A Biallelic Frameshift Mutation in Nephronectin Causes Bilateral Renal Agenesis in Humans
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
RAPID COMMUNICATIONS www.jasn.org
A Biallelic Frameshift Mutation in Nephronectin Causes Bilateral
Renal Agenesis in Humans
Lei Dai,1,2 Jingzhi Li,1,2 Liangqun Xie,1,2 Weinan Wang,1 Yang Lu,1,2 Mingkun Xie,1,2 Jingrui Huang,1
Kuifang Shen,1 Hui Yang,1 Chenlin Pei,1 Yanhua Zhao,1 and Weishe Zhang 1,2
1
Department of Obstetrics, Xiangya Hospital Central South University, Changsha, China
2
Hunan Engineering Research Center of Early Life Development and Disease Prevention, Changsha, China
ABSTRACT
Background Bilateral renal agenesis (BRA) is a lethal con genital anomaly caused by the failure of normal development of both
kidneys early in embryonic development. Oligohydramnios on fetal ultrasonography reveals BRA. Although the exact causes are
not clear, BRA is associated with mutations in many renal development genes. However, molecular diagnostics do not pick up
many clinical patients. Nephronectin (NPNT) may be a candidate protein for widening diagnosis. It is essential in kidney develop-
ment, and knockout of Npnt in mice frequently leads to kidney agenesis or hypoplasia.
Methods A consanguineous Han family experienced three cases of induced abortion in the second trimester of pregnancy, due
RAPID COMMUNICATIONS
to suspected BRA. Whole-exome sequencing (WES)–based homozygosity mapping detected underlying genetic factors, and a
knock-in mouse model confirmed the renal agenesis phenotype.
Results WES and evaluation of homozygous regions in II:3 and II:4 revealed a pathologic homozygous frameshift variant in
NPNT (NM_001184690:exon8:c.777dup/p.Lys260*), which leads to a premature stop in the next codon. The truncated NPNT
protein exhibited decreased expression, as confirmed in vivo by the overexpression of WT and mutated NPNT. A knock-in
mouse model homozygous for the detected Npnt mutation replicated the BRA phenotype.
Conclusions A biallelic loss-of-function NPNT mutation causing an autosomal recessive form of BRA in humans was confirmed
by the corresponding phenotype of knock-in mice. Our results identify a novel genetic cause of BRA, revealing a new target for
genetic diagnosis, prenatal diagnosis, and preimplantation diagnosis for families with BRA.
JASN 32: 1871–1879, 2021. doi: https://doi.org/10.1681/ASN.2020121762
Congenital anomalies of the kidney Approximately 56 genes (38 dominant For example, regulation of Glial cell
and urinary tract (CAKUTs) are a and 18 recessive) have been shown to line–derived neurotrophic factor
group of diseases with various pheno- be associated with CAKUT,5–12 but ex- (GDNF)/RET signaling is a key link in
types and clinical features caused by plain only 20%–25% of cases. the development of metanephros.13,14
embryonic defects of the kidney and As the most serious type of CAKUT, Disruption of GDNF/RET signaling–
urinary tract. CAKUT is a common bilateral renal agenesis (BRA) is a lethal related genes (such as GDNF,15 RET,16
birth defect, and the incidence rate in anomaly without both kidneys and ure- GFRA1,6 and ITGA817) causes a failure
neonates is approximately two in ters, and usually results in perinatal of ureteric bud (UB) formation from
1000,1 accounting for 30%–40%2 of all death or justifies pregnancy termina-
congenital malformations in children. tion. With the broad application of
Received December 19, 2020. Accepted April 3,
Most cases of CAKUT are sporadic, high-throughput sequencing tech- 2021.
but 15% are in familial clusters,3 which nology for clinical diagnosis in recent
indicates genetic factors are important years, mutations in many renal devel- Correspondence: Prof. Weishe Zhang, Department
of Obstetrics, Xiangya Hospital Central South
for the occurrence of CAKUT. To date, opment genes have been identified. University, 87 Xiangya Road, Changsha 410008,
the clinical manifestations of .500 syn- In general, different genes related to China. Email: zhangweishe@yeah.net
dromes have been found to include kidney development can lead to the oc- Copyright ß 2021 by the American Society of
renal and urinary tract deformities.4 currence of CAKUT in different ways.5 Nephrology
JASN 32: 1871–1879, 2021 ISSN : 1533-3450/1046-1871 1871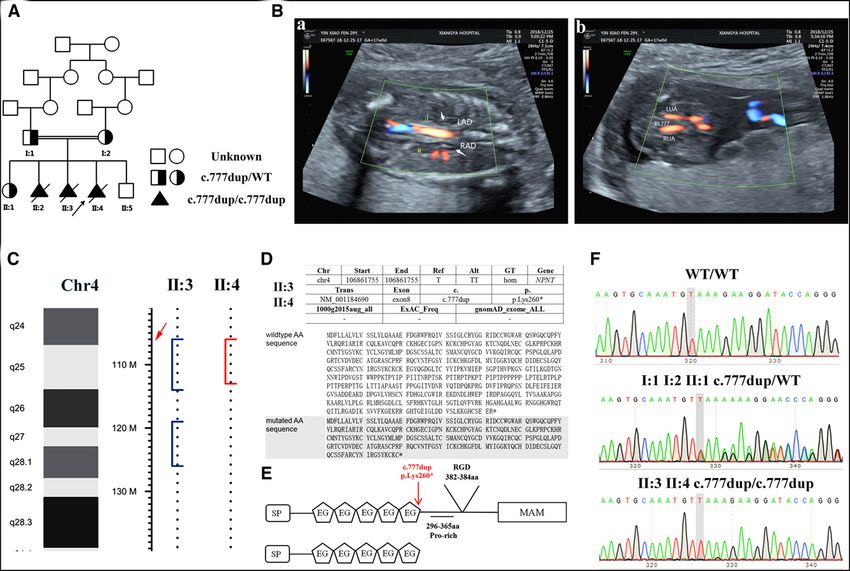
RAPID COMMUNICATIONS www.jasn.org
the Wolffian duct, or UB branching in For homozygosity mapping, the dis-
Significance Statement
the surrounding metanephric mesen- tribution of runs of homozygosity
chyme (MM), ultimately leading to the Bilateral renal agenesis (BRA) is a lethal (ROH) in the two affected fetuses (II:3,
