Atorvastatin attenuates TGF β1 induced fibrogenesis by inhibiting Smad3 and MAPK signaling in human ventricular fibroblasts
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
INTERNATIONAL JOURNAL OF MOlecular medicine 46: 633-640, 2020
Atorvastatin attenuates TGF‑β1‑induced fibrogenesis by inhibiting
Smad3 and MAPK signaling in human ventricular fibroblasts
Yanfei Du1*, Haiying Xiao2*, Jun Wan3, Xinyu Wang2, Tao Li2, Shuzhan Zheng1,
Jian Feng1, Qiang Ye1, Jiafu Li1, Guang Li2 and Zhongcai Fan1
1
Key Laboratory of Medical Electrophysiology, Ministry of Education,
Department of Cardiology, The Affiliated Hospital of Southwest Medical University;
2
Key Laboratory of Medical Electrophysiology, Ministry of Education, Institute of Cardiovascular Research,
Southwest Medical University; 3Department of Basic Medical Sciences, College of Basic Medical Sciences,
Southwest Medical University, Luzhou, Sichuan 646000, P.R. China
Received November 20, 2019; Accepted May 6, 2020
DOI: 10.3892/ijmm.2020.4607
Abstract. Excessive proliferation and myofibroblasts signal‑regulated kinase 1/2, p38 MAPK and c‑Jun N‑terminal
transformation of cardiac fibroblasts play a critical role kinase. The results indicated that ATV significantly prevented
in the process of cardiac fibrosis. Atorvastatin (ATV), a TGF‑β1‑induced cell proliferation, myofibroblast differentia-
3‑hydroxy‑3‑methyl‑glutaryl‑coenzyme A reductase inhibitor, tion and production of extracellular matrix proteins, such as
is commonly used to treat hypercholesterolemia. It has previ- matrix metalloproteinase‑2, collagen I and collagen III, in
ously been shown that ATV has potential anti‑fibrotic effects. hVFs. Furthermore, ATV effectively inhibited TGF‑β1‑induced
However, the underlying mechanisms of ATV against cardiac activation of Smad3 and MAPK signaling in hVFs. In conclu-
fibrosis remain to be fully elucidated, and to the best of our sion, the present results demonstrated that ATV prevented
knowledge, there are no reports focusing on the effects of ATV TGF‑β1‑induced fibrogenesis in hVFs, at least in part by inhib-
on transforming growth factor‑β1 (TGF‑β1)‑induced human iting the Smad3 and MAPK signaling pathways. Therefore,
ventricular fibroblasts (hVFs) activation. In the present study, these results imply that ATV may be a promising agent to treat
hVFs were stimulated with TGF‑β1 with or without pretreat- myocardial fibrosis.
ment with ATV. Subsequently, hVF proliferation, cytotoxicity,
myofibroblast differentiation and pro‑fibrotic gene expres- Introduction
sion were assessed. Canonical and non‑canonical signaling
downstream of TGF‑β1, such as Smad3 and mitogen‑activated Myocardial fibrosis is a common pathological feature
protein kinase (MAPK) signaling, were investigated by of multiple end‑stage cardiovascular diseases, including
evaluating the phosphorylation levels of Smad3, extracellular hypertension, advanced coronary heart disease and cardio-
myopathy (1). Fibrosis is defined by overproliferation and
activation of cardiac fibroblasts (CFs), and accumulation of
extracellular matrix (ECM) components secreted by activated
fibroblasts (2,3). Therefore, prevention of CF proliferation and
Correspondence to: Professor Zhongcai Fan, Key Laboratory of abrogation of CF trans‑differentiation into myofibroblasts may
Medical Electrophysiology, Ministry of Education, Department
become an effective strategy for treating cardiac fibrosis.
