MEC-12, an a -tubulin required for touch sensitivity in C. elegans
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Journal of Cell Science 112, 395-403 (1999) 395
Printed in Great Britain © The Company of Biologists Limited 1999
JCS7275
MEC-12, an α-tubulin required for touch sensitivity in C. elegans
Tetsunari Fukushige1,*, Zeba K. Siddiqui1,‡, Michael Chou2, Joseph G. Culotti3, Camelia B. Gogonea1,
Shahid S. Siddiqui1,§ and Michel Hamelin2,¶
1Toyohashi University of Technology, Laboratory of Molecular Biology, Tempaku-cho, Toyohashi, 441 Japan
2Merck Research Laboratories, PO Box 2000, Rahway, NJ 07065, USA
3Samuel Lunenfeld Research Institute of Mount Sinai Hospital, Toronto, Ontario, Canada M5G 1X5
*Present address: Department of Medical Biochemistry, University of Calgary, Calgary, Alberta, Canada T2N 4N1
‡Present address: Department of Biochemistry, Molecular Biology, and Cell Biology, Northwestern University, Evanston, IL 60208, USA
§Author for offprint requests
¶Author for correspondence (e-mail: michel_hamelin@merck.com)
Accepted 13 November 1998; published on WWW 13 January 1999
SUMMARY
mec-12 is one of a dozen genes required for touch receptor lacking the lysine-40 showed that acetylation is not
neuron function in Caenorhabditis elegans. Some mec-12 required for MEC-12 activity. Although functionally
mutants (mechanosensory-defective) lack the large- needed only in the touch neurons, mec-12 is expressed in
diameter microtubules that are characteristic of these several other neuron types. These results support the
neurons (15 protofilaments, as opposed to 11). Mutants of notion that tubulin isotype diversity contributes to the
mec-7, a β-tubulin encoding gene, have a similar phenotype. formation of distinct classes of microtubules; 15-
We have identified the nature of mec-12 by germline protofilament microtubule assembly requires MEC-12 α-
transformation rescue and characterization of a point tubulin and MEC-7 β-tubulin, which are both highly
mutation. Sequence analysis of the mec-12 encoded product expressed in the touch receptor neurons. MEC-12 is the
(MEC-12) indicates that it corresponds to a novel C. first reported α-tubulin isotype that appears to be required
elegans α-tubulin. MEC-12 is the only identified C. elegans in a single class of neuronal microtubules.
α-tubulin that contains a lysine at position 40, a known site
of post-translational acetylation. Some mec-12 mutations
eliminate microtubule acetylation as assayed immunocyto- Key words: Microtubule, Acetylation, Protofilament number,
chemically; phenotypic rescue using a MEC-12 variant Invertebrate
INTRODUCTION 1989). The touch-insensitive phenotype of these animals points
to an essential role for these microtubules in the
The response to light touch along the body of the nematode mechanotransduction process.
Caenorhabditis elegans is mediated by six touch receptor The gene mec-7 was shown to encode a novel β-tubulin,
neurons, ALML, ALMR, AVM, PVM, PLML, and PLMR MEC-7, one of the two main components (along with α-
(Chalfie and Sulston, 1981). Genetic studies on the touch tubulin) of microtubules (Savage et al., 1989). Moreover,
neurons have allowed the identification of numerous genes mec-7 was shown to be expressed almost exclusively in the
specifically affecting mechanosensation, and collectively touch neurons, in concordance with the mutant phenotype
labeled mec (mechanosensation defective) (reviewed by (Hamelin et al., 1992; Mitani et al., 1993). Since only the
Chalfie, 1993; Chalfie and Au, 1989). A unique morphological 15-protofilament microtubules of the touch neurons are
feature shared by the touch neurons is the presence of large- affected by mec-7 mutations, these data are consistent with
diameter microtubules along their axons. In most eukaryotic tubulins being the key determinants of microtubule diameter
cells, α- and β-tubulins co-assemble into 13-protofilament size.
microtubules (Tilney et al., 1973). In C. elegans, electron We now report that mec-12 corresponds to an α-tubulin
microscopy has revealed that the vast majority of microtubules (MEC-12), and that it is likely to be co-assembled with the
have 11 protofilaments; 13-protofilament microtubules are a mec-7 β-tubulin in the touch neuron 15-protofilament
subset of a few ciliated neurons (Chalfie and Thomson, 1982). microtubules. We also show that MEC-12 is the major (and
One remarkable feature of the touch receptor neurons, probably the only) acetylated α-tubulin in C. elegans. The C.
however, is the presence of 15-protofilament microtubules elegans MEC-7 and MEC-12 tubulins therefore represent a
along their axons. Mutations in two genes, mec-7 and mec-12, unique opportunity to examine parameters affecting the
specifically affect the formation of these large-diameter assembly, post-translational modification, and function of a
microtubules (Chalfie and Thomson, 1982; Chalfie and Au, structurally distinct class of microtubules.396 T. Fukushige and others
MATERIALS AND METHODS 5′ oligo: GTAATACATATCAGATATCGTAG;
3′ oligo: CATCGGATCCTCCGAGGGACTGGTCAGAAGGC.
Recombinant DNA techniques The mutation was confirmed by DNA sequencing.
