The DJ-1L166P mutant protein associated with early onset Parkinson's disease is unstable and forms higher-order protein complexes
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Human Molecular Genetics, 2003, Vol. 12, No. 21 2807–2816
DOI: 10.1093/hmg/ddg304
The DJ-1L166P mutant protein associated with
early onset Parkinson’s disease is unstable and
forms higher-order protein complexes
Maria G. Macedo1,2, Burcu Anar1,2, Iraad F. Bronner1,2, Milena Cannella3,
Ferdinando Squitieri3, Vincenzo Bonifati2,4, André Hoogeveen2,
Peter Heutink1,2,* and Patrizia Rizzu1,2
1
Department of Human Genetics, Section of Medical Genomics, VU University Medical Center, 1081 BT Amsterdam,
The Netherlands, 2Department of Clinical Genetics, ErasmusMC, 3000 DR Rotterdam, The Netherlands,
3
Neurogenetics Unit, IRCCS Neuromed, 86077, Pozzilli, Italy and 4Department of Neurological Sciences,
La Sapienza University, 00185 Rome, Italy
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
Received June 29, 2003; Revised and Accepted August 27, 2003
Parkinson’s disease (PD) is a common neurodegenerative disorder that involves the selective degeneration
of midbrain dopaminergic neurons. Recently DJ-1 mutations have been linked to autosomal-recessive early-
onset Parkinsonism in two European families. By using gel filtration assays under physiological conditions
we demonstrate that DJ-1 protein forms a dimeric structure. Conversely, the DJ-1L166P mutant protein shows
a different elution profile as compared with DJ-1WT both in overexpression cellular systems or in lympho-
blasts cells, suggesting that it might form higher order protein structures. Furthermore we observed that
the level of DJ-1L166P mutant protein in the patient’s lymphoblasts was very low as compared with the wild-
type protein. We excluded a potential transcriptional impairment by performing quantitative RT–PCR on the
patient’s material. Pulse-chase experiments in transfected COS-1 cells and cycloheximide treatment in control
and patient lymphoblasts indicated that the mutant protein was rapidly degraded. This rapid turnover and the
structural changes of DJ-1L166P mutant protein might be crucial in the disease pathogenesis.
INTRODUCTION mechanisms (4–6). Recently we reported cloning of the DJ-1
gene, for early-onset recessive PD (7). Other genes identified
Parkinson’s disease (PD) is a progressive age-related movement for familiar PD are a-synuclein (8), parkin (9) ubiquitin
disorder with a prevalence of 2% in the population aged over C-terminal hydrolase L (UCH-L1) (10), and NR4A2 (11).
65 (1). Clinically, it is characterized by bradykinesia, muscular Despite the remarkable progress in recent years, the
rigidity, resting tremor and postural instability, which are due etiopathogenesis of PD remains elusive. Several hypotheses
to the progressive and selective loss of dopaminergic neurons have been put forward. Findings in post-mortem PD brains and
in the substantia nigra pars compacta. Insoluble protein biochemical studies provide support for two main theories
aggregates known as Lewy bodies (LBs) are seen in the cyto- related to oxidative and/or proteolytic stress (12,13). Particular
plasm of dopaminergic neurons and other subcortical and interest in the concept of proteolytic stress has arisen because
cortical structures. LBs are fibrillary inclusions that mainly of the E3 ligase activity of the parkin protein. Parkin mutations
consist of a-synuclein and ubiquitin (2). associated with familial PD potentially interfere with normal
Although the etiology of the majority of PD cases is still protein degradation through the ubiquitin–proteasome system.
unknown it is likely to be a combination of genetic and Decreasing the E3 activity of parkin results in the accumulation
environmental factors. The genetic loci and genes linked with of target proteins such as CDC-cel1 (14), PaelR1 (15), or
familial PD, which account for less than 10% of all cases of PD glycosylated a-synuclein (16). Furthermore, there is growing
(3), have provided tremendous insight into the pathogenesis of evidence that the normal metabolism of dopamine in itself
this disorder by elucidating some of the molecular pathogenic might be responsible for high basal levels of oxidative stress in
*To whom correspondence should be addressed at: Department of Human Genetics, Section Medical Genomics, VU University Medical Center, Van der
Boechorststraat 7, 1081 BT Amsterdam, The Netherlands. Tel: þ31 204449962; Fax: þ31 204448285; Email: heutink@cncr.vu.nl
Human Molecular Genetics, Vol. 12, No. 21 # Oxford University Press 2003; all rights reserved2808 Human Molecular Genetics, 2003, Vol. 12, No. 21
the substantia nigra, due to auto-oxidation of dopamine, the straightforward structural explanation for the mutant’s dysfunc-
subsequent semiquinone formation and polymerization through tion: proline is a strong helix breaker and its presence in the
which radical species are produced. The hydroxyl radical mutant is therefore likely to destabilize the terminal helix,
produced can damage proteins, nucleic acids and membrane which forms part of the trimerization region, and provides a
phospholipids, potentially leading to cellular degeneration. scaffold for the loop formed by residues S155–P158, which is
By studying two families from genetically isolated commu- part of the active site cleft. The general pattern of salt bridges
nities in The Netherlands and Italy, two mutations in the DJ-1 and hydrophobic packing in the trimerization region of PH1704
gene were found to be associated with autosomal recessive is conserved in the DJ-1 model, which makes it likely that DJ-1
early-onset PD (7). In the Dutch family, a large homozygous also forms higher aggregates (7). Therefore, here we investi-
genomic deletion removes approximately 14 kb of genomic gated this hypothesis and demonstrated that, while DJ-1WT
DNA, including the first five exons of DJ-1 and approximately protein is likely to form dimers, in contrast DJ-1L166P forms
4 kb of promoter sequence. The result of this mutation is that higher-order structure. The suggested structural changes and
the expression of DJ-1 is completely abolished. In the Italian the resulting aberrant complex formation might have serious
family, a homozygous point mutation was identified, resulting consequences for the normal biological function of DJ-1.
