A Legume TOR Protein Kinase Regulates Rhizobium Symbiosis and Is Essential for Infection and Nodule Development1 OPEN
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
A Legume TOR Protein Kinase Regulates Rhizobium
Symbiosis and Is Essential for Infection and
Nodule Development1[OPEN]
Kalpana Nanjareddy 2, Lourdes Blanco 2, Manoj-Kumar Arthikala, Xóchitl Alvarado-Affantranger,
Carmen Quinto, Federico Sánchez 3, and Miguel Lara*
Ciencias Agrogenómicas, Escuela Nacional de Estudios Superiores Unidad León-Universidad Nacional
Autónoma de México, Ciudad de Mexico C.P. 37684, Mexico (K.N., M.-K.A.); Instituto de Fisiología Celular,
Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacan, Ciudad de Mexico C.P. 62210,
Mexico (L.B.); Laboratorio Nacional de Microscopía Avanzada (X.A.-A.) and Instituto de Biotecnología
(C.Q., F.S.), Universidad Nacional Autónoma de Mexico, Ciudad de Mexico C.P. 62210, Mexico; and Instituto
de Biología, Universidad Nacional Autónoma de México, Ciudad Universitaria, Coyoacan, Ciudad de Mexico
C.P. 04510, Mexico (M.L.)
ORCID IDs: 0000-0002-7052-4120 (K.N.); 0000-0002-4535-6524 (M.-K.A.); 0000-0002-3068-2036 (X.A.-A.); 0000-0002-2961-4896 (F.S.);
0000-0002-5885-7950 (M.L.).
The target of rapamycin (TOR) protein kinase regulates metabolism, growth, and life span in yeast, animals, and plants in
coordination with nutrient status and environmental conditions. The nutrient-dependent nature of TOR functionality makes this
kinase a putative regulator of symbiotic associations involving nutrient acquisition. However, TOR’s role in these processes remains
to be understood. Here, we uncovered the role of TOR during the bean (Phaseolus vulgaris)-Rhizobium tropici (Rhizobium) symbiotic
interaction. TOR was expressed in all tested bean tissues, with higher transcript levels in the root meristems and senesced nodules.
We showed TOR promoter expression along the progressing infection thread and in the infected cells of mature nodules.
Posttranscriptional gene silencing of TOR using RNA interference (RNAi) showed that this gene is involved in lateral root
elongation and root cell organization and also alters the density, size, and number of root hairs. The suppression of TOR
transcripts also affected infection thread progression and associated cortical cell divisions, resulting in a drastic reduction of
nodule numbers. TOR-RNAi resulted in reduced reactive oxygen species accumulation and altered CyclinD1 and CyclinD3
expression, which are crucial factors for infection thread progression and nodule organogenesis. Enhanced expression of TOR-
regulated ATG genes in TOR-RNAi roots suggested that TOR plays a role in the recognition of Rhizobium as a symbiont. Together,
these data suggest that TOR plays a vital role in the establishment of root nodule symbiosis in the common bean.
In plants, growth occurs as a result of cell growth postembryonic growth is sustained by specialized zones
coupled with cell proliferation and cell expansion caused (shoot meristems and root apices) and depends upon the
by nutrient availability. Unlike animals, in plants, availability of nutrients that are present externally or
generated by various cellular processes. These local and
1
This work was supported by the Dirección General de Asuntos systemic nutrient and energy sources are integrated, and
del Personal Académico (grant no. IN219916 to M.L. and grant no. information is communicated to metabolic regulators
IA205115 to M.-K.A.) and the Consejo Nacional de Ciencia y Tecno- through signaling networks to control cellular processes.
lògia (grant no. 240614 to M.L.). The evolutionarily conserved target of rapamycin
2
These authors contributed equally to the article. (TOR) is a Ser/Thr kinase that acts as a master regulator
3
Deceased April 4, 2016. of metabolism and growth. In mammals, TOR exerts
* Address correspondence to mlara@unam.mx.
translational control of cell proliferation (Dowling et al.,
The author responsible for distribution of materials integral to the
findings presented in this article in accordance with the policy de- 2010), tumorigenesis, metastasis (Hsieh et al., 2012),
scribed in the Instructions for Authors (www.plantphysiol.org) is: and insulin signaling (Hsu et al., 2011). Loss-of-function
Miguel Lara (mlara@unam.mx). TOR mutations in Saccharomyces cerevisiae and Schizo-
K.N. conducted the experiments, analyzed the data, and wrote the saccharomyces pombe (Barbet et al., 1996; Weisman and
preliminary article; L.B. isolated and cloned the PvTOR gene and per- Choder, 2001), Drosophila melanogaster (Zhang et al.,
formed the PvTOR promoter analysis; M.-K.A. was involved in technical 2000), Caenorhabditis elegans (Long et al., 2002), and mice
support, optical microscopy, documentation, and RT-qPCR analysis;
X.A.-A. assisted with confocal and electron microscopy; C.Q. and F.S.
(Murakami et al., 2004) resulted in embryo lethality and
critically evaluated the data; M.L. conceived and coordinated the study other growth-associated problems.
and finalized the article; all authors read and approved the final article. Unlike other eukaryotes, knowledge about TOR in
[OPEN]
Articles can be viewed without a subscription. plants is limited due to a lack of molecular and bio-
www.plantphysiol.org/cgi/doi/10.1104/pp.16.00844 chemical assays for endogenous TOR kinase activity.
2002 Plant PhysiologyÒ, November 2016, Vol. 172, pp. 2002–2020, www.plantphysiol.org Ó 2016 American Society of Plant Biologists. All Rights Reserved.
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.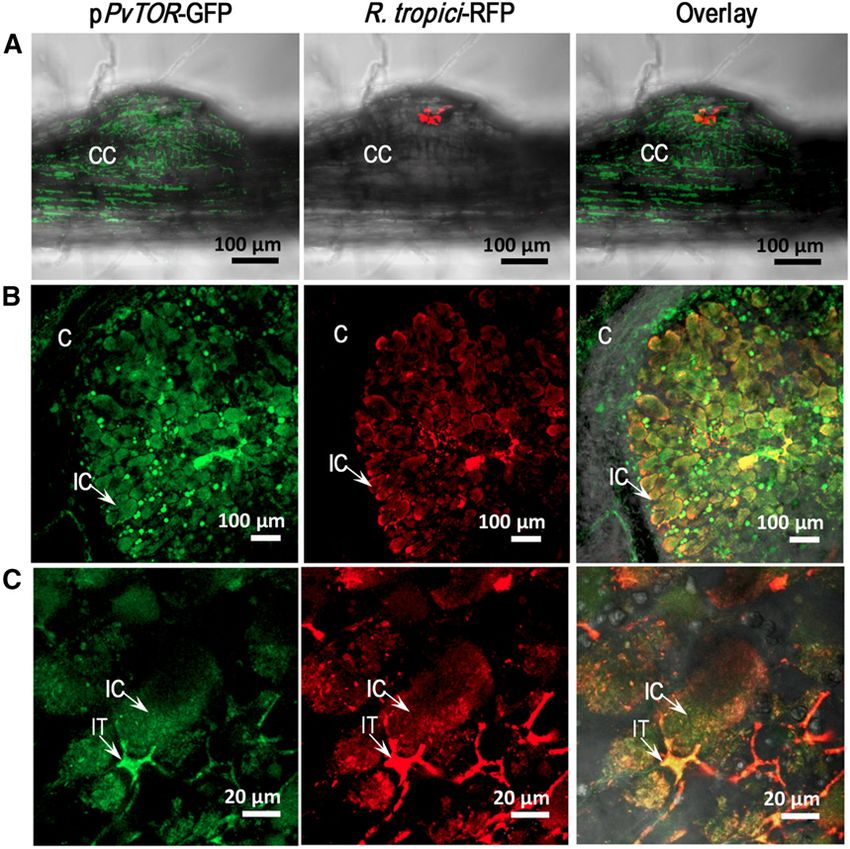
Role of TOR in the Legume-Rhizobium Symbiosis
The embryo lethality of TOR-null mutants in Arabi- endocytosis. Then, surrounded by a peribacteroid
dopsis (Arabidopsis thaliana) and the resistance of plants membrane, the rhizobia reduce atmospheric dini-
to rapamycin (Menand et al., 2002; Ren et al., 2011) have trogen to ammonium. The host plant accommodates
led to the conditional silencing of the gene using an the invading symbiont in the nodule, an organ formed
RNA interference (RNAi) approach (Deprost et al., de novo. The process of nodulation is regulated by the
2007; Xiong and Sheen, 2012; Caldana et al., 2013). The plant through cellular molecular responses. As a
repression of TOR transcript strongly retards postem- master regulator of cellular processes, the TOR sig-
bryonic developmental aspects of organ growth in naling pathway could play a pivotal role in the regu-
seedlings, including cotyledon expansion, true leaf de- lation of root nodule symbiosis. Nevertheless, TOR’s
velopment, petiole elongation, and primary and lateral role in symbiosis remains unknown.