Cosmid clones from the mec-12 region on chromosome III were To generate ‘mec-12 promoter-tba-1 coding region’ constructs, two
obtained, courtesy of A. Coulson and collaborators at the Sanger PCR fragments were ligated using an artificial XbaI site contributed
Centre (Cambridge, UK). Cosmid DNA was prepared (Coulson et al., by the PCR primers. These primers were, for mec-12:
1986) and submitted to Southern blot analysis with a 32P-labeled 5′: M13-20 primer (GTAAAACGACGGCCAGT);
probe (Feinberg and Vogelstein, 1983) made from the insert of 3′: CTTTCTAGATTGCAAAAGAGGAGCTACAAG.
cm5C5, a cDNA clone corresponding to the tba-1 tubulin gene For tba-1:
(Waterston et al., 1992; Fukushige et al., 1995). Washes were done in 5′:TTTTCTAGACGTTTTCAACATGCGTGAGG;
conditions of low stringency (0.2× SSC, 50°C). The same probe had 3′: CTTGGGCCCACTTGAAGTTGATGGTTG.
been used previously on a C. elegans genomic YAC grid (Coulson et Clone sequences were checked for integrity of the tba-1 coding
al., 1991) to determine a possible total of five α-tubulin genes (data region.
not shown; Fukushige et al., 1993; see also Results). Northern blot analysis was performed using a Nytran membrane
An EMBL3 genomic Lambda clone encompassing the entire mec- (Schleicher and Schuell), following instructions from the
12 gene was identified from a group of clones that hybridized manufacturer. 5 µg of poly(A)+-enriched RNA was used for each lane.
positively to a probe made of a C56F12 cosmid SalI fragment The probe used was the same HindIII restriction fragment mentioned
containing the early coding portion of the tba-1 gene (see Fig. 1; above. For reverse transcriptase-mediated polymerase chain reaction
Fukushige et al., 1995), under low-stringency conditions. (RT-PCR) experiments, the following oligos were used:
A mec-12 cDNA clone (approximately 1.8 kb long) was obtained 5′ oligo: GCCCACTTGTGTGTGTTCTGTTCC;
by screening a C. elegans size-selected library (P. Paress and D. Cully, 3′ oligo: CTAATACTCATCTCCTTCTTCTCC.
Merck Research Labs, unpublished) with a probe made from a These oligos delineate a region that spans from most of the 5′
C56F12 cosmid HindIII fragment encompassing the first three exons untranslated region to codon 432 of the mec-12 coding region.
of mec-12 (see Fig. 1). Starting from 100 ng C. elegans RNA, first strand cDNA was
A mec-12::lacZ reporter construct was constructed using a 4.5 kb synthesized (Superscript Preamplification System, Life Tech.). One
mec-12 SalI genomic restriction fragment that contains 2.9 kb DNA tenth of the resulting material was amplified with Pfu polymerase
upstream and 1.6 kb downstream of the start codon (from phage (Stratagene) under the following conditions: 2 minutes incubation at
SQ#TF1; see Fig. 1). This fragment, which includes the first 67 amino 94°C, then 35 cycles of: 45 seconds at 94°C, 1 minute at 60°C, 2
acids of mec-12, was fused in frame with the β-galactosidase gene of minutes at 72°C and, finally, an additional 5 minutes at 72°C. PCR
vector pPD22.04 (Fire et al., 1990). products were cloned into pCR2.1 vector (Invitrogen). PCR reactions
A 4.5 kb PstI-SalI restriction fragment covering the same region as for RFLP analyses were performed independently from those used to
described above was fused in frame at the corresponding SalI site in generate template for sequencing point mutations; RFLP analysis was
the mec-12 cDNA clone, to generate the pMM1 clone. PMM1 done by restriction digest on 10 µl (1/10 vol.) of RT-PCR product,
therefore contains a genomic fragment driving the expression of a followed by 1% TAE-agarose gel electrophoresis, and ethidium
(predominantly) cDNA coding region. This plasmid was subsequently bromide staining.
modified by PCR mutagenesis to generate a T to G nucleotide change Genomic RFLP blots were probed with a 1 kb XbaI fragment that
in codon 40. The resulting plasmid pMM4 therefore contains a lysine- contains the first exon and the flanking regions. For northern and
to-glutamine substitution (K40Q).The following oligos were used, in southern blot analyses, washes were done in high stringency
which the mutation introduced is underlined: conditions (0.2× SSC, 65°C).
Fig. 1. Identification of an α-tubulin-containing
Right Left clone in the mec-12 region. Solid bars above the
ben-1 mec-12 daf-2 dpy-1 gene structure represent relative cosmid clone
positions with regard to the YAC clone Y53G8.
mec-12 maps immediately to the right of daf-2,
1 m.u. on the left arm of chromosome III. Note that
transcription goes toward the left end of the
chromosome. The lower part of the Fig. is a
Y53G8 physical map of phage clone SQ#TF1 which
// // contains the mec-12 gene, and the localization
T21A6 R07G11 of target sites for the restriction enzymes
T10D5 HindIII and SalI. Exons are boxed, with coding
F57C5 sequence darkened; the putative proximal and
distal ends of the first and last exon respectively
C56F12 are based on sequence data from our mec-12
cDNA clone. Clone SQ#TF1 insert is 14 kb
long, and includes a 3 kb fragment downstream
of the mec-12 coding region which is not
present on cosmid C56F12. The intron positions
SQ#TF1 (with reference to Fig. 2) are located between
Sal I Hind III Sal I
Hind III
Sal I Hind III Sal I codons 1 and 2, within codon 76, between
Pst I Hind III
Start Stop codons 209 and 210, and between codons 354
and 355. Display of the relative positions of the
// cosmids is adapted from the ACeDB genome
1 kb database.The MEC-12 α-tubulin 397
Fig. 2. MEC-12 amino acid 1 MREVISIHIG QAGVQIGNAC WELYCLEHGI QPDGQMPSDK SLGGSDDSFS
sequence (as inferred from a mec- 40
12 cDNA clone). Residues that are 51 TFFSETGSGR HVPRAVMV L EPTVIDEIRT GTYRSLFHPE QLITGKEDAA
69 (N in u76 )
common to MEC-12 and the
highly similar Haemonchus 101 N N Y A R G H Y T I G K E I I D LT L D R I R R L A D N C T G L Q G F L V F H S F G G G T G S G F T
contortus α-tubulin (SwissProt 118
entry no. P50719), yet unique with 151 S L L M E R L S V D Y G K K A K L EFS IYPAPQVSTA VVEPYNSILT THTTLEHSDC
regard to other α-tubulins, are
shown in bold. Residues affected 201 SFMVDNEAIY DICRRNLDIE RPSYTNLNRL IGQIVSSITA SLRFDGALNV
by mec-12 mutations are outlined
(in addition to the substitution of 251 D L T E F Q T N L V P Y P R I H F P L A T F S P V I S A E K A Y H E Q L S V A E I T N M C F E PH N
299
an asparagine for an aspartate
(position 69; see Results), other 301 Q M V K C D P R H G K Y M A V C L L F R G D V V P K D V N A A I A T I K T K R S I Q F V D W C P TG
point mutations in this α-tubulin
351 FKV INYQPP TVVPGGDLAK VPRAVCMLSN TTAIAEAWAR LDHKFDLMYA
have now been revealed by Huang 354 (E inu241)
et al. (1995), using other mec-12 401 K R A F V H W Y V G E G M E G E F S E A R E D L A A L E K D Y E E V G V D S M E DN G E E G D E Y
mutants: they result in the 415 (K in u63 ) 443
predicted substitutions of a lysine
for a glutamate at position 415 in
mec-12[u63], and of a glutamate for a glycine at position 354 in mec-12[u241]). Lysine-40, a potential target for acetylation is also number-
tagged. The MEC-12 amino acid sequence and the cDNA sequence it was derived from are available from GenBank under accession number
U65429. Genomic sequence corresponding to the region spanning from exon 2 to exon 5 is available from the Japan Database (DDBJ) under
accession number D2-1134.