in the substitution of a highly conserved leucine residue at In addition we report that the expression of DJ-1L166P in
position 166 of the DJ-1 protein by a proline. patient lymphoblasts cells is extremely low compared with
DJ-1 is ubiquitously expressed in human tissues (7,17) and is DJ-1WT in control samples, suggesting that an abnormal
present in both the nucleus and cytoplasm of mammalian cells. turnover of the mutant protein might play an important role
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
Although its exact biological function is still largely unknown, in the pathogenic mechanism as well.
previous studies suggest several putative roles for DJ-1. In
particular there is evidence that DJ-1 might play an important
role in the post-transcriptional control of gene expression. Hod RESULTS
et al. (18) identified DJ-1 as a regulatory subunit (RS) of an
unknown 400 kDa complex (RBP) that contains an RNA- Intracellular localization of DJ-1L166P
binding activity, and demonstrated that the purified recombi-
nant RS has the capacity to bind the complex and inhibit the DJ-1WT shows diffuse cytoplasmic and nuclear immunoreac-
RNA-binding activity. DJ-1 further acts as a positive tivity in several mammalian cells including COS, HeLa and
transcriptional co-regulator of the androgen receptor (AR) by PC12 cells (7,17,18,24). DJ-1L166P, in transfected COS and
preventing PIASxa/ARIP3 (androgen receptor interacting PC12 cells, has a similar uniform nuclear staining, whereas the
protein 3) and DJBP (DJ-1 binding protein) from binding to cytoplasmic staining appeared to mostly co-localize with
AR (17,19). PIASxa/ARIP3 is a member of the PIAS (protein mitochondria (7). Since in the DJ-1 molecular model, L166
inhibitor of activated STAT) protein family that not only is placed right in the middle of a carboxyl-terminal helix, it was
interacts with SUMO-1 but also functions as an E3 type SUMO necessary to rule out the possibility that the mitochondrial
protein ligase (20). PIASxa was first characterized as an AR localization was a direct consequence of using a C-terminal
binding protein that specifically down-regulates the transcrip- tagged construct. We therefore transfected COS-1 cells with
tional activity of the AR. DJ-1 directly binds to the AR binding DJ-1WT and DJ-1L166P expression constructs without added
region of PIASxa, thereby absorbing PIASxa from the AR– tags. DJ-1 proteins were visualized by indirect immunofluor-
PIASxa complex. DJBP directly binds to both DJ-1 and AR in escence using our recently generated rabbit polyclonal antibody
a testosterone-dependent manner and it negatively modulates (SN1132) (25) or a mouse monoclonal anti-DJ-1 antibody, and
the AR transcription activity by recruiting histone–deacetylase FITC or TRITC-conjugated secondary antibodies. Both anti-
complexes. DJ-1 antagonizes this inhibition by abrogation of bodies did not stain endogenous DJ-1 in untransfected COS-1
the complexes, thereby restoring AR activity. cells. However on transfected cells the results confirmed our
Other studies suggest that DJ-1 is a protein responsive to prior studies, showing mitochondrial localization in about 50%
oxidative stress resulting from exposure to H2O2 or the of the cells transfected with DJ-1L166P (Fig. 1). The remaining
pesticide paraquat (21,22), that might function as an indicator half showed diffuse cytoplasmic and nuclear staining as seen
of oxidative stress state in vivo since it is converted into a variant for DJ-1WT protein. These results were not dependent on the
with a lower pI (pI 5.8) on two-dimensional gels in response to transfection time or cell density.