root growth (Ren et al., 2012; Xiong and Sheen, 2012, In this work, we down-regulated TOR transcripts in
2015; Caldana et al., 2013; Xiong et al., 2013; Rexin et al., common bean (Phaseolus vulgaris) via posttranscrip-
2015). Intriguingly, overexpression of the full-length tional gene silencing using an RNAi approach. As a
TOR gene or the kinase domain alone also results in consequence, primary and lateral root growth were
developmental abnormalities, including severe shoot affected and root hair elongation was altered. Fur-
and inflorescence meristem defects, a compact archi- thermore, TOR transcriptional down-regulation also
tecture with short petioles, early senescence of rosette affected root nodule symbiosis in terms of both IT
leaves, delayed flowering, and the loss of apical domi- progression and nodule organogenesis. TOR-RNAi
nance (Ren et al., 2011). These observations indicate that roots frequently exhibited aborted IT progression in
TOR is a stringent regulator of postembryonic devel- root hair cells and an absence of infection-associated
opmental processes in plants. cortical cell divisions. Here, we demonstrate the in-
In plants, the TOR kinase acts as a check point that dispensable role of TOR in root nodule symbiosis in
couples nutrient status to the regulation of root system bean.
development and associated functions. TOR senses and
transduces Glc signals to activate root meristems by
orchestrating global transcriptional reprogramming RESULTS
(Xiong et al., 2013; Xiong and Sheen, 2014). Glc-TOR Identification and Sequence Analyses of the TOR Gene in
signaling activates S-phase entry in most cells in the the Common Bean
primary root meristem by phosphorylating the tran-
scription factor E2Fa. Based on the Arabidopsis TOR sequence (Deprost
Nevertheless, plant cell growth is tightly linked to the et al., 2007), a homolog was identified in the genome of
expansion of the cell wall. TOR kinase activity also is bean via TBLASTX analysis against the bean genomic
linked to the modification of cell walls for root hair data (Phytozome version 10.2; http://phytozome.jgi.
development, which is mediated by LRR-EXTENSIN1 doe.gov/pz/portal.html). The TOR (Phvul.002G049900)
(LRX1) and REPRESSOR OF LRX1 (ROL5; Leiber et al., gene is located on chromosome 2, and an analysis of the
2010). The role of ROL5 as a downstream component of gene model revealed the presence of 55 exons and
the TOR pathway indicates that TOR signaling is in- 54 introns, with a length of 7,827 bp for the predicted
volved in cell wall formation in Arabidopsis (Leiber mature transcript, an open reading frame of 7,407 bp, a 59
et al., 2010). untranslated region (UTR) of 181 bp, and a 39 UTR of
In addition to being the appendages that facilitate an- 239 bp (Fig. 1A). The corresponding gene, PvTOR
chorage and nutrient absorption, plant roots and root (henceforth called TOR), encodes a predicted protein of
hairs also serve as a gateway for plant-microbe interac- 2,469 amino acids (Fig. 1B). The amino acid similarity
tions. Leguminous plants have the privilege of establish- deduced from the GenBank nonredundant database
ing symbiotic interactions with nitrogen-fixing soil revealed the conserved nature of TOR across plant spe-
bacteria rhizobia, which are housed in specialized struc- cies (for instance, the sequence of bean shares 93%
tures on the roots that are called nodules. The symbiotic identity with Medicago truncatula and Glycine max and
program initiates with a chemical dialogue between the 80% identity with Arabidopsis, with an E value of 0.0;
rhizobia and legumes. This interaction leads to the rhi- Supplemental Fig. S1A.) Three-dimensional modeling of
zobial production of lipochitooligosaccharide (nod factor) TOR revealed the presence of FAT, FRB, and FATC do-
molecules, which induces a series of physiological re- mains (amino acids 1,360–2,869) as well as a large Ser/
sponses in root hairs, including calcium concentration Thr kinase domain (Fig. 1C). The highest sequence con-
oscillations (Ehrhardt et al., 1996), the induction of com- servation was found in the kinase domain, which con-
mon symbiotic genes, and root hair curling to entrap the tains 11 a-helices and six b-strands separated by short
bacteria (Oldroyd and Downie, 2008). Then, local cell wall loops (Fig. 1E; Supplemental Fig. S1B.) Furthermore, a
hydrolysis and the invagination of the plasma membrane phylogenetic analysis of bean TOR and its homologs
leads to the formation of infection threads (ITs), which from several plants (legume and nonlegume), animals,
traverse to the base of the root hair cell. The ITs extend fungi, and bacteria (Tamura et al., 2011) revealed a close
from the infection and focus toward the dividing cortical relationship between bean TOR and its homologs from
cells, where they branch and ramify. The rhizobia colo- other leguminous plants (M. truncatula, Lotus japonicus,
nizing the ITs are released into plant cortical cells via and G. max; Fig. 1D).
Plant Physiol. Vol. 172, 2016 2003
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.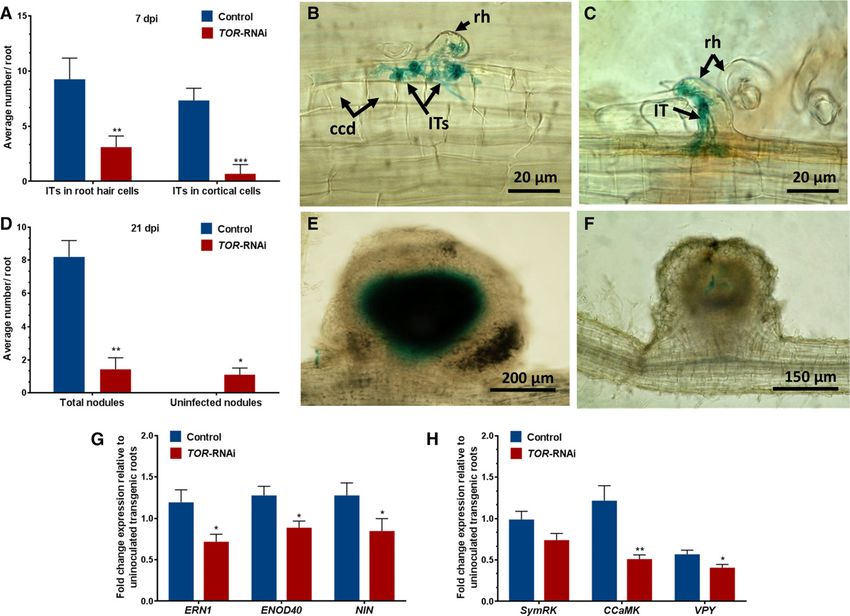
Nanjareddy et al.
Figure 1. Outline of the bean TOR gene, protein structure, and phylogenetic tree. A, Schematic representation of the TOR gene
structure showing UTRs, 55 exons, and 54 introns on chromosome 2 of bean. B, The TOR protein has typical Ser/Thr protein
kinase (phosphatidylinositol 3- and 4-kinase; red), FAT (brown), FRB (rapamycin-binding domain; gray), and FATC (black) do-
mains. aa, Amino acids. C, Three-dimensional modeling of the TOR kinase domain. D, Phylogenetic analysis was performed in
the MEGA5 program (Tamura et al., 2011) using 17 TOR amino acid sequences from G. max, bean, Vitis vinefera, Arabidopsis, M.
truncatula, Oryza sativa, Zea mays, Chlamydomonas reinhardtii, Saccharomyces cerevisiae, Homo sapiens, Mus musculus,
D. melanogaster, and Bombyx mori, which are available from the GenBank database. The evolutionary history was inferred using
2004 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.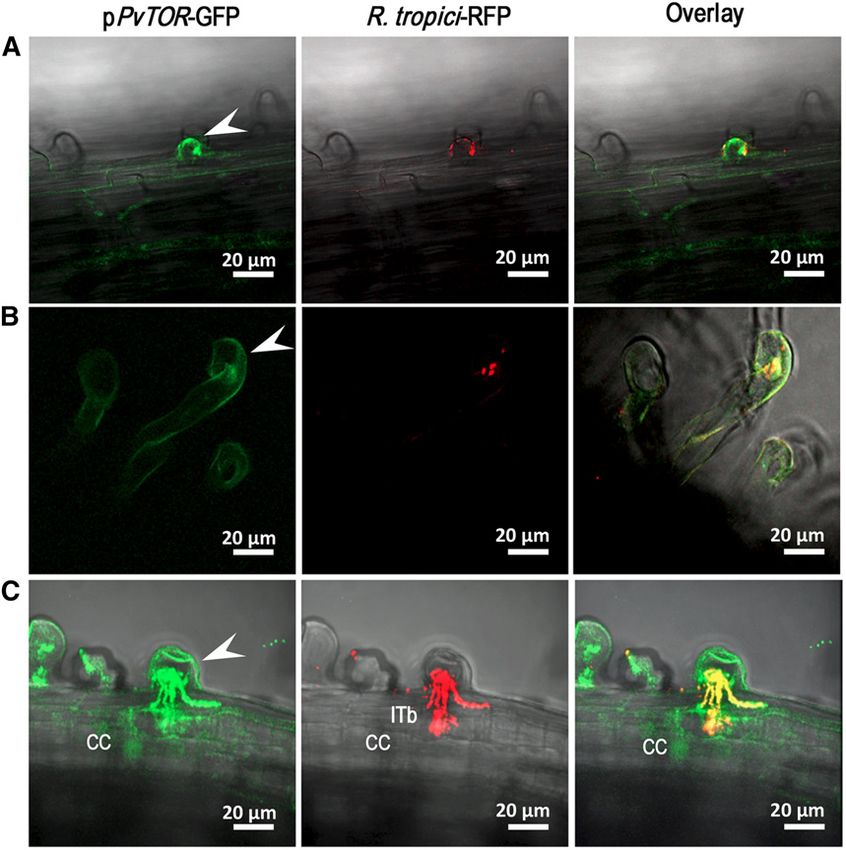
Role of TOR in the Legume-Rhizobium Symbiosis
Bean TOR Expression Is Associated with Rhizobial IT (RT-qPCR) for the TOR transcript in wild-type bean
Progression and Nodule Development tissues and observed that TOR was expressed in all
tested tissues of the plant. Within the root, stronger
When analyzed with PlantPan (http://plantpan. TOR transcription was observed in the root meristem
mbc.nctu.edu.tw/seq_analysis.php), the regulatory (root tip) than in the zone of elongation and matu-
sequence region of bean TOR was found to contain ration. In symbiotic root nodule, TOR transcript was
seven nodule-specific regulatory motifs within a 1-kb found to be higher than in the root elongation and
region. To assess the spatiotemporal expression pat- maturation zones. TOR was differentially expressed
tern of bean TOR, a 1,000-bp region upstream of the among the other vegetative (hypocotyl, vegeta-
translation initiation codon was used to generate a tive bud, stem, petiole, and leaf) and reproductive
transcriptional fusion to the chimeric reporter GUS- (flower and pod) organs. Higher transcript levels
enhanced GFP (pPvTOR::GUS-GFP). The pPvTOR:: were detected in the leaf and hypocotyl than in the
GUS-GFP reporter construct was transfected into bean flower and pod (Fig. 5A). Based on RT-qPCR, we
via hairy root transformation. Transcriptional activa- concluded that TOR is expressed in most tissues of
tion of the reporter gene in the transgenic hairy roots of bean, but the variation in transcript abundance in-
bean was monitored with and without rhizobial inoc- dicated that every tissue has its own TOR activity
ulation. In the uninoculated roots, strong GUS expres- profile.
