further enhanced thrombin-stimulated Sgk-1 pro-
tein, whereas dominant-negative Rac inhibited Sgk-1 upregu-
lation by thrombin (Figure 2D). Furthermore, transfection of
siRNA targeting the NADPH oxidase subunit p22phox down-
regulated thrombin-stimulated Sgk-1 protein levels (Figure
2E).
Thrombin-Stimulated Sgk-1 Expression Involves
p38 MAPK, PI3K, and PDK1
Because ROS and NADPH oxidases are known to activate
MAPKs5,19,23 we determined their involvement in the regu-
lation of Sgk-1. Pretreatment of PASMC with the p38 MAPK
(p38MAPK) inhibitor SB220025 (5 to 20 mol/L) dimin-
ished thrombin-induced Sgk-1 (data not shown) with a
maximal effect at 10 mol/L (Figure 3A), a concentration
that blocked p38MAPK phosphorylation by thrombin (data
not shown). In addition, overexpression of MKK3, an up-
stream kinase of p38MAPK, enhanced Sgk-1 protein levels,
whereas kinase-deficient MKK3 decreased thrombin-induced
Sgk-1 protein (Figure 3B). In contrast, application of the
c-jun N-terminal kinase (JNK) inhibitor SP600125 (5 to
Figure 1. Thrombin induces Sgk-1 expression and activity.
PASMC were incubated for different time periods with throm- 20 mol/L) did not significantly inhibit thrombin-induced
bin (3 U/mL). A, Sgk-1 mRNA levels were determined by Sgk-1 protein (Figure 3A). In addition, exposure to different
Northern blot using a Sgk-1 probe, or Sgk-1 protein levels doses of the PI3K inhibitor LY294002 (10 to 50 mol/L)
were determined by Western blot analysis using a human
Sgk-1 antibody (n⫽3; *P⬍0.05 vs Ctr). B, Representative
abolished Sgk-1 expression at a dose of 20 mol/L (Figure
Northern and Western blots. C, PASMC were pretreated with 3A). Because the PI3K downstream target PDK1 has been
cycloheximide (CH) (100 mol/L) or actinomycin D (AcD) shown to phosphorylate Sgk-1,24 we explored whether PDK1
(5 mol/L) for 30 minutes and stimulated with thrombin for 2 is also involved in the control of Sgk-1 expression by
hours. Sgk-1 protein levels were determined by Western blot
(n⫽3; *P⬍0.05 vs Ctr, #P⬍0.05 vs thrombin-stimulated Ctr). thrombin. Thrombin rapidly phosphorylated PDK1, peaking
D, PASMC were transfected with an expression vector at 2.5 minutes (data not shown). Overexpression of PDK1
encoding for Sgk-1 (Sgk-WT) and stimulated with thrombin increased Sgk-1 protein levels, whereas expression of kinase-
for different time periods. Immunoprecipitation was performed
using a GFP antibody. Western blot analyses were performed deficient PDK1 blunted the effect of thrombin on Sgk-1
with antibodies against p-ser422 or p-thr256 Sgk-1. Blots are expression (Figure 3C). Furthermore, kinase-deficient PDK1
representative of 3 experiments. prevented induction of Sgk-1 by MKK3, suggesting that
p38MAPK acts upstream of PDK1 in regulating Sgk-1 levels
(Figure 3D).
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015BelAiba et al Sgk-1 Regulates Tissue Factor 831
Figure 2. Sgk-1 regulation is redox sen-
sitive and involves Rac and p22phox.
PASMC were incubated with different
doses of H2O2 (A) or for different time
periods with 50 mol/L H2O2 (B). Sgk-1
protein levels were determined by West-
ern blot (n⫽3; *P⬍0.05 vs Ctr,). C,
PASMC were pretreated with vitamin C
(VitC) (100 mol/L), DPI (10 mol/L), or
NAC (5 mmol/L) and stimulated with
thrombin. Sgk-1 protein levels were
determined by Western blot analysis
(n⫽3; *P⬍0.05 vs Ctr, #P⬍0.05 vs
thrombin-stimulated Ctr). D, PASMC
were transfected with control vector (Ctr)
or expression vectors encoding for active
RacG12V (V12) or dominant-negative
RacT17N (N17). Sgk-1 protein levels
were determined by Western blot.
Expression of the Rac constructs was
controlled using an antibody against the
c-myc epitope (n⫽3; *P⬍0.05 vs Ctr,
#P⬍0.05 vs thrombin-stimulated Ctr). E,
PASMC were transfected with p22phox
siRNA (sip22) or nonsilencing siRNA
(siNo). Sgk-1 and p22phox protein levels
were determined by Western blot (n⫽3;
*P⬍0.05 vs siNo, #P⬍0.05 vs thrombin-
stimulated siNo).