small amounts of reactive oxygen species (ROS) produced
during aerobic metabolism in murine cells and tissues (22). DJ-1 forms dimers and L166P mutation affects
We have proposed a ‘loss of function’ mechanism to explain DJ-1 complex formation
the pathogenicity of both DJ-1 mutations in the Dutch and
Italian families (7); however the molecular mechanism under- The predicted DJ-1 molecular model led to the assumption that
lying the DJ-1L166P mutation needs to be clarified. Based on the DJ-1 (19.9 kDa) is likely to form higher order structures, and
structure of the PH1704 protease from Pyrococcus horikoshii the predicted structural consequences and the mitochondrial
(23), we predicted a molecular model of DJ-1 and assumed that localization of DJ-1L166P could represent higher-order protein
DJ-1 adopts the same a/b sandwich structure as PH1704. The complexes. This hypothesis was investigated by performing gel
latter was found to form a hexameric ring structure, which can filtration assays. As DJ-1 is highly expressed in most
be regarded as a trimer of dimers with the active sites in the mammalian cells, cytoplasmic lysates from COS, HeLa and
dimerization regions. According to our model, L166P is placed lymphoblasts cells were fractionated on a Superdex 200 PC
right in the middle of a C-terminal helix, providing a precision column using physiological conditions (150 mMHuman Molecular Genetics, 2003, Vol. 12, No. 21 2809
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
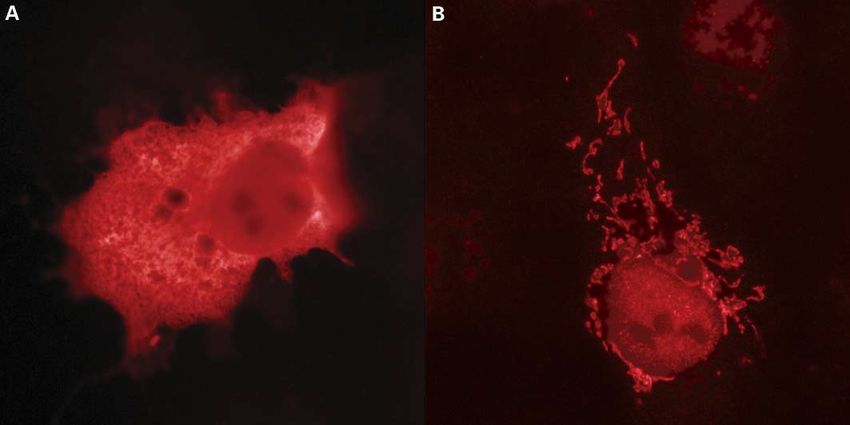
Figure 1. DJ-1L166P co-localizes with mitochondria. COS-1 cells were transfected with non-tagged DJ-1WT (A) and DJ-1L166P (B) constructs. DJ-1 expression was
observed by indirect immunofluorescence using the SN1132 antibody and TRITC-conjugated anti-rabbit IgG.
NaCl). The collected fractions were analyzed for the presence mutant physiological profiles were not affected by the 500 mM
of endogenous DJ-1 by western blotting using our SN1132 NaCl treatment, suggesting that DJ-1 interactions are strong
antibody. DJ-1 from untransfected COS and HeLa cells was enough to hold even in the presence of high salinity levels.
eluted mainly in fraction number 23, corresponding to a However, in the presence of 1 M NaCl both wild-type and
molecular weight of 35 kDa (data not shown), suggesting mutant DJ-1 ran as a 68 kDa protein complex. It appears that
therefore that DJ-1 might form a dimer, in agreement with the very high non-physiological salt concentrations induced the
predicted molecular model. The consequences on complex for- wild-type protein to behave as the mutant protein, forming
mation of the L166P mutation were then investigated. COS-1 higher order structures.
cells were transfected with a DJ-1WT/V5 or DJ-1L166P/V5
construct. The eluted fractions were visualized by using an
anti-V5 antibody. The molecular weight of DJ-1WT/V5 is L166P does not affect DJ-1 interaction with
25 kDa and was mainly present in fractions number 22 and PIASxa/ARIP3 protein
23, corresponding to a molecular weight of 35–45 kDa
(Fig. 2A). This is consistent with DJ-1WT being a dimer. In Since there is an effect of the L166P mutation on cellular
contrast, DJ-1L166P is present in fractions 19 and 20 (Fig. 2A), localization and complex formation of DJ-1, we then tested
corresponding to the elution profile of the protein marker BSA whether known protein interactions would be disrupted by
(68 kDa). These results would suggest that DJ-1L166P forms the mutation. Previous studies have shown an interaction
higher-order structures. between DJ-1 and PIASxa/ARIP3 (24). To determine whether
Since these experiments were performed using an over- DJ-1L166P retained this capacity, we transiently co-transfected
expression system, we further characterized endogenous DJ-1 COS-1 cells with either DJ-1WT/V5 or DJ-1L166P/V5 and
in lymphoblast cells (Fig. 2B) originating from the patient FLAG-PIASxa/ARIP3 or FLAG-PIASxa/ARIP3D347-418 con-
carrying the homozygous L166P point mutation and from a structs. FLAG-PIASxa/ARIP3D347-418 lacks the putative zinc-
healthy control individual. Once again DJ-1L166P was eluted binding structure containing the conserved cysteines and
one fraction earlier than DJ-1WT, strengthening the suggestion histidine necessary for its interaction with the E2 SUMO-1
that DJ-1 in normal conditions might assume a dimer conjugase (26). From total cell lysates, proteins were
conformation and that the L166P mutation affects the structure immunoprecipitated with an anti-V5 antibody and successively
in such a way that DJ-1L166P forms larger protein complexes. visualized with an anti-FLAG antibody. As shown in Figure 3,
In order to study the strength of the potential DJ-1/DJ-1 DJ-1L166P interacted efficiently with both PIASxa/ARIP3 wild-
interaction, the experiments were repeated using high salt lysis type and PIASxa/ARIP3D347-418, suggesting therefore that the
buffers. Two different salt concentrations were tested: 500 mM L166P mutation does not affect the DJ-1 capacity to bind to
and 1 M NaCl (Fig. 2C), respectively. DJ-1 wild-type and PIASxa/ARIP3 proteins.2810 Human Molecular Genetics, 2003, Vol. 12, No. 21
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
Figure 2. Distribution of DJ-1 after gel filtration. (A) and (C) elution fractions from COS-1 cells transfected with DJ-1WT/V5 or DJ-1L166P/V5 lysated by using
150 mM (A) and 1 M NaCl (C), respectively. (B) Elution fractions from lymphoblast cells from a healthy control (2.5 106) and from the Italian patient carrying the
DJ-1L166P mutation (5 106) lysated in lysis buffer containing 150 mM NaCl. DJ-1 was detected using SN1132 and V5 antibodies.
