The Solo Play of TERT Promoter Mutations - MDPI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
cells
Review
The Solo Play of TERT Promoter Mutations
François Hafezi and Danielle Perez Bercoff *
Department of Infection and Immunity, Luxembourg Institute of Health, 29, rue Henri Koch, L-4354
Esch-sur-Alzette, Luxembourg; francois.hafezi@lih.lu
* Correspondence: danielle.perezbercoff@lih.lu; Tel.: +352-6970-318
Received: 11 February 2020; Accepted: 16 March 2020; Published: 19 March 2020
Abstract: The reactivation of telomerase reverse transcriptase (TERT) protein is the principal
mechanism of telomere maintenance in cancer cells. Mutations in the TERT promoter (TERTp) are a
common mechanism of TERT reactivation in many solid cancers, particularly those originating from
slow-replicating tissues. They are associated with increased TERT levels, telomere stabilization, and
cell immortalization and proliferation. Much effort has been invested in recent years in characterizing
their prevalence in different cancers and their potential as biomarkers for tumor stratification, as well
as assessing their molecular mechanism of action, but much remains to be understood. Notably, they
appear late in cell transformation and are mutually exclusive with each other as well as with other
telomere maintenance mechanisms, indicative of overlapping selective advantages and of a strict
regulation of TERT expression levels. In this review, we summarized the latest literature on the role
and prevalence of TERTp mutations across different cancer types, highlighting their biased distribution.
We then discussed the need to maintain TERT levels at sufficient levels to immortalize cells and
promote proliferation while remaining within cell sustainability levels. A better understanding of
TERT regulation is crucial when considering its use as a possible target in antitumor strategies.
Keywords: TERT promoter mutations; telomerase; cell immortalization; GBM/glioma; melanoma;
thyroid cancer; APOBEC mutations; UV mutations
1. Introduction
Telomeres and their associated shelterin complex are located at chromosomal ends. Telomeres are
tandem repeats of TTAGGG up to 15 kb long in humans. Together, telomeres and the shelterin complex
protect chromosomal ends and preserve genomic DNA integrity [1–4]. Telomeres are shortened with
each cell division. When telomere length falls below a critical threshold, cells become replicatively
senescent and undergo apoptosis [5]. Cancer cells circumvent replicative telomere shortening by
stabilizing them [6] through one of two mechanisms: reactivation of telomerase, the enzyme that extends
telomeres (85–90% of cancers) [7–10], or homologous recombination between sister chromatids, a
phenomenon termed alternative lengthening of telomeres (ALT) (3–10% of cancers) [10–12]. Telomerase
is a ribonuclear holoenzyme composed of an RNA template (TERC) and a reverse transcriptase
catalytic subunit (TERT) [1–4,13]. TERT is silent in most somatic cells, and is reactivated in cancer cells,
endowing them with unrestricted proliferation capacity [6,14–16].
Although TERT activity is regulated principally at the transcriptional level (reviewed in
References [3,4,9,17–22]), it may also be regulated through splicing [23,24], post-translational
modifications, or intracellular trafficking [25–28]. The TERT promoter (TERTp) contains binding
sites for numerous transcriptional activators including Sp-1, c-Myc, Hypoxia Induced Factor (HIF),
AP-2, β-catenin, NF-κB, E-twenty-six (Ets)/ternary complex factors (TCF) family members, and
transcriptional repressors (Wilms’ tumor (WT1), TP53, Nuclear Transcription Factor, X-Box Binding
(NFX-1), Mad-1 and CCCTC binding factor (CTCF)) [3,4,9,17–21,29]. TERT expression can be reactivated
Cells 2020, 9, 749; doi:10.3390/cells9030749 www.mdpi.com/journal/cells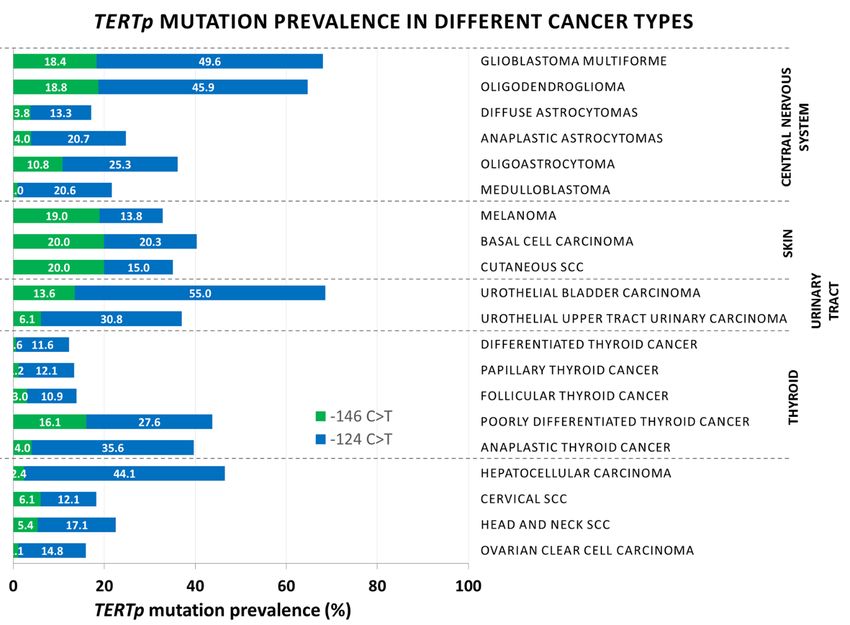
Cells 2020, 9, x FOR PEER REVIEW 2 of 25
Cells 2020, 9, 749 2 of 28
reactivated by copy number variants (CNV), TERT or TERTp structural variants, chromosomal
rearrangements
by copy numberjuxtaposing
variants (CNV), TERTpTERT to or
enhancer elements, variants,
TERTp structural cellular and viral oncogenes
chromosomal such as
rearrangements
Hepatitis
juxtaposing B virus
TERTp (HBV) X protein
to enhancer (HBx) cellular
elements, or high-risk Human
and viral papillomavirus
oncogenes (HPV)16Band
such as Hepatitis virusHPV18
(HBV)
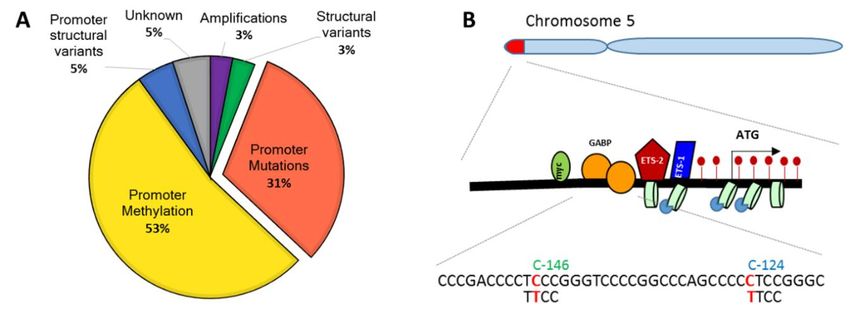
E6 oncoprotein, and, last but not least, mutations within TERTp (31% of TERT-expressing
X protein (HBx) or high-risk Human papillomavirus (HPV)16 and HPV18 E6 oncoprotein, and, last but cancers)
(Figure
not least,1A) [10,30–38]
mutations (reviewed
within in [3,4,9,18–20,39]).