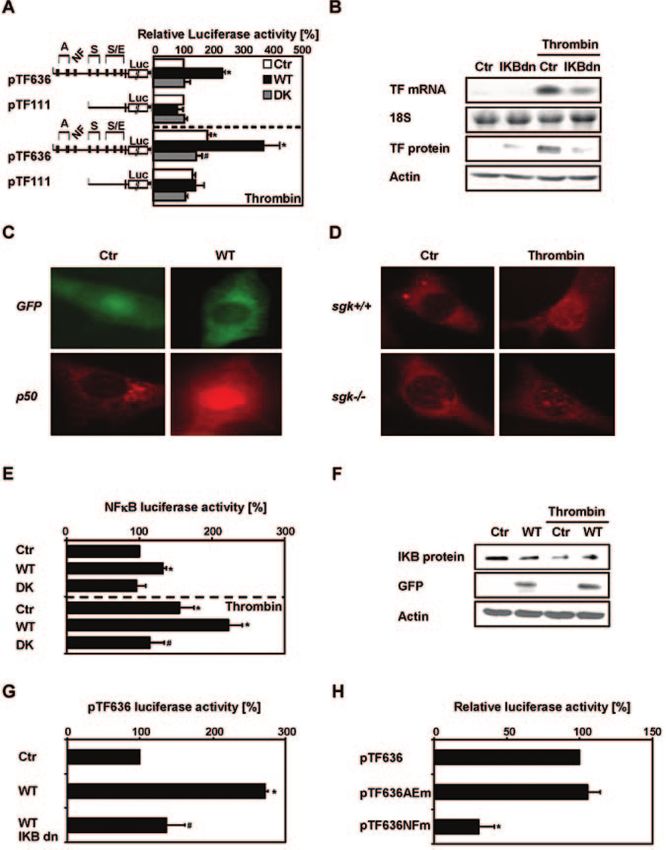
Sgk-1 Is Involved in Tissue Factor Expression and moter: pTF111 contains the promoter sequence ⫺111 to
Activity by Thrombin ⫹121 bp including a proximal enhancer with EGR-1 and Sp1
Because thrombin has been shown to upregulate TF mRNA consensus sites. pTF636 contains in addition a distal enhancer
and protein in PASMC,17 we investigated the role of Sgk-1 in containing an NFB and 2 AP-1 sites (Figure 5A).
thrombin-stimulated TF expression and activity. Sgk-1 over- Thrombin and Sgk-1-WT significantly increased luciferase
expression enhanced TF mRNA and protein levels and TF activity of pTF636, but not of pTF111, whereas in the
activity. In contrast, kinase-deficient Sgk-1 prevented presence of Sgk-1-DK thrombin-stimulated pTF636-driven
thrombin-induced TF (Figure 4A through 4C). Furthermore, luciferase activity was diminished (Figure 5A). Because
TF protein was not detected in fibroblasts from Sgk-1 pTF636 harbors a NFB site, which is not present in pTF111,
knockout (sgk1⫺/⫺) mice stimulated with thrombin for 8 hours we investigated whether Sgk-1 could regulate TF expression
but was upregulated in sgk1⫹/⫹ cells (Figure 4D). To assess via NFB. Overexpression of dominant-negative I␣B (IB-
the involvement of Sgk-1 in vivo, TF expression and activity dn), which prevents NFB activation, diminished thrombin-
were determined in lungs from sgk1⫺/⫺ or sgk1⫹/⫹ mice. stimulated TF mRNA and protein levels (Figure 5B). Expres-
Because thrombin stimulation induces multiple side effects sion of Sgk-1-WT, but not of control vector, resulted in the
and is thus not advisable for in vivo treatment, mice were translocation of the NFB subunit p50 to the nucleus (Figure
treated with dexamethasone, known to induce Sgk-1 and 5C). Thrombin induced the translocation of p50 in sgk1⫹/⫹,
TF.7,25 TF protein levels were enhanced in lung tissue derived but not in sgk1⫺/⫺ fibroblasts (Figure 5D). Consistently,
from dexamethasone-treated sgk-1⫹/⫹ but not from sgk1⫺/⫺ NFB-dependent luciferase activity was enhanced in the
mice (Figure 4E). Similarly, TF activity in lung extracts presence of Sgk-1-WT or thrombin, whereas thrombin-
derived from sgk1⫹/⫹, but not from sgk1⫺/⫺ mice, was stimulated NFB-dependent luciferase activity was reduced
significantly enhanced by dexamethasone (Figure 4F). by Sgk-1-DK (Figure 5E). Sgk-1 overexpression and throm-
bin promoted IB degradation (Figure 5F), whereas IB-dn
NFB Mediates Sgk-1–Dependent Tissue significantly diminished Sgk-1–induced pTF636-driven lucif-
Factor Expression erase activity (Figure 5G). To confirm that Sgk-1– dependent
To investigate the role of Sgk-1 in the transcriptional regu- expression of TF is mediated by NFB, Sgk-1-WT was
lation of TF by thrombin, reporter gene assays were per- coexpressed with either pTF636AEm, where the AP-1 and
formed in the presence of Sgk-1-WT or Sgk-1-DK using the EGR-1 sites were mutated, or pTF636NFm, where the
luciferase constructs containing 2 fragments of the TF pro- NFB element was mutated. No difference in luciferase
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015832 Circulation Research March 31, 2006
activity was observed between pTF636 and pTF636AEm,
whereas pTF636NFm-driven luciferase activity was signifi-
cantly diminished (Figure 5H).