sion was observed at the root tip (Fig. 2A). Promoter
activity also was found at the site of the lateral root
primordium (Fig. 2B) and the emerging lateral roots, TOR Expression Is Up-Regulated during Nodulation
and that activity subsequently became confined to the
root tip as the lateral roots emerged from the primary Next, the transcript abundance of TOR was deter-
root (Fig. 2C). TOR promoter expression in the root mined in wild-type bean root tissues inoculated with R.
hairs was localized to the tips of nascent root hairs and tropici via RT-qPCR. We observed that, upon inocula-
in a longitudinal distribution along the fully developed tion, TOR transcript abundance was induced more
root hairs (Fig. 2, E and F). strongly in nodules than in nodule-detached roots
To test promoter activation upon inoculation with (Fig. 5B). Starting from the early stage of infection
Rhizobium, hairy roots were inoculated with Rhizobium (24 h post inoculation [hpi]), the TOR transcript in-
tropici, and GUS activity was observed at periodic in- creased steadily until 48 hpi and then decreased at
tervals postinoculation. At 2 d post inoculation (dpi), 96 hpi in Rhizobium-inoculated roots (Supplemental
TOR promoter expression was observed in the tip of the Fig. S2.) In contrast, transcript induction was signif-
emerging root hair, notably around the binding site of icantly stronger in nodules from 5 dpi (120 hpi) to
the rhizobia (Fig. 3A). TOR promoter expression was 14 dpi and then decreased as the nodules matured at
found to be restricted to the growing ITs (Fig. 3, B and C). 21 dpi (Fig. 5B; Supplemental Fig. S2.) Interestingly,
Furthermore, promoter activity also was observed dur- the TOR transcript levels increased drastically with
ing nodule organogenesis in cortical cells undergoing the the onset of nodule senescence. In contrast, weak in-
initial cell divisions that form the nodule primordium duction of TOR was detected in nodule-detached
(Fig. 4A). In young and mature nodules, the promoter roots at all time points (Fig. 5B). Together, these re-
activity was restricted to infected cells. Promoter ex- sults show that TOR induction was associated with
pression also was detected in the intercellular ITs in three important stages: early infection, nodule de-
mature nodules (Fig. 4, B and C). The spatiotemporal velopment, and senescence.
expression analysis of the TOR promoter suggests that
this gene plays an important role in IT progression and
bacterial localization in the growing nodule. Down-Regulation of TOR Expression in Composite Plants
of Bean
TOR Is Differentially Expressed in Bean Tissues To functionally characterize TOR during symbio-
sis, we took advantage of the bean root transforma-
To confirm the observed spatial expression patterns, tion system that uses Agrobacterium rhizogenes. An
we performed quantitative reverse transcription-PCR RNAi construct harboring a nonconserved region of
Figure 1. (Continued.)
the neighbor-joining method (Saitou and Nei, 1987). The bootstrap consensus tree inferred from 1,000 replicates is taken to
represent the evolutionary history of the analyzed taxa (Felsenstein, 1985). Branches corresponding to partitions that were
reproduced in less than 50% of bootstrap replicates are collapsed. The percentage of replicate trees in which the associated taxa
clustered together in the bootstrap test (1,000 replicates) is shown next to the branches (Felsenstein, 1985). The evolutionary
distances were computed using the proportion-distance method (Nei and Kumar, 2000) and are presented as the number of amino
acid differences per site. All positions containing gaps and missing data were eliminated. A total of 2,190 positions were included
in the final data set. E, Secondary structure prediction using Swiss model (http://swissmodel.expasy.org) shows that the TOR
kinase domain is composed of 15 a-helices (green bars) and six b-sheets (blue arrows) joined by short coils (red lines).
Plant Physiol. Vol. 172, 2016 2005
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.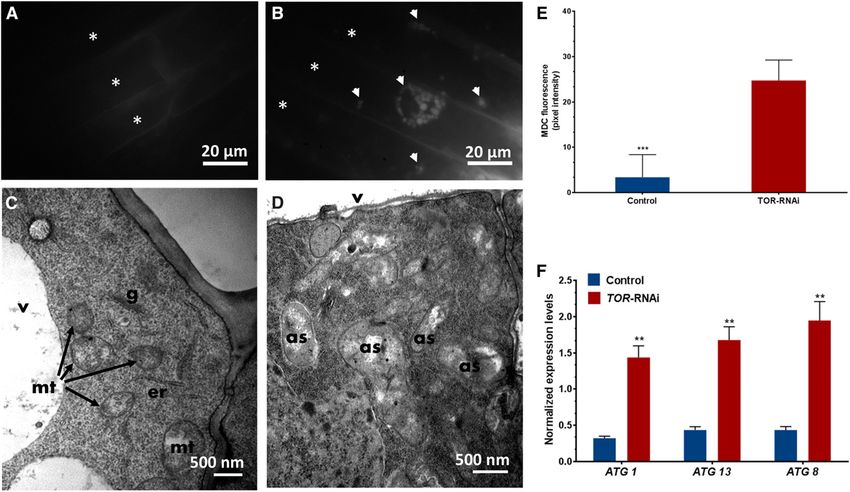
Nanjareddy et al.
Figure 2. Promoter analysis of PvTOR in
transgenic bean roots. The spatial pattern of
TOR expression was revealed by a pro-
moter::GUS-GFP construct in transgenic
hairy roots incubated with GUS as a sub-
strate. A to D, Optical microscopy images of
PvTOR::GUS principal root (A), lateral root
primordia (B), lateral root growth (C), and
fully developed lateral root (D). Confocal
microscopy images of PvTOR::GFP pro-
moter activity in root hair cells: an emerging
root hair (E) and a fully developed root hair (F).
the C terminus and 39 UTR of TOR (Fig. 1A; pTdT- (henceforth 35S-TOR-RNAi) compared with transgenic
35S-TOR-RNAi) and an empty vector (pTdT-35S- control roots containing the empty vector (henceforth
RNAi) were expressed individually in hairy roots of control roots; Fig. 6A). However, no change in the tran-
the composite plants. Considering the ubiquitous script abundance of TOR was observed in transgenic
expression of TOR in all tested tissues of bean, to roots expressing pTdT-pNIN-TOR-RNAi (henceforth
address the pleiotropic regulation of plant develop- pNIN-TOR-RNAi).
ment by TOR, we generated an RNAi construct where Furthermore, pNIN-TOR-RNAi-expressing trans-
expression was driven by a symbiosis-specific nod- genic roots were inoculated with R. tropici and TOR
ule inception (PvNIN) promoter (pTdT-pNIN-TOR- transcript abundance was analyzed at 7 dpi. TOR
RNAi), and pTdT-pNIN-RNAi vector was used as the transcripts in such roots were found to be reduced by
control. An RT-qPCR analysis of hairy roots 10 d post 40% to 50% (Supplemental Fig. S3.) Our results indi-
emergence (dpe) confirmed the reduction of TOR cated that the 35S-TOR-RNAi constructs specifically
mRNA levels, with levels ranging from 70% to 80% down-regulated TOR transcript levels in transgenic
in transgenic roots expressing pTdT-35S-TOR-RNAi roots. However, NIN promoter-driven RNAi roots
Figure 3. Promoter analysis of TOR in transgenic
bean root hair cells inoculated with Rhizobium.
TOR promoter expression is shown in R. tropici-
infected root hair cells at 2 dpi (A), 4 dpi (B), and
6 dpi (C). The left column shows TOR expression
revealed by promoter::GUS-GFP, the middle col-
umn shows red fluorescence emitted by R. tropici
CIAT899 Ds-Red, and the right column shows
overlay images. CC, Cortical cells; ITb, IT branch-
ing.