TERTp (31% of TERT-expressing Increased
cancers)TERTp
(Figuremethylation
1A) [10,30–38]is (reviewed
typically
recorded in >50% ofIncreased
in [3,4,9,18–20,39]). TERT-expressing tumors and
TERTp methylation cell linesrecorded
is typically [10,40–47]. in >50%
Epigenetic regulation of
of TERT-expressing
TERTp
tumorsisand based on altered
cell lines methylation
[10,40–47]. Epigenetic patterns of specific
regulation of TERTp regions.
is based Hypomethylation of thepatterns
on altered methylation region
between
of specific−200 and −100
regions. from the Translational
Hypomethylation Startbetween
of the region site (TSS),
−200 encompassing
and −100 from thethe
core promoter,
Translational
enables binding of c-Myc and Sp-1, thus reactivating transcription. In contrast,
Start site (TSS), encompassing the core promoter, enables binding of c-Myc and Sp-1, thus reactivatingthe region spanning
transcription. In contrast, the region spanning exon 1 (positions +1 to ±100 from the TSS) contains
exon 1 (positions +1 to ±100 from the TSS) contains a binding site for the DNA insulator CTCF.a
Hypermethylation
binding site for the DNA of thisinsulator
region disrupts binding of CTCFof
CTCF. Hypermethylation and
thistherefore allows binding
region disrupts TERT transcription
of CTCF and
[41–44].
therefore allows TERT transcription [41–44]. Similarly, the region between −600 and −200 frombinding
Similarly, the region between −600 and −200 from the TSS contains a second CTCF the TSS
site and isa partially
contains second CTCF hypermethylated
binding site andin TERT-expressing cells [41–44].
is partially hypermethylated inThe transcriptional
TERT-expressing control
cells of
[41–44].
TERT has been comprehensively
The transcriptional control of TERT reviewed
has been recently [3,4,9,18–22,29,48]
comprehensively reviewed and, as such,
recently is beyond the
[3,4,9,18–22,29,48]
scope
and, asofsuch,
this review.
is beyond In the
thisscope
review, we review.
of this focused Inonthis
thereview,
distribution and exclusiveness
we focused of TERTp
on the distribution and
mutations.
exclusiveness of TERTp mutations.
Figure 1.
Figure Mechanismsofoftelomerase
1. Mechanisms telomerasereverse
reversetranscriptase
transcriptase(TERT)
(TERT)reactivation
reactivationin
incancer
cancerand TERT
andTERT
promoter(TERTp)
promoter (TERTp) mutations.
mutations.(A)
(A)Different
Differentmechanisms
mechanismsof TERTreactivation
ofTERT reactivationin
incancer
canceraccording
accordingto
to
Reference [10]. (B) Localization of TERTp mutations on Chromosome
Reference [10]. (B) Localization of TERTp mutations on Chromosome 5. 5.
2. Telomerase Reverse Transcriptase Promoter (TERTp) Mutations
2. Telomerase Reverse Transcriptase Promoter (TERTp) Mutations
TERTp mutations were first described in congenital and sporadic melanoma in 2013 [49,50].
Subsequent mutations
TERTp large-scale werecohortfirst described
studies togetherin with
congenital
seminaland sporadic studies
mechanistic melanoma bothinascertained
2013 [49,50]. the
Subsequent large-scale cohort studies together with seminal mechanistic studies
TERTp mutation prevalence in many other forms of cancer and characterized their mode of action. both ascertained the
TERTp Themutation
two main prevalence in many other
TERTp mutations formsatofpositions
are located cancer and characterized
1,295,228 their mode
and 1,295,250 of action.
on Chromosome
The two main TERTp mutations are located at positions 1,295,228 and 1,295,250
5, and generate C to T transitions. They are located 124 and 146 base pairs upstream from the TERTp on Chromosome
5, and(Figure
TSS generate 1B). C Less
to T transitions. They are
frequent tandem located 124
mutations and 146 CC>TT
−125/−124 base pairsandupstream
−139/−138 from the TERTp
CC>TT have
TSS (Figure 1B). Less frequent tandem mutations −125/−124 CC>TT and −139/−138
been reported in cutaneous tumors (Table 1) [49,51]. While these are somatic mutations, a germline CC>TT have been
reported
mutationin at cutaneous
position −57A>Ctumorsfrom(Tablethe1)TSS[49,51]. While
has been these are
identified somaticmelanomas
in familial mutations, and a germline
showed
mutation at position −57A>C from the TSS has been identified in familial melanomas
similar effects [49]. All of these mutations have similar effects, increasing TERT expression ~2–6 and showed
fold as
similar effects [49]. All of these mutations have similar effects, increasing
measured through qRT-PCR, immunohistochemistry, TRAP, or reporter vectors in numerous cancer TERT expression ~2–6 fold
as measured through qRT-PCR, immunohistochemistry, TRAP, or reporter
types, as outlined in Table 1 [37,50,52–65]. This increased TERT expression maintains self-renewal vectors in numerous
cancer types,
potential and as outlinedininboth
telomeres Table
stem 1 [37,50,52–65]. This increased
cells and terminally TERTbladder
differentiated expression
cells,maintains
indicatingself- that
renewal potential and telomeres in both stem
these mutations are sufficient to immortalize cells [66,67]. cells and terminally differentiated bladder cells,
indicating
All ofthat
these these mutations
TERTp are (at
mutations sufficient
positions to −146,
immortalize cellsand
−124, −57, [66,67].
−139/−138) create novel Ets/TCF
All of these TERTp mutations (at positions −146, −124, −57,
transcription factor binding sites. The Ets/TCF transcription factors bind to GGAAand −139/−138) createmotifs
novel(or Ets/TCF
TTCC
transcription factor binding sites. The Ets/TCF transcription factors bind to
on the opposite strand). The 30 members of the Ets/TCF-family transcription factors are importantGGAA motifs (or TTCC
on the opposite
contributors strand). The and
to oncogenesis 30 members of the Ets-2,
include Ets-1, Ets/TCF-family transcription
and GA binding proteinfactors
(GABP) are[68].
important
So far,
contributors to oncogenesis and include Ets-1, Ets-2, and GA binding protein
GABP has been reported to selectively bind the −124 C>T and −146 C>T mutations in GBM, melanoma, (GABP) [68]. So far,
GABP has been reported to selectively bind the −124 C>T and −146 C>T mutations in GBM,Cells 2020, 9, 749 3 of 28
and urothelial bladder cancer cell lines [69–71]. Unlike the other Ets/TCF family transcription
factors, GABP is an obligate dimer of GABPA and GABPB dimers. It binds two nearby in-phase
GGAA sites [68,72–74] positioned 1, 2, or n helical turns away from each other [69], or brought
close together by DNA looping [70]. TERTp mutations are associated with epigenetically active
chromatin [54,69,75,76]. Intriguingly, whereas methylation of wild-type (wt) TERT promoter is
associated with TERT expression [10,43,44], TERTp mutations are associated with decreased TERTp
methylation [76]. The −146 C>T mutation was also shown to bind the non-canonical NF-κB-p52 and
Ets-1/2 [59].
TERTp mutations have been recorded in a wide range of solid cancers. They are
present in primary gliomas and glioblastoma multiforme (GBM), oligodendrogliomas and
astrocytomas [10,40,52–54,57,58,60,64,65,77–86], melanomas, cutaneous basal cell carcinoma (BCC)
and squamous cell carcinoma (SCC) [49–52,55,87–91], myxoid liposarcomas [77], urothelial
bladder carcinoma [50,57,78,92–94], hepatocellular carcinoma (HCC) [50,57,62,95–97], and thyroid
cancers [98–106], as well as oral and cervical SCC [36,37,57] (Table 1). Furthermore, they were
consistently found in tumor cell lines derived from these malignancies [37,50,52,54,58,62,97,100,107,108].
TERTp mutations often arise in tissues with low rates of self-renewal (brain, thyroid) [77], where they
provide an immediate competitive advantage to cells that acquire them. Conversely, they appear to
be infrequent (Cells 2020, 9, 749 4 of 28
3.2. Melanoma and Non-Melanoma Skin Carcinoma
In patients with primary melanoma, TERTp mutations have been reported in 39.2% (range
22–71%) of tumors. They arise progressively in sun-exposed sites and have been attributed to
UV radiation. They are associated with increased patient age, distal metastases, poor outcome,
and compromised OS and disease-free survival (DFS) [49–52,88,89,115]. In ~50% of cases, they are
associated with mutations in BRAF/NRAS [49,52,88,89,91,116], influencing OS in the following order:
TERTpmut +BRAF/NRASmut T occur with similar frequencies
in contrast to all other cancers, where −124 C>T is by far the most prevalent mutation (Table 1).