DNA sequence analyses as for the ability to rescue the mec-12 touch insensitive defect.
Manual DNA sequence was performed using a Sequenase kit One of these clones, C56F12, indeed displayed a clear α-
(U.S.B.); automated sequence was done with a PRISM kit (Perkin tubulin positive hybridization signal (data not shown). We then
Elmer). Inferred MEC-12 peptide sequence was compared to all other performed germline transformation of mec-12 mutants, to look
complete α-tubulin sequences available in the SwissProt Database for rescue of touch sensitivity. Phenotypic rescue of three mec-
(release no. 35) using the PrettyPlot alignment program available 12 mutant strains was obtained using DNAs from C56F12,
through GCG (Madison, WI). from the α-tubulin positive Lambda phage genomic clone
Generation and handling of transgenic nematodes SQ#TF1, and from the cDNA construct-containing plasmid
Cosmid, phage, and plasmid DNAs were injected along with the pRF4 pMM1 (Table 1); however, expression of another α-tubulin,
marker (conferring a rol-6 dominant phenotype; Fire, 1986; Mello et tba-1 (Fukushige et al., 1995), under the control of
al., 1991). For phenotypic rescue experiments, transgenic animals cosmid C56F12’s α-tubulin promoter did not lead to
were assayed for escape response to light touch with an eyelash gently mechanosensation rescue (Table 1; see Materials and Methods
drawn across the body. lacZ and DAPI staining were done as
described by Fukushige et al. (1993). Identification of touch receptor
neurons and other neurons was based on their relative position with Table 1. Transformation rescue of mec-12
regard to neighboring neurons as determined by DAPI staining, and Levels of α-
on their axonal patterns as visualized by lacZ staining (see White et tubulin
al., 1986; Wood, 1988). Strain Touch sensitivity* acetylation‡
Wild type (N2) + +
In situ immunofluorescence mec-12(e1605)§ − +
Antibody assays on whole-mounts were performed essentially as mec-12(e1607) − −
described previously (Hamelin et al., 1992). However, the reduction- mec-12(u63)§ − +
oxidation permeabilization step was found not to be necessary. mec-12(u76) − ±
Primary antibody dilution was from 1:2 to 1:10; secondary antibody Cosmid rescue attempt on mec-12(u76) + +
(rhodamine-conjugated goat anti-mouse, Jackson’s) dilution was from Cosmid rescue attempt on mec-12(e1605) + +
Phage rescue attempt on mec-12(u76) + +
1:50 to 1:200. Certain preparations were also done following the
Plasmid rescue attempt on mec-12(e1607) + +
method of Siddiqui et al. (1989). tba-1 rescue attempt on mec-12(e1607) − n.d.
Lys-40− - rescue attempt on mec-12(e1607) + −
mec-7(e1343) − +
RESULTS
See Results for details concerning the clones used for phenotypic rescue
Phenotypic rescue of mec-12 mutants attempts.
Our working hypothesis was that the mec-12 gene, based on *Mechanosensation: +, escape response to gentle touch with an eyelash; −, no
its phenotype, corresponds to an α-tubulin. A few cosmid response.
‡Acetylation levels estimated by indirect in situ immunofluorescence (see
clones have been mapped in the vicinity of mec-12, which is Materials and Methods): +, strong fluorescence signal; ±, little or no signal;
located on the left end of chromosome III, between the genes −, no signal.
daf- 2 and phm-3 (Fig. 1; Coulson et al., 1986; Hodgkin et al., §Mutants of mec-12(e1605) and mec-12(u63), although touch insensitive,
1995). DNA preparations from each of these cosmids were still display 15-protofilament microtubules (Chalfie and Au 1989; Chalfie
tested for the presence of an α-tubulin-encoding gene, as well and Thomson, 1982). n.d., not determined.398 T. Fukushige and others
and below for all clone descriptions). Finally, within the
overlap between C56F12 and SQ#TF1, the only open reading
frame seen is for the putative α-tubulin (Fig. 1); we therefore
conclude that mec-12 encodes a novel α-tubulin.
Structure of the mec-12 α-tubulin gene
mec-12 genomic and cDNA Lambda phage clones were
isolated by hybridization screens (see Materials and Methods).
Sequencing of these clones revealed the presence of 5 exons
and 4 introns (Fig. 1). mec-12 encodes a predicted α-tubulin
protein (MEC-12) of 450 residues (Fig. 2). MEC-12 shows a
high degree of conservation with other α-tubulins with
identities as high as 93% with vertebrate α-tubulins, 87% with
C. elegans tba-1 and tba-2 α-tubulins (Fukushige et al., 1993,
1995) and, interestingly, a 99% identity with an α-tubulin from
the parasitic nematode Haemonchus contortus (Klein et al.,
1992; SwissProt entry no. P50719). Finally, MEC-12 is the
only C. elegans α-tubulin that contains a lysine residue at
position 40, a potential target for acetylation (see below; note:
the latter statement also takes into account comparisons with
two additional, unpublished C. elegans α-tubulin potential
transcripts that were revealed by the Genome Sequencing
Center (Washington University, MO) under entries nos Z
78062 and U 40420).
