The spatial structure of а snow leopard population (Panthera uncia, Felidae, Carnivora) in east Kyrgyzstan - Research Article
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Ecologica Montenegrina 33: 17-28 (2020)
This journal is available online at: www.biotaxa.org/em
http://dx.doi.org/10.37828/em.2020.33.4
The spatial structure of а snow leopard population (Panthera uncia,
Felidae, Carnivora) in east Kyrgyzstan
VICTOR S. LUKAREVSKIY1, ALEXANDER P. VERESHCHAGIN2 &
SVYATOSLAV V. LUKAREVSKIY3
1
Informational and Analytical Center for Support of Conservation, 123242, Moscow, Russia
2
Sarychat-Ertash Reserve, Karakol, Kyrgyzstan
3
The Center for Research and Conservation of Wild Cats, Chernogolovka, Moscow Oblast, Russia
E-mail: vlukarevsky@mail.ru
Received 2 June 2020 │ Accepted by V. Pešić: 12 July 2020 │ Published online 18 July 2020.
Abstract
The distribution of the snow leopard (Panthera uncia uncia (Shreber, 1775) across its geographic range is associated
exclusively with the mountain landscapes. The most important factor in maintaining the integrity and viability of a
population is its structure.
Our studies were conducted in 2010, 2011 and 2012 and covered most of the range of snow leopard in East
Kyrgyzstan. Each territory was surveyed through series of routes. The main methods used in field research were:
searching for traces of vital activity, conducting short trail surveys, and using the SLIMS (Snow Leopard Information
Management System) technique.
In order to study the spatial structure of the snow leopard population, we conducted individual DNA-based
identification and GPS registration of bioassays collected from all individuals inhabiting this territory. Biological
samples (tissue, fur, and excrement) were obtained following the protocols for collecting bioassays. A total of 146
samples (2 of wool, 2 of tissue, and 142 of excrement) were collected in nature in the Central Tien Shan Mts within
Kyrgyzstan (in the Sarychat-Ertash Reserve, and in the area of the Kainar River).
Snow leopard DNA was identified in 70 samples. A total of 21 individual snow leopards (8 females and 13
males) were identified. All females of a local population of the Sarychat-Ertash Reserve, with the exception of one,
were directly related to each other. The same is true for most of males (11 of 13). Two females were accompanied by
kittens. All females settled near the maternal site, while almost all males, which were born there, move away. There is a
direct correlation between population density and marking activity.
Key words: population, scrapings, identification, marking, Panthera uncia, Kyrgyzstan.
Introduction
The distribution of the snow leopard (Panthera uncia uncia (Shreber, 1775) throughout the range is
associated exclusively with the mountains. Unlike in other large cats, most of the world's snow leopards live
at the altitudes over 2,000-2,500 m above sea level (Jackson and Hunter, 1996). Other authors consider
altitude as a secondary factor (Heptner and Sludsky, 1972; Zavatsky, 2004; Lukarevskiy and Poyarkov 2008;
Ecologica Montenegrina, 33, 2020, 17-28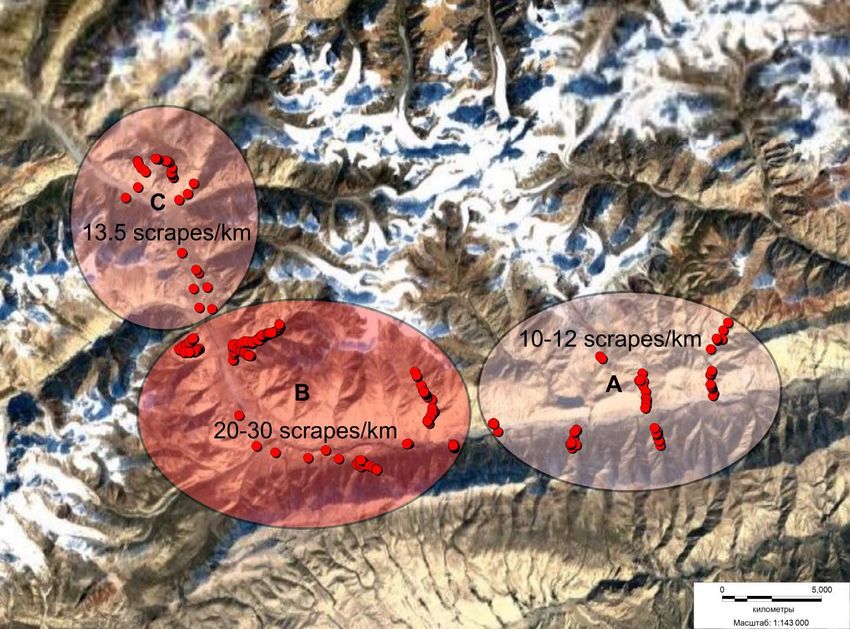
THE SPATIAL STRUCTURE OF А SNOW LEOPARD POPULATION IN EAST KYRGYZSTAN
Istomov, 2013). In the choice of habitats by the snow leopard, the relief structure appears to be more
important than the elevation. They live in the mountainous terrain with highly dissected slopes, with a lot of
protected habitats, and a large number of wild ungulates (Koshkarev, 1988; Ahlborn and Jackson, 1988;
Jackson, Hunter, 1996; Lukarevskiy and Poyarkov, 2008). In the studied part of the range in Kyrgyzstan,
snow leopards are found exclusively within the highlands (Koshkarev, 1988, 1989; Matyushkin and
Koshkarev, 1990).
Most of the studies of the snow leopard analyzed distribution and number of animals, while little
attention was given to the spatial structure of local populations. At the same time, in our opinion, it is the
fully developed, spatial-ethological structure of a population of any species that determines its well-being
(Hornocker and Bailey, 1986; Wolff, 1997; Goodrich et al., 2005).