Second, −139/−138 CC>TT and −125/−124 CC>TT tandem mutations are often reported. Third, TERTp
mutations were detected in 9/10 melanomas with ALT in one study [117] and together (−124 C>T +
−146 C>T) in two patients with BCC in another study [89], indicating that more than one telomere
maintenance mechanism can, unusually, coexist in skin cancers.
3.3. Urothelial Bladder Cancer
TERTp mutations have been detected in 64.6% (range 29.5–100%) of urothelial bladder
and upper urinary tract cancers. They are the most common somatic lesions in this cancer
type [52,57,61,77,92,94,118,119]. They have been associated with reduced survival, disease recurrence,
and distal metastases [61,118,119], although there appears to be no difference between early- and
late-stage patients [52,94].
3.4. Thyroid
Among thyroid cancers, TERTp mutations have been reported mainly in follicular-cell-derived
thyroid malignancies (papillary thyroid carcinoma (PTC): 13.4%, range 4.1–37.7%; follicular thyroid
carcinoma (FTC): 13.9%, range 5.9–66.7%; poorly differentiated thyroid carcinoma (PDTC): 43.7%, range
21–51.7%; and anaplastic thyroid carcinomas (ATC): 39.7%, range 13–50%). The presence of TERTp
mutations is significantly associated with increased age, tumor size and stage, distal metastases, tumor
recurrence, and shorter OS and DFS in PTC and FTC. Their prevalence increases from differentiated
PTC and FTC to the more aggressive poorly differentiated ATC (Table 1) [98–106]. The association of
TERTp mutations with the common BRAF-V600E mutation is a powerful predictor of poor OS and
DFS [52,98,99,104–106,108]. As in glioma, TERTp mutations compromise the outcome of radioiodine
therapy [101,105].
3.5. Hepatocellular Carcinoma (HCC)
TERTp mutations are an early event in hepatocellular tumorigenesis [57,62,77,95]. They are not
only seen in established HCC (47.1%, range 29.3–65.4%). As hepatocellular adenomas transform into
HCC, TERTp mutations are the first gene recurrently mutated after β-catenin (CTNNB1) in preneoplastic
cirrhotic lesions [62,95]. Together with the CTNNB1 mutation, TERTp mutations are considered critical
effectors of malignant transformation. As such, they have been proposed as early biomarkers for
hepatocellular transformation [62,77,95,96,120,121].
TERTp mutations appear to be more frequent in HCV-associated HCC [62,77,95,96,122]
and less frequent or excluded from HBV-associated HCC [62,96,121,122], although this remains
controversial [63,77,95]. HBV DNA insertion in the TERTp is a recurrent mechanism of TERT
transcriptional reactivation in HBV-associated HCC [34,123,124], and a genetic screen of TERT in HCC
found TERTp mutations to be mutually exclusive with HBV integration, TERT CNVs, and ATRX
mutations [121].Cells 2020, 9, 749 5 of 28
3.6. Cervical and Oral Head and Neck Squamous Cell Carcinoma (HNSCC)
TERTp mutations were detected in cervical SCC (20.1%, range 4.5–21.8%) and HNSCC (22.5%,
range 17–31.7%) [36,37]. These malignancies are often associated with high-risk-HPV16/18 E6 and
E7 viral oncoproteins and with APOBEC mutations [125–127]. High-risk HPV–E6 transactivates
TERT [30,32,33,128,129]. TERTp mutations have a notably higher prevalence in HPV-negative
cervical and oral SCC. This gives distinct patterns of TERT reactivation through mutually
exclusive mechanisms [36,37]. In cervical SCC, they are associated with higher TERT levels than
HPV16/18-E6-positive tumors and with advanced cervical cancer [36,37]. Broader studies are needed
to evaluate the added value of screening for the molecular mechanism underlying TERT reactivation
in cervical and oral SCC.
3.7. The rs2853669 Polymorphism
Among TERT polymorphisms, a common polymorphism (rs2853669 A>G) which disrupts a
pre-existing Ets/TCF binding site located 245 bp upstream of the TERT TSS has been reported to modify
the effect of TERTp mutations. It decreases TERT transcription in vitro and reverses TERT upregulation
by TERTp mutations [56,61,81,85,130]. Controversial clinical impacts have been reported, from a
beneficial effect on OS and limited tumor recurrence in TERTp-mutated urothelial bladder cancer, renal
clear cell carcinoma, melanoma, and GBM [56,61,81,85,116,131], to unchanged or worsened clinical
outcome in GBM, melanoma, or differentiated thyroid carcinomas [64,65,84,91,102,103]. In HCC, the
rs2853669 polymorphism in combination with TERTp mutations has been associated with decreased OS
and DSF, and increased TERTp methylation and expression [47]. Possible reasons for these conflicting
reports could be homozygosity versus heterozygosity of the variant, or its occurrence on the same
allele as TERTp mutations. Further studies are needed to assess the relevance of screening for this
polymorphism for prognostic and treatment purposes.Cells 2020, 9, 749 6 of 28
Table 1. Prevalence and distribution of telomerase reverse transcriptase promoter (TERTp) mutations in cancer genomes. The prevalence of TERTp mutations is given
as percentage and as total number of cases.
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Central nervous system (CNS)
DNA
62% 25% 75% Patients
GBM Yes sequencing, Associated with older age. [52]
24/39 6/24 18/24 (Portugal)
qRT-PCR, IHC,
DNA
83.9% 34% 65.9% sequencing, Patients
GBM IV Yes Associated with older age. [57]
47/55 16/47 31/47 qRT-PCR, TRAP, (China)
reporter assays
Patients
GBM 83% 24.6% 75.4% DNA Associated with shorter OS,
IV N/A (US [77]
(Primary) 65/78 16/65 49/65 sequencing IDH-wt, ATRX-wt, exclusively in EGFRmut samples.
American)
DNA
Associated with late-stage disease and patient age.
44.6% 26.7% 73.3% sequencing, Patients
GBM I–IV Yes Only in gliomas, not in pituitary adenocarcinomas, [60]
45/101 12/45 33/45 qRT-PCR, (China)
meningiomas or secondary metastases.
reporter assays
Associated with shorter OS and with EGFRmut .
55% 27% 73% DNA Patients Negatively associated with mutant IDH and TP53.
GBM N/A [111]
197/358 54/197 144/197 sequencing (Switzerland) More frequent in primary (58%) than in secondary GBM (28%).
One patient with both −146 C>T + −124 C>T mutation.
GBM Associated with shorter OS in patients without rs2853669 TERT
80.3% DNA
(primary & IV * * N/A Patients -245 A>G polymorphism. [81]
143/178 sequencing
secondary) Detected in 4/14 (28%) secondary GBM.
Patients Associated with older age, poor prognosis, and shorter
66.9% 25.5% 74.5% DNA
GBM IV N/A (Portugal & survival. [85]
141/211 36/141 105/141 sequencing
Brazil) Reversed by rs2853669 TERT −245 A>G polymorphism.
Associated with older age.
Not associated with OS or DFS.
DNA Associated with MGMT methylation and EGFR amplification.
60.4% 24.1% 75.8% Patients
GBM Yes sequencing, Associated with rs2853669 TERT −245 A>G polymorphism [64]
29/48 7/29 22/29 (Korea)
qRT-PCR (21/29 patients).
rs2853669 TERT −245 A>G polymorphism reversed TERT
upregulation by TERTp mutations.
Mutually exclusive with IDH-1 mutations.
DNA Associated with shorter telomeres.
73% 28% 82% sequencing, Associated with lower OS in IDH-1wt patients.
GBM Yes [65]
92/126 26/92 66/92 qRT-PCR, TRAP, rs2853669 TERT -245 A>G polymorphism associated with
qPCR improved OS in patients without TERTp mutations, and with
worse OS in patients with TERTp mutations.Cells 2020, 9, 749 7 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Associated with older age and shorter OS.
GBM 86% 25% 75% DNA Homozygous rs2853669 TERT −245 A>G polymorphism
[84]
(primary) 79/92 20/79 69/79 sequencing associated with worse OS in patients without and with TERTp
mutations.