occurrence of BRA.18–24 form of congenital anomaly of the kidney II:4) was searched using PLINK32 soft-
and urinary tract, usually resulting in peri-
Nephronectin (NPNT), also known natal death or justifying termination of
ware on the basis of the WES data
as preosteoblast EGF-like repeat pro- pregnancy. The exact causes of this dis- cording to established criteria.29 The
tein with meprin or EGFL6L, is an ex- ease are not clear, although genetic fac- variants were filtered under the
tracellular matrix component with a tors are considered important. In this acconditions of sequencing depth $30
study, we describe a pathological homo-
molecular weight of 70–90 kDa and a and allele frequency #0.1 in gnoma-
zygous frameshift variant
novel ligand for integrin a8b1 that was (NPNT:NM_001184690:exon8:c.777dup/ d_EAS_AF and 1000 g_EAS. For homo-
recently identified in mouse embryonic p.Lys260*) that is responsible for a reces- zygosity mapping, we identified ROH
kidneys.25,26 The NPNT gene is located sive form of BRA within a family. A regions according to the following pa-
on chromosome 4q24 and contains 13 knock-in mouse model mimicking the rameters: length of ROH $1 Mb, den-
mutation we found in the family was es-
exons. The encoded protein is com-
tablished and replicated the phenotype
sity of SNPs in ROH $1/1200 kb, and
posed of 561 amino acids, including of BRA. These results reveal a new ge- number of SNPs in ROH $4.
five EGF-like domains, one RGD motif netic cause of BRA and identify a new
(Arg-Gly-Asp), and one MAM domain target for genetic diagnosis, prenatal di- WES Data Analysis and
at the COOH terminus, and is consis- agnosis, and preimplantation diagnosis
for families with BRA.
Sanger Sequencing
tently highly expressed during kidney The genetic variants were functionally
development.25,27 Studies by Linton et annotated by ANNOVAR.33 The ho-
al. (2007)23 reported that Npnt2/2 platform (Illumina, San Diego, CA)
mozygous SNVs (nonsynonymous,
mice frequently display kidney agene- using the IDT XGen Exome Research
frameshift, and splice variants with allele
sis, due to a delay of invasion of the Panel (Coralville, IA) according to the
frequency ,1%) and Indels in the two
MM by the UB during the early stages manufacturers’ protocols. The genomic
affected fetal shared ROH regions were
of metanephric development, similar to DNA used was obtained from the amni-
selected for analysis.
that of Itga82/2 mice. Gdnf expression otic fluid of II:3 (residual DNA sample
The variant was confirmed by per-
is decreased in both Npnt2/2 and taken during the third pregnancy for
forming direct Sanger sequencing for
Itga82/2 mice, specifically in the MM chromosomal microarray analysis), the
all available family members for segre-
at the time of UB invasion.23 These fetal skin tissue of II:4 (taken after preg-
nancy termination), and the peripheral gation analysis. Primers (Human
findings suggested NPNT, which regu- NPNT-P, Supplemental Table 2) were
lates GDNF expression by binding blood cells of I:1, I:2, and II:1.
Read mapping and variant filtering used to amplify the PCR fragments
integrin a8b1, is essential for renal de- covering NPNT exon 8, and the prod-
velopment.23,28 Therefore, NPNT is were performed according to previously
described criteria.29 In brief, filtered ucts were subsequently sequenced us-
thought to be a possible pathogenic gene ing an ABI-A3500 genetic analyzer.
raw FASTQ reads were aligned to the
for BRA. Nevertheless, NPNT mutations The raw WES data were submitted
human reference genome (build hg19)
have not yet been described in patients to the National Center for Biotechnolo-
with a Burrows-Wheeler Alignment
with renal agenesis. gy Information Sequence Read Archive
Tool.30 The Picard software tool
In this study, we present one consan- database under the accession codes
(http://broadinstitute.github.io/picard/)
guineous Han family with recurrent fe- SRR13239865 (I:1), SRR13239864 (I:2),
was then used to remove duplicate
tal loss due to renal agenesis. A NPNT SRR13239863 (II:1), SRR13239862
reads. We obtained 18.51 GB and 13.28
recessive loss-of-function mutation was (II:3), and SRR13239866 (II:4). The re-
GB of data for II:3 and II:4, respective-
identified and confirmed in knock-in viewer link before release is https://
ly. The average coverage rate of the tar-
mice. Our results revealed NPNT is a dataview.ncbi.nlm.nih.gov/object/PRJN
get sequence was 99.8%, and the mean
novel genetic cause of BRA in humans. A684351?reviewer=tm5m hdk1ja06bmf
read depth was 153.8. The 20x mean
depth coverage rate for the samples was sg2lurgnm34.