For 20 years (1990s-2010s) the population of the Central Tien Shan snow leopard has undergone
serious changes. In the 1990s, the number of snow leopards decreased sharply throughout its range in
Kyrgyzstan (Koshkarev, 2002; Koshkarev and Vyrypaev, 2002). In the 2000s, in fact, only the local group in
the Sarychat-Ertash Reserve regained its numbers (Lukarevskiy and Umetbekov, 2011). Our studies show
that a significant part of the Tien Shan groups disappeared, which led to a strong fragmentation in the
geographic range of this species. Other groups were severely degraded. Snow leopard tracks were registered
in only three out of ten regions we surveyed in East Kyrgyzstan,
The goal of this work is to show the local structure of the snow leopard population and its role in
maintaining the integrity of metapopulations.
Materials and methods
Our studies, covering most of East Kyrgyzstan (Fig. 1), were conducted from July 29 to August 12, 2010;
from June 10 to 22, 2011; and in March 2012. The choice of territories for the survey was based on the map
analysis and surveys of local residents, taking into account the habitat preferences of the snow leopard
(Koshkarev, 1988; Matyushkin and Koshkarev, 1990; Jackson and Hunter, 1996). We ignored easily
accessible areas, because we believed that if the snow leopard is absent in hard-to-reach areas, then it is
cannot be found in the areas to which the locals can have easy access. The survey data were treated with
great caution, especially if these areas were used for livestock grazing. To test of such intermediate areas, we
made small routes. For each territory we surveyed a series of routes. Usually a route length was about 10-15
km (Lukarevskiy and Umetbekov, 2011).
Climate of the Central Tien Shan (dry/cold or dry/warm air) allows for longer preservation of DNA
in excrement (i. e., excrement is mummified). We paid special attention to protocols for collecting DNA
bioassays. Samples of excrement were chosen from the sites that were not exposed to UV radiation (e.g.,
were located under the rock ledges). For primary species identification, we carefully examined the
appearance of excrement, its localization and freshness. In order to study the feeding habits of the snow
leopard, the species composition of the prey remains in the excrement was identified. We registered only
those excrement samples that were definitely identified as belonging to snow leopards.
As we designed survey routes, we took into consideration that the snow leopard uses certain types of
landforms where it moves and makes marks (Koshkarev, 1988; Ahlborn and Jackson, 1988; Jackson and
Hunter, 1996; Lukarevskiy and Poyarkov, 2008; Lukarevskiy and Purevsuren, 2011; Lukarevskiy and
Umetbekov, 2011). As the main method of field research, we used searching for traces of vital activity;
where possible, we conducted short trail surveys, and also used the technique of SLIMS (Snow Leopard
Information Management System) (Jackson and Hunter, 1996). The placement of scrapes and excrement by
the snow leopard, as well as other leopard species, is typical (Koshkarev, 1988; Jackson and Hunter, 1996;
Lukarevskiy, 2001, 2005; Lukarevskiy and Poyarkov, 2008; Lukarevskiy et al., 2004, et al.). Most of them
are recorded on the crests of mountain ridges, often in saddles near large stones or rocks, near large, clearly
distinguished trees. Similar marks are also observed at the bottom of gorges: at large stones, trees; along
riverbeds and in the points where two or three mountain spurs come together, which are the places of
movement of the ungulates that constitute their prey: Siberian mountain goat (Capra sibirica Pallas, 1776) &
mountain sheep (Ovis ammon Linneus, 1758). Very often the snow leopard leaves scrapes, urinating points
and excrement under an overhanging rock, in small grottoes located on the slopes of gorges. In a situation
where there is no pronounced trail on the slope of the gorge, it is not marked, except the cases when the snow
leopard gets its prey there. In such a case, scrapes and excrement can be found near the closest rocks or large
18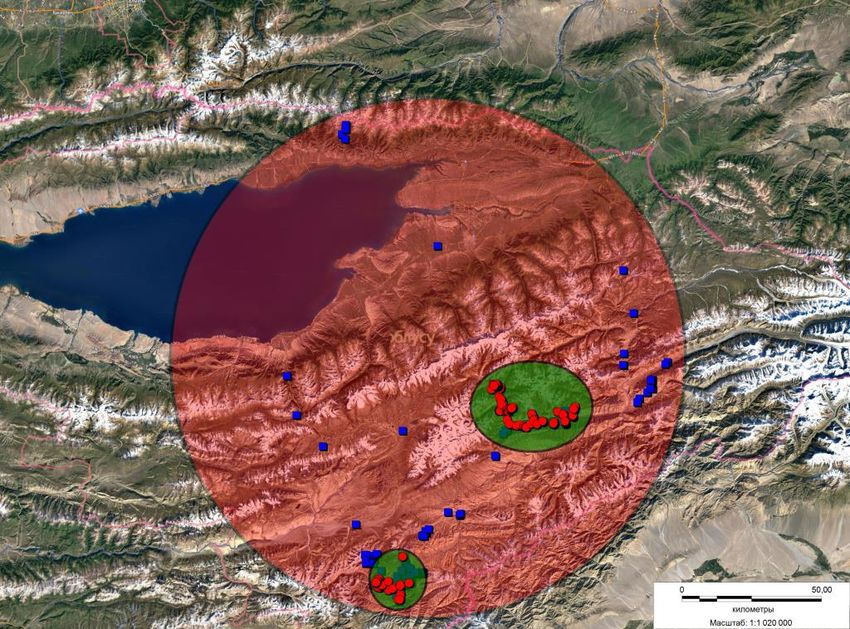
LUKAREVSKIY ET AL.
stones. As a rule, the snow leopard leaves scrapes, urinary points and excrement, at the location it marked
earlier. This demonstrates a rigid spatial ordering and conservatism of the marking system, and as a result,
clusters of scrapes are formed. As a rule, a cluster of scrapes is formed when snow leopard makes its mark
over the mark of another individual; it looks like soil loosened by numerous deep furrows (scrapes) taking an
area of 2-10 m2 (Lukarevskiy et al., 2019). Existence of such clusters is directly connected to snow leopard
population density (Ahlborn and Jackson, 1988; Jackson and Hunter, 1996; Lukarevskiy et al., 2019).