of Cardiology, The Affiliated Hospital of Southwest Medical
Transforming growth factor‑ β1 (TGF‑ β1) is a major
University, 25 Taiping Street, Luzhou, Sichuan 646000, P.R. China
E‑mail: fanzhongcaixnyd@126.com pro‑fibrotic factor (2,4). It can stimulate the proliferation of
CFs and the differentiation of CFs into myofibroblasts, which
Professor Guang Li, Key Laboratory of Medical Electrophysiology, are characterized by the upregulation of α‑smooth muscle actin
Ministry of Education, Institute of Cardiovascular Research,
(α‑SMA) and the secretion of ECM proteins, such as collagen I
Southwest Medical University, 319 Zhongshan Road, Luzhou,
and collagen III (5,6). TGF‑β1 ligand initiates a signaling
Sichuan 646000, P.R. China
E‑mail: liguang7645@126.com cascade through cell‑surface receptors and intracellular Smad
signal proteins, such as the canonical signal transducer‑Smad2/3
*
Contributed equally protein, whose activation is associated with the transcription of
numerous pro‑fibrotic genes (7,8). Furthermore, TGF‑β1 can
Key words: cardiac fibrosis, human ventricular fibroblasts, induce other non‑canonical signaling cascades independently
transforming growth factor‑β1, atorvastatin, extracellular matrix, of Smads, such as mitogen‑activated protein kinase (MAPK)
signaling pathways signaling pathways, including extracellular signal‑regulated
kinase 1/2 (ERK1/2), p38 MAPK and c‑Jun N‑terminal kinase
(JNK) (9). Accumulating evidence has demonstrated that634 DU et al: ATORVASTATIN ATTENUATES TGF-β1-INDUCED FIBROGENESIS IN HUMAN VENTRICULAR FIBROBLASTS
MAPK signaling plays a critical role in ECM synthesis and in Next, the absorbance of each well at 450 nm was examined using
the proliferation of CFs (10,11). Thus, pharmacological inter- an automatic microplate reader (TECAN M200; Tecan Group,
ventions in these signaling pathways could be considered as a Ltd.). The results were expressed as percentages of the control
promising therapeutic strategy against cardiac fibrosis. group (cells were treated with an equal amount of DMSO).
Statins, which are inhibitors of the 3‑hydroxy‑3‑methyl‑
glutaryl‑coenzyme A reductase, have been extensively applied Cell proliferation assay. hVFs were seeded into 96‑well plates
to treat patients with hypercholesterolemia in the past decade at 2x103 cells/well and cultured overnight. Cells were deprived
due to their lipid‑reducing effects (12). Furthermore, statins of serum for 16 h before being pretreated with ATV (2, 5
have also been reported to possess pleiotropic cardiovascular and 10 µM) or DMSO for 3 h, followed by stimulation with
effects, including reduced oxidative stress and inflammation, TGF‑β1 (5 ng/ml) for 24 h in a humidified atmosphere with
decreased platelet adhesion, improved endothelial function 5% CO2 at 37˚C. Subsequently, the medium was removed from
and enhanced atherosclerotic plaque stability (13‑15). In addi- each well, and CCK‑8 solution (10 µl/well) was added into the
tion, previous studies have shown that atorvastatin (ATV), a wells, followed by incubation for an additional 4 h at 37˚C.
member of the statin family, exerts potential anti‑myocardial Finally, the optical density of each well was detected at an
fibrotic properties (16‑20). Nevertheless, to the best of our absorbance wavelength of 450 nm using the aforementioned
knowledge, the molecular mechanism involved in the effect microplate reader.
of ATV on cardiac fibrosis remains to be clarified, and limited
information is available concerning the possible impact of Western blotting. hVFs (5x105 cells/well) were plated into
ATV on TGF‑β1‑induced human ventricular fibroblast (hVF) 6‑well plates, cultured to ~80% confluence and starved of
proliferation and myofibroblast differentiation. Thus, the aim serum for 16 h. Then, the cells were pretreated for 3 h with
of the present study was to investigate the impact of ATV on ATV (10 µM) or DMSO before treatment with 5 ng/ml
TGF‑β1‑stimulated fibrogenesis and its underlying molecular TGF‑β1 for 24 h at 37˚C. Subsequently, the cells were washed
mechanism in hVFs. The findings indicated that ATV prevents twice with cold PBS and lysed in RIPA lysis buffer (Beyotime
TGF‑ β1‑induced fibrogenesis in hVFs, at least in part, by Institute of Biotechnology) at 4˚C. Lysates were centrifuged at
inhibiting the Smad3 and MAPK signaling pathways. 13,000 x g for 15 min at 4˚C, and protein concentrations were
determined by BCA assay (cat. no. 23227; Pierce; Thermo
Materials and methods Fisher Scientific, Inc.). Cell lysates (30 µg) were separated
by 8‑10% SDS‑PAGE and transferred onto a polyvinylidene
Cells, chemicals and reagents. Adult hVFs (cat. no. 6310) difluoride membrane (Merck KGaA). The membranes were
and complete medium for their culture (fibroblast medium‑2; blocked with blocking buffer (5% nonfat dry milk in TBS
FM‑2; cat. no. 2331) were obtained from ScienCell Research plus 0.1% Tween‑20) for 1 h at room temperature and incu-
Laboratories, Inc. Antibodies against α‑SMA, matrix metal- bated with primary antibodies overnight at 4˚C, including
loproteinase‑2 (MMP‑2), collagen I and collagen III were antibodies against α‑SMA (1:1,000; cat. no. 14395‑1‑AP),
purchased from ProteinTech Group, Inc. Antibodies against MMP‑2 (1:1,000; cat. no. 10373‑2‑AP), collagen I (1:2,000; cat.