When the mec-12 cDNA clone was used to probe a C.
elegans northern blot of RNA isolated from a mixed stage
population, a single messenger of approximately 2 kb was
detected (Fig. 3A).
Point mutation in a mec-12 mutant
We have sequenced a mec-12-specific RT-PCR product from
the mec-12(u76) mutant strain. Results from three
independent clones allowed the detection of a ‘G to A’ point
mutation in codon 69 (Fig. 3B), resulting in the predicted
substitution of an asparagine for an aspartate. This mutation
also predicted the loss of a SalI restriction site: this was
subsequently verified by restriction digest of a mec-12(u76)
RT-PCR product generated independently, under similar
conditions (Fig. 3C). In addition restriction fragment length
analysis of genomic DNA from mec-12(u76) also revealed the
loss of a SalI site in the same region (i.e. within the second
exon; Figs 1 and 3D).
mec-12 pattern of expression
A mec-12::lacZ reporter gene was constructed to examine the
pattern of expression of mec-12. Histochemical staining of
transgenic animals suggests strong levels of expression in the Fig. 3. Northern blot detection of mec-12 mRNA and identification
six touch receptor neurons, as expected from the mec-12 of a point mutation and of a restriction fragment length
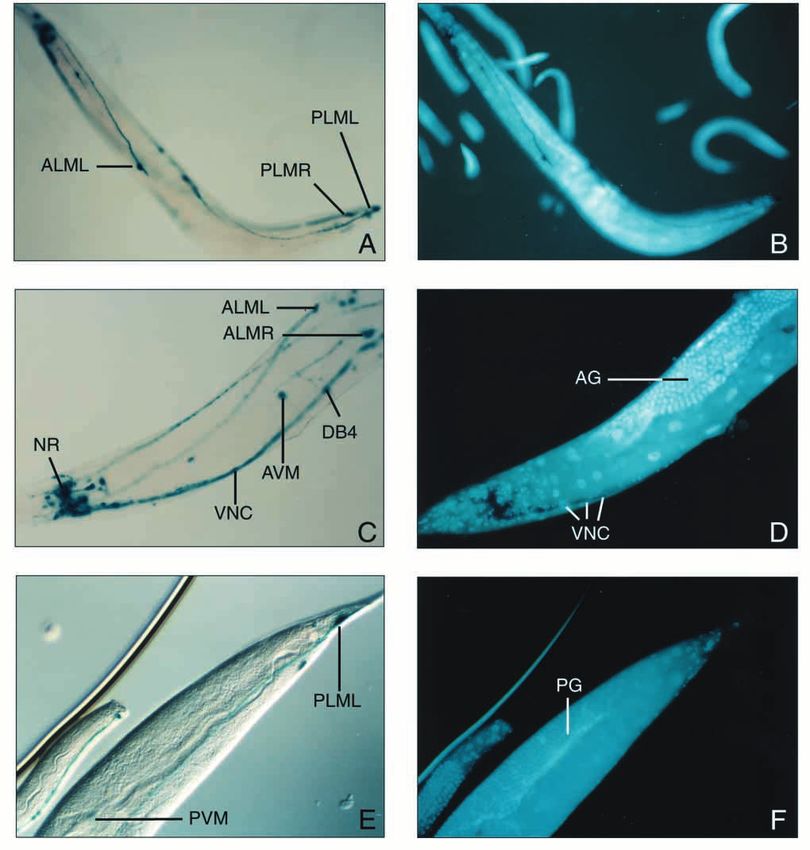
phenotype (Fig. 4A-F). In addition, staining was detectable in polymorphism. (A) Detection of mRNA band of approximately 2 kb
several other neurons, including motorneurons of the ventral by northern blot analysis; ribosomal marker sizes are indicated.
cord such as members of the DB class (Fig. 4C), various (B) Portion of a graphic diplay from automated sequencing of mec-
neurons of the nerve ring region (Fig. 4C,E), and a few neurons 12-specific RT-PCR products from wild-type (N2) and mec-12(u76);
of the lumbar ganglion such as PVR (not shown). No displayed above the electropherograms are the corresponding
expression was detected in ALN and FLP neurons; these are sequences, with codon 69 bracketed (C) Restriction fragment length
polymorphism (RFLP) analysis of mec-12-specific RT-PCR products
the cells, other than the touch neurons, that display mec-7 from wild-type (N2) and mec-12(u76), following a SalI digest; U:
expression (Hamelin et al., 1992) but do not contain 15- uncut PCR product from wild-type; arrow points to mec-12(u76)
protofilament microtubules. restriction band that resisted SalI cleavage, as predicted by sequence
analysis (see B; see also Materials and Methods for additional
MEC-12 acetylation and distribution details). (D) RFLP analysis of genomic DNA samples from wild-
Acetylation of microtubules is widespread among eukaryotes type (N2), mec-12(e1607), and mec-12(u76), following cleavage with
and is confined to a lysine residue at position 40 of the α- SalI, and probing with a DNA fragment spanning the first exon.The MEC-12 α-tubulin 399 Fig. 4. Expression pattern of a mec-12::lacZ reporter gene. (A,C,E) Histochemical (X-Gal) staining of mec-12::lacZ/rol-6 transgenic adult hermaphrodites; (B,D,F) DAPI nuclear staining visualization of the same animals (respectively). ALML, ALMR, AVM, PLML, PLMR, PVM: the six body touch receptors neurons; AG: anterior gonad arm; DB4: ventral cord motorneuron DB4; NR: nerve ring; PG: posterior gonad arm; VNC: ventral nerve cord. Magnifications: 1/100 (A,B); 1/200 (C,D,E,F). tubulin subunit (L’Hernault and Rosenbaum, 1985; LeDizet Fuller, 1985). In C. elegans, 6-11B-1 recognizes a single α- and Piperno, 1987; Edde et al., 1991). This lysine-40 is tubulin species identified by isoelectric focusing (Siddiqui et conserved in at least one α-tubulin gene in almost every al., 1989). 6-11B-1 also stains several neurons, including eukaryote investigated so far. A monoclonal antibody, 6-11B- motorneurons, but the most intensely stained cells are the touch 1, detects specifically acetylated α-tubulins in a variety of neurons (ALMR, ALML, AVM, PVM, PLMR, PLML) and a species ranging from Chlamydomonas to humans (Piperno and posterior interneuron, PVR (Siddiqui et al., 1989; Fig. 4A,B;
400 T. Fukushige and others
Fig. 5D). Therefore, the sequence, isoelectric focusing, and in
situ immunofluorescence data are consistent with MEC-12
being the only acetylated α-tubulin in C. elegans.