Figure 1. Areas of snow leopard study in the East Kyrgyzstan. Blue squares-surveyed areas; red points, traces of snow
leopard activity.
As a characteristic of the density of the elements of marking behavior, we will use only such signs as
a scrapes. The scrapes are the most typical and easily recognizable traces made by the snow leopard. They
persist much longer than other life signs (up to a year or more, depending on location). However, in summer,
during drought, and in winter during frosts, in the open areas, scrapes may be less visible. Such scrapes are
usually difficult to recognize and less durable, since they are destroyed by the first heavy rain or snowfall;
some experience is needed to recognize them. At the same time, scrapes located under an overhanging rock
can be preserved for several years. In the studied parts of the snow leopard's range, no other animals exist
capable of leaving similar scrapes. Sometimes, on a cursory inspection, the snow leopard's scrapes can be
confused with the dust "baths" of gallinaceous birds, with the small ruts of wild boars (Sus scrofa Linneus,
1758) or brown bears (Ursus arctos Linneus, 1758) that graze along the trail or a ridge crest. Tracks and
track chains do not always show the attachment of animals to a given territory, but can only indicate the
movements or temporary use of a territory. Spray urination and "urinary points" in a scrape can only be
recognized while very fresh or only in winter on the snow. These two forms of marking are often associated
with the physiological state of the animal, namely, estrus for females or urination. Excrement is not a reliable
sign, since in the frost-free period of the year, a lot of excrement gets destroyed or eaten by coprophages and
Ecologica Montenegrina, 33, 2020, 17-28 19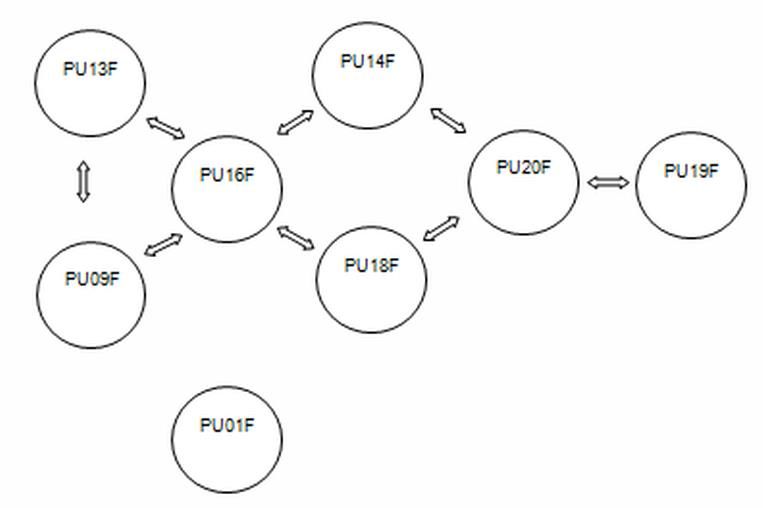
THE SPATIAL STRUCTURE OF А SNOW LEOPARD POPULATION IN EAST KYRGYZSTAN
bird lice, so their quantitative estimates are very difficult. Rubbings by the cheeks and preorbital glands are
almost impossible to identify in the field.
To identify the freshness of traces of snow leopard activity, we always took into account weather
conditions: strong wind, fog, rain, snow powder, the condition of damaged plants. The condition of damaged
plants during their growth season allows the most reliable determination of the age of scrapes. Determination
of the age of traces of vital activity was necessary to estimate the frequency of visits to a certain site by snow
leopards, as well as the freshness of excrement. As in other areas, such as Mongolia, the intensity of marking
increases from the periphery to the center of the group’s range (Lukarevskiy and Purevsuren, 2011;
Lukarevskiy et al., 2019). Our field observations in the Sayano-Shushensky Reserve, and on some ranges in
the Tien Shan showed that the snow leopard marks the territory only if there is a female present. At a site
where only one male remained, his marking activity gradually began to fade, and after a year we did not
register a single scratch (our unpublished data).
The intensity of the marking of the territory is in direct correlation with the condition of the snow
leopard population (Ahlborn and Jackson, 1988; Jackson and Hunter, 1996; Lukarevskiy et al., 2019). We
believe that the intensity of marking (by scrapes) also indicates the degree of development of the spatial-
ethological structure of the population, and that the scrapes indicate the claim of an individual leopard to a
given territory. An increase in animal density leads to an increase in social tension in the population, and as a
consequence to an increase in marking activity.
In places where only a few footprints were registered, without scrapes, they were considered only as
a result of accidental movement of males. In most of the territory that we surveyed in Tien Shan (Fig. 1), the
traces of snow leopards were not available.
In order to investigate the spatial distribution of the snow leopard within the studied group in the
Sarychat-Ertash reserve, we had to conduct identification of all individuals inhabiting this territory. For this
purpose, we organized the collection of biological samples (tissue, fur, and excrement) using non-invasive
research methods (Janecka et al., 2020). A total of 146 samples (2 of wool, 2 of tissue and 142 of
excrement), collected in nature in the Central Tien Shan in Kyrgyzstan within the Sarychat-Ertash Reserve,
were studied. The samples were fixed in 96% ethanol. The genetic analysis of the samples was carried out in
the Laboratory of Molecular Diagnostic Methods of the IPEE RAS by E.Yu. Zvychaynaya & M.P. Korablev.