To investigate whether the L166P mutation might have an subset of such tau aggregates in the brains of several
effect on the nuclear co-localization of PIASxa/ARIP3 and DJ-1 tauopathies including Pick’s disease and Alzheimer’s disease
in mammalian cells (24), transfected COS-1 cells were stained (25), demonstrating a possible role of DJ-1 in the pathogenesis
with a mouse monoclonal anti-V5 and a rabbit polyclonal anti- of both dementia and Parkinsonism. Here we performed co-
FLAG antibodies, DJ-1 and PIASxa/ARIP3 proteins were transfection experiments in COS-1 cells with both DJ-1WT/V5
subsequently detected with FITC and TRITC-conjugated and DJ-1L166P/V5 and tau constructs to determine if the
secondary antibodies, respectively. No difference was observed observed co-localization and/or interaction between DJ-1 and
in the nuclear co-localization of DJ-1L166P/V5 with PIASxa/ tau might also occur in non-pathological conditions. After 24
ARIp3 as compared with DJ-1 wild-type (data not shown). or 48 h of transfection, cytoplasmic lysates were immunopre-
cipitated with a phosphorylation-independent anti-tau antibody
DJ-1 does not directly interact with tau and analyzed by immunoblotting with anti-V5 and anti-tau
and alpha-synuclein antibodies. No co-immunoprecipitation of DJ-1 proteins was
observed (data not shown) suggesting that the co-localization
Tau protein aggregates such as neurofibrillary tangles and Pick seen in tau aggregates might be dependent on the hyperpho-
bodies are the pathological hallmark of many neurodegenerative sphorylation state of tau protein in brain aggregates or is
disorders. We recently observed co-localization of DJ-1 in a mediated by a third as yet unknown protein.Human Molecular Genetics, 2003, Vol. 12, No. 21 2811
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
Figure 3. DJ-1wt and DJ-1L166P interact with PIAS. COS-1 cells were co-transfected with DJ-1WT/V5 or DJ-1L166P/V5 and FLAG-PIASxa/ARIP3 or PIASxa/
ARIP3D347-418 constructs. Five percent of the cell extracts were immunoblotted with the anti-V5 antibody (lanes 1–4) and anti-FLAG antibody (lanes 5–8),
and the rest of the samples were immunoprecipitated with the anti-V5 antibody followed by immunoblotting with the anti-FLAG antibody (lanes 10–13). IP, immu-
noprecipitation; WB, western blotting.
To investigate whether DJ-1 is able to interact with the homozygous mutation. Immunoblot experiments were
a-synuclein, the major component of Lewy bodies, we performed with lymphoblast cells (1 and 2.5 106 cells) from
co-transfected COS-1 cells with either DJ-1WT/GFP or the Dutch patient who does not express DJ-1, from the Italian
DJ-1L166P/GFP and a-synuclein/V5 constructs. Cytoplasmic patient carrying the L166P mutation and a control sample. As
lysates were fractionated by gel filtration using physiological shown in Figure 5, the endogenously expressed DJ-1L166P
conditions. The collected fractions were visualized with anti-GFP detected with SN1132 was significantly less than DJ-1WT
and anti-V5 antibodies to detect DJ-1 and a-synuclein, present in the control. To confirm that the same amount of
respectively. As previously shown by Xu et al. (27), a-synuclein protein was loaded in each lane the blot was stripped and
forms 54–83 kDA soluble protein complexes and this pattern relabeled with an antibody against a-tubulin (Fig. 5). To rule
essentially does not change after DJ-1 co-expression (Fig. 4). out individual differences in the expression level of DJ-1
Furthermore we were not able to co-immunoprecipitate DJ-1 protein, we repeated the experiments with several control
and a-synuclein in co-transfection experiments on COS-1 cells samples (data not shown), with similar results. The different
with either DJ-1WT or DJ-1L166P with GFP-tag and alpha- expression level of DJ-1WT and DJ-1L166P might be explained
synuclein-V5 constructs (data not shown), suggesting therefore by differences at transcriptional level or alternatively by
that DJ-1 and a-synuclein do not directly interact. differences in the stability of the normal and mutated protein.