2006 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.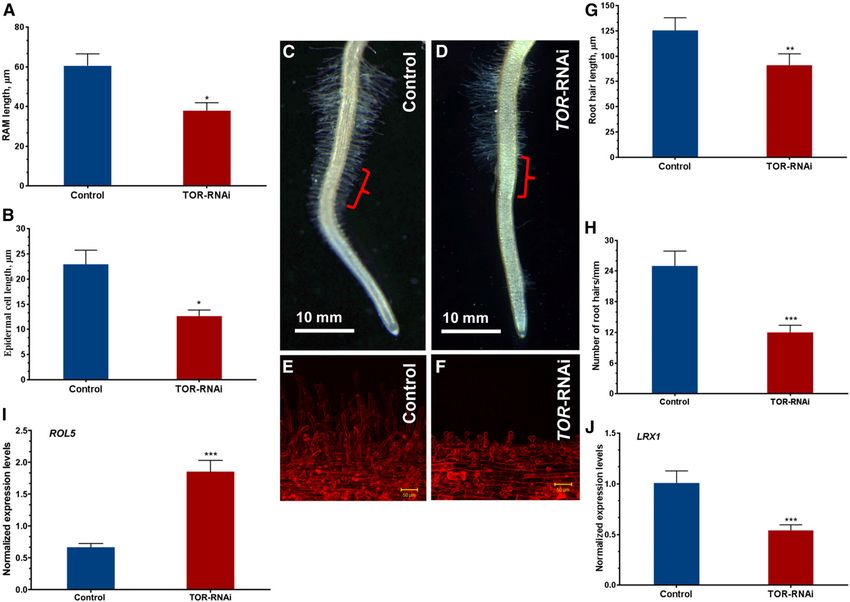
Role of TOR in the Legume-Rhizobium Symbiosis
Figure 4. Bean TOR promoter expression and
Rhizobium localization in transgenic bean nod-
ules. A and B, Confocal microscopy images of a
nodule meristem at 8 dpi (A) and a central infected
zone of a mature nodule at 14 dpi (B). C, Closer
views of a nodule infected zone. The left column
shows PvTOR expression revealed by promoter::
GUS-GFP, the middle column shows red fluores-
cence emitted by R. tropici CIAT899 Ds-Red, and
the right column shows overlay images. C, Inner
cortex; CC, cortical cells; IC, infected cells.
were found to down-regulate the TOR transcripts only plants (Fig. 6B) and exhibited a significant increase in the
in the presence of Rhizobium in bean. root-shoot ratio (Fig. 7A). The primary root length and the
lateral root density of 35S-TOR-RNAi plants were signif-
icantly smaller than those of the control (Fig. 7, B and C).
Interfering with TOR Modifies Root and Root Hair
We then examined root cell size and organization using
Morphology in Bean
confocal microscopy. Similar to the knockout mutant
In this study, plant biomass was analyzed in the (AtTOR of Arabidopsis), the root apical meristem (RAM)
transgenic composite plants at 10 dpe. The 35S-TOR- of the 35S-TOR-RNAi plants also showed significant re-
RNAi composite plants were smaller than the control ductions (37.9 6 4.3 mm) in cell size and cell numbers
Figure 5. Expression patterns of the TOR gene in wild-type bean tissues by RT-qPCR. A, Expression profiles of the TOR gene in
different vegetative and reproductive organs. Transcript accumulation was normalized based on the expression of Elongation
factor 1 alpha (EF1a) and Insulin degrading enzyme (IDE) which were used as reference genes. B, Transcript abundance of TOR in
R. tropici-inoculated roots (1 dpi), nodules (7, 14, 21, and 28 dpi), and nodule-detached roots at different time points. The
presented data are averages of three biological replicates (n . 9) and represent means 6 SE.
Plant Physiol. Vol. 172, 2016 2007
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.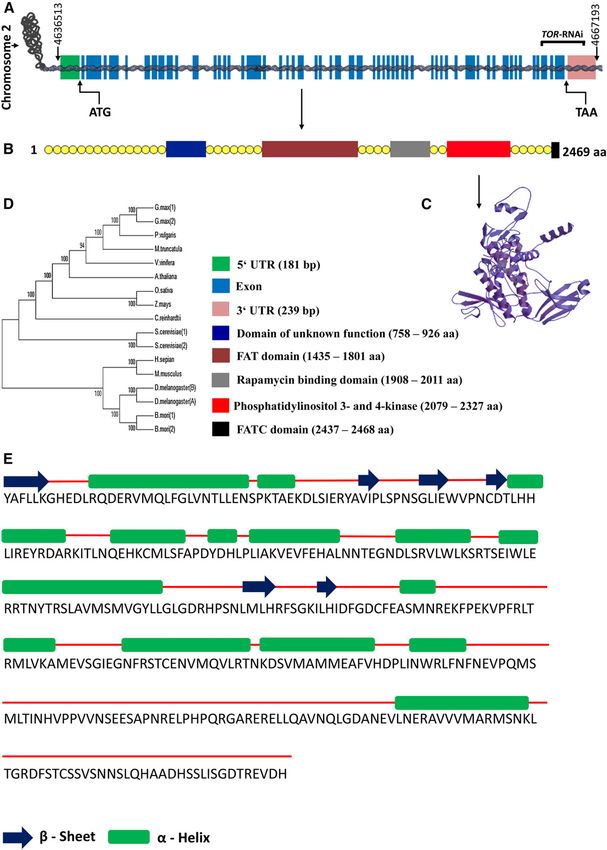
Nanjareddy et al.
Figure 6. Phenotype of TOR-RNAi composite plants and RT-qPCR analysis of the TOR transcript. A, Results of TOR down-regulation in
uninoculated TOR-RNAi transgenic roots relative to transgenic control (empty vector) roots. The numbers in parentheses indicate the
composite bean plants used in this study. Transcript accumulation was normalized based on the expression of Ef1a and IDE, which were
used as reference genes. Statistical significance was determined using an unpaired two-tailed Student’s t test (***, P , 0.001), and the
data are presented as means 6 SD. B, Representative images of bean transgenic control and TOR-RNAi composite plants at 10 dpe.
relative to the control RAM (60.64 6 6.1 mm; Fig. 8A). TOR Down-Regulation Is Associated with Altered
Furthermore, the root epidermal cell size of 35S-TOR- Reactive Oxygen Species Production
RNAi plants was reduced by approximately 45% in
comparison with the control roots (Fig. 8B). As TOR is a central player in reactive oxygen species
We also found that, as a consequence of TOR down- (ROS) sensing and signaling (Niles et al., 2014) and
regulation, the root hair cells grew shorter and less genes involved in ROS generation (e.g. respiratory
densely than the control roots at 10 dpe (Fig. 8, C–F and burst oxidase homolog [Rboh] genes) are essential for
H). Interestingly, in the susceptible zone (the zone in root development (Torres et al., 2002; Montiel et al.,
which root nodules typically form), the lengths of the 2013), we assessed ROS production in the TOR-down-
regulated roots. At 15 dpe, the transgenic roots were
root hair cells were reduced drastically (Fig. 8G).
stained using the fluorescent probe Chloromethyl de-
Moreover, an RT-qPCR analysis of 35S-TOR-RNAi
rivative of 29, 79-dichlorodihydrofluorescein diacetate
roots showed a significant reduction of transcript ac-
(CM-H2DCFDA) (Duan et al., 2010) and were examined
cumulation for the gene LRX1 and an enhancement of
with a confocal microscope. The CM-H2DCFDA-
transcript accumulation for the gene ROL5; both of
treated TOR roots exhibited weak fluorescence (Fig. 9,
these genes are involved in root hair cell wall formation.
E and F) in comparison with the control roots (Fig. 9, B
LRX1, which is a cell wall chimeric Leu-rich repeat/
and C). A comparative analysis of the relative fluores-
extensin protein, and ROL5, a repressor of LRX1, exhibited
cence intensity of TOR versus control transgenic roots
a 200% increase and a 46% reduction in transcript levels,
is shown in Figure 9G. Furthermore, the transcripts of
respectively, in TOR-silenced roots compared with control
the RbohB gene, which is involved in root growth and
roots (Fig. 8, I and J). The overall anatomy of the root and
lateral root development (Montiel et al., 2013), were
root hair did not differ between 35S-TOR-RNAi and
reduced significantly in TOR-down-regulated roots
control plants (data not shown). relative to controls (Fig. 9H). Similarly, the transcripts
We then quantified the transcripts of root growth fac- of antioxidant metalloproteins, such as SOD and CAT
tors (RGFs) and the UPBEAT1 transcription factor (UPB1), (Fridovich, 1998), also were reduced, by 32% and 67%,
which are the key TOR integrated regulatory genes for the respectively, upon TOR down-regulation compared
local promotion of the root meristem (Xiong et al., 2013). with control roots (Fig. 9H). Together, these results
Overexpression of the transcription factor UPB1 is known imply that the reduction of RbohB and antioxidant
to inhibit root meristem expansion through redox control transcripts reduces ROS production in 35S-TOR-RNAi
(Tsukagoshi et al., 2010). We found increased transcript roots.
abundance of UPB1 in 35S-TOR-RNAi plants relative to
controls (Fig. 7D). Other genes, such as the root meristem
growth factors RGF6 and RGF9 and the S-assimilation TOR Is a Negative Regulator of Autophagy in Bean
genes glutathione synthetase, adenosine-59-phospho-
sulfate kinase2, and sulfite reductase, were repressed TOR has been identified as a negative regulator
upon TOR down-regulation. Taken together, these ob- of autophagy in yeast, mammals, and Arabidopsis
servations indicate that TOR is required for cell division (Noda and Ohsumi, 1998; Pattingre et al., 2008; Liu and
and elongation in bean. Bassham, 2010). Here, the impact of TOR down-regulation
2008 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.