Total DNA was isolated with QIAamp DNA Stool Mini Kit (QIAGEN), which proved to be the best
method for DNA isolation from cat excrement (Bhagavatula and Singh, 2006). DNA identification consists
of two stages: the primary establishment of species belonging to the animal tissue sample or traces of its life
and subsequent individualization. The method of species and individual identification used followed Janecka
et al. (2008), Zvychaynaya et al. (2011), and Rozhnov et al. (2011). Eight microsatellite loci were selected
for individual identification of individual snow leopards (Janecka et al., 2008; Zvychaynaya et al., 2011)
(primary data available from the authors). Samples in which the genetic profile was identical and coincided
in all loci and alleles were attributed to one individual. For each sample, its geographic coordinates are
known, which allowed us to visualize the spatial distribution of individuals (Fig. 2.)
Another issue that we wanted to solve using the data of DNA analysis was to determine the
relationship between individuals of the local group. Samples, in which the genetic profile coincided in all
loci at least for one allele of each locus of an investigated segment of DNA, were considered related. In cases
where the genetic profile of an individual coincided with the genetic profiles of two unrelated females and
males, such individuals we considered their offspring. Individuals whose genetic profile coincided at all loci
and differed only in a few alleles and were spatially located in the same place, were considered close
relatives.
For all the collected materials, we had an exact geographic location, which allowed us to understand
how the animals are distributed in space; in many cases, the collected data allowed us to outline their
hypothetical ranges. Individual ranges of snow leopards were determined by the MCP (minimum convex
polygon) method (Hayne, 1949); all registrations were entered in the MapInfo program.
Another problem we wanted to solve was to find a correlation between the marking activity and the
density of the snow leopard population. Therefore, we compared the number of the identified individuals and
the number of markings for each specific territory. As a characteristic of the density of the elements of
marking behavior, we used only a "scrape", which is the most typical and pronounced form of marking
behavior. It should be noted that, despite the seasonality of reproduction, the marking activity of the snow
leopard practically does not change during the year (Lukarevskiy and Purevsuren, 2011), so the main routes
of the animals remain well marked by scrapes year-round.
20LUKAREVSKIY ET AL.
Figure 2. Sections 1–3 of different intensity of marking activity of the snow leopard. For a description, see text
(polygons A, B, C).
Results
We surveyed a significant part of the potential habitats of the snow leopard in East Kyrgyzstan (Fig.1). Out
of 10 regions surveyed in East Kyrgyzstan, traces of the snow leopard were registered in only three of them,
and one region showed only paw prints, without showing signs of territoriality of individuals. A small group
was located on the border with China near the Kainar site; there were various life traces of different age, but
judging by the number of registered signs of traces of life of the snow leopard, the grouping is insignificant
in size. One snow leopard has been identified for this area.
Our study was focused mostly on the territory of the Sarychat-Ertash Reserve, Issyk-Kul Province,
Kyrgyzstan (аreа 134,140 hectares), where the traces of snow leopard activity are registered on all the routes
surveyed. A stable snow cover was established here in November at the altitude of over 3500 m a.s.l. The
ungulates were distributed over the territory from the river valley to the snow line. The traces of life of the
snow leopard followed this distribution; almost all the scrapes of the snow leopard were concentrated in the
lower portion of the river valleys close to their mouths and to the base of the lateral spurs of the ranges. In
summer, the distribution of ungulates and therefore the distribution of snow leopards is significantly different
from their winter distribution. The material collected here in March 2012 characterizes the winter
distribution of the snow leopards in the mountains of the Central Tien Shan. Their marking activity here was
the highest in the region.
At the same time, the life traces of the snow leopard in this territory are unevenly distributed. We
have identified several sites where the marking activity was the highest (Fig. 2).
In the area of Lake Baskel (Fig. 2, polygon A), the density of marking features was the lowest at 10-
12 scrapes/km of the route.
Ecologica Montenegrina, 33, 2020, 17-28 21THE SPATIAL STRUCTURE OF А SNOW LEOPARD POPULATION IN EAST KYRGYZSTAN
In the Solomo-Sary-Echki section (Fig. 2, polygon B), marking and trace activity was the highest,
from 20 to 30 and more scrapes per km. In the Solomo site, at the lateral spurs of the range, within 0.9 km
we recorded marked snow leopard tracks, 4 excrement samples and 30 scrapes (33.3 scrapes/km) including
4 clusters (2-3 or more scrapes of different age). Within the area from the Eshegart-Sary-Echki road, up the
river, at its left tributary, the left side spurs of the ridge (from the riverbed to 3000-4000 m a.s.l.), on a site of
2.5 km we recorded snow leopard tracks of different age, 58 scrapes of different age (23.2 scrapes km) and 3
excrement samples.
However, in March 2012, on a monitoring route where in August 2011 there was a high marking
activity, there were practically no fresh traces. At the boundary of the Solomo-Sary-Echki and Koilyu, we
found the remains of the two kittens a little less than one year old. The kittens died around the end of January
– beginning of February, or possibly earlier. However, we attribute the low marking activity on this route to
the death of an adult female that was recorded here in 2010 and 2011 but was not found anywhere in March
2012.
The next location is Boroko-Bordo Koilyu (Fig. 2, polygon C). In the Koilyu River watershed, at the
left side spur of the range within the route of about 12 km along the range, 20 scrapes of different age were
registered. Here, on a spur of the right slope of the range in a route of about 6 km, were recorded snow
leopard traces of different age, 81 scrapes (13.5 per km) and 10 excrement samples. Within the same site on
another route in the valley of the Sarychat River (13 km), under the ledges of the first level terrace, we
recorded 22 scrapes of different age.
We estimate the effective area of winter habitats of the snow leopard within these three areas in the
Sarychat-Ertash Reserve and its protection zone at 40-45,000 hectares. In March 2012, this territory was
inhabited by at least 10 adult snow leopards and 5 kittens. The total population was about 17-18 individuals.
Table. 1. The genetic profile of the identified individual snow leopards. Materials of the 2011-2012 collection in
Kyrgyzstan.