96.37%. In total, 101.23 M mapped
METHODS
reads with a duplication rate of 19.01% Lentiviral Vectors Expressing Wild-
were obtained. The detailed quality type NPNT or K260ter-NPNT
Whole-exome Sequencing–based control data for all samples are listed in To further investigate the effect of the
Homozygosity Mapping Supplemental Table 1. For single nucle- mutation, we constructed wild type
Whole-exome sequencing (WES) was otide polymorphisms (SNP) calling, we (WT)–NPNT (Sp-3flag-WT-NPNT) and
performed for five members of one fam- used the Genome Analysis Toolkit31 for K260ter-NPNT (Sp-3flag-Lys260*-NPNT)
ily with BRA (I:1, I:2, II:1, II:3, II:4; Fig- single-nucleotide variant (SNV) and in- plasmids (Figure 2A). Full-length NPNT
ure 1A) on the Illumina NovaSeq6000 sertion-deletion (Indel) detection. cDNA with a 33 FLAG sequence
1872 JASN JASN 32: 1871–1879, 2021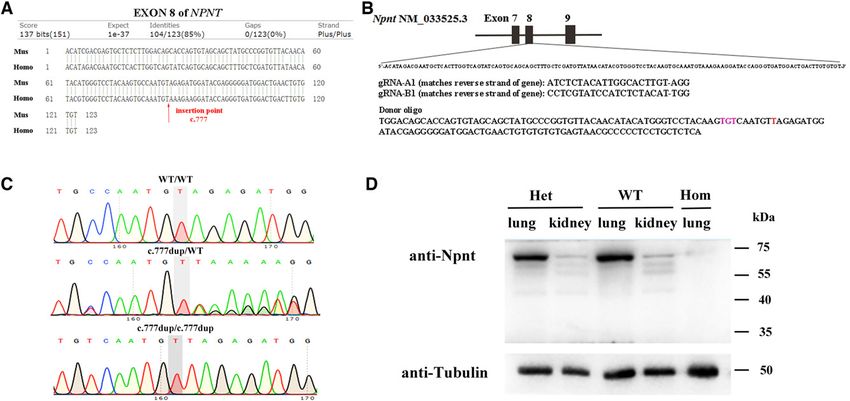
www.jasn.org RAPID COMMUNICATIONS Figure 1. Mutated NPNT in a consanguineous Han family with recurrent loss of fetuses with oligohydramnios due to bilateral renal agenesis. (A) Pedigree with the NPNT mutation. (B) Ultrasound images of affected fetus II:4. (a) Examination of prenatal ultrasound and color Doppler showed “lying-down” adrenal glands, and the kidneys could not be visualized. Abdominal aorta blood flow was detected, but bilateral renal artery blood flow was not. The arrows show the adrenal glands. (b) The urinary bladder could not be vi- sualized between umbilical arteries. (C) WES-based homozygosity mapping analysis revealed one 6.317 Mb homozygous genomic region (4q24-q25) shared by the two affected fetuses (II:3 and II:4) in the family. The arrow shows the position of the human NPNT gene. (D) Information and functional predictions for the NPNT mutation. (a) The variant is not recorded in the Freq_ExAC_ALL, Freq_1000g2015aug_all and gnomAD databases. (b) Mutated AA sequence predicted by MutationTaster. The mutation introduces a premature stop codon 1 position downstream. http://www.mutationtaster.org/. (E) WT (upper) and deduced mutant (lower) NPNT proteins; the Pro-rich, RGD, and MAM domains are absent in the mutated protein. (F) Chromatograms of NPNT mutation genotyp- ing by Sanger sequencing. LAD, left adrenal gland; RAD, right adrenal gland; L, left kidney region; R, right kidney region; LUA, left umbilical artery; RUA, right umbilical artery. after the signal peptide was chemically WT-NPNT and K260ter-NPNT plasmids produce recombinant lentivirus, 293T cells synthesized by using NM_001184690 as were identified by using vector and insert were cotransfected with WT-NPNT a template. A site-directed mutagenesis cDNA sequence primers (WT-NPNT-P or K260ter-NPNT and packaging plasmids method34 was used to introduce the mu- and pEGFP-N-3 for WT-NPNT plasmid, (pHelper 1.0 and 2.0, Shanghai GeneChem tation into K260ter-NPNT using the K260ter-NPNT-P, and pEGFP-N-3 for Co., China). The fluorescence method was WT-NPNT cDNA as a template, with K260ter-NPNT plasmid, Supplemental used to determine the virus titer. the primers listed in Supplemental Table Table 2). The cDNA sequence inserted 2 (NPNT-P1, NPNT-P2, NPNT-P3, and into the plasmids was confirmed by PCR- NPNT-P4). The cDNA sequence was based sequencing using a specific primer Cell Culture and Transfection cloned into the lentiviral vector GV367 set (Ubi-F-P, WT-NPNT-P, Plasmid- with Lentivirus (Shanghai GeneChem Co., China) for NPNT-P, and pEGFP-N-3 for HEK293T cells (GDC187) were pur- cocistronic expression of GFP by the re- WT-NPNT plasmid; Ubi-F-P, K260ter- chased from the China Center for Type striction enzyme–mediated integration NPNT-P, Plasmid-NPNT-P, and pEGFP- Culture Collection (Wuhan, China). The method. Transformants positive for the N-3 for K260ter-NPNT plasmid). To cells were infected with recombinant JASN 32: 1871–1879, 2021 NPNT in Human Bilateral Renal Agenesis 1873
RAPID COMMUNICATIONS www.jasn.org
lentivirus with enhanced infection The experimental protocols were ap- bands were probed with different prima-
solution (GeneChem, China) and cul- proved by the Animal Ethics Commit- ry antibodies. Primary antibodies against
tured in DMEM containing 10% FBS at tee of Xiangya Hospital (Changsha, NPNT/Npnt (mouse anti-human/mouse,
5% CO2 and at 37 C. Transfection effi- China). 1:1000, sc-393033, SCBT), FLAG (mouse
ciency was monitored by cotransfection anti-flag, 1:1500, 8146, CST), and
of an expression plasmid for GFP and Histology Assay b-tubulin (rabbit anti-b-tubulin, 1:1500,
was consistently .90%. Tissue from II:4, a fetus with BRA from AC008, ABclonal) were used. The
the affected family, was fixed in forma- membranes were incubated with corre-
Generation of Npnt-c.777dup/ lin during autopsy at 3 months after in- sponding Horseradish Peroxidase-labeled
p.Lys260* Mice duced labor. Mouse tissues (brain, secondary antibodies (rabbit anti-mouse
Although Npnt-knockout mice are re- heart, liver, spleen, lung, pancreas, or mouse anti-rabbit IgG, 1:20000; AB-
ported to display renal agenesis at stomach, large intestine, small intestine, clonal), and finally visualized by West-
birth, it is unclear whether the frame- and eye) were fixed in 4% paraformal- ernBright ECL (Advansta). Beta-tubulin
shift mutation we found in the family dehyde at 0 dpp after dissection. Paraf- was used as the reference.