phosphorylated (p)‑Smad3, total (t)‑Smad3, p‑ERK1/2, no. 14695‑1‑AP), collagen III (1:1,000; cat. no. 22734‑1‑AP),
t‑ERK1/2, p‑JNK, t‑JNK, p‑p38 and t‑p38 were purchased p‑Smad3 (1:1,000; cat. no. 9520), t‑Smad3 (1:1,000; cat.
from Cell Signaling Technology, Inc. Anti‑GAPDH antibody no. 9523), p‑ERK1/2 (1:1,000; cat. no. 9101), t‑ERK1/2
and Cell Counting Kit‑8 (CCK‑8) were purchased from (1:1,000; cat. no. 9102), p‑JNK (1:1,000; cat. no. 4671), t‑JNK
Beyotime Institute of Biotechnology. ATV was obtained from (1:1,000; cat. no. 9252), p‑p38 (1:1,000; cat. no. 4511), t‑p38
Sigma‑Aldrich (Merck KGaA). Recombinant human TGF‑β1 (1:1,000; cat. no. 9212) and GAPDH (1:5,000; cat. no. AG019).
cytokine was purchased from PeproTech, Inc. Following washing three times with washing buffer (TBS
containing 0.1% Tween‑20), the membranes were incubated
Cell culture. hVFs were maintained in FM‑2 supplemented with horseradish peroxidase (HRP)‑conjugated secondary
with 5% fetal bovine serum (cat. no. 0025; ScienCell Research antibodies [HRP‑conjugated goat anti‑rabbit IgG, 1:2,000, cat.
Laboratories, Inc.) and 1% penicillin‑streptomycin in a humidi- no. D110058; HRP‑conjugated goat anti‑mouse IgG, 1:2,000,
fied atmosphere with 5% CO2 at 37˚C. The cells were passaged cat. no. D110087; all from Sangon Biotech (Shanghai) Co.,
≤8 times and were cultured for 16 h in serum‑free medium Ltd.] for 1 h at 25˚C. The protein signals were detected using
before treatment. In the present study, an in vitro cardiac Immobilon ECL HRP Substrate (Merck KGaA), and images
fibrosis model was established by treatment of hVFs with were acquired using Chemidoc XRS (Bio‑Rad Laboratories,
TGF‑β1 (5 ng/ml) for 24 h at 37˚C in a humidified atmosphere Inc.). Image Pro‑Plus 6.0 software (Media Cybernetics, Inc.)
with 5% CO2, according to the previous literature (3,21). was used to quantify the density of the bands.
Cell viability assays. The survival rate of the cells was deter- Immunofluorescence staining. In order to detect the effect of
mined by CCK‑8 assay according to the manufacturer's protocol. ATV on the trans‑differentiation of hVFs into activated myofi-
In brief, fibroblasts (1x104 cells/well) were seeded into 96‑well broblasts, cells were stained with the fibroblast activation marker
plates until ~90% confluence. Subsequently, cells were subjected α‑SMA. The fibroblasts (1x104 cells/well) were cultured on
to serum starvation for 16 h, followed by treatment with ATV coverslips in the plate and were subjected to different treatments
(0, 2, 5, 10, 20 and 30 µM) for 24 h at 37˚C. Following treat- as follows: i) Control group (DMSO treatment); ii) TGF‑β1
ment, the cells were washed with PBS, and then 10 µl CCK‑8 was group; iii) ATV group; and iv) TGF‑β1 combined with ATV
added to each well and incubated for an additional 4 h at 37˚C. group. Following treatment, cells were washed three timesINTERNATIONAL JOURNAL OF MOlecular medicine 46: 633-640, 2020 635
Table I. Sequences of primers used for reverse transcription‑quantitative PCR.