Code/ PUN894 PUN132 PUN272 PUN834 PUN229 PUN124 PUN935 PUN1157 Gender
Locus* (r6g) (tam-ra) (fam) (rox) (r6g) (tam-ra) (fam) (rox)
PU01F 115 123 117 127 121 127 114 116 111 111 96 106 120 120 107 109 ♀
PU02M 115 119 123 127 123 125 112 116 109 117 96 96 120 120 107 111 ♂
PU03M 119 123 125 127 121 123 116 116 111 111 96 102 116 120 107 107 ♂
PU04M 119 123 125 127 121 123 118 118 111 113 96 102 116 120 107 107 ♂
PU05M 119 121 125 127 125 127 114 116 111 115 102 106 116 120 107 107 ♂
PU06M 115 115 125 127 121 127 112 114 115 115 96 96 116 120 109 115 ♂
PU07M 121 123 125 127 121 127 114 118 111 115 96 98 120 120 107 114 ♂
PU08M 115 115 125 127 121 127 114 116 115 115 96 96 116 120 109 115 ♂
PU09F 115 123 125 127 123 127 118 118 111 115 96 96 116 116 111 111 ♀
PU10M 115 121 125 127 123 127 116 118 115 115 96 96 116 116 107 111 ♂
PU11M 115 115 125 127 125 127 116 118 113 115 96 102 116 120 111 111 ♂
PU12M 115 115 125 127 125 127 116 116 115 115 96 102 116 120 107 111 ♂
PU13F 115 123 125 127 123 127 118 118 111 115 96 96 110 116 109 111 ♀
PU14F 115 121 123 125 123 125 116 118 109 115 102 106 116 120 107 111 ♀
PU15M 115 121 123 127 121 127 114 116 113 115 96 98 120 120 107 114 ♂
PU16F 115 121 123 127 125 127 116 118 115 115 96 102 116 116 111 111 ♀
PU17M 121 123 125 127 127 127 114 118 111 115 96 102 120 120 107 114 ♂
PU18F 115 119 117 123 119 125 114 116 115 115 96 106 116 120 111 111 ♀
PU19F 115 119 123 125 121 125 116 118 107 109 96 106 120 120 107 111 ♀
PU20F 115 119 123 125 121 125 116 118 109 115 96 106 120 120 107 111 ♀
PU21M 115 115 127 127 125 127 118 118 115 115 96 102 116 120 111 111 ♂
* - Code of the identified individual / loci with primers (in brackets-fluorescent paint).
22LUKAREVSKIY ET AL.
Snow leopard DNA was identified in 70 samples. In 16 samples, DNA was severely degraded and
did not allow unambiguous interpretation of the results, therefore results from 54 samples were used for
individual identification. A total of 21 snow leopards (8 females and 13 males) were individually identified
(Table 1). 20 individuals were identified in the local population of the Sarychat-Ertash Reserve. All females
of the local population of the Sarychat-Ertash Reserve are directly related to each other, with the exception
of PU01F (Fig. 3). Almost all males (9 out of 11) are also are directly related (Table 1). Two males (PU02M
and PU04M) are not directly related to any other males (Fig. 4); perhaps their father either remained
unrecorded by us, or he is no longer in this group, as well as one of the females (PU18F). We assume that
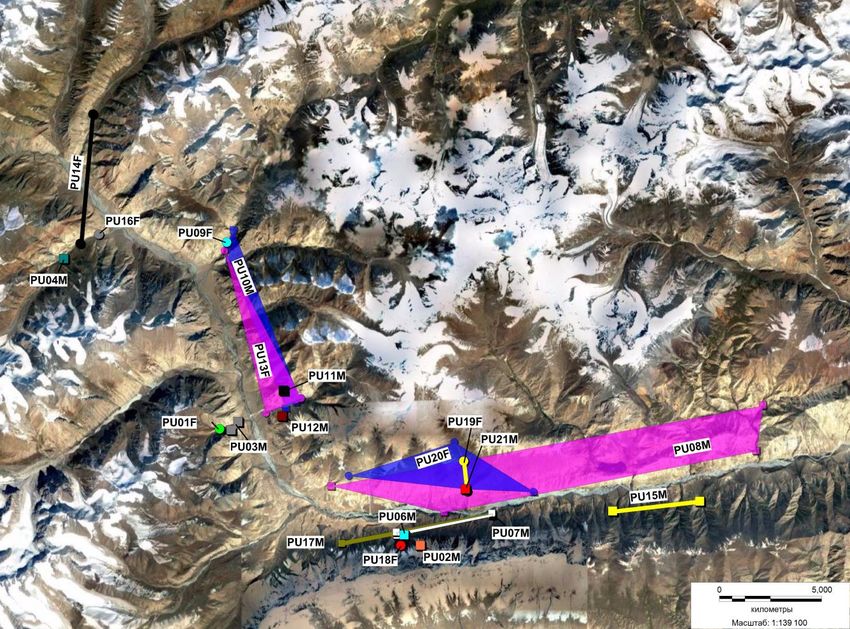
both of these males came from outside of this area. Their spatial placement is shown in Fig. 5. Three males
registered in August 2011, related to different females of this group, were not registered in the spring of
2012. It can be assumed that these males moved outside of the study group, as well as the male PU05M,
whose parents (according to the genetic profile) were in the Sarychat-Ertash Reserve in March 2012. This
male was recorded in another group 70 km away at the Kainar site, on the border with China.
Figure 3. Relationship between the females of the Sarychat –Ertash Reserve (as inferred from the DNA microsatellite
profiles).
Female PU01F most likely has died, since in March 2012, we did not discover her life traces, and
within her habitat we found the remains of two young kittens. It was the only female recorded in the Reserve
that was not related to other females.
Thus, the snow leopard group of the Sarychat-Ertash Reserve in March 2012 included at least 16
individuals: 7 females and 9 males. Judging from the genetic profile and spatial distribution, it can be
assumed that in April 2012, two females were accompanied by kittens (female PU13F was accompanied by
three kittens: two sons PU10M and PU11M and daughter PU09F, and female PU09F was accompanied by
daughter PU19F and son PU21M).