Role of TOR in the Legume-Rhizobium Symbiosis
Figure 7. Evaluation of TOR-RNAi composite plant growth and transcripts of root meristem regulatory genes. A, Root-shoot (R/S)
ratio of uninoculated composite plants at 10 dpe. Root growth parameters were assessed at 10 dpe in TOR-RNAi composite plants
and compared with the transgenic control roots. B and C, Primary root length (B) and lateral root density (C). D, RT-qPCR analysis
to measure the transcript levels of root meristem regulatory genes, such as the transcription factor UPB1, glutathione synthetase
(GSS), RGF6, RGF9, adenosine-59-phosphosulfate kinase2 (APK2), and sulfite reductase (SIR), in control and TOR-RNAi trans-
genic roots. The statistical significance of differences between transgenic control and TOR-RNAi roots was determined using an
unpaired two-tailed Student’s t test (**, P , 0.01; and ***, P , 0.001). The data are presented as averages of three biological
replicates (n . 27) for B to D and represent means 6 SE.
on basal autophagy was analyzed in 10-dpe trans- are involved in phagosome formation, and since ATG8
genic roots stained with the fluorescent dye mono- is involved in autophagosomal membrane formation,
dansylcadaverine (MDC), which selectively labels we performed RT-qPCR for these genes and observed
autophagosomes (Biederbick et al., 1995). In vivo that the down-regulation of TOR significantly induced
imaging of 35S-TOR-RNAi root cells using a UV ATG gene expression (Fig. 10F). These results suggest
fluorescence microscope showed rapidly moving that TOR negatively regulates autophagy genes in
fluorescent puncta in the cytoplasm (Fig. 10B). In bean roots.
contrast, no fluorescent puncta were observed in the
root cells of control plants (Fig. 10A). However, after
carbon deprivation (in composite plants that were The Down-Regulation of TOR Impairs IT Progression in
dark treated for 48 h), the control root cells showed Root Hair Cells
fewer fluorescent puncta than the 35S-TOR-RNAi
plants, which had denser and larger fluorescent To assess the role of TOR in nodulation, transgenic
puncta (Supplemental Fig. S4, A and B.) A compara- hairy roots expressing 35S-TOR-RNAi were inoculated
tive analysis of the relative fluorescence intensity of with R. tropici expressing a GUS marker (Vinuesa et al.,
the puncta in 35S-TOR-RNAi versus control transgenic 2003). Light microscopic observations revealed that the
roots is shown in Figure 10E and Supplemental Figure Rhizobium-infected root hair cells of both control and
S2C. Ultrastructural observations of transverse sec- 35S-TOR-RNAi plants show the typical root hair curl-
tions of the root tips revealed an accumulation of ing and rhizobial microcolonies of wild-type roots. In
double membrane structures (autophagosomes) in contrast, the total number of infection events (i.e. ITs in
TOR-down-regulated plants (Fig. 10D). However, the root hair cells and in dividing cortical cells) was re-
cytoplasm of the control root tip cells showed abun- duced significantly in 35S-TOR-RNAi roots compared
dant mitochondria, Golgi apparatus, and endoplasmic with control roots (Fig. 11A). When examined closely,
reticulum but was devoid of autophagosomes (Fig. 10C). the IT progression was arrested within the root hair
As ATG1 and ATG13 are phosphorylated by TOR and cells of 35S-TOR-RNAi plants, and these ITs were
Plant Physiol. Vol. 172, 2016 2009
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.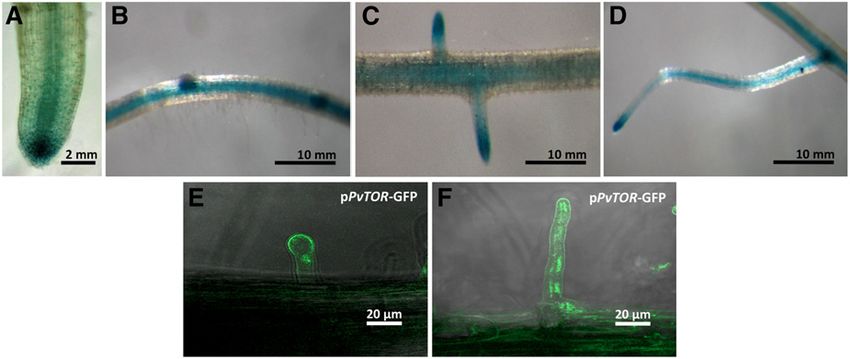
Nanjareddy et al.
Figure 8. Phenotypic and expression analyses of root hair cell wall formation genes in TOR-RNAi roots. A and B, Quantitative
analysis of the RAM (A) and epidermal cell length (B) of transgenic control and TOR-RNAi roots at 10 dpe. C and D, Repre-
sentative images showing root hair density on the primary roots of transgenic control (C) and TOR-RNAi (D) plants. The red
brackets indicate the susceptible zones for nodulation. E and F, Confocal images showing the length of root hairs at the susceptible
zones of transgenic control roots (E) and TOR-RNAi roots (F). G and H, Quantitative analysis of the average root hair length (G)
and root hair numbers (H) in transgenic control and TOR-RNAi roots. I and J, RT-qPCR analyses of the root hair cell wall formation
genes ROL5 (I) and LRX1 (J) in transgenic control and TOR-RNAi roots. Transcript accumulation was normalized based on the
expression of Ef1a and IDE, which were used as reference genes. The data are presented as averages of three biological replicates
(n . 21 for A, B, G, and H and n . 9 for I and J). The statistical significance of differences between transgenic control and TOR-
RNAi roots was determined using an unpaired two-tailed Student’s t test (*, P , 0.05; **, P , 0.01; and ***, P , 0.001). Data
represent means 6 SE.
broader (Fig. 11C) than the ITs found in the root hair cortical cell divisions, and nodule infection (Supplemental
cells of the controls (Fig. 11B; Supplemental Fig. S3, A Fig. S6, A and C). These results suggest that both
and B.) However, in contrast to the controls, cortical cell constitutive (35S-TOR-RNAi) and symbiosis-specific
divisions were not associated with the majority of (pNIN-TOR-RNAi) promoter-driven TOR down-
aborted ITs in 35S-TOR-RNAi roots (Fig. 11A). Never- regulation similarly impairs IT progression; hence,
theless, in 35S-TOR-RNAi roots, the ITs occasion- further analysis was carried out in 35S-TOR-RNAi
ally spread beyond the epidermal cells into dividing roots alone.
cortical cells, resulting in nodule bumps (Fig. 11F; Because ROS accumulation is the primary response
Supplemental Fig. S5D); however, the majority of these to nod factor perception (Cárdenas et al., 2008), we
nodules were uninfected (Fig. 11D). Furthermore, analyzed the ROS response during the Rhizobium in-
pNIN-TOR-RNAi-expressing hairy roots also demon- teraction. The CM-H2DCFDA-stained nodule primor-
strated a similar phenotype of IT abortion within dium and young nodules showed reduced ROS
the root hair cells and uninfected nodule bumps production during nodule development in 35S-TOR-
(Supplemental Fig. S6, B and D); however, pNIN-RNAi RNAi roots (Supplemental Fig. S7, B and C) compared
empty vector controls showed typical IT progression, with controls (Supplemental Fig. S4A.) Furthermore,
2010 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.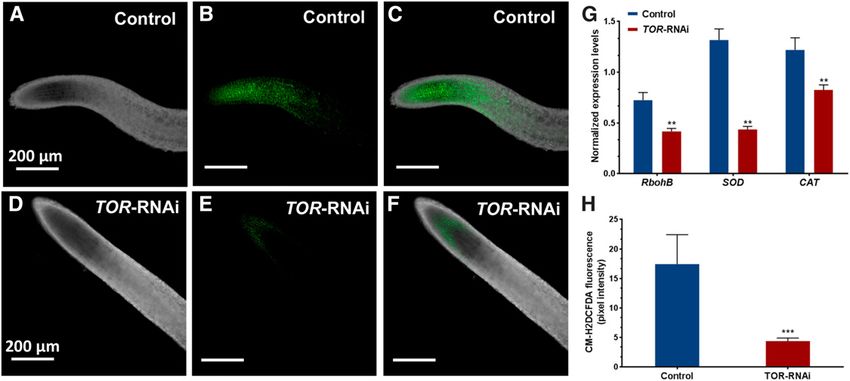
Role of TOR in the Legume-Rhizobium Symbiosis
Figure 9. Analysis of ROS production in bean TOR-RNAi roots. Transgenic control and TOR-RNAi composite plants were grown
under identical conditions and treated with CM-H2DCFDA to monitor ROS levels. A to C, Confocal microscopy images showing
similar root zones under transmitted light (A), fluorescence light (B), and merged images of A and B (C) for transgenic control roots.