Quantitative RT–PCR experiments were therefore performed
Low level of DJ-1L166P protein compared to on total RNA isolated from lymphoblast cells of the L166P
DJ-1 wild-type patient and control samples using DJ-1 primers that span the
exons 3 and 5 and exons 5 and 6. Primers specific for the HPRT
Throughout our experiments, DJ-1L166P appeared to be poorly gene were also included in the assay to provide a normalization
expressed. To investigate whether this was a biologically control for each sample. We found no significant difference in
relevant finding, COS-1 cells were transfected with either DJ-1 transcription levels between the patient and control
DJ-1WT/V5 or DJ-1L166P/V5 and immunoblotted. Immunoblots samples (data not shown).
of cell lysates showed that the expression level of DJ-1L166P To study possible differences in the stability of the normal and
was significantly lower than the wild-type form. However these mutated protein, we performed pulse chase-experiments and
experiments are not strictly quantitative (data not shown). determined the half-life of DJ-1WT/V5 and its mutant L166P
To exclude the possibility that this finding was artificially counterpart in transiently transfected COS-1 cells. After 3 h
determined by the C-terminal tag present in the eukaryotic labeling with [35S] trans-labeled methionine, newly synthesized
expression vectors used, we investigated the direct effect of the DJ-1 protein was collected at different time points (0, 3, 6 and
L166P mutation on lymphoblast cells from the patient carrying 24 h), and immunoprecipitated with anti V5 antibody. While2812 Human Molecular Genetics, 2003, Vol. 12, No. 21
Figure 4. Distribution of the DJ-1 and a-synuclein proteins after gel filtration. COS-1 cells were transfected with a-synuclein/V5 construct (A) and co-transfected
with a-synuclein/V5 and DJ-1WT/GFP (B) or DJ-1L166P/GFP constructs. Subsequently the samples were separated by gel filtration, and each collected fraction was
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
resolved by 12% SDS–PAGE and tested for the presence of a-synuclein and DJ-1 with anti-V5 and anti-GFP antibodies, respectively. The approximate molecular
size of DJ-1/GFP is 50 kDA and a-synuclein/V5 is 25 kDa.
DJ-1WT/V5 was stable over time, DJ-1L166P/V5 disappeared Although it might be relevant to determine the exact structure
almost completely after 3 h (Fig. 6A). A similar turnover rate for of DJ-1L166P, the structural change might not be the crucial
DJ-1L166P was observed when we inhibited protein synthesis in event for the disease pathogenesis as we have demonstrated that
lymphoblast cells using cycloheximide. A rapid drop of the the mutation does not alter the known protein interaction with
mutant protein was seen between 90 and 120 min, confirming PIASxa/ARIP3.
therefore that DJ-1L166P is a short-lived protein while DJ-1WT Recently, two independent studies reported the crystal
appeared to be a more stable protein whose abundance hardly structure of DJ-1, revealing that the wild-type protein indeed
varied throughout the time points analyzed. forms dimers and that this dimer formation is probably
correlated to DJ-1 biological function (28,29), in agreement
with our results. Any attempt to obtain the crystal structure of the
mutant protein failed, but light scattering experiments showed
DISCUSSION that it might exist in solution as monomer (28).
The results of the analyses of DJ-1WT by gel fractionating Interestingly, in our earlier study (7) we saw a change in the
assays on either lymphoblast cells and overexpression cellular localization of DJ-1L166P as compared with the wild-type
systems strongly suggest that DJ-1WT forms dimers, as was protein. In both COS-1 cells and PC12 cells we observed a
predicted from the molecular model (7). It was also demon- co-localization of the mutated protein with mitochondria, while
strated that DJ-1L166P is present in a higher order protein DJ-1WT showed a diffuse localization in cytoplasm and nuclei.
complex and that its expression levels in transfection studies We confirmed this result after ruling out the possibility that such
and patients’ lymphoblasts are very low compared with the co-localization was artificially determined by the presence of a
wild-type protein. C-terminal tag in the eukaryotic expression constructs used.
The L166 residue is highly conserved in evolution among the Although the biological relevance of this finding is not yet
DJ-1 proteins and, based on the sequence homology between clear, particularly because DJ-1 does not have a canonic
DJ-1 and the PH1704 from Pyrococcus horikoshi, it was mitochondrial localization signal, we cannot exclude DJ-1L166P
previously suggested that the L166 amino acid is located right being shuttled to mitochondria after binding to other proteins.