The marking activity of the snow leopard, as mentioned above, differed at different sites. We
observed a correlation between population density and marking activity: as the population density increased,
the marking activity increased as well. The same results were recorded in the Jargalant Mountains in
Mongolia (Lukarevskiy et al., 2019). In the Bashkel section of the Sarychat-Ertash Reserve, the marking
activity was 10-12 scrapes/km, while in the Solomo-Sary-Echki section it was from 23 to 33 scrapes/km. In
this area, the highest density of the snow leopard was noted, where almost two-thirds of the group was
concentrated. In fact, only in this area the clusters of scrapes have been recorded.
Ecologica Montenegrina, 33, 2020, 17-28 23THE SPATIAL STRUCTURE OF А SNOW LEOPARD POPULATION IN EAST KYRGYZSTAN
Figure 4. Relationship between the males of the Sarychat-Ertash Reserve (as inferred from the DNA microsatellite
profiles).
Discussion
The genetic diversity and structure of the snow leopard population in the Sarychat-Ertash Reserve could
testify to the "bottleneck" that this group passed through in the 1990s-2000s (Koshkarev, 2002; Koshkarev
and Vyrypaev, 2002). Our study shows that (according to the genetic profile) all females settled near the
maternal site, while almost all males born here move away, i.e. they provide a continuous gene flow within
the larger population, and thus prevent inbreeding. We see a deep biological meaning in this pattern.
Nomadic males are at greater risk, while females who remain within the same territory are more
protected from deadly accidents. Such strategy allows a population to avoids the risk of loss of its
reproductive potential, because it provides greater safety to the females.
Our data on the spatial structure of the population are fully consistent with those obtained at other
points of the snow leopard’s range (Jackson and Ahlborn, 1989; McCarthy et al., 2010; Sharma et al., 2014).
In the Sarychat-Ertash Reserve, we observe a "clan" structure of the snow leopard population based on the
relationship among females. It differs from a classical clan structure known in lions, where (Grinell et al.,
1995), in which a group of females lives within the same territory, jointly raising offspring obtained from
one or more related males. In the snow leopard clan, each female owns an individual site and independently
raises her offspring, and the male normally "owns" only one female—in fact, forming a family that can
persist for many years. Zoo observations (Koshkarev, Kuzminykh, 1988) show that the breeding success of
the snow leopard depends on the individual characteristics of each animal. Similar patterns have been
recorded for the Amur tiger (Panthera tigris altaica (Temminck, 1844) and the Far East leopard (Panthera
pardus orientalis (Schlegel, 1857); our data, unpublished). In these cases, the male actually provides a living
space for the female and her young. Our earlier results (Lukarevsky and Poyarkov, 2008), which showed
male owning several females, refer to the territories with a disturbed population structure.
Like other territorial mammals, the snow leopard has a complex of species-specific elements of
marking behavior that allow for regulation of territorial relationships. We believe that under a high
population density, social tension increases. This leads to increasing stress, and a sharp increase in marking
activity, which indirectly allows for reduction of territorial conflicts between animals.
Ovsyanikov & Badridze (1989) suggested animals affect and change the elements of the
environment in order to achieve a state of comfort; Naumov (1973, 1977) addressed this concept as a
biological signal field. The intensity of this signal field is determined by the structure and condition of a
24LUKAREVSKIY ET AL.
parcel group, which, as defined by Naumov (1967), is an elementary structural unit of a population. In our
case, such parcel groups, which can be considered a reproductive nucleus, are family groups consisting of
related females, occupying a single territory but having individual habitat areas. Metaphorically speaking, a
biological signal field is actually an “information field” and its "scanning" by snow leopards allows for
regulation of social relations and spatial distribution, In other words, the structure and level of development
of the biological signal field reflects the structure and condition of the population.
Figure 5. Spatial distribution of snow leopards in the Sarychat –Ertash Reserve.
In the study area, we observed the a developed spatial-ethological structure of the snow leopard
population. It is expressed as a high density of animals, with ratio of males and females close to 1:1. In this
group, there is an almost instantaneous replacement of the individual habitat area of one individual by
another. A similar example with a developed spatial-ethological structure of the population was observed in
the leopards (Panthera pardus) in the Central Kopet Dag and the Far East (Lukarevsky, 2005; unpublished
data), as well as in the population of the Amur tiger (Goodrich et al., 2005; our unpublished data, 2010-
2013). Goodrich et al. (2010) demonstrated that females own larger territories than they actually need, and
that they "pass" part of the land to their daughters. We suggest that in populations with a developed spatial-
ethological structure, formed on the basis of maternal clans, the reproductive potential is much higher than in
groups with a disturbed population structure.
Acknowledgments
Our field studies was conducted with the financial support of WWF Russia, for which the authors express their sincere
gratitude. The authors are grateful to O. B. Pereladova for support and assistance in conducting field research, as well as
to A. Alamanov, F. Balbakova and A. Umetbekov, and the rangers of the Sarychat-Ertash Reserve Akyl Kydyraliev,
Ecologica Montenegrina, 33, 2020, 17-28 25THE SPATIAL STRUCTURE OF А SNOW LEOPARD POPULATION IN EAST KYRGYZSTAN
Ulan Аbylgaziev and Toktobek uulu Urmat for assistance in the expedition and collection of field material. The authors
express special gratitude to E. Yu. Zvychaynаya and M. Korablev for carrying out DNA analysis of biological samples.
The authors are sincerely grateful to Victor Fet & Elinor Pate for the translation of this paper.
References
Ahlborn, G. G. & Jackson, R. M. (1988) Marking in free-ranging snow leopards in West Nepal: preliminary
assessment. Proceedings of the Fifth International Snow Leopard Symposium, pp. 25–49.