GBM and
100% 10% 90% DNA In primary GBM, characterized by 10q deletion EFGR
gliomas N/A Patients [58]
10/10 1/10 9/10 sequencing amplification.
(primary)
2.2–286-fold
DNA
94% 36% 64% compared to
GBM sequencing, Cell lines [58]
33/35 12/33 21/33 normal
qRT-PCR
astrocytes
905/1331 206/762 567/762
Total GBM
(68%) (27%) (73%)
DNA
45% 20% 80% Patients
OligodendrogliomaII Yes sequencing, [52]
10/22 2/10 8/10 (Portugal)
qRT-PCR, IHC
DNA
70% 14.3% 85.7% sequencing, Patients
Oligodendroglioma
II–III Yes Associated with older age. [57]
7/10 1/7 6/7 qRT-PCR, TRAP, (China)
Reporter Assays
Patients
46.3% 24% 76% DNA Associated with older age at diagnosis.
Oligodendroglioma
II–III N/A (Portugal & [85]
25/54 6/25 19/25 sequencing Not associated with lower survival.
Brazil)
DNA Associated with total 1p19q loss and IDH-1/2 mutations (98%)
73.5% 20% 80% Patients
Oligodendroglioma Yes sequencing, but [53]
25/34 5/25 20/25 (Japan)
qRT-PCR exclusive with IDH-1mut if not total loss of 1p19q.
Patients
66.81% DNA Associated with shorter OS.
Oligodendroglioma
II–IV * * N/A (US [80]
151/226 sequencing Can be associated with ATRX mutations or IDHmut /1p19q loss.
American)
IDH-wt only.
Patients
63.2% 41.7% 58.3% DNA Associated with worse prognosis in IDH-wt.
Oligodendroglioma
II–III N/A (US [77]
12/19 5/12 7/12 sequencing Associated with older age.
American)
Mutually exclusive with ATRX mutations.
DNA
Anaplastic 54% 30.8% 69.2% Patients
III Yes sequencing, Associated with older age. [52]
oligodendroglioma 13/24 4/13 9/13 (Portugal)
qRT-PCR, IHC,
DNA Associated with total 1p19q loss and IDH-1/2 mutations (98%)
Anaplastic 74.2% 30.4% 69.6% Patients
Yes sequencing, but [53]
oligodendroglioma 23/31 7/23 16/23 (Japan)
qRT-PCR exclusive with IDH-1 if not total loss of 1p19q.Cells 2020, 9, 749 8 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Associated with older age.
Patients
Anaplastic 88.5% 43.5% 56.5% DNA IDH-wt only.
III N/A (US [77]
oligodendroglioma 23/26 10/23 13/23 sequencing Associated with worse prognosis in IDH-wt.
American)
Mutually exclusive with ATRX mutations.
Total 289/446 40/138 98/138
Oligodendroglioma (64.7%) (29%) (71%)
DNA Associated with total 1p19q loss and IDH-1/2 mutations (98%)
Diffuse 19.2% 20% 80% Patients
Yes sequencing, but [53]
astrocytomas 10/52 2/10 8/10 (Japan)
qRT-PCR exclusive with IDH-1 if not total loss of 1p19q.
DNA
Diffuse 15% 33,3% 66,6% Patients
II Yes sequencing, Associated with older age. [52]
astrocytoma 3/20 1/3 2/3 (Portugal)
qRT-PCR, IHC
DNA
Diffuse 20% 25% 62.5% sequencing, Patients
II Yes Associated with age. [57]
astrocytoma 8/40 2/8 5/8 qRT-PCR, TRAP, (China)
reporter assays
Patients
Diffuse 15.2% 16.7% 83.3% DNA
II N/A (Portugal & Frequency increased with grade. [85]
astrocytoma 7/46 1/7 6/7 sequencing
Brazil)
Total Diffuse 28/158 6/28 21/28
Astrocytoma (17.7%) (21.4%) (75%)
Patients
62.5% DNA Associated with shorter OS.
Astocytoma II–IV N/A N/A N/A (US [80]
416/665 sequencing Can be associated with ATRX mutations or IDHmut/1p1q loss.
American)
Patients
Anaplastic 10% 0% 100% DNA
III N/A (Portugal & Frequency increased with grade. [85]
Astrocytomas 1/10 0/1 1/1 sequencing
Brazil)
DNA
Anaplastic 33.3% 0% 100% sequencing, Patients
III Yes Correlation with age. [57]
Astrocytoma 4/12 0/4 4/4 qRT-PCR, TRAP, (China)
reporter assays
DNA
Anaplastic 25.3% 20% 80% Patients Associated with total 1p19q loss and IDH-1/2 mutations (98%)
III Yes sequencing, [53]
Astrocytomas 20/79 4/20 16/20 (Japan) but exclusive with IDH-1 if not total loss of 1p19q.
qRT-PCR
Total
25/101 4/25 21/25
Anaplastic
(24.7%) (16%) (84%)
AstrocytomasCells 2020, 9, 749 9 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Patients
Mixed 32.3% DNA Associated with shorter OS.
II–IV * * N/A (US [80]
Oligoastocytoma 63/195 sequencing Can be associated with ATRX mutations or IDHmut /1p1q loss.
American)
DNA
40% 28.6% 71.4% Patients Associated with total 1p19q loss and IDH-1/2 mutations (98%)
Oligoastrocytoma Yes sequencing, [53]
14/35 4/14 10/14 (Japan) but exclusive with IDH-1 if not total loss of 1p19q.
qRT-PCR
Patients
40.0% 50% 50% DNA
OligoastrocytomaII–III N/A (Portugal & Not associated with lower survival. [85]
4/10 2/4 2/4 sequencing
Brazil)
DNA
Anaplastic 48.9% 27.3% 72.7% Patients Associated with total 1p19q loss and IDH-1/2 mutations (98%)
Yes sequencing, [53]
Oligoastrocytoma 22/45 6/22 16/22 (Japan) but exclusive with IDH-1 if not total loss of 1p19q.
qRT-PCR
Total 103/285 12/40 28/40
Oligoastrocytoma (36.1%) (30%) (70%)
33.3% 50% 50% DNA Patients
Medulloblastoma N/A Associated with age. [57]
2/6 1/2 1/2 sequencing (China)
IDH-wt and ATRX-wt only.
Patients
20.9% 100% DNA Associated with worse prognosis in IDH-1-wt.
Medulloblastoma 0%0/19 N/A (US [77]
19/91 19/19 sequencing Associated with older age.
American)
Mutually exclusive with ALT.
Total 21/97 1/21 20/21
Medulloblastoma (21.6%) (4.7%) (95.3%)
Skin
DNA
71% 46% 54% Patients
Melanoma Yes sequencing, [50]
50/70 23/50 27/50 & cell lines
reporter vectors
-57 C>T germline mutation in family with history of
melanoma.
32.5% 20% 28% DNA High prevalence in metastatic cell lines (85%) compared to
Melanoma N/A Patients [49]
25/77 5/25 7/25 sequencing primary melanoma (32.5%).
CC>TT −139/−138 tandem mutation in 10.4% patients.
Concomitant with BRAF mutations in 47% of cases.
29% 50% 50% DNA Patients
Melanoma N/A Associated with BRAF mutations. [52]
16/56 8/16 8/16 sequencing (Portugal)
DNA
34% 52.5% 36% Patients CC>TT −139/−138 tandem mutations in 4/97 (4.1%) patients.
Melanoma Yes sequencing, [88]
97/287 51/97 35/97 (Spain) Associated with BRAF mutations in 50% cases.
qRT-PCRCells 2020, 9, 749 10 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Associated with shorter telomeres in tumor and with
accelerated telomere shortening rate.
41.6% DNA Patients
Melanoma * * N/A Associated with BRAF/NRAS mutation in 75/243 cases. [115]
121/291 sequencing (Spain)
Telomere shortening rate:
BRAF/NRASmut +TERTpmut >TERTpmut >BRAF/NRASmut
Associated with reduced OS & DFS.