in the middle of a carboxyl terminal helix. This carboxyl Interestingly, it has been suggested that DJ-1 has antioxidant
terminal helix is thought to be involved in the formation of activity as it is modified by oxidative stress generated by H2O2
DJ-1 high order structures (7). Therefore the change of the and paraquat. Several studies suggested a prominent role for
166 residue from leucine to proline might have a destabilizing oxidative stress and oxidizing toxins in inducing nigral cell
effect, forcing the dimer to assume a different structure as degeneration. In this respect it is tempting to explain the
compared with the wild-type protein. The shift in molecular detrimental consequences of the L166P mutation by its inability
size observed for the mutant protein in our gel filtration to maintain a dimer formation and by its sequestration in protein
experiments might be a direct consequence of protein complexes co-localized with mitochondria resulting in an
misfolding that affects migration of the protein under native absence of functional protein in the cytoplasm. By developing
conditions. Alternatively, the extended elution pattern might be and using specific DJ-1 antibodies on post-mortem brain from
explained by DJ-1 present as a monomer, forming complexes patients with several neurodegenerative disorders, including
with other unidentified protein. Such binding would be quite Alzheimer’s disease and Pick’s disease, co-localization of DJ-1
strong, as it was not disrupted under high salt conditions. was observed in a subset of tau aggregates that suggests aHuman Molecular Genetics, 2003, Vol. 12, No. 21 2813
Figure 5. Steady-state level of DJ-1. Lymphoblast cells, 1 106 cells (lanes 1,
3 and 5) and 2.5 106 cells (lanes 2, 4 and 6) from a healthy control (DJ-1), a
patient carrying the deletion of exons 1–5 (DJ-1D1-5) and a patient carrying the
L166P point mutation (DJ-1L166P) were lysed in sample buffer and resolved by
12% SDS–PAGE. DJ-1 expression was visualized with SN1132 antibody. After
stripping the blot was labeled with anti-a-tubulin antibody.
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
potential chaperone activity for DJ-1 (25). These results are in
agreement with the finding that the DJ-1 yeast homolog is
strongly up-regulated after treatment with azetidin-2-carboxylic
acid, a toxic analog of proline that activates heat shock factors via
accumulation of thermally misfolded proteins (30).
DJ-1 therefore may perform a chaperone-like role that
protects cells against toxicity from abnormally aggregated
proteins and it may no longer be able to perform this function
when mutated.
Strengthening the arguments for the detrimental effects of the
L166P mutation is the observation of the low expression level
for DJ-1L166P protein in both an overexpression system and in
patient’s lymphoblast cells. By pulse-chase experiments and Figure 6. DJ-1L166P is a short-lived protein. (A) [35S]methionine pulse chase of
cycloheximide assays it was demonstrated that the mutant COS-1 cells transfected with DJ-1WT /V5 or DJ-1L166P /V5. Four time points
were taken at 0, 3, 6 and 24 h. Lysates were immunoprecipitated with anti-
protein is highly unstable as it is rapidly degraded in contrast to V5 antibody, samples were run on 12% SDS–PAGE gel and exposed on phos-
the wild-type protein, which is highly stable. If the steady-state phoimager. (B) Lymphoblast cells (2.5 106 cells) from a control sample and a
level of DJ-1 protein has biological relevance, then it is likely patient carrying the L166P mutation were treated with cycloheximide (final
that an aberrant turn over of the mutant protein might be concentration 50 mg/ml) for the indicated time points. Total lysates were run
responsible for the loss of function of DJ-1. Studies on cystic on 12% and labeled with SN1132 antibody. Different exposure times have been
used to detect DJ-1WT and DJ-1L166P, respectively.
fibrosis (31), a-1-antitrypsin deficiency (32), mitochondrial
acyl-CoA dehydrogenase (33) deficiencies and many other
diseases have indeed shown that enhanced proteolytic degrada- MATERIALS AND METHODS
tion of mutant proteins is a common molecular mechanism
(34–36) and might play a key role here as well. Interestingly, Plasmids and their construction
recently the involvement of the ubiquitin–proteosome pathway
in the degradation of DJ-1L166P has been reported (37). DJ-1WT/V5 and DJ-1L166P/V5 have been previously described
In this context, then, we might also reinterpret the results (7). Coding sequence of DJ-1, DJ-1L166P and alpha-synuclein
demonstrating the interaction of DJ-1L166P with PIASxa/ were amplified by PCR and cloned in pcDNA3.1/CT-GFP-
ARIP3, particularly if the interaction is dose-dependent. Low TOPO and pcDNA.3.1/V5-His TOPO (Invitrogen), respec-
level of expressed DJ-1 might not be sufficient to efficiently tively. For the untagged constructs, DJ-1WT and DJ-1L166P
absorb PIASxa from the AR-PIASxa complex, consequently cDNAs were amplified by reverse transcriptase-treated total
leading to down-regulation of the AR. Furthermore the RNA from a control sample and from a patient carrying L166P
potential antioxidant property and chaperone activity of DJ-1 mutation using the following forward and reverse primers:
would be also jeopardized by the high instability demonstrated 50 gggtgcaggcttgtaaacat 30 and 50 tgacttccatacttccgaa 30 . PCR
by the mutant protein. products were cloned in TOPO TA cloning vector (Invitrogen),
In conclusion, our results would suggest that the reduced and ECO RI fragments were then transferred to pcDNA3.1
stability of DJ-1L166P could well explain the loss of function expression vector (Invitrogen). Fidelity of the constructs was
proposed for the L166P mutation. The dramatic reduction verified by sequencing using the Big Dye terminator version 3
observed in the steady-state level of DJ-1 would consequently (Applied Biosystems). The longest tau isoform containing four
impair the protein capability to perform its normal biological microtubules-binding repeats (4R tau) cloned into pcDNA3.1
functions, leading therefore to Parkinsonism. has been described elsewhere (38).2814 Human Molecular Genetics, 2003, Vol. 12, No. 21
pFLAG-PIASxa/ARIP3 and pFLAG-PIASxa/ARIP3D347-418 Gel electrophoresis and immunoblotting
were kindly provided by Jorma J. Palvimo (26).