Bhagavatula, J. & Singh, L. (2006) Genotyping faecal samples of Bengal tiger Panthera tigris tigris for
population estimation: A pilot study. BMC Genetics, 7 (48), 1–12.
Geptner V. G. & Sludskiy A. A. (1972) Mlekopitayushchiye Sovetskogo Soyuza. [Mammals Soviet Union].
T. 2. Part 2. Carnivorous (hyenas and cats). M.: «Vysshaya shkola», 552 pp. (in Russian).
Goodrich, J. M., Kerley, L. L., Miquelle, D. G., Smirnov, E. N., Schleyer, B., Quigley, H. B., Hornocker, M.
G. & Ufyrkina, O. V. (2005) Sotsial'naya struktura populyatsii amurskogo tigra v Sikhote-Alinskom
biosfernom zapovednike) [Social structure of Amur tiger population in Sikhote-Alin biosphere
reserve]. In: Tigry Sikhote-Alinskogo zapovednika: ekologiya i sokhraneniye [Tigers of the Sikhote-
Alin Reserve: ecology and conservation], Vladivostok: PSP, 224 pp. (in Russian).
Goodrich, J.M., Miquelle, D. G., Smirnov, E. N., Kerley, L. L., Seredkin, I. V., Hornoker, M. G. & Quigley,
H. B. (2010) Razmer individual'nogo uchastka, kharakteristiki prostranstvennogo raspredeleniya i
raschetnaya plotnost' populyatsii amurskogo tigra.) [Individual plot size, spatial distribution
characteristics and estimated population density of the Amur tiger]. In: Amurskiy tigr v Severo-
Vostochnoy Azii: problemy sokhraneniya v XXI veke. Materialy mezhdunarodnoy nauchno-
prakticheskoy konferentsii 15-18 marta 2010 g.) [The Amur tiger in northeast Asia: conservation
challenges in the twenty-first century. Proceedings of the international scientific and practical
conference 15-18 March 2010], Vladivostok: Dal'nauka, pp. 49–59. (in Russian).
Grinell, J., Packer, C. & Pusey, A.E. (1995) Cooperation in male lions: kinship, reciprocity or mutualism?
Animal Behaviour, 49, 95–105.
Jackson, R. & Ahlborn, G. (1989) Snow leopards (Panthera uncia) in Nepal – Home Range and Movements.
National geographic research, 5 (2), 161–175.
Jackson, R.M. & Hunter, D.O. (1996) Snow Leopard Survey and Conservation Handbook. International
Snow Leopard Trust. Seattle, Second Edition, 189 pp.
Janeka, J. E., Jackson, R., Yuquang, Z. et al. (2008) Population monitoring of snow leopards using
noninvasive collection of scat samples: a pilot study. Animal Conservation, 11 (5), 401–411.
Janeka, J. E., Hacker, Ch., Broderick, J., Pulugulla S., Auron, Ph., Ringling, Mc. et al. (2020) Noninvasive
Genetics and Genomics Shed Light on the Status, Phylogeography, and Evolution of the Elusive
Snow Leopard In: Conservation Genetics in Mammals, pp. 83-120. DOI:10.1007/978-3-030-33334-
8_5
Hayne, D.W. (1949) Calculation of size of home range. Journal of Mammalogy, 30, 1–18.
Hornocker, M. G. & Bailey, T. N. (1986) Natural regulation in three species of Felids. In Cats of the World:
Biology, Conservation, and Management: 211–220. Miller, S. D. and Everett, D. D. (Eds.).
Washington, D.C.: National Wildlife Federation.
Istomov, S. V. (2013) Irbis Zapadnogo Sayana. [The Snow Leopard of The Western Sayan Mountains]
Abakan. LLC Cooperative Journalist, 128 pp. (in Russian).
Koshkarev, E. P. (1988) Territorial'noye razmeshcheniye, chislennost' i mestoobitaniya irbisa v Severnom i
Vnutrennem Tyan'-Shane) [Territorial distribution, numbers and habitats of the snow leopard in the
Northern and Inner Tien–Shan]. Bulletin of the Moscow Society of Naturalists. Div. Biol., 93(2), 3–1.
(in Russian).
Koshkarev, E. P. (1989) Snezhnyy bars v Kirgizii (struktura areala, ekologiya, okhrana) [Snow leopard in
Kirgizia (the structure of the habitat, ecology, conservation)]. Frunze. Publishing house "Ilim". 99
pp. (in Russian).
Koshkarev, E. (2002) Valyutnyye barany i ministerskiye volki Kirgizii [The hard-currency sheep and the
ministerial wolves of Kyrgyzstan]. Wildlife Protection, Moscow, 1(24), 26–32. (in Russian).
Koshkarev, E. P. & Kuzminykh, I. A. (1988) Prichiny izmeneniya chislennosti snezhnogo barsa i otsenka
yego populyatsii v prirode i nevole [Causes of the changes in snow leopard numbers and assessment
26LUKAREVSKIY ET AL.
of its population in the wild and in captivity]. In: Ekologicheskiye issledovaniya bioty ekosistem
Severnoi Kirgizii [Ecological studies of the biota of ecosystems of North Kirghizia]. Frunze, Ilim, pp.
95–124. (in Russian).
Koshkarev, E. P. & Vyrypaev, V. A. (2002) Sostoyaniye populyatsiy redkikh i promyslovykh zverey Kirgizii
vo vtoroy polovine XX veka) /The state of populations of rare and commercial animals of
Kyrgyzstan in the second half of the 20th century. Bulletin of the Moscow Society of Naturalists. Div.
Biol., 107(3), 3–11. (in Russian).
Lukarevsky V. S. (1993) Leopard (Panthera pardus) na Zapadnom Kopetdage: razmeshcheniye,
chislennost', osobennosti ekologii i povedeniya) [The leopard (Panthera pardus) of the Western
Kopetdag: distribution, numbers, features of ecology and behavior]. Zoological Journal, 72(1), 131–
141. (in Russian).