Sgk-1 Is Expressed in Remodeled
Pulmonary Vessels
Finally, we assessed whether Sgk-1 is also associated with
pulmonary vascular remodeling. To this end, lung tissue was
obtained from patients with PH (n⫽5) or lung carcinoma
(n⫽1) and evaluated for media hypertrophy as sign of
pulmonary vascular disease. Immunohistochemistry revealed
that Sgk-1 was expressed in the bronchial epithelium (Figure
6A), as described.26 In addition, in all tissue samples with
apparent vascular disease (n⫽3), strong expression of Sgk-1
was found in the media of small vessels (Figure 6A). In
contrast, in tissue samples lacking vascular remodeling
(n⫽3), Sgk-1 protein was almost undetectable in the smooth
muscle cell layer (Figure 6B). Similarly, Sgk-1 could not be
detected in airway smooth muscle (Figure 6A), suggesting
that Sgk-1 is specifically expressed in remodeled pulmonary
vessels. In addition, TF protein colocalized with Sgk-1 in the
media of small vessels (Figure 6C), further emphasizing a
role of Sgk-1 in pulmonary vascular remodeling.
Discussion
In this study, we identified Sgk-1 as a novel element in the
regulation of TF expression and activity by thrombin which
required activation of NFB, whereby thrombin induced
Sgk-1 by stimulating NADPH oxidases, PI3K, p38MAPK,
and PDK1. This was based on the following findings. (1)
Thrombin stimulated phosphorylation as well as de novo
synthesis of Sgk-1. (2) Sgk-1 induction by thrombin was
redox sensitive and diminished by dominant-negative Rac
and depletion of p22phox. (3) Inhibition of PI3K, MKK3/
p38MAPK, and PDK1 prevented Sgk-1 induction by throm-
bin. (4) Sgk-1 overexpression increased TF expression and
activity, whereas TF expression and activity were diminished
in PASMC expressing kinase-deficient Sgk-1, in cultivated
sgk1⫺/⫺ cells or in lungs derived from sgk1⫺/⫺ mice treated
with dexamethasone. (5) Sgk-1 and thrombin enhanced deg-
radation of IB and promoted NFB nuclear translocation
and activity. (6) Sgk-1–induced TF promoter activity was
diminished by dominant-negative I␣B or a mutation in its
NFB binding site. Because Sgk-1 and TF proteins were
Figure 3. Thrombin-stimulated Sgk-1 expression is dependent detected in the media of vessels with pulmonary vascular
on p38MAPK, PI3K, and PDK1. A, PASMC were pretreated with disease these pathways may be involved in pulmonary vas-
LY294002 (LY) (10 mol/L), SP600125 (SP) (20 mol/L), or
SB220025 (SB, 20 mol/L) for 30 minutes and stimulated with cular remodeling in PH.
thrombin for 2 hours. Sgk-1 protein levels were determined by
Western blot (n⫽3; *P⬍0.05 vs Ctr, #P⬍0.05 vs thrombin- Thrombin Stimulates Sgk-1 Activity
stimulated Ctr). B, Sgk-1 protein levels were determined in and Expression
PASMC expressing MKK3 (MKK) or kinase-deficient MKK3
(MKKKD) (n⫽3; *P⬍0.05 vs Ctr, #P⬍0.05 vs thrombin- Pulmonary vascular remodeling is a common complication of
stimulated Ctr). Expression of the constructs was controlled PH and is associated with in situ thrombosis and increased
using an antibody against the FLAG epitope. C, Sgk-1 protein procoagulant activity.1,2 The coagulation factors thrombin
levels were determined in PASMC expressing wild-type PDK1
(PDK), or kinase-deficient PDK1 (PDKKD) (n⫽3; *P⬍0.05 vs Ctr,
and TF have been shown to interact with the vascular wall
#P⬍0.05 vs thrombin-stimulated Ctr). Expression of the con- and to activate signaling pathways and gene expression thus
structs was controlled using an antibody against the c-myc being key players in promoting vascular remodeling
epitope. D, PASMC were transfected with vectors encoding for processes.27
MKK3 (MKK), kinase-deficient PDK (PDKKD), or control vector
(pcDNA3). Sgk-1 levels were determined by Western blot analy- In this study, we identified Sgk-1 as a target for thrombin.