D to F, Similar transmitted light (D), fluorescence light (E), and merged images of D and E (F) are shown for TOR-RNAi roots. All
images were acquired using the image-acquisition conditions of the control. G, RT-qPCR analysis showing a decrease in the
expression levels of RbohB, SOD, and CAT in TOR-RNAi transgenic roots relative to transgenic control roots. The statistical
significance of differences between transgenic control and TOR-RNAi roots was determined using an unpaired two-tailed Stu-
dent’s t test (**, P , 0.01). The data are presented as averages of three biological replicates (n . 9) and represent means 6 SE. H,
ROS level quantification using pixel intensity. The statistical significance of differences between transgenic control and TOR-
RNAi roots was determined using an unpaired two-tailed Student’s t test (***, P , 0.001). The data represent means 6 SD of ROS
intensity measured from n . 18 roots.
transcript accumulation of the ROS-related early TOR Down-Regulation Results in Altered
nodulin gene Rhizobium-induced peroxidase 1 (RIP1) Nodule Morphology
(Ramu et al., 2002) was reduced significantly in 35S-
TOR-RNAi roots relative to controls (Supplemental We next assessed the physiological characteristics
Fig. S7D.) of sparse root nodules in TOR-down-regulated plants
To determine whether this decreased number of in comparison with controls. At 21 dpi, TOR-down-
infection events is associated with changes in the regulated roots showed an 80% reduction in nodule
expression of genes involved in early nodulin sig- numbers in comparison with control roots (Fig. 11D).
naling, we used RT-qPCR to analyze the expression The distribution of nodule bumps/nodules along the
profiles of the ERN1, ENOD40, and NIN transcripts transgenic TOR-RNAi roots was significantly lower
in Rhizobium-inoculated transgenic roots (Fig. 11G). (approximately two nodules per root) than in the
Transcriptional activation of ERN1 (Middleton et al., transgenic control roots (approximately nine nodules
2007) and NIN (Madsen et al., 2010) regulates the per root). Furthermore, the 35S-TOR-RNAi nodules
early steps of nodulation, such as nod factor-induced were small and white, in contrast to the round control
gene expression and IT formation; in contrast, nodules, which were pink as a result of the presence of
ENOD40 is up-regulated during cortical cell division the leghemoglobin protein, which is required for
and nodule development (Stougaard, 2000). After oxygen-sensitive nitrogenase activity. An acetylene re-
rhizobia inoculation, we observed that the ERN1, duction assay of the nodules further indicated the loss
ENOD40, and NIN transcripts decreased in 35S-TOR- of nitrogen-fixing ability in the 35S-TOR-RNAi lines
RNAi roots relative to controls (Fig. 11G). Similarly, (Fig. 12A).
the transcripts of common symbiotic pathway (CSP) We used optical microscopy and transmission elec-
genes, such as Symbiosis Receptor Kinase (SymRK), tron microscopy to examine the structural and ultra-
Calcium and Calmodulin dependent protein Kinase structural characteristics of the sparse nodules formed
(CCamK), Vapyrin (VPY), and Remorin, which are by the 35S-TOR-RNAi lines. The semithin sections of
essential for successful rhizobial infection, also were the 35S-TOR-RNAi nodules exhibited a cell organiza-
decreased in rhizobia-inoculated 35S-TOR-RNAi tion similar to that of control nodules. In both control
roots in comparison with control roots (Fig. 11H). and 35S-TOR-RNAi nodules, an outer cortex surrounded
Taken together, our observations indicate that TOR an inner cortex containing the nodule vascular bundles
is required for the progression of ITs and nodule and the central tissue, which contained infected and
organogenesis. uninfected cells (Fig. 12, C and D). However, the infected
Plant Physiol. Vol. 172, 2016 2011
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Nanjareddy et al.
Figure 10. Analysis of autophagosomes and expression profile of autophagy genes in TOR-RNAi roots. A and B, Visualization of
autophagosomes in 10-dpe roots using an MDC fluorescence probe: transgenic control (A) and TOR-RNAi (B) showing fluo-
rescent puncta (white arrowheads). Asterisks indicate cells. C and D, Transmission electron microscopy images of roots showing
different cell organelles in the transgenic control (C) and autophagosomes (as) in TOR-RNAi (D) samples. er, Endoplasmic re-
ticulum; g, Golgi apparatus; mt, mitochondria; v, vacuole. E, MDC fluorescence quantification using pixel intensity. The statistical
significance of differences between transgenic control and TOR-RNAi roots was determined using an unpaired two-tailed Stu-
dent’s t test (***, P , 0.001). The data represent means 6 SD of the ROS intensity measured from n . 10 roots. F, RT-qPCR analysis
showing an increase in the expression levels of ATG1, ATG13, and ATG8 in TOR-RNAi transgenic roots relative to transgenic
control roots. The statistical significance of differences between transgenic control and TOR-RNAi roots was determined using an
unpaired two-tailed Student’s t test (**, P , 0.01). The data are presented as averages of three biological replicates (n . 9) and
represent means 6 SE.
cells in the 35S-TOR-RNAi nodules showed an abundant, Interestingly, the 35S-TOR-RNAi nodules contained
clear, multivacuolated cytoplasm (Figs. 12, F and H, and abundant cytoplasmic vesicles and autophagosomes
13B) in comparison with controls (Figs. 12E and 13A). (Fig. 13H). Furthermore, in contrast to the control nod-
Furthermore, a drastic reduction in the size of infected cells ules, amyloplasts were absent from the 35S-TOR-RNAi
was seen in 35S-TOR-RNAi nodules relative to controls nodules (Supplemental Fig. S8, C and D.)
(Fig. 12, G and H). To confirm this finding, the size of the
infected cells was measured; this quantitative analysis
is depicted in Figure 12B. Ultrastructural observations TOR Down-Regulation Affected the Expression of Cyclin
revealed that the 35S-TOR-RNAi lines hosted fewer Genes in Rhizobium-Inoculated Roots
symbiosomes per infected cell, and the majority of these
symbiosomes contained one bacteroid (Fig. 13, C–F). In mammals, TOR controls G1/S-phase progression
Some symbiosomes had enlarged perisymbiotic mem- via the transcriptional regulation of G1 cyclins (D-type
branes but still contained fewer bacteria. Intriguingly, the and E-type cyclins) through the downstream effectors
intercellular ITs in 35S-TOR-RNAi nodules also showed a 4E-BP and S6K1 (Fingar et al., 2004). It was reported re-
sparse distribution of bacteria (Fig. 13J) compared with cently that the Arabidopsis S6K protein is associated with
the controls (Fig. 13I). The presence of a degraded peri- the cell cycle regulation of root meristems (Henriques
bacteroid membrane was a significant observation in the et al., 2010; Shin et al., 2012; Rexin et al., 2015).
35S-TOR-RNAi nodules (Fig. 13H). In this study, the 35S-TOR-RNAi roots showed a re-
The functionally active 21-d-old control nodules had duction in cellular organization and RAM size, which
fibrous cytosols and were devoid of double-membrane resulted in shorter roots (Fig. 8A). Thus, we investigated
autophagic structures, or autophagosomes (Fig. 13G). whether TOR affects cell cycle regulation during the course
2012 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Role of TOR in the Legume-Rhizobium Symbiosis
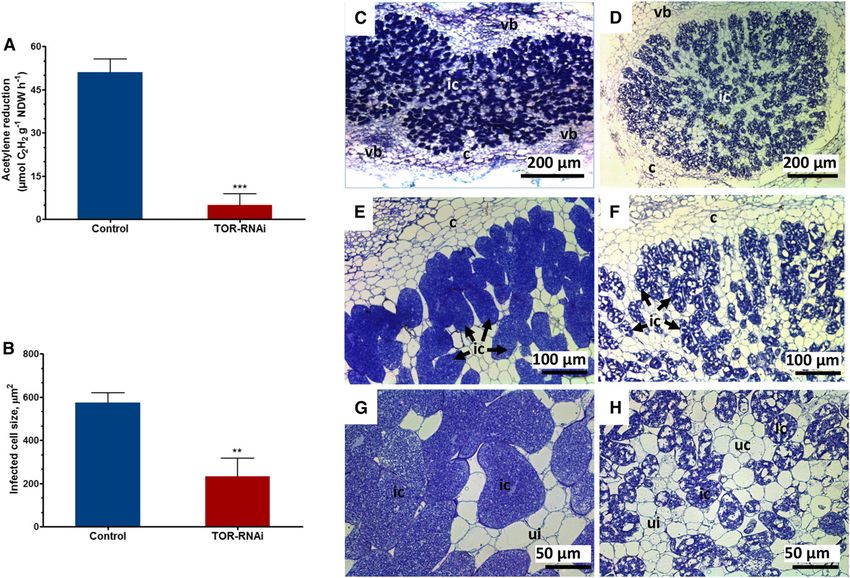
Figure 11. IT phenotype and expression profile of IT-associated genes. Seven days after inoculation with R. tropici expressing a
GUS marker, GUS-stained roots were observed using a light microscope. A, Average number of ITs found in root hair cells and
dividing cortical cells of transgenic control and TOR-RNAi roots. B, Representative image showing normal IT progression, with
branching and cortical cell divisions, in transgenic control roots. C, In contrast, in TOR-RNAi roots, the IT was broader and was
arrested within the root hair cell. D, Average nodule numbers in transgenic control and TOR-RNAi roots at 21 dpi. E and F,
Representative young nodule images showing fully colonized central tissue in transgenic control roots (E) and uninfected central
tissue in TOR-RNAi roots (F). ccd, Cortical cell division; rh, root hair. G and H, RT-qPCR analysis showing decreases in the
transcript abundance of early nodulins (ERN1, ENOD40, and NIN; G) and CSP genes (SymRK, CCamK, and VPY; H) in TOR-RNAi
transgenic roots relative to transgenic control roots. The data are presented as averages of three biological replicates (n . 30 for A
and D and n . 9 for G and H). The statistical significance of differences between transgenic control and TOR-RNAi roots was
determined using an unpaired two-tailed Student’s t test (*, P , 0.05; **, P , 0.01; and ***, P , 0.001). The data represent
means 6 SE.
of nodule organogenesis in 35S-TOR-RNAi roots using and after the initial infection process. This complex in-
RT-qPCR to analyze the transcripts of CyclinD1, CyclinD3 teraction involves cellular and molecular responses in
(G1/S-phase progression) and CyclinB1-1 (G1/M-phase different cell layers of the host root, which ultimately
progression). The relative expression levels of all of these give rise to the nitrogen-fixing organ, the nodule. In
cyclin genes were reduced significantly in 35S-TOR-RNAi plants, TOR kinase activity is associated with growth
roots compared with controls (Fig. 14), indicating that TOR and development from embryogenesis to senescence
decreases the transcripts of cyclins that might affect cortical (for review, see Xiong and Sheen, 2014). However, be-
cell divisions during nodule organogenesis. yond postembryonic growth regulation, the role of
TOR during root nodule symbiosis has remained un-
explored. This study demonstrates that TOR is required
DISCUSSION
for both early and late symbiotic processes in bean.