Proteins present in total lysates and protein fractions were
released in 2 sodium dodecyl sulfate (SDS) sample buffer,
Antibodies resolved by 12% SDS–polyacrylamide gel electrophoresis
(PAGE) and transferred onto a nitrocellulose membrane
The following primaries antibodies were used: mouse mono- (Schleicher and Schuell). Residual protein-binding sites were
clonal anti-V5 antibody from Invitrogen; mouse monoclonal blocked by incubation of the membrane in PBS supplemented
anti-FLAG antibody M2 from Sigma; mouse monoclonal against with 5% (w/v) dry milk and 0.1% Tween 20 for 1 h at room
green fluorescent protein from Clontech; mouse monoclonal temperature. Immunodetections were made using the appro-
anti-tau (H-7) from Innogenetics; mouse monoclonal anti DJ-1 priate primary antibodies: rabbit polyclonal SN1132 (1 : 1000),
from Stressgen; mouse monoclonal anti a-tubulin from Sigma; mouse monoclonal anti-V5 antibodies (1 : 5000), mouse
and rabbit polyclonal anti DJ-1 SN1132 was developed and monoclonal anti GFP (1 : 1000) for 1 h at room temperature.
characterized by us as described elsewhere (25). Following washing, the membranes were incubated with the
appropriate secondary antibodies conjugated with horseradish
peroxidase and the reaction products were visualized by using
Secondary antibodies. Horseradish peroxidase-conjugated
the enhanced chemiluminescence detection reagents (ECL kit,
anti-rabbit and anti-mouse IgG were purchased from
Amersham Biosciences) according to the manufacturer’s
Amersham and goat anti-mouse FITC-conjugated, and goat
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
instructions.
anti-rabbit TRITC-conjugated were from Sigma.
Co-immunoprecipitation assays
Cell culture and transfection
DJ-1 and PIAS. COS-1 cells were co-transfected with DJ-1/
COS-1 and HeLa cells were cultured in Dulbecco’s modified V5 or DJ-1L166P/V5 and FLAG-tagged PIASxa/ARIP3 or
Eagle’s medium (DMEM) supplemented with 10% (v/v) fetal PIASxa/ARIP3D347-418 constructs. Twenty-four hours after
calf serum (FCS), 100 units/ml penicillin and 100 mg/ml transfection, cells were collected and lysed in physiological
streptomycin (all from Invitrogen) and were kept at 37 C in buffer as previously described (24) and cell homogenates were
5% CO2. Lymphoblast cells from control individuals and the prepared as described above. Five percent of the cell extracts
Italian patient carrying the L166P mutation and from the Dutch were immunoblotted with the anti-FLAG M2 monoclonal
patient carrying the deletion of exons 1–5 were cultured in mouse antibody (1 : 1000), and the rest of the samples were
RPMI 1640 medium supplemented with 10% FCS and immunoprecipitated with 2 mg of monoclonal mouse anti-V5
antibiotics (Invitrogen). The day before transfection COS-1 antibody, overnight at 4 C followed by addition of protein G
cells were seeded on 6 cm-diameter dishes. Cells were Sepharose-beads (Amersham). Bound proteins were detected
transiently transfected with an expression plasmid containing with anti-FLAG antibody (1 : 1000).
DJ-1WT or DJ-1L166P either with V5, GFP or untagged, and
p-FLAG-PIASxa/ARIP3 or p-FLAF-PIASxa/ARIP3D347-418 or DJ-1 and tau. COS-1 cell were co-transfected with DJ-1WT/
a-synuclein/V5, or 4R tau using Lipofectamine Plus reagent V5 or DJ-1 L166P/V5 and 4R tau in pcDNA3. Twenty-four
(Invitrogen) according to the manufacturer’s recommendations. and 48 h after transfection, cells were lysed in physiological
buffer containing 1% NP40 and immunoprecipitated with
either anti-V5 or anti-tau (H-7) antibodies. Detection was per-
Gel filtration formed by using anti-V5 (1 : 5000) and anti-tau (1 : 2000)
A Precision column PC 3.2/30 pre-packed with Superdex 200 antibodies.
was used in a SMART system (Pharmacia) to determine by gel
filtration the molecular mass of DJ-1. The optimal range for DJ-1 and a-synuclein. COS-1 cells were co-transfected with
separation of globular proteins in this column is 10–600 kDa, DJ-1WT/GFP or DJ-1L166P/GFP and a-synuclein/V5. Twenty-
with an exclusion limit of 1300 kDa. In order to calibrate the four hours after transfection, cells were lysed in physiological
column and to determine the molecular masses of the eluting buffer containing 1% NP40 or 0.5% sodium deoxycholate
fractions, four protein markers were applied in each of the three and immunoprecipitated with anti-V5 antibody. Detection
buffers used for protein fractionation: 150 mM NaCl (physio- was performed by using anti-GFP antibody (1 : 1000).
logical buffer), 500 mM and 1 M NaCl (high-salt buffers).