sis. Blots are representative of 3 independent experiments. In PASMC, thrombin rapidly phosphorylated overexpressed
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015BelAiba et al Sgk-1 Regulates Tissue Factor 833
Figure 4. Sgk-1 induces TF expression
and procoagulant activity. PASMC were
transfected with control vector (Ctr) or
expression vectors for Sgk-1 (WT) or
kinase-deficient Sgk-1 (DK) and stimu-
lated with thrombin. A, TF mRNA levels
were determined by Northern blot analy-
sis using a human TF cDNA probe. B, TF
protein levels were determined by West-
ern blot analysis using an antibody
against human TF. Expression of the
constructs was controlled using a GFP
antibody. C, TF activity was evaluated
using a chromogenic assay. Values⫾SD
represent the percentage of induction of
TF mRNA, protein, or procoagulant activ-
ity, respectively (n⫽3; *P⬍0.05 vs Ctr,
#P⬍0.05 vs thrombin-stimulated Ctr). D,
Fibroblasts from control (sgk1⫹/⫹) or
sgk-1 knock-out (sgk1⫺/⫺) mice were
stimulated with thrombin for 8 hour.
Western blot analysis was performed
using antibodies against TF or Sgk-1.
Blots are representative of 3 experi-
ments. E, sgk1⫹/⫹ or sgk1⫺/⫺ mice were
treated with dexamethasone (10 g/g
body weight) for 4 days or with vehicle
(daily SC injection). TF protein levels
were determined in lung extracts by
Western blot using a TF antibody (n⫽3;
*P⬍0.05 vs vehicle [sgk1⫹/⫹]). F, TF
activity was determined in lung extracts
from sgk1⫹/⫹ or sgk1⫺/⫺ mice treated
with dexamethasone (n⫽3; *P⬍0.05 vs
vehicle [sgk1⫹/⫹]).
as well as endogenous (data not shown) Sgk-1 at serine 422 p38MAPK was involved in Sgk-1 upregulation by hyperos-
and threonine 256, which is required for activation of Sgk- motic stress and transforming growth factor- in different
1.7,24 In addition, thrombin transiently enhanced Sgk-1 cell types7,8,10,12,13,30 and by interleukin 6 in cholangiocarci-
mRNA and protein levels, consistent with previous findings noma.31 Because p38MAPK can be activated by thrombin
that Sgk-1 is regulated at the level of activity and involving ROS and NADPH oxidases,19,23 stimulation of
expression.7,8 NADPH oxidases by thrombin may activate p38MAPK and
subsequently induce Sgk-1. Although inhibition of ERK1/2
NADPH Oxidases and ROS Mediate Sgk-1 also reduced Sgk-1 consistent with previous findings,32 rela-
Upregulation by Thrombin tively high doses of PD98059 were required to downregulate
Our results further indicate that ROS are critically involved in Sgk-1 (data not shown). Together with the observation that
the upregulation of Sgk-1 because H2O2 time- and dose- JNK inhibition did not affect Sgk-1 induction by thrombin,
dependently enhanced Sgk-1 levels similar to previous ob- these data further emphasize the role of p38MAPK in
servations in murine mammary glandular cells.28 Consis- regulating Sgk-1 expression by thrombin.
tently, the antioxidants vitamin C and NAC as well as the In addition, inhibition of PI3K blunted Sgk-1 upregulation
unspecific NADPH oxidase inhibitor DPI diminished by thrombin, consistent with previous observations.21,30 Fur-
thrombin-induced upregulation of Sgk-1 expression in thermore, PDK1, a downstream kinase of PI3K, which can
PASMC. In addition, depletion of the NADPH oxidase phosphorylate Sgk-1,21 was also involved in thrombin induc-
subunit p22phox or dominant-negative Rac, known to inhibit tion of Sgk-1. Thus, in addition to PI3K and p38MAPK,
thrombin-induced ROS formation,17,19,22 prevented upregula- which have been shown to both activate and induce Sgk-
tion of Sgk-1 by thrombin, indicating that NADPH oxidases 1,7,8,21,24,31,33 our data indicate that PDK1 is able to activate
are the source of ROS promoting induction of Sgk-1. Simi- and upregulate Sgk-1. Because activation of PI3K and PDK1
larly, RacG12V activated Sgk-1 in MDCK cells, although was shown to be redox sensitive,33,34 MKK3-stimulated
RacT17N failed to prevent activation of Sgk-1 in response to Sgk-1 expression was inhibited by kinase-deficient PDK1 in
integrins.29 PASMC, and LY294002 inhibited phosphorylation of PDK1,
but not of p38MAPK, in PASMC (data not shown), our
Thrombin-Induced Sgk-1 Is Controlled by PI3K, results suggest a model whereby thrombin induces Sgk-1 via
PDK1, and p38MAPK stimulation of NADPH oxidases and activation of PI3K or
Our studies further showed that MKK3 and p38MAPK MKK3/p38MAPK followed by PDK1 activation (Figure 7).