More prevalent in sun-exposed regions.
DNA
22% 35% 46% Patients Associated with increased mitotic rates.
Melanoma Yes sequencing, [89]
26/116 9/26 12/26 (Portugal) −138/−138 CC>TT tandem mutation in 2/26 (7.7%) patients.
IHC
−125/−124 CC>TT tandem mutation in 3/26 (11.5%) patients.
Associated with BRAF-V600E mutation (58% of cases).
Associated with shorter OS and DFS.
−139/−138 CC>TT & −125/−124 CC>TT tandem mutations in
38.6% 50% 32.8% DNA Patients 16/116 cases (13.8%).
Melanoma N/A [116]
116/300 58/116 32/116 sequencing (Spain) Associated with BRAF/NRAS mutations in 126/283 (44.5%)
cases.
Reversed by rs2853669 TERT -245 A>G polymorphism.
−139/−138 CC>TT tandem mutations in 4/63 (6.3%) patients.
54.8% 61.9% 30.2% DNA Patients −125/−124 CC>TT tandem mutation in 1/63 (1.6%) patient.
Melanoma N/A [91]
63/115 39/63 19/63 sequencing (Austria) Associated with BRAF/NRAS mutation in 75/243 cases.
Associated with rs2853669 TERT -245 A>G polymorphism.
Total 514/1312 193/398 140/398
Melanoma (39.2%) (48.5%) (35.1%)
Basal cell 55.6% 55.6% 22.2% DNA Patients
N/A [55]
carcinoma 18/32 10/18 4/18 sequencing (Germany)
Mostly homozygous.
−139/−138 CC>TT tandem mutation in 7/31 (22.6%) patients.
Basal cell
−125/−124 CC–TT tandem mutation in 5/31 (16.1%) patients.
carcinoma 74% 35.5% 45.1% DNA
N/A Patients 1 patient with −139/−138 CC>TT + −125/−124 CC>TT tandem [90]
(sporadic & 31/42 11/31 14/31 sequencing
mutations.
nevoid)
Mutations more frequent in basal cell carcinoma than in
squamous cell carcinoma.
No correlation with clinical parameters.
Higher prevalence in patients not exposed to X-irradiation:
48/94 (51%) vs. 28/102 (27%) in X-irradiated patients.
DNA
Basal cell 38.7% 43% 49% Patients −124 C>T more frequent than −146 C>T in non-X-irradiated
no sequencing, [89]
carcinoma 76/196 33/76 37/76 (Portugal) patients;
IHC
−146 C>T more frequent in X-irradiated patients.
−139/138 CC>TT tandem mutation in 2/76 (2.6%) patients,
2 patients with −146 C>T + −124 C>T mutations.Cells 2020, 9, 749 11 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Total Basal 125/270 54/125 55/125
cell carcinoma (46.2%) (43.2%) (44%)
Cutaneous 50% 29.4% 29.4% DNA Patients
N/A [55]
SCC 17/34 5/17 5/17 sequencing (Germany)
Mostly homozygous.
Cutaneous 50% 54% 31% DNA −139/−138 CC>TT tandem mutation in 2/13 (15.4) patients.
N/A Patients [90]
SCC 13/26 7/13 4/13 sequencing Mutations more frequent in basal cell carcinoma than in
squamous cell carcinoma.
Total
30/60 12/30 9/30
Cutaneous
(50%) (40%) (30%)
SCC
Bladder/urinary tract cancers
Bladder 85% 4.5% 95.5% DNA Patients
N/A [78]
Cancer 44/52 2/44 42/44 sequencing (China)
Urothelial Patients
80% 17% 83% DNA
bladder III N/A (US [93]
12/15 2/12 10/12 sequencing
carcinoma American)
Urothelial Patients
66.7% 28.6% 71.4% DNA
bladder N/A (US [77]
14/21 4/14 10/14 sequencing
carcinoma American)
Urothelial
61.7% 25% 58.8% DNA Patients
bladder N/A Not associated with age. [57]
148/240 37/148 87/148 sequencing (China)
carcinoma
Urothelial DNA Not associated with age.
59% 37.5% 62.5% Patients
bladder N/A sequencing, Low-grade bladder cancer: 67%, [52]
48/82 18/48 30/48 (Portugal)
carcinoma qRT-PCR high-grade bladder cancer: 56%.
Associated with shorter telomeres and worse OS.
DNA
Urothelial Associated with FGFR3 mutation in 45% of tumors.
65.4% 17.8% 81.8% sequencing, Patients
bladder N/A FGFR3 mutations found in low-grade tumors, TERTp [61]
214/327 38/214 175/214 relative (Sweden)
carcinoma mutations in low-grade and high-grade tumors.
telomere length
Reversed by rs2853669 TERT −245 A>G polymorphism.
Urothelial DNA
77.1% 17% 83% Not associated with OS, DFS, or clinical outcome.
bladder Not increased sequencing, Patients [94]
361/468 62/361 299/361 Associated with FGFR3mut .
carcinoma qRT-PCR
Urothelial
100% 12% 85% DNA Pure micropapillary carcinoma and urothelial cancer with
bladder N/A Patients [92]
33/33 5/33 28/33 sequencing focal micropapillary features.
carcinoma
Urothelial
upper tract 76.9% 12.5% 72.5% DNA Patients
N/A Not associated with age. [57]
urinary 40/52 5/40 29/40 sequencing (China)
carcinomaCells 2020, 9, 749 12 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Urothelial
Patients
upper tract 47.4% 11.1% 88.9% DNA
N/A (US [77]
urinary 9/19 1/9 8/9 sequencing
American)
carcinoma
Urothelial DNA
upper tract 29.5% 18.5% 81.5% sequencing, Patients
N/A Associated with distant metastases. [118]
urinary 65/220 12/65 53/65 Detection in (China)
carcinoma urine
Total
Urothelial
bladder & 988/1529 186/988 771/988
upper tract (64.6%) (18.8%) (78%)
urinary
carcinoma
Thyroid
Differentiated 12.2% 4.9% 95.1% DNA
N/A Patients Only in malignant lesions. [108]
thyroid cancer 41/336 2/41 39/41 sequencing
DNA
Papillary 8% 7.7% 84.6% Patients
Yes sequencing, [52]
thyroid cancer 13/169 1/13 11/13 (Portugal)
qRT-PCR, IHC
Associated with older age, larger tumor size, extrathyroid
Papillary 11.3% 15.2% 85.8% DNA Patients
III/IV N/A invasion, advanced clinical stage. [99]
thyroid cancer 46/408 7/46 39/46 sequencing (China)
Associated with BRAF-V600E mutation.
Only in patients >45.
Papillary 27% 7.7% 92.3% DNA Patients
N/A Correlated with shorter telomeres and distal metastases. [98]
thyroid cancer 13/51 1/13 12/13 sequencing (Sweden)
PTC: 27% (25/332); FTC: 22% (12/70); ATC: 50% (12/36).
Associated with BRAF/RAS mutations.
Papillary 4.1% DNA Patients Associated with tumor size, stage III-IV, recurrence, decreased
III/IV * * N/A [106]
thyroid cancer 18/432 sequencing (Korea) OS and DFS with BRAF/RAS mutations: RAS/BRAF >TERTp >
RAS/BRAF+TERTp.
Papillary 11.7% 0% 100% DNA Only in malignant lesions.
N/A Patients [108]
thyroid cancer 30/257 0/30 30/30 sequencing −124 C>T associated with BRAF-V600E mutation.
Papillary 37.7% 10% 90% DNA Patients No TERTp mutation found in 192 well differentiated cancers
N/A [105]
thyroid cancer 10/27 1/10 9/10 sequencing (Korea) without distant metastasis.
Papillary 22% 44% 66% DNA Patients
N/A More frequent in BRAF-wt patients than in BRAFmut . [100]
thyroid cancer 18/80 8/18 10/18 sequencing (US & Japan)
Associated with older age (>45 years), larger tumor size, stage
III–IV, distant metastases, decreased OS and DFS.