causes the same phenotype, because the fin-embedded blocks of tissues were
protein is produced but truncated. The prepared, and sections of 5 mm thick- Statistical Analysis
CRISPR/Cas9 system can precisely in- ness were cut and stained with hema- All data are presented as the
troduce stable mutations at specific toxylin-eosin. mean6SD. Data were analyzed using
sites with a cleaner genetic background, the SPSS software package, version 25
making it suitable for studying the ef- RNA Extraction and Real-time PCR (IBM Software). The independent t test
fect of the gene mutation on disease. Total RNA was extracted by TRIzol was applied to compare differences be-
Therefore, to investigate the pheno- (Invitrogen, Carlsbad, CA) from tween groups. P,0.01 was considered
type and effect of the mutation, we HEK293T cells at 3 days after transfec- statistically significant.
constructed knock-in mice with the tion. RNA was reverse transcribed into
Npnt-c.777dup/p.Lys260* mutation cDNA using the PrimeScript RT Re- Human Clinical Data
(below described NpntK260*) in C57BL/ agent Kit with gDNA Eraser (TaKaRa, A consanguineous Han family residing
6 mice via the CRISPR/Cas9 system Dalian, China). PCR was performed in Jiangxi Province, China, with recur-
(Cyagen, China). The Npnt gene (En- with a 7500 Real Time PCR System rent loss of fetuses with oligohydram-
sembl: ENSMUSG00000040998; GEN- (Applied Biosystems) using SYBR nios due to BRA participated in this
BANK_ACCESSION: NM_033525.3) is Green Real-time PCR Master Mix study (Figure 1A). The maternal grand-
located on mouse chromosome 3. The (Toyobo, Osaka, Japan) with primers mothers of the couple were sisters. The
c.777dup mutation was introduced in against NPNT and b-actin (Human couple was referred during their fourth
exon 8 of the donor oligonucleotide. NPNT-QPCR, Human Actb-QPCR; pregnancy. A healthy girl was born in
Microinjection of gRNA, Cas9 mRNA, Supplemental Table 2). Expression of the first pregnancy (II:1). However, the
and the donor oligonucleotide into zy- NPNT mRNA was normalized to that second (II:2) and third (II:3) pregnan-
gotes was carried out, and then the zy- of b-actin. cies were terminated at 21–22 weeks of
gotes were transferred into the uteri of gestational age in another hospital after
pseudopregnant C57BL/6 females (Fig- Western Blotting ultrasound testing indicated oligohy-
ure 3, A and B). Direct sequencing of Protein lysates were prepared from dramnios (August 2016 in Dongguan
PCR-amplified fragments from the tail HEK293T cells at 3 days after transfec- Maria Maternity Hospital, May 2017 in
DNA was performed for genotyping of tion and lung and kidney tissues from Maternal and Child Health Hospital of
F0 mice (Mouse-NPNT-P, Supplemental mice at 0 dpp. Because the kidney tis- Ji’an County) (Supplemental Figure 1).
Table 2). After intercrossing F0-tar- sues of homozygous mice were not Chromosomal microarray of the third
geted mice and WT C57BL/6 mice, we enough to extract the protein, the lung pregnancy (II:3) revealed no obvious
obtained F1 heterozygous targeted tissues were chosen to detect the ex- chromosome abnormalities. The fourth
mice. F2 homozygous targeted mice pression of Npnt proteins. The Brad- pregnancy (II:4) was conceived sponta-
were generated by intercrossing hetero- ford method was used to determine the neously, and the first diagnosis of oligo-
zygous mice from the F1 generation, total protein content. Protein separa- hydramnios was made by ultrasound at
and the genotypes were confirmed by tion was performed by SDS-PAGE, and the Maternal and Child Health Hospi-
direct sequencing (Figure 3C). Expres- the separated proteins were transferred tal of Ji’an City at the 16th week of ges-
sion of Npnt was analyzed by Western to polyvinylidene difluoride mem- tation (December 21, 2018). Level two
blotting (WB) in the lung and kidney branes. The membrane was incubated ultrasonography scan at our hospital 4
tissues from all three groups of F2 mice with blocking solution for 1 hour at days later showed a “lying-down” adre-
at 0 days postpartum (dpp) (Figure 3D). room temperature, and the protein nal gland and absent kidneys
1874 JASN JASN 32: 1871–1879, 2021www.jasn.org RAPID COMMUNICATIONS
bilaterally, consistent with the suspi- homozygosity mapping, we identified genomic region (4q24-q25) shared by
cion of BRA (Figure 1B). Fetal growth 191 and 181 ROH regions ($1 Mb) in the two affected fetuses (II:3, II:4) was
was normal (Supplemental Table 3). II:3 and II:4, respectively. A total of identified (Figure 1C) as the first candi-
Anatomic and histopathological exami- 35.05 Mb genomic intervals (17 ROH date to test as causative for BRA.