contribute to upregulation of Sgk-1 by thrombin. Similarly, This model is further supported by our previous observations
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015834 Circulation Research March 31, 2006
Figure 5. Sgk-1 induces TF via NFB. A, rSMC were cotransfected with expression vectors encoding wild-type (WT) or kinase-deficient (DK)
Sgk-1 or control vector (Ctr) and the human TF promoter luciferase constructs pTF636 or pTF111 (A indicates AP-1; S, Sp-1; NF, NFB; E,
EGR-1) and stimulated with thrombin (n⫽3; *P⬍0.05 vs Ctr, #P⬍0.05 vs thrombin-stimulated Ctr). B, PASMC were transfected with a vector
encoding dominant-negative I␣B (IBdn) or control vector (Ctr) and stimulated with thrombin. TF mRNA and protein levels were determined
by Northern and Western blot analysis, respectively. Blots are representative for 3 experiments. C, PASMC were transfected with control vec-
tor (Ctr) or expression vectors for Sgk-1 (WT). Immunofluorescence was performed with antibodies against the NFB subunit p50 and GFP.
D, Fibroblasts from sgk1⫹/⫹ or sgk1⫺/⫺ mice were stimulated with thrombin. Immunofluorescence was performed using a p50 antibody. Each
image is representative for 3 experiments. E, rSMC were cotransfected with a luciferase construct containing five NFB elements in front of
the SV40 promoter and expression vectors encoding for Sgk-1 (WT), kinase-deficient Sgk-1 (DK), or with control vector (Ctr) and stimulated
with thrombin. Luciferase activity was measured (n⫽3; *P⬍0.05 vs Ctr, #P⬍0.05 vs thrombin-stimulated Ctr). F, PASMC were transfected
with an expression vector for Sgk-1 (WT) or control vector (Ctr) and stimulated with thrombin for 2 hours. IB levels were investigated by
Western blot. Expression of the constructs was controlled using a GFP antibody. Blots are representative for 3 experiments. G, rSMC were
cotransfected with vectors for Sgk-1 (WT), dominant-negative I␣B (IBdn), or control vector (Ctr) and pTF636 (n⫽3; *P⬍0.05 vs Ctr,
#P⬍0.05 vs Sgk-1– overexpressing cells). H, rSMC were cotransfected with wild-type Sgk-1 (WT) and pTF636 or the corresponding con-
structs pTF636AEm, where all AP-1 and EGR-1 sites were mutated, or pTF636NFm, where the NFB element was mutated (n⫽3; *P⬍0.05
vs Sgk-1-WT transfected with pTF636).
that inhibition of MKK3 prevents thrombin-stimulated PDK1 by thrombin in PASMC. However, further studies, which are
phosphorylation.18 beyond the scope of this study, are required to dissect the
Although to date only limited data exist with regard to the transcription factors involved in Sgk-1 regulation by
transcriptional machinery regulating Sgk-1,7 it has been thrombin.
suggested that p38MAPK either directly or indirectly acti-
vates a Sp1 transcription factor protein complex on the Sgk-1 Tissue Factor Expression Is Mediated by Sgk-1
promoter.30 Because Sp1 is redox sensitive and can be and Involves NF-B
activated by thrombin35 and PDK1,36 one might thus specu- We further showed that Sgk-1 is involved in the regulation of
late that such a pathway also contributes to Sgk-1 regulation TF expression and activity by thrombin in PASMC. More-
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015BelAiba et al Sgk-1 Regulates Tissue Factor 835
Figure 7. Proposed model of regulation of Sgk-1 and TF by
thrombin. Thrombin stimulates ROS production by NADPH oxi-
dases, leading via activation of PI3K, p38MAPK, and PDK1 to
the induction of Sgk-1, resulting in activation of NFB and
increased TF expression and activity, which may lead to the for-
mation of thrombin (dashed line) and thus a thrombogenic
cycle.
thrombin stimulation17 and contributes to TF expression.
Recently it has been shown that Sgk-1 interacts with and
activates IK in MDA231 cells by phosphorylating serine
181, thus allowing the degradation of IB.37 Together with
the findings that Rac is able to activate NFB and to control
TF expression,17 this suggests that activation of NADPH
oxidases, PI3K, p38MAPK, and PDK1 is required for induc-
tion of Sgk-1, which then phosphorylates IK, allowing
degradation of IB, nuclear translocation of NFB, and
transcriptional activation of TF (Figure 7).