Gene Forward sequence (5'‑3') Reverse sequence (5'‑3')
α‑SMA CTATGAGGGCTATGCCTTGCC GCTCAGCAGTAGTAACGAAGGA
MMP‑2 GATACCCCTTTGACGGTAAGGA CCTTCTCCCAAGGTCCATAGC
COL‑I GAGGGCCAAGACGAAGACATC CAGATCACGTCATCGCACAAC
COL‑III GCCAAATATGTGTCTGTGACTCA GGGCGAGTAGGAGCAGTTG
GAPDH GGAGCGAGATCCCTCCAAAAT GGCTGTTGTCATACTTCTCATGG
α‑SMA, α‑smooth muscle actin; MMP‑2, matrix metalloproteinase‑2; COL‑I, type I collagen; COL‑III, type III collagen.
with cold PBS, treated with 4% paraformaldehyde for 20 min
at room temperature and permeabilized with 0.2% Triton‑X
100 for 15 min at room temperature. Following blocking with
5% bovine serum albumin [cat. no. A602449; Sangon Biotech
(Shanghai) Co., Ltd.] in PBS for 1 h at room temperature, the
slides were stained with the aforementioned primary antibody
against α‑SMA (1:100) overnight at 4˚C. Subsequently, an
Alexa Fluor 488‑conjugated secondary antibody (1:400; cat.
no. A11008; Invitrogen; Thermo Fisher Scientific, Inc.) was
used to probe the primary antibody, and the nuclei were stained
with DAPI at room temperature for 5 min. Lastly, the cells were
covered with antifade mounting medium (cat. no. MM1401;
Shanghai Maokang Biotechnology Co., Ltd.), and images were
captured using an Olympus fluorescence microscope (Olympus
Corporation; magnification, x200).
Reverse transcription‑quantitative PCR (RT‑qPCR). Total
RNA was extracted from cells using NucleoZOL reagent (cat.
no. 740404.200; Macherey‑Nagel GmbH) and was reverse
transcribed into cDNA with PrimeScript™ RT Master mix (cat.
no. RR036A; Takara Biotechnology Co., Ltd.) according to the
manufacturer's protocol. Firstly, ordinary PCR was performed
using Rapid Taq Master mix (cat. no. P222; Vazyme Biotech Co.,
Ltd), according to the manufacturer's protocol, to ensure that
the PCR product of each primer pair was specific. The temper-
ature conditions were 95˚C for 3 min, followed by 30 cycles
of 95˚C for 15 sec, 60˚C for 15 sec and 72˚C for 15 sec, and a
final extension at 72˚C for 5 min. Then, qPCR was performed
Figure 1. Effect of ATV on TGF‑β1‑induced proliferation of hVFs. (A) Cells
on a Bio‑Rad CFX Connect Real‑Time PCR Detection system were exposed to different concentrations of ATV, and then the viability of
(Bio‑Rad Laboratories, Inc.) using SYBR Green Supermix (cat. the cells was assessed using a CCK‑8 assay. (B) hVFs were pretreated with
no. RR820A; Takara Biotechnology Co., Ltd.). The thermocy- various concentrations of ATV (0, 2, 5 and 10 µM) for 3 h, and then exposed
cling conditions for qPCR were as follows: Initial denaturation to 5 ng/ml TGF‑β1 for 24 h. The proliferation of hVFs was then assessed by
CCK‑8 assay. All data are presented as the mean ± standard error of mean of
at 95˚C for 30 sec; followed by 40 cycles of 95˚C for 5 sec, 60˚C three independent experiments and described as a percentage of the control
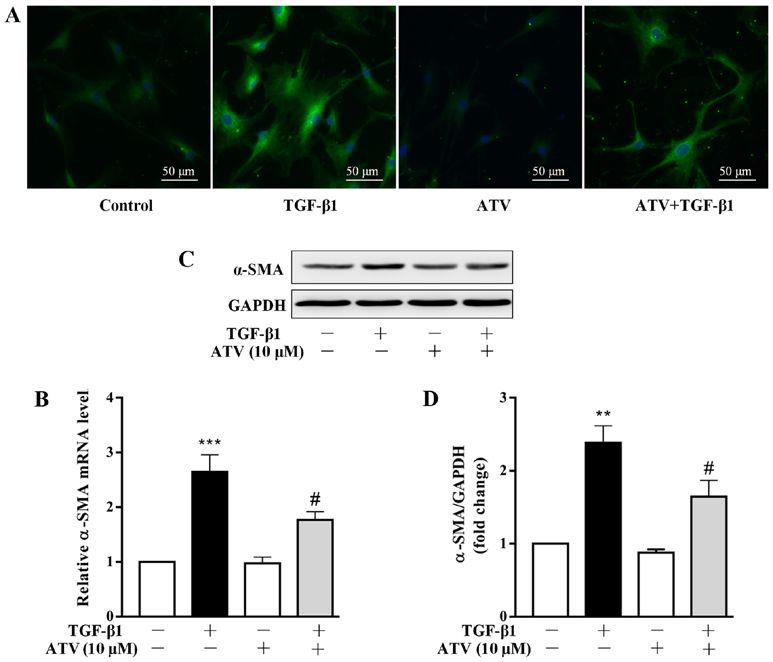