nation of the terminated pregnancy $1 Mb) were shared by the two fetuses This variant is predicted by Muta-
confirmed the absence of kidneys with- (Supplemental Table 4) and we found tionTaster36 to result in a truncated pro-
out any other obvious structural 310 variants in those shared ROH re- tein that was generated by a premature
anomalies (Supplemental Figure 2), but gions. After screening and filtering ac- stop codon of TAA in the next codon
functional abnormalities or minor struc- cording to the parameters described in (260aa). The RGD motif (at positions
tural abnormalities were not excluded. the Methods, only two variants 382–384) and MAM domain (at positions
remained. After removing benign or 420–563) located in the region between
likely benign variants predicted by In- 260 aa and the C-terminus are absent in
WES-based Homozygous terVar,35 only one variant remained the truncated protein. The RGD motif is
Mapping Analysis (Supplemental Table 5). Therefore, a already known as the primary integrin
WES was carried out for five individu- homozygous nucleotide variant, recognition site in extracellular matrix
als in consanguineous family (two af- c.777dup in exon 8 of the NPNT gene proteins, and the MAM domain is essen-
fected fetuses, II:3 and II:4; their sister, (NM_001184690:exon8:c.777dup/ tial for localization to the cell surface,27
II:1; and the parents, I:1 and I:2). By p.Lys260*) in a 6.317Mb homozygous which is important for the function of
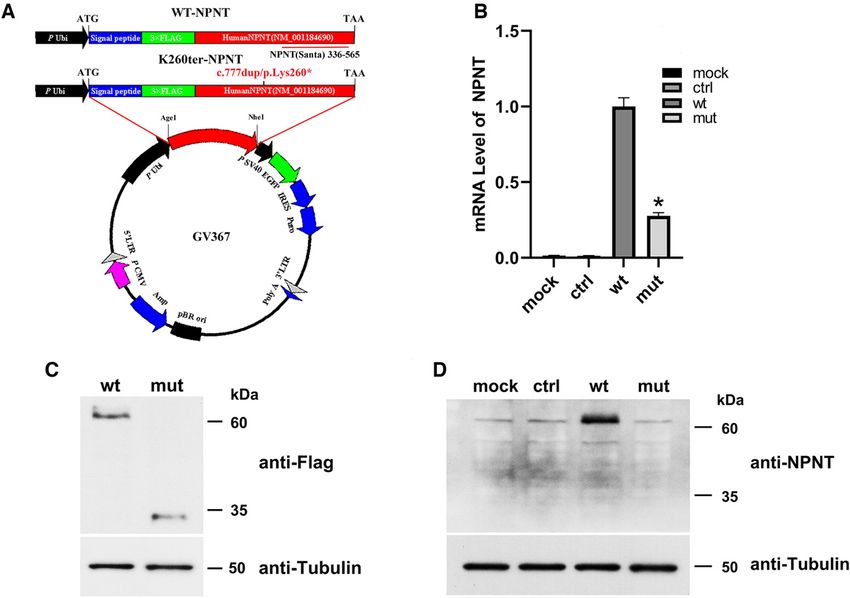
Figure 2. In vitro overexpression of WT-NPNT and K260ter-NPNT in cultured 293T cells. (A) Construction of the NPNT lentiviral vec-
tors. The human NPNT cDNA (WT-NPNT and K260ter-NPNT) sequences were amplified and ligated to the Ubi-MCS-SV40-EGFP-
IRES-puromycin plasmid at the AgeI/NheI site. After producing recombinant lentivirus, the plasmids were transfected into 293T cells.
Transfection efficiency was monitored by cotransfection of a GFP expression plasmid. After the cells were cultured for 72 hours, they
were harvested and subjected to quantitative RT-PCR and WB. The red bar indicates the position of the NPNT antibody immuno-
genic sequence. (B) NPNT mRNA levels were significantly lower in K260ter-NPNT–transfected cells than in WT-NPNT–transfected
cells, according to quantitative real-time PCR (*P,0.01). The NPNT expression level was normalized to that of endogenous b-actin.
(C) WB performed using a FLAG antibody showed a truncated protein of 35 kDa with decreased expression in K260ter-NPNT–trans-
fected cells. Tubulin was used as the control. (D) WB performed with an NPNT antibody showed obviously increased expression of
NPNT after WT-NPNT transfection, but only endogenous NPNT expression was detected only in mock, control, and in K260ter-
NPNT–transfected cells.
JASN 32: 1871–1879, 2021 NPNT in Human Bilateral Renal Agenesis 1875RAPID COMMUNICATIONS www.jasn.org
Figure 3. Generation of knock-in mice mimicking the mutation of the homozygous frameshift variant (NPNT:NM_001184690:ex-
on8:c.777dup/p.Lys260*) in the family by CRISPR-Cas9 technology. (A) Sequence homology comparison of NPNT exon 8 between
humans and mice. The arrow shows the insertion point corresponding to the mutation. (B) Schematic diagram of the exon 8 target
site in knock-in mice. The localization and sequences of gRNAs and donor DNA are indicated. (C) Chromatograms for Npnt mutation
genotyping of F2 offspring by Sanger sequencing from mouse tail genomic DNA. (D) Detection of Npnt proteins by WB in mice at 0
dpp. The expected sized bands of Npnt proteins are detected in the lung and kidney tissues from WT and heterozygous mice, but
no bands were detected in lung tissue from homozygous mice.
NPNT. The mutation was not recorded in endogenous expression in the control no external defects) but died the day af-
any SNP databases (Figure 1, D and E). and mock cells. As expected, the truncat- ter birth. The ratio of homozygous
Sanger direct sequencing verified the vari- ed protein was undetected by this anti- (21.7%, 15 of 69), heterozygous (53.6%,
ant in the affected fetuses (II:3, II:4, homo- body in K260ter-NPNT–transfected cells, 37 of 69), and WT (24.6%, 17 of 69)
zygous), the sister (II:1, heterozygous), except endogenous NPNT (Figure 2D). mice was in line with the expected
and the parents (I:1, I:2, heterozygous) These results demonstrated the Mendelian ratio, and there was no ob-
(Figure 1F). p.Lys260* mutation results in both the vious sex bias (Supplemental Tables
degradation of NPNT mRNA by non- 6–8). Through dissection at 0 dpp, we
Expression of WT-NPNT and sense-mediated mRNA decay37 and the found the vast majority of homozygous
K260ter-NPNT in HEK293T cells production of a truncated protein. mice had no kidneys on either side (14
In vitro overexpression of WT-NPNT of 15), accompanied by absence of the
and K260ter-NPNT in HEK293T cells Npnt K260*/K260* Mice Display Renal ureter and normal development of the
showed significantly decreased Agenesis at Birth bladder adrenal glands and internal
K260ter-NPNT mRNA levels com- Knock-in mice with the Npnt-c.777dup/ genitalia (Figure 4A); the remaining
pared with those of WT-NPNT after p.Lys260* mutations were obtained by the NpntK260*/K260* mouse had two primor-
transfection (Figure 2B). WB analysis CRISPR/Cas9 system (Figure 3, A and B). dial kidneys that were smaller in size
using anti-Flag antibody detected a Homozygous mutants (NpntK260*/K260*) (Figure 4B). There are some differences
truncated protein of 35 kDa, with were generated by mating the heterozy- in the phenotypic severity of renal
reduced expression in K260ter- gous (NpntK260*/WT) mice. No NPNT pro- agenesis compared with Npnt2/2 mice.