Papillary 31.8% 0% 100% DNA Patients
N/A rs2853669 TERT −245 A>G polymorphism (46.7% (113/242)of [103]
thyroid cancer 77/242 0/77 77/77 sequencing (US)
patients) increases OS & DFS in patients without TERTp
mutations and with BRAF-V600E.Cells 2020, 9, 749 13 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Associated with older age and poor prognosis.
DNA
Papillary 12% 14.6% 86.4% Patients Increased cytoplasmic localization of TERT.
Yes sequencing, [102]
thyroid cancer 22/182 3/22 19/22 (Italy) No impact of rs2853669 TERT -245 A>G polymorphism on
WB, and IHC
outcome.
Total Papillary 247/1848 21/229 207/229
thyroid cancer (13.4%) (9.2%) (90.4%)
Follicular
13.9% 18.2% 81.8% DNA
Thyroid N/A Patients Only in malignant lesions. [108]
11/79 2/11 9/11 sequencing
Cancer
Follicular
66.7% 50% 50% DNA Patients No TERTp mutation found in 192 well-differentiated cancers
Thyroid N/A [105]
2/3 1/2 1/2 sequencing (Korea) without distanst metastasis.
Cancer
Follicular DNA
14% 22.2% 77.8% Patients
thyroid Yes sequencing, [52]
9/64 2/9 7/9 (Portugal)
Cancer qRT-PCR, IHC
Follicular 22% 12.5% 87.5% DNA Patients Increased prevalence in ATC: PTC: 27% (25/332); FTC: 22%
N/A [98]
thyroid cancer 8/36 1/8 7/8 sequencing (Sweden) (12/70); ATC: 50% (12/36).
Associated with older age, larger tumor size, extrathyroid
Follicular 36.4% 12.5% 87.5% DNA Patients
N/A invasion, advanced clinical stage. [99]
thyroid cancer 8/22 1/8 7/8 sequencing (China)
Associated with BRAF-V600E mutation.
Associated with BRAF/RAS mutations.
Follicular 5.9% DNA Patients Associated with tumor size, stage III-IV, recurrence, decreased
III/IV * * N/A [106]
thyroid cancer 7/119 sequencing (Korea) OS and DFS with BRAF/RAS mutations: RAS/BRAF >TERTp >
RAS/BRAF+TERTp.
Associated with older age and poor prognosis.
DNA
Follicular 14% 38.5% 62.5% Increased cytoplasmic TERT.
Yes sequencing, Patients (Italy) [102]
thyroid cancer 8/58 3/8 5/8 No impact of rs2853669 TERT -245 A>G polymorphism on
WB, and IHC
outcome.
Total
53/381 10/46 36/46
Follicular
(13.9%) (21.7%) (78.2%)
thyroid cancer
Poorly DNA
21% 33.3 66.7 Patients
differentiated Yes sequencing, [52]
3/14 1/3 2/3 (Portugal)
thyroid cancer qRT-PCR, IHC
Poorly
37.5% 0% 100% DNA
differentiated N/A Patients Only in malignant lesions. [108]
3/8 0/3 3/3 sequencing
thyroid cancer
Poorly
29% 50% 50% DNA Patients No TERTp mutation found in 192 well-differentiated cancers
differentiated N/A [105]
2/7 1/2 1/2 sequencing (Korea) without distanst metastasis.
thyroid cancerCells 2020, 9, 749 14 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Poorly
51.7% 40% 60% DNA Patients More prevalent in advanced cancer patients with
differentiated N/A [100]
30/58 12/30 18/30 sequencing (US & Japan) BRAF/RASmut .
thyroid cancer
Total Poorly
38/87 14/38 24/38
differentiated
(43.7%) (36.8%) (63.2%)
thyroid cancer
Anaplastic 46.3% 8% 92% DNA
N/A Patients Only in malignant lesions. [108]
thyroid cancer 25/54 2/25 23/25 sequencing
DNA
Anaplastic 13% 50% 50% Patients
Yes sequencing, [52]
thyroid cancer 2/16 1/2 1/2 (Portugal)
qRT-PCR
Anaplastic 50% 0% 100% DNA Patients More prevalent in advanced cancer patients with
N/A [100]
thyroid cancer 10/20 0/10 10/10 sequencing (US & Japan) BRAF/RASmut .
Anaplastic 50% 20% 80% DNA Patients
N/A PTC: 27% (25/332); FTC: 22% (12/70); ATC: 50% (12/36). [98]
thyroid cancer 10/20 2/10 8/10 sequencing (Sweden)
Associated with older age, larger tumor size, distant
Patients metastases and disease-related death in FTC.
Anaplastic 33.3% DNA
* * N/A (Portugal & PTC: 7.5% (25/332); FTC: 17.1% (12/70); PDTC: 29% (9/31); ATC: [101]
thyorid cancer 12/36 sequencing
Spain) 33.4% (12/36).
PTC associated with BRAF-V600E mutation in 60.3% of cases.
Anaplastic 38.7% 10% 90% DNA Patients Associated with older age and distal metastases.
N/A [104]
thyroid cancer 41/106 4/41 37/41 sequencing (US & China) −124 C>T found in 56.3% of BRAF-V600E mutated cases.
Total
100/252 9/88 79/88
anaplastic
(39.7%) (10.2%) (89.7%)
thyroid cancer
Thyroid
91.7% 27.3% 72.7% DNA
Cancer cell N/A Cell lines [108]
11/12 3/11 8/11 sequencing
lines
Thyroid
75% 17.7% 83.3% DNA
Cancer cell N/A ATC cell lines [98]
6/8 1/6 5/6 sequencing
lines
Liver-Hepatocellular Carcinoma (HCC)
31.4% 18,2% 81,8% DNA Patients
HCC N/A [57]
11/35 2/11 9/11 sequencing (China)
Patients
34% 33.3% 66.7% DNA Higher TERTp mutation prevalence in African (53%) compared
HCC N/A (Africa, Asia, [97]
15/44 5/15 10/15 sequencing to non-African (24%) populations.
Europe)
Patients
44.3% 3.7% 96.3% DNA Detected in both HBV-associated and HBV-independent HCC
HCC N/A (US [77]
27/61 1/27 26/27 sequencing Frequent in HCV-associated HCC.
American)Cells 2020, 9, 749 15 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
41% of mutations in HBV-associated HCC.
48.5% 3.1% 96.9% DNA 53.6% mutations in HCV-associated HCC.
HCC N/A Patients (Italy) [95]
65/131 2/65 63/65 sequencing All heterozygous.
No −57 A>C.
DNA HBV-associated HCC.
31% 1.1% 98.9% Patients
HCC Yes sequencing, Correlated with age, not with HBV status. [63]
85/275 1/85 84/85 (China)
IHC Found in 4/7 preneoplastic lesions (HBV-associated HCC).
Associated with older age.
65.4% 3% 97% DNA Patients Associated with shorter OS and DFS.
HCC Yes [122]
68/104 2/68 66/68 sequencing (Japan) Associated with HCV infection and excluded from HBV+
HCC.
Detected in cirrhotic preneoplastic macronodules (25%) and
DNA
58.6% 6.1% 92.7% Yes Patients cirrhotic adenomas (44%), at last step of malignant
HCC sequencing, [62]
179/305 11/179 166/179 2–10-fold (French) transformation into HCC.
qRT-PCR
Absent from HBV-associated tumors 2/179 (1%) −146 C>T.
Associated with older age.
DNA
29.3% 5.3% 94.7% No impact on overall survival.
HCC No sequencing, [96]
57/195 3/57 54/57 Excluded from HBV-associated HCC.
qRT-PCR
Higher frequency in HCV-associated HCC.
−57 A>C mutation detected in 1.6%.
Patients Present in 37% HBV-associated HCC but mutually exclusive
54% 4.3% 93% DNA (Japan, with HBV sequence integration.
HCC N/A [121]
254/469 11/254 236/254 sequencing US-European Mutually exclusive with TERT CNV and ATRX mutations.
ancestry) Associated with HCV infection (64% or TERTp mutations).