To determine whether the presence of an acetylated lysine
at position 40 is essential for the role of MEC-12 in
mechanosensation, we generated a plasmid construct that
contains the mec-12 promoter region fused with the coding and
the 3′-untranslated regions of a mec-12 cDNA clone but also
includes a K40Q substitution. This plasmid construct, with a
deleted acetylation site, was nevertheless able to rescue a
normal touch response in mec-12(e1607) mutants (Table 1).
DISCUSSION
We have identified a novel α-tubulin isotype that corresponds
to mec-12, a gene required for the normal function of the touch
receptor neurons in C. elegans. This finding is consistent with
the observation that several mec-12 mutants lack the large-
diameter microtubules that are found exclusively in the touch
neurons (Chalfie and Au, 1989). Similarly, the MEC-7 β-
tubulin is essential for the formation of those same
microtubules (Savage et al., 1989). 15-protofilament
microtubules in more complex organisms have been reported
only in a few instances such as in insect wings, epidermal cells,
or crustacean sperm (Burton et al., 1975; Nagano and Suzuki,
1975; Mogensen et al., 1989). It is not known if any specific
tubulin isotype is required in these cases. To our knowledge,
this is the first report of a given class of microtubule that
requires a specific isotype for both the α- and the β-tubulin
subunit. It is also the first reported α-tubulin mutation that
specifically affects neuronal microtubules.
The α-tubulin gene on cosmid C56F12 is mec-12
We have shown that DNA from a cosmid that maps to a region
of the C. elegans genome containing mec-12 can rescue the
touch-insensitive phenotype of two different mec-12 mutations.
Since this cosmid contains an α-tubulin gene, we hypothesized
Fig. 5. Whole mount in situ indirect immunofluorescence, using 6- that this gene is mec-12. This was confirmed by: (i) germline
11B-1, a monoclonal antibody specific to acetylated microtubules. rescue of mec-12 mutants using DNA from this cosmid, from
(A) anterior portion of a wild-type (N2) animal; (B) posterior end of an overlapping Lambda phage, and from a cDNA construct,
a wild-type animal; (C) anterior portion of a mec-12(u76) mutant; and (ii) our findings and the related findings of another group
(D) anterior portion of a rescued mec-12(u76) animal. VNC: ventral (Huang et al., 1995, with whom we shared unpublished data)
cord; NR: nerve ring; ALM: ALM neuron processes; PLM: PLM that the α-tubulin gene on C56F12 is mutated in at least three
neuron processes. Magnifications: 1/200. different independently-derived mec-12 alleles.
MEC-12 and MEC-7: two essential components of
PVR staining not shown). This 6-11B-1 antibody detection 15-protofilament microtubules
array is reminiscent of the pattern of expression of the mec-12 What is needed to generate 15-protofilament microtubules? In
reporter gene shown in Fig. 4. MEC-12 is likely the only α- principle any of several requirements can be evoked, including
tubulin with a lysine residue at position 40 (K40) in C. elegans, proper physiological conditions, the presence of appropriate
suggesting that it represents the single acetylated α-tubulin microtubule-associated proteins (MAPs), or the presence (or
identified by isoelectric focusing. We further examined this absence) of certain specific tubulin isotypes. In addition, the
possibility by carrying out in situ immunofluorescence analysis absolute levels of those specific tubulin isotypes may also be
of several mec-12 mutants. We found that, indeed, 6-11B-1 a crucial factor.
staining is greatly reduced in mec-12(u76) and completely In vitro polymerization experiments suggest that solution
absent in mec-12(e1607), a mutation also resulting in the lack conditions affect microtubule diameter size (Pierson et al.,
of 15-protofilament microtubules (Table 1; Fig. 5C; Chalfie and 1978; Scheele et al., 1982; Chretien et al., 1992). On the other
Au, 1989). Cosmid-, phage-, or plasmid-rescued mec-12(u76) hand, Aamodt and Culotti (1986) noted that for a given
and mec-12(e1607) transgenics (see above) were found to stain organism, the final diameter of microtubules assembled in vitro
with 6-11B-1 to the same level seen in the wild type (Table 1; from purified tubulins was predominantly the same as that seenThe MEC-12 α-tubulin 401
in vivo (i.e. 13 protofilaments for bovine and 11 protofilaments MEC-12 and MEC-7 conterparts. If this were true, the potential
for nematodes). role of any of the residues mentioned above in determining the
Some tubulin isotypes are known to be interchangeable, and number of protofilaments should be addressed. Nevertheless,
a clear demonstration that a tubulin isotype can affect the the aspartate residue at position 69 in MEC-12 appears to be
number of protofilament was reported recently by Raff et al. essential for the assembly of 15-protofilament microtubules, as
(1997), in which ectopic expression of a moth β-tubulin we have shown it to be the target of the mec-12(u76) mutation,
isoform into Drosophila testes resulted in the formation of which leads to the absence of 15-protofilament microtubules.
axonemal microtubules having 16 protofilaments, instead of Replacement by an asparagine (see Fig. 2) results in the
13. absence of these microtubules in the touch neurons (Chalfie
There are also clear examples in which ‘intra-isotypic’ ratios and Au, 1989). Likewise, substitution of a glutamate for a
(i.e. the ratio between α isotypes or between β isotypes) can glycine at position 354 results in the same phenotype (Huang
dictate what type of microtubule will be formed in a given cell. et al., 1995). Both of these residues are invariant in all α-
This is exemplified in Drosophila oocytes, where altering the tubulins heretofore sequenced, and may therefore be essential
α-tubulin ratio selectively affects microtubules required for for the assembly of microtubule of any diameter size.
cleavage stage mitoses (Matthews et al., 1993). This indicates Interestingly, the known point mutations in MEC-12 are
that specific tubulin isotypes, if present in sufficient quantities located in each of the three different domains defined by
relative to others, can dictate microtubule type and that some Nogales et al. (1998).
tubulin isotypes cannot functionally substitute for others.