Figure 6. Sgk-1 is expressed in the media of remodeled pulmo-
nary vessels. Immunohistochemistry was performed using an
antibody against Sgk-1 in lung tissue with media hypertrophy Sgk-1 Is Associated With Pulmonary
and vascular remodeling (A) or without signs of remodeling (B), Vascular Remodeling
obtained from a lung tumor resection. Smooth muscle cells In addition to our findings in cultivated PASMC, Sgk-1
were identified using a SMC-␣ actin antibody. C, Immunolocal-
ization of TF was determined in remodeled pulmonary tissue.
protein was detected in the media of small vessels in lung
tissue with vascular remodeling and PH. In contrast, Sgk-1
over, TF expression was not detectable in sgk1⫺/⫺ fibroblasts was not expressed in airway smooth muscle cells or in lung
vessels without signs of pulmonary disease, suggesting the
stimulated with thrombin or in lung tissue from sgk1⫺/⫺ mice
specific upregulation of Sgk-1 in remodeled vessels. Interest-
treated with dexamethasone, known to stimulate TF25 and
ingly, Sgk-1 has been implicated in vasculogenesis in mouse
Sgk-1.7 The importance of Sgk-1 for thrombin-induced TF
development during embryonic days 8.5 to 12.5.26 Moreover,
expression is further supported by our previous findings that
although Sgk-1 was increased in fibrotic lungs in particular in
TF mRNA and protein expression peak at 4 and 8 hours of
bronchi, lung epithelial cells or lung fibroblasts, Sgk-1 was
stimulation with thrombin in PASMC, respectively,17 thus
barely detectable in nondiseased lung vessels.13 In addition,
clearly following the upregulation of Sgk-1. In addition,
TF protein colocalized with Sgk-1 in the media of small
treatment with the antioxidants vitamin C or NAC (data not
vessels, further indicating that Sgk-1 and TF are involved in
shown), expression of dominant-negative Rac or depletion of
pulmonary vascular remodeling in PH.
p22phox prevented thrombin-induced TF expression in
Taken together, our data provide evidence that thrombin
PASMC and other smooth muscle cells.17,19 Moreover, sim-
activates NADPH oxidases to generate ROS, thus leading to
ilar concentrations of PI3K or p38MAPK inhibitors were activation of PI3K or p38MAPK followed by activation of
required to prevent TF upregulation by thrombin, whereas PDK1 and subsequent induction of Sgk-1. Furthermore,
inhibition of ERK1/2 was not effective,18,19 and MKK3 and Sgk-1 is involved in upregulation of TF by activating NFB.
PDK1 have been shown to regulate TF expression by throm- Because activated TF increases the formation of thrombin,
bin in PASMC.18 and Sgk-1 and TF proteins were present in pulmonary vessels
The importance of Sgk-1 for controlling TF expression was with vascular disease, this pathway may contribute to the
confirmed by our findings that Sgk-1 activates the NFB prothrombotic state and vascular remodeling in PH.
pathway in PASMC by promoting IB degradation, thus
allowing translocation of NFB to the nucleus and that NFB Acknowledgments
is required for Sgk-1–mediated activation of the TF promoter. This work was supported by Deutsches Forschungsgemeinschaft
Consistently, NFB activation is sensitive to ROS and GO709/4-4 and GRK438 and Fondation Leducq (to A.G.). We
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015836 Circulation Research March 31, 2006
acknowledge the expert technical assistance of Kerstin Diemer and 19. Herkert O, Diebold I, Brandes RP, Hess J, Busse R, Gorlach A. NADPH
Stefan Würth and scientific advice of Isabel Diebold, MD. oxidase mediates tissue factor-dependent surface procoagulant activity by
thrombin in human vascular smooth muscle cells. Circulation. 2002;105:
2030 –2036.
References 20. Salvi R, Peclat V, So A, Busso N. Enhanced expression of genes involved
1. Humbert M, Morrell NW, Archer SL, Stenmark KR, MacLean MR, Lang in coagulation and fibrinolysis in murine arthritis. Arthritis Res. 2000;2:
IM, Christman BW, Weir EK, Eickelberg O, Voelkel NF, Rabinovitch M. 504 –512.
Cellular and molecular pathobiology of pulmonary arterial hypertension. 21. Park J, Leong ML, Buse P, Maiyar AC, Firestone GL, Hemmings BA.
J Am Coll Cardiol. 2004;43:13S–24S. Serum and glucocorticoid-inducible kinase (SGK) is a target of the PI
2. Chaouat A, Weitzenblum E, Higenbottam T. The role of thrombosis in 3-kinase-stimulated signaling pathway. EMBO J. 1999;18:3024 –3033.
severe pulmonary hypertension. Eur Respir J. 1996;9:356 –363. 22. Gorlach A, Diebold I, Schini-Kerth VB, Berchner-Pfannschmidt U, Roth
3. Mann KG. Thrombin formation. Chest. 2003;124:4S–10S. U, Brandes RP, Kietzmann T, Busse R. Thrombin activates the hypox-
4. Eto M, Kozai T, Cosentino F, Joch H, Luscher TF. Statin prevents tissue ia-inducible factor-1 signaling pathway in vascular smooth muscle cells:
factor expression in human endothelial cells: role of Rho/Rho-kinase and role of the p22(phox)-containing NADPH oxidase. Circ Res. 2001;89:
Akt pathways. Circulation. 2002;105:1756 –1759. 47–54.