Lukarevsky, V. S. (2001) Leopard, polosataya giyena i volk v Turkmenistane. [The leopard, the striped
hyena, and the wolf in Turkmenistan] Moscow: Signar, 128 pp. (in Russian).
Lukarevsky, V. S. (2005) Kommunikativnoye povedeniye leoparda (Panthera pardus) v Tsentral'noy Azii i
na Kavkaze [Communicative behavior of the leopard (Panthera pardus) in Central Asia and the
Caucasus]. In: Povedeniye i povedencheskaya ekologiya mlekopitayushchikh. Materialy nauchnoy
konferentsii (4–8 oktyabrya 2005 g., Chernogolovka) [Behavior and behavioral ecology of
mammals. Proceedings of the Research Conference (4-8 October 2005, Chernogolovka)], pp. 191–
194. (in Russian)
Lukarevskiy, V. S. & Poyarkov, A.D. (2008) Sovremennoye sostoyaniye populyatsii irbisa (Uncia uncia,
Carnivora) v Rossii / The current state of the population of the snow leopard (Uncia uncia,
Carnivora) in Russia. Zoological Journal, 87(1), 114–121. (in Russian).
Lukarevskiy, V. S. & Umetdbekov, A. (2011) O sostoyanii nekotorykh gruppirovok irbisa v Kyrgyzstane.
Selevinia Journal, 162–167. (in Russian).
Lukarevskiy, V. S. & Purevsuren, S. (2011) Sostoyaniye gruppirovok irbisa Uncia uncia na
okhranyayemykh territoriyakh severo-zapadnoy Mongolii. Selevinia Journal, 167–174. (in Russian).
Lukarevskiy, V., Askerov, E. & Hazaryan, Gr. (2004). Condition of the Leopard Population in the Caucasus.
Beitrage zur Jagd & Wildforshung, 29, 303–319.
Lukarevskiy, V.S., Dalannast, M., Lukarevskiy, S.V. & Damdin E. (2019) Factors Determining the
Distribution and Status of the Snow Leopard Population (Panthera uncia) in Western Mongolia.
Animal and Veterinary Sciences, 7 (6), 127–132. doi: 10.11648/j.avs.20190706.12
Matyushkin, E. N. & Koshkarev, E. P. (1990) Sledy snezhnogo barsa [Traces of the snow leopard]. In:
Okhota i okhotn. khoz–vo [Hunting and game management], 2, 14–17 (in Russian).
Naumov, N. P. (1967) Struktura populyatsii i dinamika chislennosti nazemnykh pozvonochnykh)
[Population structure and population dynamics of terrestrial vertebrates]. Zoological Journal, 46(10),
1470–1486. (in Russian).
Naumov, N. P. (1973) Signal'nyye (biologicheskiye) polya i ikh znacheniye dlya dikikh zhivotnykh [Signal
(biological) fields and their significance for wild animals)]. Journal of General Biology (Moscow),
34(6), 808–817. (in Russian).
Naumov, N. P. (1977) Biologicheskie (signal'nye) polya i ikh znachenie v zhizni mlekopitayushchikh
[Biological (signal) fields and their significance in the life of mammals]. In: Uspekhi sovremennoi
teriologii [Advances in modern theriology]. Moscow, Nauka, pp. 93–110. (in Russian).
Ovsyannikov, N. G. & Badridze, Ya. K. (1989) Ponyatiye psikhologicheskogo komforta v interpretatsii
dvizhushchikh sil povedeniya [The concept of psychological comfort in the interpretation of the
driving forces of behavior]. Reports of the USSR Academy of Sciences, 4, 1015–1018. (in Russian).
Pikunov, D. G. & Korkishko,V. G. (1992) Leopard Dal'nego Vostoka [The Leopard in the Far East].
Moscow, Nauka, 192 pp. (in Russian).
Rozhnov, V. V., Zvychaynaya, E. Yu., Kuksin, A. N. & Poyarkov, A.D. (2011) Neinvazivnyy
molekulyarno-geneticheskiy analiz v issledovaniyakh ekologii irbisa: problemy i perspektivy [Non-
invasive molecular genetic analysis in the studies of snow leopard ecology: problems and prospects].
Ecology, 6, 403–408. (in Russian).
Wolff, J. O. (1997) Population regulation in mammals: an evolutionary perspective. Journal of Animal
Ecology, 66, 1–13.
Yudakov, A. G. & Nikolaev, I. G. (1987) Ekologiya amurskogo tigra [Ecology of the Amur Tiger]. Moscow,
Nauka, 153 pp. (in Russian).
Ecologica Montenegrina, 33, 2020, 17-28 27THE SPATIAL STRUCTURE OF А SNOW LEOPARD POPULATION IN EAST KYRGYZSTAN
Zavatsky, B. P. (2004) Snezhnyi bars, buryy medved' i volk Sayano-Shushenskogo zapovednika [The snow
leopard, the brown bear and the wolf in the Sayano-Shushensky Reserve]. Shushenskoe, 127 pp. (in
Russian).
Zvychaynaya, E. Yu., Rozhnov, V. V. & Poyarkov, A.D. (2011) Genotipirovaniye i pasportizatsiya osobey
redkikh vidov mlekopitayushchikh na primere irbisa (Uncia uncia) [Genotyping and certification of
individuals of rare species of mammals using the example of the snow leopard (Uncia uncia)]. In:
Tekhnologii sokhraneniya redkikh vidov zhivotnykh. Materialy nauchnoy konferentsii, 21–23
noyabrya 2011 g. [Technologies of conservation of rare animal species. Materials of the research
conference. 21–23 November 2011], Moscow, IPEE RAS, p. 16. (in Russian).
28You can also read

















