In the touch receptor neurons, α- and β-tubulin isotypes Microtubule acetylation may not be essential in C.
other than MEC-7 and MEC-12 are presumably also present, elegans
since 11-protofilament microtubules substitute for 15- Post-translational modifications of α-tubulins include
protofilament in the touch neuron axons of both mec-7 and tyrosination (Barra et al., 1973), acetylation (L’Hernault and
mec-12 mutants (Chalfie and Thomson, 1982; Chalfie, 1993; Rosenbaum, 1985), glutamylation (Edde et al., 1990),
see also Fukushige et al., 1993, 1995). This argues against the polyglycylation (Redeker et al., 1994), and palmitoylation
involvement of specific MAPs that would dictate the formation (Zambito and Wolff, 1997). Tubulin acetylation generally
15-protofilament microtubules in these cells. correlates with incorporation of α/β dimers into a microtubule
The phenotypes of mec-7 and mec-12 support the notion that (L’Hernault and Rosenbaum, 1983) and with the subsequent
it is tubulin subunits that determine microtubule diameter. stability of the polymer (Piperno and Fuller, 1985; Gundersen
There are two ways in which expression of tubulins from these and Bulinski, 1986; Schulze et al., 1987; Lim et al., 1989; Baas
genes might lead to the formation of 15-protofilament and Black, 1990). There is evidence for only five α-tubulin
microtubules. One is that MEC-7 and MEC-12 tubulin proteins genes in C. elegans (see Results). All of them have now been
per se are specifically required for the assembly or stability of sequenced, and only MEC-12 has Lys-40, the sole residue that
these microtubules; the other is that MEC-7 and MEC-12 are becomes acetylated post-translationally (L’Hernault and
not specifically required (i.e. other isotypes can functionally Rosenbaum, 1985; LeDizet and Piperno, 1987; Edde et al.,
substitute for them), but their high level of expression is what 1991). Given the absence of the 6-11B-1 signal in mec-
dictates the formation of 15-protofilament microtubules in the 12(e1607) animals, we conclude that MEC-12 alone is
touch neurons. The latter model implies that other tubulin responsible for the acetylation of microtubules seen in a large
isotypes could form 15-protofilament microtubules if portion of the nervous system. α-tubulin acetylation has been
expressed at sufficiently high levels together in a given cell shown not to be essential in the lower eukaryote Tetrahymena
type. The failure of the TBA-1 α-tubulin (under mec-12 thermophila (Gaertig et al., 1995). Similarly, acetylation may
promoter-driven expression) to rescue touch sensitivity in a not be essential for growth of mammalian cells in vitro
mec-12 mutant (Table 1) argues against this ‘high tubulin level’ (Piperno et al., 1987). Since mec-12 mutants do not display
model (although the possibility that non-functional 15- behavioral defects other than touch insensitivity, it appears that
protofilament microtubules are made in these transgenics can microtubule acetylation is not essential in C. elegans.
not be entirely excluded). The simplest hypothesis is therefore Moreover, a mec-12 point mutant construct lacking the
that it is the sole presence of MEC-7 and MEC-12 together in acetylation site also rescued touch sensitivity, indicating that
the same cells that dictates the assembly of 15-protofilament acetylation was not essential for that function either.
microtubules. Finally, the gene expression and acetylation
patterns strongly suggest that MEC-12 takes part in the 15-protofilament microtubules and
assembly of 11-protofilament microtubles in cells other than mechanosensation
the touch neurons (which lack MEC-7). This further supports The exact role played by the 15-protofilament microtubules in
the notion that neither MEC-12 nor MEC-7 can determine the touch response remains unclear. It is possible that these
protofilament number alone. specialized microtubules are able, via an intermediate protein,
to induce a functional spatial arrangement of mechano-
Features of MEC-12 receptors and/or ion channels. Such a scheme has been
At the sequence level, MEC-12 and an α-tubulin from the proposed to take place in rat spinal cord synapses, with
parasitic nematode H. contortus (P50719) share 3 residues gephyrin as the intermediate link between microtubules and
found in no other α-tubulins (threonine-118, histidine-299, and glycine receptors (Prior et al., 1992; Kirsch et al., 1993). In the
asparagine-443; see Fig. 2). The extreme identity between C. elegans touch neurons, a putative ‘mechanosensation
these homologues raises the interesting question of whether H. receptor channel’ made of MEC-4 and MEC-10, two
contortus contains 15-protofilament microtubules comprised of transmembrane degenerins, has been proposed (Chalfie, 1993;402 T. Fukushige and others
Huang and Chalfie, 1994; Hong and Driscoll, 1994). Edde, B., Rossier, J., LeCaer, J.-P., Berwald-Netter, Y., Koulakoff, A.,
According to this model, dominant mutations in mec-4 and Gros, F. and Denoulet, P. (1991). A combination of postranslational
mec-10 would lead to a constitutively ‘open state’ of the modifications is responsible for the production of neuronal α-tubulin
heterogeneity. J. Cell. Neurochem. 46, 134-142.
channel, causing an osmotic imbalance and ultimately the Feinberg, A. P. and Vogelstein, B. (1983). A technique for radiolabelling
death of the touch neuron. Interestingly, mutations in mec-12 DNA restriction endonuclease fragments to high specific activity. Anal.
partially suppress dominant mutations of mec-10 (Huang and Biochem. 132, 6-13.