for 30 sec and 72˚C for 1 min. GAPDH was used as a reference group. *P636 DU et al: ATORVASTATIN ATTENUATES TGF-β1-INDUCED FIBROGENESIS IN HUMAN VENTRICULAR FIBROBLASTS Figure 2. ATV inhibits the trans‑differentiation of hVFs into myofibroblasts. Cells were exposed to ATV (10 µM) for 3 h and then co‑treated with 5 ng/ml TGF‑β1 for a further 24 h. (A) Cells were then immunostained with anti‑α‑SMA antibody (green) and stained with DAPI (nuclei; blue). Magnification, x200. (B) The α‑SMA mRNA level was assessed by reverse transcription‑quantitative PCR assay. (C) The protein expression of α‑SMA was measured by western blotting. (D) Quantitative analysis of α‑SMA protein level in hVFs. The relative value was normalized to GAPDH expression. The data are presented as the mean ± standard error of mean of three independent experiments. **P
INTERNATIONAL JOURNAL OF MOlecular medicine 46: 633-640, 2020 637 Figure 3. ATV inhibits TGF‑β1‑stimulated ECM production in hVFs. Fibroblasts were pretreated with ATV (10 µM) for 3 h and then co‑stimulated with 5 ng/ml TGF‑β1 for a further 24 h. (A) The mRNA levels of MMP‑2, type I collagen and type III collagen were assessed by reverse transcription‑quantitative PCR. (B) The protein expression levels of MMP‑2, collagen I and collagen III were detected by western blotting. (C) Quantitative analysis of the western blot results was performed by evaluating the protein band densities with Image Pro‑Plus version 6.0 software. The relative value was normalized to GAPDH expression. All data are expressed as the mean ± standard error of mean of three independent experiments. **P
638 DU et al: ATORVASTATIN ATTENUATES TGF-β1-INDUCED FIBROGENESIS IN HUMAN VENTRICULAR FIBROBLASTS Figure 4. ATV inhibits TGF‑β1‑induced Smad3 and MAPK signaling activation in hVFs. Fibroblasts were pre‑incubated with ATV (10 µM) for 3 h and then co‑treated with 5 ng/ml TGF‑β1 for 30 min. The protein levels of (A) p‑Smad3 and t‑Smad3, (B) p‑ERK1/2 and t‑ERK1/2, (C) p‑p38 and t‑p38, and (D) p‑JNK and t‑JNK were detected using western blotting. The relative density was expressed as the ratio of p‑protein to the corresponding t‑protein. Values represent the mean ± standard error of mean of three independent experiments. **P
INTERNATIONAL JOURNAL OF MOlecular medicine 46: 633-640, 2020 639
also been shown to play an essential role in the progression Patient consent for publication
of cardiac fibrosis. For example, ERK1/2 has been reported
to play an important signaling role in driving CF prolifera- Not applicable.
tion (44), and it is required for TGF‑β1‑induced pro‑fibrotic
phenotypes (3). Molkentin et al (11) reported that transgenic Competing interests
mice with fibroblast‑specific activation of p38 MAPK devel-
oped interstitial and perivascular fibrosis in the heart. Thus, The authors declare that they have no competing interests.
inhibiting these signaling pathways may imply a promising
therapeutic strategy against cardiac fibrosis. The present study References
evaluated the importance of Smad3 and MAPK signaling
in TGF‑ β1‑induced cardiac fibrosis in hVFs. The results 1. Rockey DC, Bell PD and Hill JA: Fibrosis‑a common pathway to
indicated that TGF‑β1 significantly activated both Smad3 organ injury and failure. N Engl J Med 373: 96, 2015.