5. Herkert O, Djordjevic T, BelAiba RS, Gorlach A. Insights into the redox 23. Brandes RP, Viedt C, Nguyen K, Beer S, Kreuzer J, Busse R, Gorlach A.
control of blood coagulation: role of vascular NADPH oxidase-derived Thrombin-induced MCP-1 expression involves activation of the
reactive oxygen species in the thrombogenic cycle. Antioxid Redox p22phox-containing NADPH oxidase in human vascular smooth muscle
Signal. 2004;6:765–776. cells. Thromb Haemost. 2001;85:1104 –1110.
6. Webster MK, Goya L, Ge Y, Maiyar AC, Firestone GL. Characterization 24. Kobayashi T, Cohen P. Activation of serum- and glucocorticoid-regulated
of sgk, a novel member of the serine/threonine protein kinase gene family protein kinase by agonists that activate phosphatidylinositide 3-kinase is
which is transcriptionally induced by glucocorticoids and serum. Mol Cell mediated by 3-phosphoinositide-dependent protein kinase-1 (PDK1) and
Biol. 1993;13:2031–2040. PDK2. Biochem J. 1999;339(pt 2):319 –328.
7. Firestone GL, Giampaolo JR, O’Keeffe BA. Stimulus-dependent regu- 25. Reddy KV, Bhattacharjee G, Schabbauer G, Hollis A, Kempf K, Tencati
lation of serum and glucocorticoid inducible protein kinase (SGK) tran- M, O’Connell M, Guha M, Mackman N. Dexamethasone enhances LPS
scription, subcellular localization and enzymatic activity. Cell Physiol induction of tissue factor expression in human monocytic cells by
Biochem. 2003;13:1–12. increasing tissue factor mRNA stability. J Leukoc Biol. 2004;76:145–151.
8. Lang F, Cohen P. Regulation and physiological roles of serum- and 26. Lee E, Lein ES, Firestone GL. Tissue-specific expression of the tran-
glucocorticoid-induced protein kinase isoforms. Sci STKE. 2001; scriptionally regulated serum and glucocorticoid-inducible protein kinase
108:RE17. (Sgk) during mouse embryogenesis. Mech Dev. 2001;103:177–181.
9. Waldegger S, Barth P, Raber G, Lang F. Cloning and characterization of 27. Singh R, Pan S, Mueske CS, Witt T, Kleppe LS, Peterson TE, Slobodova
a putative human serine/threonine protein kinase transcriptionally A, Chang JY, Caplice NM, Simari RD. Role for tissue factor pathway in
modified during anisotonic and isotonic alterations of cell volume. Proc murine model of vascular remodeling. Circ Res. 2001;89:71–76.
Natl Acad Sci U S A. 1997;94:4440 – 4445. 28. Leong ML, Maiyar AC, Kim B, O’Keeffe BA, Firestone GL. Expression
10. Kumar JM, Brooks DP, Olson BA, Laping NJ. Sgk, a putative serine/ of the serum- and glucocorticoid-inducible protein kinase, Sgk, is a cell
threonine kinase, is differentially expressed in the kidney of diabetic mice survival response to multiple types of environmental stress stimuli in
and humans. J Am Soc Nephrol. 1999;10:2488 –2494. mammary epithelial cells. J Biol Chem. 2003;278:5871–5882.
11. Friedrich B, Warntges S, Klingel K, Sauter M, Kandolf R, Risler T, 29. Shelly C, Herrera R. Activation of SGK1 by HGF, Rac1 and integrin-
mediated cell adhesion in MDCK cells: PI-3K-dependent and
Muller GA, Witzgall R, Kriz W, Grone HJ, Lang F. Up-regulation of the
-independent pathways. J Cell Sci. 2002;115:1985–1993.
human serum and glucocorticoid-dependent kinase 1 in glomerulone-
30. Bell LM, Leong ML, Kim B, Wang E, Park J, Hemmings BA, Firestone
phritis. Kidney Blood Press Res. 2002;25:303–307.
GL. Hyperosmotic stress stimulates promoter activity and regulates
12. Fillon S, Klingel K, Warntges S, Sauter M, Gabrysch S, Pestel S, Tanneur
cellular utilization of the serum- and glucocorticoid-inducible protein
V, Waldegger S, Zipfel A, Viebahn R, Haussinger D, Broer S, Kandolf R,
kinase (Sgk) by a p38 MAPK-dependent pathway. J Biol Chem. 2000;
Lang F. Expression of the serine/threonine kinase hSGK1 in chronic viral
275:25262–25272.
hepatitis. Cell Physiol Biochem. 2002;12:47–54.