Chalfie, 1994), suggesting an involvement of 15-protofilament Fire, A. (1986). Integrative transformation of Caenorhabditis elegans. EMBO
J. 5, 2673-2680.
microtubules in the regulation of the channel. More recently, Fire, A., Harrison, S. W. and Dixon, D. (1990). A modular set of lacZ fusion
Huang and co-workers (1995) have found that two mec-12 vectors for studying gene expression in Caenorhabditis elegans. Gene 93,
mutations affect the axonal distribution of MEC-2, a stomatin- 189-198.
related gene product. In the same work it is shown that some Fire, A. (1992). Histochemical techniques for locating Escherichia coli β-
mec-2 mutations can suppress mec-4 and mec-10 mutations. galactosidase activity in transgenic organisms. GATA 9, 151-158.
Fukushige, T., Yasuda, H. and Siddiqui, S. S. (1993). Molecular cloning and
Evidence is therefore growing for the existence of a ‘MEC-7/ developmental expression of the Alpha-2 tubulin gene of Caenorhabditis
MEC-12:MEC-2:MEC-4/MEC-10’ complex of ‘microtubule: elegans. J. Mol. Biol. 234, 1290-1300.
linker protein: channel proteins’ involved in Fukushige, T., Yasuda, H. and Siddiqui, S. S. (1995). Selective expression
mechanosensation. The mec-12 mutants u63 and e1605 are of of the tba-1 α-tubulin gene in a set of mechanosensory and motor neurons
during the development of Caenorhabditis elegans. Biochim. Biophys. Acta
particular interest, since they affect the touch response and not 3, 1261 401-416.
the formation of microtubules. Gaertig, J., Cruz, M. A., Bowen, J., Pennock, D. G. and Gorovsky, M. A.
(1995). Acetylation of lysine 40 in α-tubulin is not essential in Tetrahymena
We thank the following for providing material essential to this thermophila. J. Cell Biol. 129, 1301-1310
work: A. Coulson (MRC, UK), for his help with clone localization Gundersen, G. G. and Bulinski, J. C. (1986). Microtubule arrays in
and for providing a C. elegans YAC grid; G. Piperno (Mt Sinai differentiated cells contain elevated levels of a postranslationally modified
Medical School, NY) for the 6-11B-1 antibody; M. Huang and M. form of tubulin. Eur. J. Cell Biol. 42, 288-294.
Chalfie (Columbia U., NY), for the mec-12(e1607) strain; P. Paress Hamelin, M., Scott, I. M., Way, J. C. and Culotti, J. G. (1992). The mec-7
β-tubulin gene of Caenorhabditis elegans is expressed primarily in the touch
and D. Cully (Merck Research Laboratories – MRL), for the cDNA receptor neurons. EMBO J. 11, 2885-2893.
library; A. Fire (Carnegie Inst., MD) for providing C. elegans vectors; Hodgkin, J., Durbin, R. and Martinelli, S. (1995). Genetic map of
J. Shockey and C. Morse (MRL), for preparation of figures, K. Korsun Caenorhabditis elegans. Caenorhabditis Genetics Center.
for technical assistance, and J. Schaeffer (MRL) for support. S.S.S. Hong, K. and Driscoll, M. (1994). A transmembrane domain of the putative
thanks Monbusho (Japan), the Human Genome Project; and NEC channel subunit MEC-4 influences mechanotransduction and
Corp. (Japan) for research grants. neurodegeneration in C. elegans. Nature 367, 470-473.
Huang, M. and Chalfie, M. (1994). Gene interactions affecting
mechanosensory transduction in Caenorhabditis elegans. Nature 367, 467-
470.
REFERENCES Huang, M., Gu, G., Ferguson, E. L. and Chalfie, M. (1995). A stomatin-
like protein necessary for mechanosensation in C. elegans. Nature 378, 292-
Aamodt, E. J. and Culotti, J. G. (1986). Microtubules and microtubule- 295
associated proteins from the nematode Caenorhabditis elegans: periodic Kirsch, J., Wolters, I., Triller, A. and Betz, H. (1993). Gephyrin antisense
cross-links connect microtubules in vitro. J. Cell Biol. 103, 23-31. oligonucleotides prevent glycine receptor clustering in spinal neurons.
Baas, P. W. and Black, M. M. (1990). Individual microtubules in the axon Nature 366, 745-748.
consist of domains that differ in both composition and stability. J. Cell Biol. Klein, R. D., Nulf, S. C., Alexander-Bowman, S. J., Mainone, C. B., and
111, 495-509. Geary, T. G. (1992). Cloning of a cDNA encoding α-tubulin from
Barra, H. S., Arce, C. A., Rodriguez, J. A. and Caputto, R. (1973). A Haemonchus contortus. Mol. Biochem. Parasitol. 56, 345-348.
soluble preparation from rat brain that incorporates into it’s own proteins L’Hernault, S. W. and Rosenbaum, J. L. (1983). Chlamydomonas α-tubulin
[14C]-arginine by a ribonuclease-sensitive system and [14C]-tyrosine by a is postranslationally modified in the flagella during flagellar assembly. J.
ribonuclease insensitive system. J. Neurochem. 20, 97-108. Cell Biol. 97, 258-253.
Burton, P. R., Hinkley, R. E. and Pierson, G. B. (1975). Tannic acid- L’Hernault, S. W. and Rosenbaum, J. L. (1985). Chlamydomonas α-tubulin
stained microtubules with 12, 13, and 15 protofilaments. J. Cell Biol. 65, is postranslationally modified by acetylation on the e-amino group of a
227-233. lysine. Biochemistry 24, 473-478.
Chalfie, M. and Sulston, J. (1981). Developmental genetics of the LeDizet, M. and Piperno, G. (1987). Identification of an acetylation site in
mechanosensory neurons of Caenorhabditis elegans. Dev. Biol. 82, 358- Chlamydomonas α-tubulin Proc. Nat. Acad. Sci. USA 84, 5720-5724.
370. Lim, S. S., Sammak, P. J. and Borisy, G. G. (1989). Progressive and spatially
Chalfie, M. and Thomson, J. N. (1982). Structural and functional diversity differentiated stability of microtubules in developing neuronal cells. J. Cell
in the neuronal microtubules of Caenorhabditis elegans. J. Cell Biol. 93, Biol. 109, 253-263.