NPNT–transfected cells (Figure 2C). tein was detected by WB analysis using The anatomy of the homozygous mice
To further confirm the domains absent Npnt antibody in lung tissue from homo- at birth showed a high frequency
in the truncated protein, we used the zygous mice (Figure 3D), demonstrating (93.3%) of bilateral kidney absence,
NPNT antibody that binds the epitope the mutation introduced in a mouse mod- compared with the frequency of 46%
(336–565aa) downstream of the muta- el affected the expression of Npnt protein, previously reported in Npnt2/2 mice.23
tion (Figure 2A). The higher expression similar to humans. One potential explanation for this ob-
of NPNT was detected after WT-NPNT All mice were born alive with a nor- servation is that another ligand (or oth-
transfection compared with the mal appearance (size, body weight, and er ligands) expressed by the UB may
1876 JASN JASN 32: 1871–1879, 2021www.jasn.org RAPID COMMUNICATIONS
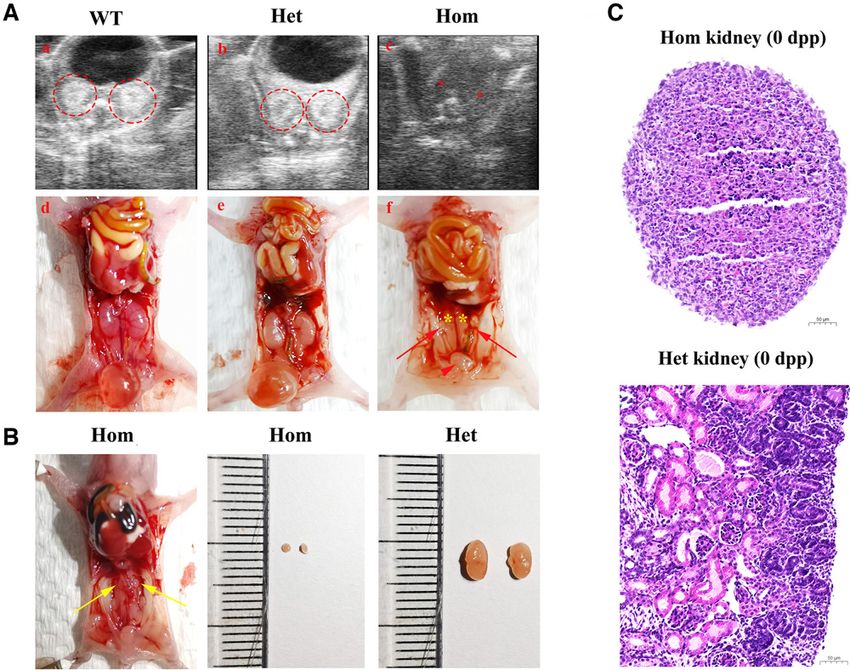
Figure 4. Kidney phenotypes of NpntK260*/K260* mice at 0 dpp. (A) (a), (b), and (c) Small animal ultrasound before dissection showed ab-
sent kidneys bilaterally in homozygous mutants. The red dotted circles show the kidney signals in WT and heterozygous mice. The red
asterisks show the missing kidney signals. (d), (e), and (f) Morphology and anatomy changes in mice. In WT and heterozygous mice, the
adrenal glands, kidneys, ureter, and bladder were well developed. Most of the homozygous mutants (14 out of 15) had no ureter or kid-
neys, but the bladder, adrenal glands, and internal genitalia were unaffected. The yellow asterisks show the absence of kidneys, the red ar-
rowhead shows the bladder, and the red arrows show the uterus and fallopian tubes. (B) One homozygous mutant showed renal dysplasia,
and the size of the kidneys (yellow arrows) was significantly smaller than that in heterozygous mice. (C) Histologic analysis of the kidney of
homozygous and heterozygous mice at 0 dpp. There were few nephrons in the homozygous primordial kidneys. Scale bar, 50 mm.
functionally compensate for the absent without obvious morphologic anomalies BRA in humans, and disclose a new
Npnt protein, mediating the respon- in other tissues (Supplemental Figure 3). target for genetic diagnosis, prenatal
siveness of the MM in Npnt2/2 mice. Consistent with this finding, Linton et al. diagnosis, and preimplantation diag-
In our mutated mice, the truncated (2007) reported that Npnt2/2 mice that nosis in BRA families.
Npnt protein residues may have nega- survive after birth are healthy and fertile
tive effects on the alternative ligand. with an apparently normal lifespan.23 The DISCLOSURES
Another possible explanation is the dif- authors hypothesized that EGFL6 (also
ferent genetic backgrounds. Linton et named MAEG),38,39 as an alternative can- All authors have nothing to disclose.
al. (2007) stated the frequency of agen- didate, compensated for the absence of
esis was dramatically decreased in NPNT in those tissues. In contrast, no
FUNDING
Itga82/2 mice in an outbred back- agenesis was observed in heterozygous or
ground.23 Therefore, our NpntK260*/ WT offspring (Figure 4, A and B). This study was supported by The Science and
K260*
mice inbred with a pure C57BL/6 In conclusion, we describe that ho- Technology Innovation Program of Hunan Pro-
background may exhibit less trait vari- mozygous loss-of-function NPNT vince grants 2019SK1010 and 2020SK2072 (to
ability. It is also possible the homozy- mutations in the affected family of W. Zhang).