The symbiotic association between legume roots and In TOR-silenced roots, the frequent failure of IT pro-
Rhizobium involves regulatory cross talk before, during, gression beyond the epidermis and the lack of nodule
Plant Physiol. Vol. 172, 2016 2013
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Nanjareddy et al.
Figure 12. Quantitative analysis of nitrogen fixation and light micrographs showing the structural characteristics of transgenic
control and TOR-RNAi nodules. A, Nitrogenase activity, as determined using an acetylene reduction assay, in 21-dpi transgenic
control and TOR-RNAi nodules. The data are presented as averages of three biological replicates (n . 33). The statistical sig-
nificance of differences between data from transgenic control and TOR-RNAi nodules was determined using an unpaired two-
tailed Student’s t test (***, P , 0.001). NDW, Nodule dry weight. B, Quantification of the infection cell size of nodules from
transgenic control and TOR-RNAi roots. The data are presented as averages of two biological replicates (n . 48). The statistical
significance of differences between control and TOR-RNAi-infected cells was determined using an unpaired two-tailed Student’s
t test (**, P , 0.01). For A and B, the data represent means 6 SE. C and D, Toluidine Blue-stained transverse sections of an
R. tropici-inoculated nodule at 21 dpi showing the morphology and organization of representative samples collected from
transgenic control (C) and TOR-RNAi (D) roots. E and G, Higher magnification images showing a dense cytoplasm without
vacuoles in infected cells of transgenic control nodules. F and H, In contrast, the infected cells of TOR-RNAi nodules show a
multivacuolated cytoplasm. c, Cortex; ic, infected cell; ui, uninfected cell; vb, vascular bundle.
organogenesis show that cellular responses are regu- gene, unlike in yeast, where endoreduplication led to
lated by TOR. Furthermore, the reduction of early the presence of two functionally similar TOR genes
nodulin transcripts upon rhizobial infection demon- (Helliwell et al., 1994). In yeast and mammals, TOR
strates the lack of molecular responses in different cell forms two multiprotein complexes: the rapamycin-
layers, resulting in the reduction of the total nodule sensitive TORC1 (Martin and Hall, 2005) and the
number by 80%. Thus, we propose that TOR is required rapamycin-insensitive TORC2 (Wullschleger et al., 2006)
for IT progression and nodule organogenesis in bean. complexes. Although the precise compositions of the
TOR kinase complexes in plants have not been system-
Bean TOR Has High Similarity to TOR Genes from atically characterized, some of the components and
Other Species downstream effectors of mTORC1 have been identified
through sequence similarity searches in organisms
Bean TOR shares a high sequence similarity with other ranging from the unicellular green alga Chlamydomonas
eukaryotic TOR genes. Here, based on bioinformatics reinhardtii to the eudicot Arabidopsis (Ahn et al., 2011;
analysis, the functional component of the TOR path- Moreau et al., 2012; Ren et al., 2012; Xiong and Sheen,
way (the TOR protein) in bean was found to be a single 2012). TORC1 translationally regulates cell proliferation
2014 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Role of TOR in the Legume-Rhizobium Symbiosis
Figure 13. Ultrastructural characteristics of TOR-RNAi nodules. A and B, Transmission electron microscopy images of R. tropici-
inoculated 21-d-old nodules showing the architecture of infected cells of transgenic control (A) and TOR-RNAi (B) roots. C,
Symbiosomes and bacteroids of the infected cells appear similar to wild-type bean nodules (Cermola et al., 2000). D, Infected
cells of TOR-RNAi roots show an abundant, clear, multivacuolated cytoplasm. E and F, Higher magnification images showing the
size of symbiosomes in transgenic control (E) and TOR-RNAi (F) nodules. G and H, Nodules showing different cell organelles in
the cytoplasm of infected cells from transgenic control roots (G) and several autophagosomes in the cytoplasm of infected cells
from TOR-RNAi samples (H). I, Intercellular IT harboring densely packed bacteria. J, In contrast, TOR-RNAi had sparsely dis-
tributed bacteria. as, Autophagosomes; b, bacteroid; cw, cell wall; db, degrading (ghost) bacteria; er, endoplasmic reticulum;
g, Golgi apparatus; ic, infected cell; mt, mitochondria; n, nucleus; s, symbiosome; ui, uninfected cell; v, vacuole.
and temporal cell growth by promoting anabolic pro- Arabidopsis have shown that TOR is expressed in em-
cesses, and TORC2 regulates spatial cell growth by reg- bryos, the endosperm, and the primary meristems. Here,
ulating cytoskeletal structure and polarity (Wullschleger we demonstrated the expression of the PvTOR promoter
et al., 2006). The bean TOR protein showed a domain in root nodules at different developmental stages.
structure similar to that observed in other eukaryotes
and Arabidopsis, including FAT, FRB, kinase, and FATC
domains and a short C-terminal sequence. Among these TOR Modulates Growth and Development in Bean Roots
domains, the kinase domain showed maximum conser- In this study, a marked reduction of TOR mRNA
vation across species; this domain is known to regulate levels in bean produced a noticeable root phenotype,
development and rRNA expression in Arabidopsis (Ren including reductions in root length and lateral root
et al., 2011). The phylogenetic analysis of bean TOR density, which is consistent with previous observations
showed that this gene belongs to the legume group and in Arabidopsis (Deprost et al., 2007; Ren et al., 2012;
is closely related to the G. max and M. truncatula TOR Xiong and Sheen, 2012, 2015; Rexin et al., 2015). TOR-
genes. Previously, promoter and expression studies in RNAi roots had short RAMs and reduced epidermal
Plant Physiol. Vol. 172, 2016 2015
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Nanjareddy et al.
Figure 14. RT-qPCR analysis of cyclin genes in TOR-RNAi roots after inoculation with R. tropici. Transcript accumulation of G1/
S-phase progression genes, such as CyclinD1 (CycD1) and CycD3 (A), and CycB1-1, a G1/M-phase progression gene (B), was
analyzed in 7-dpi transgenic roots of control and TOR-RNAi plants. Transcript accumulation was normalized based on the ex-
pression of Ef1a and IDE, which were used as reference genes. The data are presented as averages of three biological replicates
(n . 9). The statistical significance of differences between transgenic control and TOR-RNAi roots was determined using an
unpaired two-tailed Student’s t test (***, P , 0.001). The data represent means 6 SE.
cell lengths in the root elongation zone. Because TOR progression. IT abortions at the base of the root hair
has been reported to sense and transduce shoot-derived cells have been documented in several gene-silenced
Glc signals to activate the root meristem (Xiong et al., and mutant legume lines (Popp and Ott, 2011). These
2013; Xiong and Sheen, 2014), we presume that a similar phenotypes are most often associated with impaired
explanation exists for the root phenotype. The tran- nodulin induction. Similarly, TOR-silenced roots in this
scriptional reduction of RbohB, SOD, and CAT in TOR- study also showed reduced transcript levels of nodulins
RNAi plants also explains the root length reduction, as and common symbiotic genes.
recent evidence from bean has shown that RbohB- Autophagy is a survival mechanism in cellular ho-
mediated ROS induction is critical for primary root meostasis and a variety of stress responses (Liu and
growth (Montiel et al., 2012) and lateral root devel- Bassham, 2012; Boya et al., 2013), and its protective
opment (Montiel et al., 2013). Furthermore, the down- function has been implicated in innate immune re-
regulation of TOR affected the expression of ROL5 and sponses to pathogen infection (Deretic, 2012). Here, the
LRX1, the genes responsible for the cell wall modula- up-regulation of autophagic processes could indicate
tion required for root hair cell expansion (Leiber et al., the possibility that a symbiont is recognized as a
2010). Thus, we propose that TOR regulates root length pathogen under TOR-suppressed conditions, leading to
and development by coordinating root meristematic IT abortions within the root hair cell.
activity and cellular enzymatic and nonenzymatic ROS Furthermore, ROS accumulation in response to rhi-
production. zobial infection is one of the primary requirements
for IT progression in bean (Montiel et al., 2012), and
the IT phenotype in this study could be attributed to the
TOR Down-Regulation Impairs the Infection Process impaired ROS accumulation in TOR-silenced plants.