15-23. Matthews, K. A., Rees, D. and Kaufman, T. C. (1993). A functionally
Chalfie, M. and Au, M. (1989). Genetic control of differentiation of the specialized α-tubulin is required for oocyte meiosis and cleavage mitoses in
Caenorhabditis elegans touch receptor neurons. Science 243, 1027-1033. Drosophila. Development 117, 977-991.
Chalfie, M. (1993). Touch receptor development and function in Mello, C. C., Kramer, J. M., Stinchcomb, D. and Ambros, V. (1991).
Caenorhabditis elegans. J. Neurobiol. 24, 1433-1441. Efficient gene transfer in C. elegans: extrachromosomal maintenance and
Chretien, D., Metoz, F., Verde, F., Karsenti, E. and Wade, R. H. (1992). integration of transforming sequences. EMBO J. 10, 3959-3970.
Lattice defects in microtubules: protofilament numbers vary within Mitani, S., Du, H., Hall, D. H., Driscoll, M. and Chalfie, M. (1993).
individual microtubules. J. Cell Biol. 117, 1031-1040. Combinatorial control of touch receptor neuron expression in
Coulson, A., Sulston, J., Brenner, S. and Karn, J. (1986). Toward a physical Caenorhabditis elegans. Development 119, 773-783.
map of the genome of the nematode Caenorhabditis elegans. Proc. Nat. Mogensen, M. M., Tucker, J. B. and Stebbings, H. (1989). Microtubule
Acad. Sci. USA 83, 7821-7825. polarities indicate that nucleation and capture of microtubules occurs at cell
Coulson, A., Kozono, Y., Lutterbach, B., Shownkeen, R., Sulston, J. and surfaces in Drosophila. J. Cell Biol. 108, 1445-1452.
Waterston, R. (1991). YACs and the C. elegans genome. BioEssays 13, 413- Nagano, T. and Suzuki, F. (1975). Microtubules with 15 subunits in
417. cockroach epidermal cells. J. Cell Biol. 64, 242-245.
Edde, B., Rossier, J., Le Caer, J.-P., Desbruyeres, E., Gros, F. and Denoulet, Nogales, E., Wolf, S. G. and Downing, K. H. (1998). Structure of the αβ
P. (1990). Postranslational glutamylation of α-tubulin. Science 247, 83-85. tubulin dimer by electron crystallography. Nature 391, 199-203The MEC-12 α-tubulin 403 Pierson, G. B., Burton, P. R. and Himes, R. H. (1978). Alterations in number structural fidelity of microtubules by initiation sites. J. Mol. Biol. 154, 485- of protofilaments in microtubules assembled in vitro. J. Cell Biol. 76, 223- 500. 228. Schulze, E., Asai, D. J., Bulinski, J. C. and Kirschner, M. (1987). Piperno, G. and Fuller, M. T. (1985). Monoclonal antibodies specific for an Postranslational modification and microtubule stability. J. Cell Biol. 105, acetylated form of α-tubulin recognize the antigen in cilia and flagella from 2167-2177. a variety of organisms. J. Cell Biol. 101, 2085-2094. Siddiqui, S. S., Aamodt, E., Rastinejad, F. and Culotti, J. (1989). Anti- Piperno, G., LeDizet, M. and Chang, X.-J. (1987). Microtubules containing tubulin monoclonal antibodies that bind to specific neurons in acetylated α-tubulin in mammalian cells in culture. J. Cell Biol. 104, 289-302. Caenorhabditis elegans. J. Neurosci. 9, 2963-2972. Prior, P., Schmitt, B., Grenningloh, G., Pribilla, I., Multhaup, G., Tilney, L. G., Bryan, J., Bush, D. J., Fujiwara, K., Mooseker, M. S. Beyreuther, K., Maulet, Y., Werner, P., Langosch, D., Kirsch, J. and Betz, Murphy, D. B. and Snyder, D. H. (1973). Microtubules: evidence for 13 H. (1992). Primary structure and alternative splice variants of gephyrin, a protofilaments. J. Cell Biol. 59, 267-275. putative glycine receptor-tubulin linker protein. Neuron 8, 1161-1170. Waterston, R., Martin, C., Craxton, M., Huynh, C., Coulson, A., Hillier, Raff, E. C., Fackenthal, J. D., Hutchens, J. A., Doyle, H. D. and Turner, L., Durbin, R., Green, P., Shownkeen, R., Halloran, N., Metzstein, M., F. R. (1997). Microtubule architecture specified by a β-tubulin isoform. Hawkins, T., Wilson, R., Berks, M., Du, Z., Thomas, K., Thierry-Mieg, Science 275, 70-73. J. and Sulston, J. (1992). A survey of expressed genes in Caenorhabditis Redeker, V., Levilliers, N., Schmitter, J.-M., Le Caer, J.-P., Rossier, J., elegans. Nature Genet. 1, 114-123. Adoutte, A., and Bre, M.-H. (1994). Polyglycylation of tubulin: a White, J. G., Southgate, E., Thomson, J. N. and Brenner, S. (1986). The postranslational modification in axonemal microtubules. Science 266, 1688- structure of the nervous system of the nematode Caenorhabditis elegans. 1691. Phil. Trans. R. Soc. Lond. B314, 1-340. Savage, C., Hamelin, M., Culotti, J. G., Coulson, A., Albertson, D. G. and Wood, W. B. (1988). The Nematode Caenorhabditis elegans. Cold Spring Chalfie, M. (1989). mec-7 is a β-tubulin gene required for the production Harbor Laboratory Press, Cold Spring Harbor, NY. of 15-protofilament microtubules. Genes Dev. 3, 870-881. Zambito, A. M. and Wolff, J. (1997). Palmytoylation of tubulin. Biochem. Scheele, R. B., Bergen, L. G. and Borisy, G. G. (1982). Control of the Biophys. Res. Commun. 239, 650-654.
You can also read

















