gote we obtained was not sufficient to this study are responsible for the re-
ACKNOWLEDGMENTS
express the full variability in the trait of cessive form of BRA. The mouse
kidney development. model mimicking the mutation we
The authors gratefully acknowledge the con-
Histologic analysis revealed develop- found in the human family confirmed tributions of the family to this study. We also
mental arrest in the nephron in the homo- the clinical phenotype of BRA. Our thank Mr. Quanfei Huang, Mrs. Lisi Liang, and
zygous primordial kidneys (Figure 4C), results reveal a new genetic cause of Miss. Yang Wang from CapitalBio Genomics
JASN 32: 1871–1879, 2021 NPNT in Human Bilateral Renal Agenesis 1877RAPID COMMUNICATIONS www.jasn.org
Co., Ltd. for next-generation sequencing techni- 2. Baum M: Overview of chronic kidney dis- migration and chemoattraction of epithe-
cal support and scientific suggestions. ease in children. Curr Opin Pediatr 22: lial cells. J Cell Biol 142: 1337–1345, 1998
Dr. L. Dai designed the study, provided su- 158–160, 2010 16. Skinner MA, Safford SD, Reeves JG, Jack-
pervision of the research workflow and pro- 3. Kari J: Epidemiology of chronic kidney son ME, Freemerman AJ: Renal aplasia in
cedures, conducted the animal experiments, disease in children. J Nephropathol 1: humans is associated with RET mutations.
and wrote the manuscript; Dr. J. Huang ana- 162–163, 2012 Am J Hum Genet 82: 344–351, 2008
lyzed the next-generation sequencing data; 4. dos Santos Junior AC, de Miranda DM, 17. Humbert C, Silbermann F, Morar B,
Mrs. J. Li performed the experiments of in Sim~oes e Silva AC: Congenital anomalies Parisot M, Zarhrate M, Masson C, et al.:
vivo overexpression and the detection of of the kidney and urinary tract: An em- Integrin alpha 8 recessive mutations are
mRNA and protein expression; Mrs. Y. Lu bryogenetic review. Birth Defects Res C responsible for bilateral renal agenesis in
collected the samples and information from Embryo Today 102: 374–381, 2014 humans. Am J Hum Genet 94: 288–294,
5. van der Ven AT, Vivante A, Hildebrandt F: 2014
patients; Prof. C. Pei, Dr. W. Wang, and
Novel insights into the pathogenesis of 18. Sariola H, Saarma M: Novel functions and
Prof. Y. Zhao were the treating physicians;
monogenic congenital anomalies of the signalling pathways for GDNF. J Cell Sci
Prof. K. Shen and Mrs. H. Yang operated the
kidney and urinary tract. J Am Soc Neph- 116: 3855–3862, 2003
level two ultrasonography scan and provided
rol 29: 36–50, 2018 19. Keefe Davis T, Hoshi M, Jain S: Stage
ultrasound images of the affected fetuses; Dr.
6. Arora V, Khan S, El-Hattab AW, Dua Puri specific requirement of Gfra1 in the ure-
L. Xie performed the cell culture experi-
R, Rocha ME, Merdzanic R, et al.: Biallelic teric epithelium during kidney develop-
ments; Dr. M. Xie performed the histological
pathogenic GFRA1 variants cause autoso- ment. Mech Dev 130: 506–518, 2013
analysis experiments; and Prof. W. Zhang
mal recessive bilateral renal agenesis. J 20. Moore MW, Klein RD, Fari~ nas I, Sauer H,
was the primary treating physician, designed
Am Soc Nephrol 32: 223–228, 2020 Armanini M, Phillips H, et al.: Renal and
the study, and edited the manuscript.
7. De Tomasi L, David P, Humbert C, Silber- neuronal abnormalities in mice lacking
mann F, Arrondel C, Tores F, et al.: Muta- GDNF. Nature 382: 76–79, 1996
tions in GREB1L cause bilateral kidney 21. Pichel JG, Shen L, Sheng HZ, Granholm
SUPPLEMENTAL MATERIAL agenesis in humans and mice. Am J Hum AC, Drago J, Grinberg A, et al.: Defects
Genet 101: 803–814, 2017
in enteric innervation and kidney develop-
8. van der Ven AT, Kobbe B, Kohl S, Shril S,
This article contains the following supplemen- ment in mice lacking GDNF. Nature 382:
Pogoda HM, Imhof T, et al.: A homozy-
tal material online at http://jasn.asnjournals.org/ 73–76, 1996
gous missense variant in VWA2, encoding
lookup/suppl/doi:10.1681/ASN.2020121762/-/ 22. Sanchez MP, Silos-Santiago I, Fris en J, He
an interactor of the Fraser-complex, in a
DC Supplemental. B, Lira SA, Barbacid M: Renal agenesis
patient with vesicoureteral reflux. PLoS
Supplemental Table 1. Quality data of WES for and the absence of enteric neurons in
One 13: e0191224, 2018
the investigated family. mice lacking GDNF. Nature 382: 70–73,
9. Kitzler TM, Schneider R, Kohl S, Kolven-
Supplemental Table 2. Primers used for PCR, 1996
bach CM, Connaughton DM, Dai R, et al.:
quantitative PCR, cloning, and vector construc- 23. Linton JM, Martin GR, Reichardt LF: The
COL4A1 mutations as a potential novel
tion were in the list. ECM protein nephronectin promotes
cause of autosomal dominant CAKUT in
Supplemental Table 3. Fetal growth index of kidney development via integrin alpha8-
humans. Hum Genet 138: 1105–1115,
the affected fetus (II:4). beta1-mediated stimulation of Gdnf ex-
2019
Supplemental Table 4. The LOH regions pression. Development 134: 2501–2509,
10. Yang N, Wu N, Dong S, Zhang L, Zhao Y,