31. Meng F, Yamagiwa Y, Taffetani S, Han J, Patel T. IL-6 activates serum
13. Waerntges S, Klingel K, Weigert C, Fillon S, Buck M, Schleicher E,
and glucocorticoid kinase via p38alpha mitogen-activated protein kinase
Rodemann HP, Knabbe C, Kandolf R, Lang F. Excessive transcription of pathway. Am J Physiol Cell Physiol. 2005;289:C971–C981.
the human serum and glucocorticoid dependent kinase hSGK1 in lung 32. Mizuno H, Nishida E. The ERK MAP kinase pathway mediates induction
fibrosis. Cell Physiol Biochem. 2002;12:135–142. of SGK (serum- and glucocorticoid-inducible kinase) by growth factors.
14. Busjahn A, Aydin A, Uhlmann R, Krasko C, Bahring S, Szelestei T, Feng Genes Cells. 2001;6:261–268.
Y, Dahm S, Sharma AM, Luft FC, Lang F. Serum- and glucocorticoid- 33. Barthel A, Klotz LO. Phosphoinositide 3-kinase signaling in the cellular
regulated kinase (SGK1) gene and blood pressure. Hypertension. 2002; response to oxidative stress. Biol Chem. 2005;386:207–216.
40:256 –260. 34. Prasad N, Topping RS, Zhou D, Decker SJ. Oxidative stress and vanadate
15. Delmolino LM, Castellot JJ Jr. Heparin suppresses sgk, an early response induce tyrosine phosphorylation of phosphoinositide-dependent kinase 1
gene in proliferating vascular smooth muscle cells. J Cell Physiol. 1997; (PDK1). Biochemistry. 2000;39:6929 – 6935.
173:371–379. 35. Edmead C, Kanthou C, Benzakour O. Thrombin activates transcription
16. Wulff P, Vallon V, Huang DY, Volkl H, Yu F, Richter K, Jansen M, factors sp1, NF-kappaB, and CREB: importance of the use of phosphatase
Schlunz M, Klingel K, Loffing J, Kauselmann G, Bosl MR, Lang F, Kuhl inhibitors during nuclear protein extraction for the assessment of tran-
D. Impaired renal Na(⫹) retention in the sgk1-knockout mouse. J Clin scription factor DNA-binding activities. Anal Biochem. 1999;275:
Invest. 2002;110:1263–1268. 180 –186.
17. Djordjevic T, Herkert O, Hess J, Gorlach A, BelAiba RS. Rac regulates 36. Morris RC, Morris GZ, Zhang W, Gellerman M, Beebe SJ. Differential
thrombin-induced tissue factor expression in pulmonary artery smooth transcriptional regulation by the alpha- and gamma-catalytic subunit
muscle cells involving the nuclear factor kappa B pathway. Antioxid isoforms of cAMP-dependent protein kinase. Arch Biochem Biophys.
Redox Signal. 2004;6:713–720. 2002;403:219 –228.
18. Gorlach A, BelAiba RS, Hess J, Kietzmann T. Thrombin activates the 37. Zhang L, Cui R, Cheng X, Du J. Antiapoptotic effect of serum and
p21-activated kinase in pulmonary artery smooth muscle cells. Role in glucocorticoid-inducible protein kinase is mediated by novel mechanism
tissue factor expression. Thromb Haemost. 2005;93:1168 –1175. activating I{kappa}B kinase. Cancer Res. 2005;65:457– 464.
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015The Serum- and Glucocorticoid-Inducible Kinase Sgk-1 Is Involved in Pulmonary
Vascular Remodeling: Role in Redox-Sensitive Regulation of Tissue Factor by Thrombin
Rachida S. BelAiba, Talija Djordjevic, Steve Bonello, Ferruh Artunc, Florian Lang, John Hess
and Agnes Görlach
Circ Res. 2006;98:828-836; originally published online February 16, 2006;
doi: 10.1161/01.RES.0000210539.54861.27
Circulation Research is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231
Copyright © 2006 American Heart Association, Inc. All rights reserved.
Print ISSN: 0009-7330. Online ISSN: 1524-4571
The online version of this article, along with updated information and services, is located on the
World Wide Web at:
http://circres.ahajournals.org/content/98/6/828
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published
in Circulation Research can be obtained via RightsLink, a service of the Copyright Clearance Center, not the
Editorial Office. Once the online version of the published article for which permission is being requested is
located, click Request Permissions in the middle column of the Web page under Services. Further information
about this process is available in the Permissions and Rights Question and Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Circulation Research is online at:
http://circres.ahajournals.org//subscriptions/
Downloaded from http://circres.ahajournals.org/ by guest on October 25, 2015You can also read

















