2. Davis J and Molkentin JD: Myofibroblasts: Trust your heart and
and MAPK signaling in hVFs. However, ATV pretreatment let fate decide. J Mol Cell Cardiol 70: 9‑18, 2014.
significantly decreased the TGF‑β1‑induced phosphorylation 3. Scha fer S, Viswa natha n S, Widjaja A A, Lim W W,
levels of Smad3, ERK1/2, JNK and p38 MAPK in hVFs. This Moreno‑Moral A, DeLaughter DM, Ng B, Patone G, Chow K,
Khin E, et al: IL‑11 is a crucial determinant of cardiovascular
suggested that ATV inhibited TGF‑β1‑induced fibrogenesis fibrosis. Nature 552: 110‑115, 2017.
in hVFs at least in part through inhibition of the Smad3 and 4. Akhurst RJ and Hata A: Targeting the TGFβ signalling pathway
MAPK signaling pathways. in disease. Nat Rev Drug Discov 11: 790‑811, 2012.
5. Wynn TA: Cellular and molecular mechanisms of fibrosis.
There are also some limitations in the present study. J Pathol 214: 199‑210, 2008.
Firstly, the results were obtained only from a series of in vitro 6. Eghbali M, Tomek R, Woods C and Bhambi B: Cardiac fibro-
experiments, and were not validated in vivo. Secondly, the blasts are predisposed to convert into myocyte phenotype:
Specific effect of transforming growth factor beta. Proc Natl
exact targets of ATV in cardiac fibrosis remain unclear. Acad Sci USA 88: 795‑799, 1991.
Further investigations need to be performed to resolve these 7. Shi Y and Massague J: Mechanisms of TGF‑beta signaling from
limitations. cell membrane to the nucleus. Cell 113: 685‑700, 2003.
8. Schiller M, Javelaud D and Mauviel A: TGF‑beta‑induced SMAD
Taken together, the results of the present study report signaling and gene regulation: Consequences for extracellular
a protective role of ATV on TGF‑ β1‑induced fibrogenesis matrix remodeling and wound healing. J Dermatol Sci 35: 83‑92,
2004.
in hVFs and the potential mechanism involved. The results 9. Evangelia P and Peter TD: TGFβ signaling and cardiovascular
demonstrated that ATV prevented TGF‑β1‑induced fibrogen- diseases. Int J Bio Sci 8: 195‑213, 2012.
10. Chung CC, Kao YH, Yao CJ, Lin YK and Chen YJ: A compar-
esis in hVFs at least in part by inhibiting Smad3 and MAPK ison of left and right atrial fibroblasts reveals different collagen
signaling activation. These novel findings suggest a potential production activity and stress‑induced mitogen‑activated protein
therapeutic effect of ATV against fibrotic disease in clinical kinase signalling in rats. Acta Physiol (Oxf) 220: 432‑445,
2017.
practice. 11. Molkentin JD, Bugg D, Ghearing N, Dorn LE, Kim P, Sargent MA,
Gunaje J, Otsu K and Davis J: Fibroblast‑specific genetic manip-
Acknowledgements ulation of p38 mitogen‑activated protein kinase in vivo reveals
its central regulatory role in fibrosis. Circulation 136: 549‑561,
2017.
Not applicable. 12. McFarlane SI, Muniyappa R, Francisco R and Sowers JR:
Clinical review 145: Pleiotropic effects of statins: Lipid reduction
and beyond. J Clin Endocrinol Metab 87: 1451‑1458, 2002.
Funding 13. Wang CY, Liu PY and Liao JK: Pleiotropic effects of statin
therapy: Molecular mechanisms and clinical results. Trends Mol
This study was supported by The Natural Science Foundation Med 14: 37‑44, 2008.
14. Ludman A, Venugopal V, Yellon DM and Hausenloy DJ: Statins
of Southwest Medical University and The Foundation of The and cardioprotection‑more than just lipid lowering? Pharmacol
Affiliated Hospital of Southwest Medical University (grant Ther 122: 30‑43, 2009.
no. 2017-PT-43). 15. Liao JK: Effects of statins on 3‑hydroxy‑3‑methylglutaryl
coenzyme a reductase inhibition beyond low‑density lipoprotein
cholesterol. Am J Cardiol 96: 24F‑33F, 2005.