Associated with Wnt pathway mutations.
60% 11.1% 88.9% DNA
HCC N/A Cell lines [97]
9/15 1/9 8/9 sequencing
770/1634 39/770 722/770
Total HCC
(47.1%) (5%) (93.7%)
Cervical
Patients
21.8% 31.8% 45.5%
Cervical SCC Yes qRT-PCR (Italian [37]
22/101 7/22 10/22
women)
DNA
sequencing, Patients 75% TERTp mutations in HPV-negative samples.
21.4% 26.7% 73.3%
Cervical SCC N/A Association (Indian −124 C>T 6/22 were TT homozygous. [36]
30/140 8/30 22/30
with clinical women) −146 C>T 2/8 were TT homozygous.
statusCells 2020, 9, 749 16 of 28
Prevalence of Tert Sample
Cancer Type Stage −146 C>T −124 C>T Methods Remarks Ref.
Mutations Upregulation Origin
Patients
4.5% 100% 0% DNA
Cervical SCC N/A (US 1 patient with −125 C>A mutation.
1/22 1/1 0/1 sequencing
American)
Total Cervical 53/263 16/53 32/53
[77]
SCC (20.1%) (30.2%) (60.4%)
Head and Neck Squamous Cell Carcinoma (HNSCC)
Patients
31.7% 30.8% 69.2% DNA
HNSCC N/A (Indian Association with clinical status. [36]
13/41 4/13 9/13 sequencing
women)
Patients 11/12 HNSCC with TERTp mutations were in the oral tongue,
17% 16.7% 83.3% DNA
HNSCC N/A (US and 11/23 (47.8%) of HNSCC of the oral tongue harbored [77]
12/70 2/12 10/12 sequencing
American) TERTp mutations.
25/111 6/25 19/25
Total HNSCC
(22.5%) (24%) (76%)
Ovarian cancer
Patients
Ovarian clear 15% 0% 10% DNA
N/A (US 1 patient with −124 C>A mutation. [77]
cell carcinoma 3/20 0/3 2/3 sequencing
American)
DNA
sequencing, No link with survival or age.
Ovarian clear 16.5% 8.1% 91.9%
N/A IHC, telomere Patients TERTp mutations tended to be mutually exclusive with loss of [132]
cell carcinoma 37/233 3/37 34/37
length ARID1A protein expression and PIK3CA mutation.
evaluation
Ovarian clear 30% 0% 100%
Yes qRT-PCR Cell lines [132]
cell carcinoma 3/10 0/3 3/3
Total ovarian
43/263 3/43 39/43
clear cell
(16.3%) (6.9%) (90.7%)
carcinoma
N/A: not assessed; *: data not available. TERT: telomerase reverse transcriptase; GBM: glioblastoma multiforme; SCC: squamous cell carcinoma; HNSCC: head and neck squamous cell
carcinoma; HCC: hepatocellular carcinoma; GI: gastrointestinal; UC: urothelial cancer; MPC: micropapillary carcinoma; HPV: Human papilloma virus; HBV: Hepatitis B virus; HCV:
Hepatitis C virus; PTC: papillary thyroid cancer; FTC: follicular thyroid cancer; ATC: anaplastic thyroid cancer.Cells 2020, 9, x FOR PEER REVIEW 14 of 25
4. Cancer Bias of TERTp Mutations
Cells 2020, 9, 749 17 of 28
TERTp mutations have been recorded in individuals of Caucasian, African, and Asian descent,
with no race-related bias. The −124 C>T mutation has an overwhelmingly higher prevalence than the
4. Cancer Bias of TERTp Mutations
−146 C>T mutation in all cancers, with the exception of skin cancers, where both hotspots are mutated
withTERTp mutations
comparable have been
frequencies recorded
(Figure 2 andinTable
individuals of Caucasian,
1). Although African,
both −124C>T andand Asianmutations
−146C>T descent,
with no race-related
generate bias. The −124
identical sequences, enable C>T mutation
binding has an overwhelmingly
of GABPA, higher prevalence
and are equally efficient thanTERT
in increasing the
−146 C>T mutation
transcription in [57,69],
in vitro all cancers, withthe
in vivo, the−exception of skin cancers,
124 C>T mutation where both
was associated withhotspots are mutated
higher TERT mRNA
with comparable
in GBM [57,112].frequencies
This would (Figure
suggest 2 and
thatTable 1). Although
the Ets/TCF both
binding −124C>T
site and−−146C>T
at position 124 providesmutations
a more
generate
favorable identical sequences,
or accessible hotspotenable binding
for the of GABPA,
transcriptional and are equally
machinery efficient
[109]. The in increasing TERT
overrepresentation of the
transcription in vitroin
−146 C>T mutation [57,69], in vivo,hints
skin cancers the −124 C>T mutation
at different etiologieswasofassociated with higher
TERTp mutations. TERT
TERTp mRNA
mutations
ininGBM [57,112].and
melanoma This would suggestskin
non-melanoma thatcancers
the Ets/TCFhavebinding to UV−124
site at position
been attributed provides
damage a more
[49,51,55,88–
favorable or accessible
91,116], which triggershotspot for the transcriptional
CT transitions machinery
at CC dinucleotides [109]. The
[55,127]. overrepresentation
Nevertheless, of the
CT transitions
−146
whereC>T mutation
C is preceded in skin
by Ccancers hints attodifferent
also conform etiologies
the preferred of TERTp
target mutations. TERTp
of Apolipoprotein B mRNA mutations
Editing
inCatalytic
melanomaPolypeptide-like
and non-melanoma skin cancers have
(APOBEC)3A/B been attributed
de-aminations andto UV
to damage [49,51,55,88–91,116],
aging mutations [127,133].
which
APOBEC3triggers C→T transitions
mutations are highlyatprevalent
CC dinucleotides
in ovarian [55,127]. Nevertheless,cervical
and HPV-associated C→T transitions where
and oral SCC C
[125–
is127],
preceded
as wellbyasC in
also
HCCconform
and intocirrhotic
the preferred
lesionstarget of Apolipoprotein
[121,134]. A role for APOBEC B mRNA andEditing Catalytic
aging-associated
Polypeptide-like
de-aminations is (APOBEC)3A/B
consistent with de-aminations
potentially and to agingaccessibility
increased mutations [127,133].
of the −124APOBEC3
positionmutations
to DNA
binding
are highlyproteins
prevalentandinwith the association
ovarian of TERTp cervical
and HPV-associated mutations andwith
oralolder
SCCage at diagnosis
[125–127], in as
as well GBM,
in
melanoma,
HCC and in and PTC [52,57,60,63,64,77,79,80,82,86,88,98,100–102].
cirrhotic lesions [121,134]. A role for APOBEC andThese observationsde-aminations
aging-associated therefore raise
isthe possibility
consistent withthat UV-driven
potentially lesions account
increased for TERTp
accessibility of themutations in skin
−124 position cancers,
to DNA while proteins
binding APOBEC
andwith
and age-driven de-aminations
the association of TERTpaccount
mutations for with
the older
−124 age
C>Tatmutation
diagnosisin in other cancers. Further
GBM, melanoma, and
epidemiological
PTC and mechanistic studies are needed
[52,57,60,63,64,77,79,80,82,86,88,98,100–102]. Theseto shed light on this
observations point. raise the possibility
therefore
The −139/−lesions
that UV-driven 138 CC>TT tandem
account mutation
for TERTp is very in
mutations infrequent, limited
skin cancers, whileto skin
APOBECcancers,
andand has been
age-driven
associated with
de-aminations lowerfor
account DFS.
the This
−124 tandem mutation
C>T mutation has been
in other cancers.suggested
Furthertoepidemiological
favor chromosomal and
instability [51].
mechanistic studies are needed to shed light on this point.
Figure2.2.Distribution
Figure DistributionofofTERT
TERTpromoter
promotermutations
mutationsinindifferent
differentcancers.
cancers.Cells 2020, 9, 749 18 of 28
The −139/−138 CC>TT tandem mutation is very infrequent, limited to skin cancers, and has
been associated with lower DFS. This tandem mutation has been suggested to favor chromosomal
instability [51].