Interestingly, in S. cerevisiae, TOR complex 2-YPK1
Rhizobium nod factor signals initiate a cascade of signaling regulates actin polarization by controlling
events in root hair cells, chiefly calcium spikes in and ROS accumulation (Niles and Powers, 2014). In plants,
around the nucleus and the reorganization of the actin although the TORC2 homolog has not yet been iden-
cytoskeleton. This process also triggers two signal tified, the change in ROS accumulation that occurs upon
transduction pathways: one pathway leads to the ex- the suppression of the TOR transcript suggests the pos-
pression of nodulins, resulting in the formation of sibility that the unique TOR complex described in plants
nodules via the differentiation of cortical cells, whereas could participate in both cell growth and cytoskeletal
the other pathway leads to the deformation of root hairs organization. Hence, we hypothesize that TOR silencing
and the subsequent entrapment of the rhizobia. TOR in bean could lead to an impaired cytoskeleton and
gene silencing driven either by a constitutive promoter consequently affect the IT progression.
(35S) or a symbiosis-specific NIN promoter in bean
leads to normal root hair curling; however, ITs were
aborted within the root hair cells. These results dem- TOR Down-Regulation Alters Nodule Morphology
onstrated that, although TOR is a ubiquitous regulator
of plant development (Deprost et al., 2007; Ren et al., During nodule organogenesis, while the ITs pro-
2012; Xiong and Sheen, 2012), given the rhizobial gress, the cortical cells dedifferentiate and reenter the
symbiotic conditions, it has a direct role to play in IT cell cycle to initiate nodule primordia. An intriguing
2016 Plant Physiol. Vol. 172, 2016
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.Role of TOR in the Legume-Rhizobium Symbiosis
feature of TOR silencing in Rhizobium-inoculated bean sequences identified in this study were aligned with highly similar sequences
from GenBank using ClustalW. Phylogenetic analysis was inferred using the
roots was the absence of infection event-associated neighbor-joining method (Saitou and Nei, 1987) and was calculated using the
cortical cell divisions under both 35S and NIN promoter- Kimura (Kimura, 1980) and parsimony methods. A bootstrap confidence
driven silencing conditions. The roles of several core analysis was performed in MEGA 5.0 (Tamura et al., 2011) with 1,000 replicates
cell cycle regulatory genes that function in nodule for- (Felsenstein, 1985). The TOR sequences were aligned and analyzed using
the PRALINE program (http://www.ibi.vu.nl/programs/pralinewww/;
mation in M. truncatula have been revealed (Foucher
Simossis et al., 2005).
and Kondorosi, 2000). For instance, CyclinD3s have
been implicated as the set of cyclins responsible for the
mitotic activation of cortical cells within the first 24 h of Plant Growth and Inoculation
rhizobial inoculation during nodule primordia orga- Common bean (‘Negro Jamapa’) seeds were surface sterilized and germi-
nogenesis. Incidentally, TOR’s role as a cell cycle reg- nated in the dark for 2 d. The 2-d-old seedlings were transferred to pots with
ulator through the translational activation of the sterile vermiculite, inoculated with Rhizobium tropici (strain CIAT899) at OD600,
and irrigated with B&D medium (Broughton and Dilworth, 1971) without
downstream targets 4E-BP and S6K1 is widely under- nitrate.
stood in mammals (Dowling et al., 2010; Ben-Sahra
et al., 2013). In Arabidopsis, E2Fa phosphorylation by
TOR was reported recently (Xiong et al., 2013). In this Plasmid Construction and Composite Plants
study, the TOR-silenced roots showed an inability to Upstream of the TOR translation start site, 1,000 bp of the promoter was
induce CyclinD1 and CyclinD3 expression in response amplified from bean genomic DNA using primer-specific oligonucleotides
to Rhizobium. Hence, we hypothesize that the absence of (Supplemental Table S1) and cloned into the pENTR/SD/D-TOPO vector
(Invitrogen). The Gateway LR reaction was performed between the entry vector
cortical cell divisions in response to infection could be pENTR/SD/D-TOPO-TOR and the destination vector pBGWSF7.0 (Karimi
caused by the lack of CyclinD1 and CyclinD3 during et al., 2002), according to the manufacturer’s instructions (Invitrogen).
nodule organogenesis. To develop the RNAi construct, we first developed pTdT-pNIN-RNAi
Nevertheless, the sparse nodules found on TOR-RNAi vector. This was derived from pTdT-DC-RNAi vector (here referred to as
pTdT-35S-RNAi; Valdés-López et al., 2008) by replacing the 35S promoter (cut
plants were smaller and had a cellular organization with XhoI and SacII) with the PvNIN promoter amplified from gDNA of bean
similar to those of the controls. Furthermore, the ultra- root tips (Supplemental Fig. S9, A and B.) A fragment corresponding to the
structure revealed the presence of a large number of nonconserved region of the C terminus and 39 UTR of TOR was amplified from
vacuoles and fewer symbiosomes, which each typically the cDNA isolated from the root tips of 2-d-old germinated bean using specific
oligonucleotides (Supplemental Table S1.) The PCR product was recombined
house only one bacteroid. Considering the fact that TOR
with the pTdT-35S-RNAi vector and pTdT-pNIN-RNAi vector using the
silencing reduced root and root hair growth, we specu- Gateway system (Invitrogen). The correct orientation of the clone was con-
late that the plant might sense a nutritional deficiency. firmed by sequencing the plasmid insert. The empty pTdT-RNAi vector and
Because root nodule symbiosis is a high-cost biological pTdT-pNIN-RNAi vectors were used as controls.
process for both partners (Libault, 2014), under TOR- The Agrobacterium rhizogenes K599 strain carrying the corresponding con-
structs was used to generate hairy root formation on bean tissues and to form
down-regulated conditions, the host plant’s photosyn- composite plants after transformation (Estrada-Navarrete et al., 2007). Transgenic
thetic carbon output is probably not sufficient to support hairy roots expressing the TOR-RNAi vectors and the TOR::GFP-GUS promoter
the energy requirements of the symbiont. This argument were selected using an epifluorescence stereomicroscope based on the fluores-
is supported by the presence of very few amyloplasts cence of red fluorescent protein (RFP) and GFP, respectively. RFP fluorescence
was excited at 561 nm using a solid-state laser, and the emission was filtered using
in uninfected nodule cells and fewer poly-b-hydroxy- a 640/50-nm band-pass filter. GFP fluorescence was excited with a blue argon ion
butyrates in bacteroids. Furthermore, the nodules de- laser (488 nm), and the emitted fluorescence was collected from 510 to 540 nm.
rived from TOR-silenced roots fix negligible amounts of
nitrogen, indicating that the nodules are nonfunctional. Physiological Analysis
Taken together, these observations show that TOR reg-
ulates nodule organogenesis and functionality in bean. Composite plants grown in pots containing vermiculite or in glass tubes
Based on the symbiotic phenotype observed in TOR- (15 cm) containing B&D medium were used to determine growth parameters,
such as root hair, root growth, and nodulation. Root hair parameters were
RNAi plants, especially when the silencing was driven recorded from uninoculated transgenic roots grown in glass tubes at 10 dpe. The
by the NIN promoter, where the effect of TOR silencing root growth parameters, such as the root-shoot ratio, root length, and lateral
was specific toward symbiotic processes, we propose root density, were obtained from individual uninoculated TOR-RNAi and
the possibility that TOR functions through multiple control hairy roots. Lateral root density was calculated using the formula D =
LR/L9, where D = density, LR = the number of lateral roots, and L9 = the length
pathways as a facilitator of rhizobial symbiotic pro- of the primary root between the first and last lateral roots (Dubrovsky et al.,
cesses ranging from IT formation to the differentiation 2006). To determine the effect(s) of TOR-RNAi on root symbiosis, transgenic
of mature nodules. Hence, TOR is essential for root roots were inoculated with R. tropici harboring a GUS (Vinuesa et al., 2003) or a
nodule symbiosis in bean. GFP reporter gene. The number of infection events was determined at 7 dpi, and
nodule quantification analyses were performed periodically up to 21 dpi in the
roots of vermiculite-grown transgenic plants. GUS activity was analyzed
according to the protocol described by Jefferson (1987).
MATERIALS AND METHODS
Phylogenetic Analysis and Protein Sequence Alignment RNA Isolation and Expression Analysis Using RT-qPCR
Based on the Arabidopsis (Arabidopsis thaliana) TOR sequence (Deprost et al., Total RNA was extracted from tissues ground in liquid nitrogen using the
2007), a homolog was identified in the genome of common bean (Phaseolus TriPure Isolation Reagent (Roche) according to the manufacturer’s instructions.
vulgaris) via TBLASTX analysis against the bean genomic data (Phytozome Genomic DNA contamination was eliminated from the RNA samples by in-
version 10.2; http://phytozome.jgi.doe.gov/pz/portal.html). The gene cubating the samples with RNase-free DNase (1 unit mL21). RNA integrity and
Plant Physiol. Vol. 172, 2016 2017
Downloaded on March 21, 2021. - Published by https://plantphysiol.org
Copyright (c) 2020 American Society of Plant Biologists. All rights reserved.You can also read

















