Availability of data and materials 16. Chen M, Li H, Wang G, Shen X, Zhao S and Su W: Atorvastatin
prevents advanced glycation end products (AGEs)‑induced
cardiac fibrosis via activating peroxisome proliferator‑activated
All data used or analyzed during the present study are available receptor gamma (PPAR‑γ). Metabolism 65: 441‑453, 2016.
from the corresponding author on reasonable request. 17. Choi SY, Park JS, Roh MS, Kim CR, Kim MH and Serebruany V:
Inhibition of angiotensin II‑induced cardiac fibrosis by atorvas-
tatin in adiponectin knockout mice. Lipids 52: 415‑422, 2017.
Authors' contributions 18. Fang T, Guo B, Xue L and Wang L: Atorvastatin prevents
myocardial fibrosis in spontaneous hypertension via inter-
YD, HX, JW, XW, TL, JF, SZ, QY and JL performed the leukin‑6 (IL‑6)/signal transducer and activator of transcription
3 (STAT3)/endothelin‑1 (ET‑1) pathway. Med Sci Monit 25:
experiments. ZF and GL conceived and designed the research. 318‑323, 2019.
YD, HX, GL and ZF analyzed the data and drafted the manu- 19. Wang Q, Cui W, Zhang HL, Hu HJ, Zhang YN, Liu DM and
script. QY and JL reviewed and edited the manuscript. All Liu J: Atorvastatin suppresses aldosterone‑induced neonatal
rat cardiac fibroblast proliferation by inhibiting ERK1/2 in
authors read and approved the final version of the manuscript. the genomic pathway. J Cardiovasc Pharmacol 61: 520‑527,
2013.
Ethics approval and consent to participate 20. Gao G, Jiang S, Ge L, Zhang S, Zhai C, Chen W and Sui S:
Atorvastatin improves doxorubicin‑induced cardiac dysfunction
by modulating Hsp70, Akt, and MAPK signaling pathways.
Not applicable. J Cardiovasc Pharmacol 73: 223‑231, 2019.640 DU et al: ATORVASTATIN ATTENUATES TGF-β1-INDUCED FIBROGENESIS IN HUMAN VENTRICULAR FIBROBLASTS
21. Xiao H, Ma X, Feng W, Fu Y, Lu Z, Xu M, Sheng Q, Zhu Y 34. Travers JG, Kamal FA, Robbins J, Yutzey KE and Blaxall BC:
and Zhang Y: Metformin attenuates cardiac fibrosis by inhibiting Cardiac fibrosis: The fibroblast awakens. Circ Res 118: 1021‑1040,
the TGFbeta1‑Smad3 signalling pathway. Cardiovasc Res 87: 2016.
504‑513, 2010. 35. Petrov VV, Fagard RH and Lijnen PJ: Stimulation of collagen
22. Livak KJ and Schmittgen TD: Analysis of relative gene expres- production by transforming growth factor‑beta1 during differen-
sion data using real‑time quantitative PCR and the 2(‑Delta Delta tiation of cardiac fibroblasts to myofibroblasts. Hypertension 39:
C(T)) method. Methods 25: 402‑408, 2001. 258‑263, 2002.
23. Yi X, Li X, Zhou Y, Ren S, Wan W, Feng G and Jiang X: 36. Li B, Chen H, Yang X, Wang Y, Qin L and Chu Y: Knockdown of
Hepatocyte growth factor regulates the TGF‑ β1‑induced eIF3a ameliorates cardiac fibrosis by inhibiting the TGF‑β1/Smad3
proliferation, differentiation and secretory function of cardiac signaling pathway. Cell Mol Biol (Noisy‑le‑grand) 62: 97‑101,
fibroblasts. Int J Mol Med 34: 381‑390, 2014. 2016.