5. Exclusiveness of TERTp Mutations
Aside from non-melanoma skin cancers [90], TERTp mutations are mostly monoallelic. This
suggests that TERT reactivation on one allele is probably sufficient to ensure telomere maintenance or
elongation in cancer cells [54]. In line with this observation, TERTp mutations appear to be mutually
exclusive [50]. Likewise, TERTp mutations are generally absent from cancers where telomere elongation
is ensured by ALT [77,79,80,98] or TERT copy-number duplications [38,121]. TERTp mutations are also
less frequent in cancers where viral transformation or viral oncogenes reactivate TERT transcription,
such as HBV-DNA or high-risk HPV16/18 E6 [30,32,33,36,37,62,95,96,121,122]. These observations
reinforce the concept that, despite some exceptions [38,89,111,117], tumors generally rely on one
mechanism for telomere maintenance. The reasons for such selectivity remain speculative to date.
One possible explanation is that there is a threshold for TERT expression, above which the biological
advantage is lost.
Consistent with this view, Phosphatidyl Inositol Kinase 3 (PIK3) CA and PIK3 Receptor 1 (PIK3R1)
mutations are recorded in 50% of GBM with wt TERTp and tend to be mutually exclusive with TERTp
mutations in ovarian clear cell carcinoma [79,86,132]. The PIK3CA/Akt signaling pathway is involved in
cellular self-renewal in embryonic stem cells and cancer stem cells [135], as well as in TERT Ser227 and
Ser824 phosphorylation, subsequent nuclear translocation, and cellular transformation [25–28]. Mutual
exclusion of PIK3CA and TERTp mutations suggests that activation of the PIK3CA/Akt pathway or of
TERT confer cells a similar growth and proliferative advantage. In the absence of TERT reactivation,
other telomere maintenance mechanisms, such as ALT, can achieve immortalization [27]. Indeed,
TERT also contributes to cell survival and proliferation through telomere-independent mechanisms;
it facilitates Wnt/β-catenin-dependent [136,137], c-myc-dependent [138,139], and NF-κB-dependent
gene transcription [140,141], thereby sustaining both oncogenic signaling pathways and its own
transcription in a feedforward loop [29,142]. It also regulates methylation [48,143] and DNA damage
responses [144,145], and protects cells from Endoplasmic Reticulum (ER) stress and apoptosis by
buffering Reactive Oxygen Species (ROS) and modulating mitochondrial function [145–151]. It is
highly likely that TERT homeostasis is also tuned by these functions within a given tumor type and
microenvironment, and by related metabolic alterations that need to be preserved.
6. Discussion
Hints for a model come from the observation that overall, TERTp mutations
are associated with late-stage disease in GBM, melanoma, urothelial, and thyroid
carcinoma [49,52,60,61,66,85,98,100,101,103–105,112,118] and with the last steps of hepatocellular
transformation [62,95]. They often occur with or after mutations in pathways associated with cell
growth and proliferation. In GBM, TERTp mutations coexist with EGFR amplification [64,77,111], and
in urothelial bladder carcinoma, they are associated with FGFR3 (Fibroblast Growth Factor Receptor 3)
mutations [61,94]. In ~50% of melanoma, urothelial, and thyroid cancers, TERTp mutations coexist with
the common BRAF-V600E mutation [52,88,89,105,106,108,116,152]. GFR and BRAF/RAS kinases control
the MAPK and PI3K-Akt pathways that lead to cell growth, survival, and angiogenesis. Constitutive
activation of the GFR/FGFR-BRAF/RAS pathway leads to constitutive cell growth and division [153].
Mutations in these oncogenes are often detectable in low-grade tumors and probably precede TERTp
mutations [22,61,77,112]. The picture is even more clear-cut in HCC, where mutations in β-catenin
(CTNNB1) neatly precede TERTp mutations during the process of malignant transformation [62,95,120].
β-catenin is involved in cell adhesion and interacts with Wnt, promoting cell growth and division.
The proliferative advantage conferred by driver mutations in these pathways leads to accelerated
telomere erosion. Accordingly, most tumors display telomere dysfunction and shortened telomeres,Cells 2020, 9, 749 19 of 28
which leads to chromosome instability [10,22,61,66,98,112,115]. In this scenario, TERT reactivation
regenerates telomeres sufficiently to maintain them above the critical threshold and to stabilize the
tumor genome [3,18,145]. This interpretation is consistent with the association of TERTp mutations with
shortened telomeres and with age as in PTC, melanoma, and GBM/glioma, since cells from younger
patients or with sufficiently long telomeres do not need to rely on telomerase reactivation to overcome
telomeric crisis [10,29,57,77,85,98,101,115]. Partial telomere healing is coherent with a modest increase
in TERT expression (2- to 4-fold) and with a single genetic mechanism of telomere elongation. It likely
reflects an exquisite balance between escape from apoptosis resulting from telomere attrition and
genomic instability, and cell sustainability in terms of oxygen and nutrient supplies.
Intriguingly, it was recently reported that GABPA controls the cell cycle and induces cell
differentiation, thus acting as a tumor suppressor regulating cell proliferation, stemness, and adhesion.
It decreased tumor invasiveness and distal metastases in PTC, HCC, and bladder carcinoma [154–156].
GABPA levels were decreased and even negatively associated with TERT expression in PTC [154–156].
One possible explanation is that other Ets/TCF family transcription factors bind TERTp mutations.
Alternatively, the decrease in GABPA expression may follow rather than precede TERTp mutations.
In this case, it would be a cellular adaptation which confers a selective advantage to TERTp-mutated (and
GFR/BRAF/RAS-mutated) cells by containing TERT reactivation within sustainable limits. Decreased
GABPA could also be an adaptation to the TERT-induced proliferation, stemness, and invasion to
avoid contradictory signals. Further studies establishing the order of emergence of these mutations
would be needed to shed light on this matter.
Taken together, these observations point to a fine tuning of TERT homeostasis and suggest that
there is a narrow kinetic and quantitative window for TERT expression. Below that window, cells
succumb to telomere crisis and DNA damage. Above that window, cells succumb to overwhelming
genetic alterations or metabolic needs. This frailty could be exploited through strategies aiming to
push cells either way beyond the threshold of TERT tolerability.
7. Concluding Remarks
TERTp mutations have only been described recently; however, they have prompted an impressive
number of studies which draw a comprehensive picture of their prevalence across cancers, as well
as providing clues on their mechanisms of action and their associated constraints. They have been
proposed as potential biomarkers with predictive and treatment-orienting value. However, more
structured studies are needed to validate their clinical potential, particularly since they appear at
different stages in different malignancies, ranging from preneoplastic cirrhotic lesions to late stage
GBM or melanoma with distal metastases. Cancer cells only require one mechanism of telomere
maintenance. This underscores the key role of telomere stabilization in the process of transformation,
as well as the necessity of maintaining an exquisitely balanced TERT homeostasis to achieve tumor
cell selection, adaptation, and sustainability. TERT is a target of choice in antitumor strategies due
to its reactivation in numerous cancers. A better understanding of TERT regulation, homeostasis,
and functions could help to overcome the shortcomings of prior genetic and immunotherapy-based
approaches targeting TERT.
Author Contributions: F.H. and D.P.B. wrote the manuscript and approved the final version. All authors have
read and agreed to the published version of the manuscript.
Funding: This work was supported by the Ministère de l’Education et de la Recherche du Luxembourg;
FH is supported by the Fonds National de la Recherche du Luxembourg FNR-PRIDE scheme
(PRIDE/11012546/NEXTIMMUNE).
Acknowledgments: The authours are deeply grateful to Dr Jonathan D Turner for his thorough revision of this
manuscript and for his insightful advise.
Conflicts of Interest: The authors declare no conflict of interest.You can also read

















