shared by the two affected fetuses. 2007
Chen W, et al.: Human and mouse studies
Supplemental Table 5. The candidate homozy- 24. Schuchardt A, D’Agati V, Larsson-Blom-
establish TBX6 in Mendelian CAKUT and
gous variants identified in the family. berg L, Costantini F, Pachnis V: Defects in
as a potential driver of kidney defects as-
Supplemental Table 6. Data of nine litters the kidney and enteric nervous system of
sociated with the 16p11.2 microdeletion
transmitted from the F1 heterozygote (NpntK260*/ syndrome. Kidney Int 98: 1020–1030, mice lacking the tyrosine kinase receptor
WT
) mice intercross. 2020 Ret. Nature 367: 380–383, 1994
Supplemental Table 7. Statistics in body weight 11. Saygili S, Atayar E, Canpolat N, Elicevik 25. Brandenberger R, Schmidt A, Linton J,
and height between the NpntWT/WT and M, Kurugoglu S, Sever L, et al.: A homo- Wang D, Backus C, Denda S, et al.: Iden-
NpntK260*/WT mice. zygous HOXA11 variation as a potential tification and characterization of a novel
Supplemental Table 8. Statistics in body weight novel cause of autosomal recessive con- extracellular matrix protein nephronectin
and height between the NpntWT/WT and genital anomalies of the kidney and uri- that is associated with integrin alpha8-
NpntK260*/K260* mice. nary tract. Clin Genet 98: 390–395, 2020 beta1 in the embryonic kidney. J Cell Biol
Supplemental Figure 1. Ultrasound images of 12. Rieke JM, Zhang R, Braun D, Yilmaz O, € 154: 447–458, 2001
affected fetuses (II:2, II:3) Japp AS, Lopes FM, et al.: SLC20A1 is in- 26. Huang JT, Lee V: Identification and char-
Supplemental Figure 2. Histological analysis of volved in urinary tract and urorectal devel- acterization of a novel human nephronec-
third affected fetus (II:4) at 17th week of gesta- opment. Front Cell Dev Biol 8: 567, 2020 tin gene in silico. Int J Mol Med 15:
tion after anatomy. 13. Costantini F, Shakya R: GDNF/Ret signal- 719–724, 2005
Supplemental Figure 3. Histological analysis of ing and the development of the kidney. 27. Morimura N, Tezuka Y, Watanabe N,
homozygote mice at 0 dpp. BioEssays 28: 117–127, 2006 Yasuda M, Miyatani S, Hozumi N, et al.:
14. Jing S, Wen D, Yu Y, Holst PL, Luo Y, Molecular cloning of POEM: A novel ad-
Fang M, et al.: GDNF-induced activation hesion molecule that interacts with al-
REFERENCES
of the ret protein tyrosine kinase is medi- pha8beta1 integrin. J Biol Chem 276:
ated by GDNFR-alpha, a novel receptor 42172–42181, 2001
1. Caruana G, Bertram JF: Congenital anom- for GDNF. Cell 85: 1113–1124, 1996 28. M€ uller U, Wang D, Denda S, Meneses JJ,
alies of the kidney and urinary tract genet- 15. Tang MJ, Worley D, Sanicola M, Dressler Pedersen RA, Reichardt LF: Integrin al-
ics in mice and men. Nephrology (Carlton) GR: The RET-glial cell-derived neurotro- pha8beta1 is critically important for epi-
20: 309–311, 2015 phic factor (GDNF) pathway stimulates thelial-mesenchymal interactions during
1878 JASN JASN 32: 1871–1879, 2021www.jasn.org RAPID COMMUNICATIONS
kidney morphogenesis. Cell 88: 603–613, A tool set for whole-genome association prediction for the deep-sequencing age.
1997 and population-based linkage analyses. Nat Methods 11: 361–362, 2014
29. Yang Y, Guo J, Dai L, Zhu Y, Hu H, Tan L, Am J Hum Genet 81: 559–575, 2007 37. Hentze MW, Kulozik AE: A perfect mes-
et al.: XRCC2 mutation causes meiotic ar- 33. Wang K, Li M, Hakonarson H: ANNOVAR: sage: RNA surveillance and nonsense-
rest, azoospermia and infertility. J Med Functional annotation of genetic variants mediated decay. Cell 96: 307–310,
Genet 55: 628–636, 2018 from high-throughput sequencing data. 1999
30. Li H, Durbin R: Fast and accurate long- 38. Yeung G, Mulero JJ, Berntsen RP, Loeb
Nucleic Acids Res 38: e164, 2010
read alignment with Burrows-Wheeler DB, Drmanac R, Ford JE: Cloning of a
34. Kunkel TA, Bebenek K, McClary J: Effi-
transform. Bioinformatics 26: 589–595, novel epidermal growth factor repeat
cient site-directed mutagenesis using ura-
2010 containing gene EGFL6: Expressed in tu-
cil-containing DNA. Methods Enzymol
31. McKenna A, Hanna M, Banks E, Sivachen- mor and fetal tissues. Genomics 62:
ko A, Cibulskis K, Kernytsky A, et al.: The 204: 125–139, 1991 304–307, 1999
Genome Analysis Toolkit: A MapReduce 35. Li Q, Wang K: InterVar: Clinical interpreta- 39. Buchner G, Orfanelli U, Quaderi N, Bassi
framework for analyzing next-generation tion of genetic variants by the 2015 MT, Andolfi G, Ballabio A, et al.: Identifi-
DNA sequencing data. Genome Res 20: ACMG-AMP Guidelines. Am J Hum Genet cation of a new EGF-repeat-containing
1297–1303, 2010 100: 267–280, 2017 gene from human Xp22: A candidate for
32. Purcell S, Neale B, Todd-Brown K, Thom- 36. Schwarz JM, Cooper DN, Schuelke M, developmental disorders. Genomics 65:
as L, Ferreira MA, Bender D, et al.: PLINK: Seelow D: MutationTaster2: Mutation 16–23, 2000
JASN 32: 1871–1879, 2021 NPNT in Human Bilateral Renal Agenesis 1879You can also read

















