24. Van Nieuwenhoven FA and Turner NA: The role of cardiac fibro- 37. Zhang M, Pan X, Zou Q, Xia Y, Chen J, Hao Q, Wang H and
blasts in the transition from inflammation to fibrosis following Sun D: Notch3 Ameliorates cardiac fibrosis after myocardial
myocardial infarction. Vascul Pharmacol 58: 182‑188, 2013. infarction by inhibiting the TGF‑β1/Smad3 pathway. Cardiovasc
25. Abdalla M, Goc A, Segar L and Somanath PR: Akt1 mediates Toxicol 16: 316‑324, 2016.
α‑smooth muscle actin expression and myofibroblast differentia- 38. Zhao M, Zheng S, Yang J, Wu Y, Ren Y, Kong X, Li W and
tion via myocardin and serum response factor. J Biol Chem 288: Xuan J: Suppression of TGF‑ β1/Smad signaling pathway by
33483‑33493, 2013. sesamin contributes to the attenuation of myocardial fibrosis in
26. Lan TH, Huang XQ and Tan HM: Vascular fibrosis in atheroscle- spontaneously hypertensive rats. PLoS One 10: e0121312, 2015.
rosis. Cardiovasc Pathol 22: 401‑407, 2013. 39. Khalil H, Kanisicak O, Prasad V, Correll RN, Fu X, Schips T,
27. Park S, Nguyen NB, Pezhouman A and Ardehali R: Cardiac Vagnozzi RJ, Liu R, Huynh T, Lee SJ, et al: Fibroblast‑specific
fibrosis: Potential therapeutic targets. Transl Res 209: 121‑137, TGF‑ β ‑Smad2/3 signaling underlies cardiac fibrosis. J Clin
2019. Invest 127: 3770‑3783, 2017.
28. Jugdutt BI: Remodeling of the myocardium and potential targets 40. Massagué J: TGF β signalling in context. Nat Rev Mol Cell
in the collagen degradation and synthesis pathways. Curr Drug Biol 13: 616‑630, 2012.
Targets Cardiovasc Haematol Disord 3: 1‑30, 2003. 41. Schmierer B and Hill CS: TGFbeta‑SMAD signal transduction:
29. Souders CA, Bowers SL and Baudino TA: Cardiac fibroblast: Molecular specificity and functional flexibility. Nat Rev Mol Cell
The renaissance cell. Circ Res 105: 1164‑1176, 2009. Biol 8: 970‑982, 2007.
30. Li P, Wang D, Lucas J, Oparil S, Xing D, Cao X, Novak L, 42. Lee SJ, Park K, Ha SD, Kim WJ and Moon SK: Gleditsia
Renfrow MB and Chen YF: Atrial natriuretic peptide inhibits sinensis thorn extract inhibits human colon cancer cells: The role
transforming growth factor beta‑induced Smad signaling and of ERK1/2, G2/M‑phase cell cycle arrest and p53 expression.
myofibroblast transformation in mouse cardiac fibroblasts. Circ Phytother Res 24: 1870‑1876, 2010.
Res 102: 185‑192, 2008. 43. Yin Y, Guan Y, Duan J, Wei G, Zhu Y, Quan W, Guo C, Zhou D,
31. Rizvi F, Siddiqui R, DeFranco A, Homar P, Emelyanova L, Wang Y, Xi M and Wen A: Cardioprotective effect of Danshensu
Holmuhamedov E, Ross G, Tajik AJ and Jahangir A: Simvastatin against myocardial ischemia/reperfusion injury and inhibits
reduces TGF‑β1‑induced SMAD2/3‑dependent human ventric- apoptosis of H9c2 cardiomyocytes via Akt and ERK1/2 phos-
ular fibroblasts differentiation: Role of protein phosphatase phorylation. Eur J Pharmacol 699: 219‑226, 2013.
activation. Int J Cardiol 270: 228‑236, 2018. 44. Zhang W and Liu HT: MAPK signal pathways in the regulation
32. Bujak M and Frangogiannis NG: The role of TGF‑beta signaling of cell proliferation in mammalian cells. Cell Res 12: 9‑18, 2002.
in myocardial infarction and cardiac remodeling. Cardiovasc
Res 74: 184‑195, 2007.
33. Li RK, Li G, Mickle DA, Weisel RD, Merante F, Luss H, This work is licensed under a Creative Commons
Rao V, Christakis GT and Williams WG: Overexpression of Attribution-NonCommercial-NoDerivatives 4.0
transforming growth factor‑beta1 and insulin‑like growth International (CC BY-NC-ND 4.0) License.
factor‑I in patients with idiopathic hypertrophic cardiomyopathy.
Circulation 96: 874‑881, 1997.You can also read

















