The more salt present in the running buffer, the longer was the DJ-1 half-life. For pulse chase experiments COS-1 cells were
retention time of the protein in the column. Before running, the transfected with DJ-1WT/V5 or DJ-1L166P/V5. Twenty-four
column was equilibrated with lysis buffer [50 mM Tris–HCl hours after transfection, cells were starved in methionine/
(pH 8.0), 5 mM EDTA, 150 NaCl and protease inhibitor cysteine/cystine/glutamine-free DMEM medium (ICN) for
cocktail (Complete from Roche)]. Lysates were homogenized 1 h, pulsed for 3 h with 20 mCi/ml of [35S]methionine/cysteine
by sonication and then centrifuged at 13 000g at 4 C for 10 min (ICN), rinsed and chased for the indicated periods of time in
to obtain a cytoplasmic supernatant. Cytoplasmic lysate (2/3) DMEM complete plus 10% FCS. Cell lysates were obtained
was injected into the SMART system and the protein profile in lysis buffer with 1% NP-40 and were immunoprecipitated
was monitored at 280 nm with a column flow rate of 50 ml/min. with 2 mg anti-V5 antibody and protein G sepharose. Immuno-
Fractions (50 ml each) were collected separately. precipitates were resolved by 12% SDS–PAGE, visualized byHuman Molecular Genetics, 2003, Vol. 12, No. 21 2815
phosphoimaging and quantified with ImageQuant analysis soft- 8. Polymeropoulos, M.H., Lavedan, C., Leroy, E., Ide, S.E., Dehejia, A.,
ware. DJ-1 half-life was investigated in lymphoblast cells by Dutra, A., Pike, B., Root, H., Rubenstein, J., Boyer, R. et al. (1997)
Mutation in the alpha-synuclein gene identified in families with Parkinson’s
inhibiting protein synthesis with 50 mg/ml cycloheximide disease. Science, 276, 2045–2047.
(Sigma) at the time points indicated. Cell pellets were resus- 9. Kitada, T., Asakawa, S., Hattori, N., Matsumine, H., Yamamura, Y.,
pended in sample buffer and after sonication, were analyzed Minoshima, S., Yokochi, M., Mizuno, Y. and Shimizu, N. (1998) Mutations
by immunoblot with SN1132 antibody. in the parkin gene cause autosomal recessive juvenile parkinsonism.
Nature, 392, 605–608.
10. Leroy, E., Boyer, R., Auburger, G., Leube, B., Ulm, G., Mezey, E.,
Harta, G., Brownstein, M.J., Jonnalagada, S., Chernova, T. et al. (1998)
Quantitative RT–PCR The ubiquitin pathway in Parkinson’s disease. Nature, 395, 451–452.
11. Le, W.D., Xu, P., Jankovic, J., Jiang, H., Appel, S.H., Smith, R.G. and
For the quantitative assay the following primer combinations Vassilatis, D.K. (2003) Mutations in NR4A2 associated with familial
were used to amplify DJ-1 and HPRT genes: forward I Parkinson disease. Nat. Genet., 33, 85–89.
50 agagggaccatatgatgtgg 30 and reverse I 50 ctgctccttcagtatctcc 30 12. Jenner, P. (2003) Oxidative stress in Parkinson’s disease. Ann. Neurol., 53,
and forward II 50 ggagatactgaaggagcagg 30 and reverse II 50 S26-36; discussion S36–38.
13. McNaught, K.S. and Olanow, C.W. (2003) Proteolytic stress: a unifying
gcaagagggtgtgttgtaac 30 spanning exons 3 and 5 and exons 5 concept for the etiopathogenesis of Parkinson’s disease. Ann. Neurol., 53,
and 6, respectively for DJ-1; forward 50 gctgaggatttggaaagggtg S73-84; discussion S84–86.
30 and reverse 50 ttgagcacacagagggctaca 30 for HPRT. PCR 14. Chung, K.K., Zhang, Y., Lim, K.L., Tanaka, Y., Huang, H., Gao, J.,
reactions were prepared by using qPCR Core Kit for SYBR Ross, C.A., Dawson, V.L. and Dawson, T.M. (2001) Parkin ubiquitinates
the alpha-synuclein-interacting protein, synphilin-1: implications for
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015
Grenn from Eurogentec and the iCycler iQ termocycler from Lewy-body formation in Parkinson disease. Nat. Med., 7, 1144–1150.
Biorad. 15. Imai, Y., Soda, M., Inoue, H., Hattori, N., Mizuno, Y. and Takahashi, R.
(2001) An unfolded putative transmembrane polypeptide, which can
lead to endoplasmic reticulum stress, is a substrate of Parkin. Cell, 105,
Immunofluorescence 891–902.
16. Shimura, H., Schlossmacher, M.G., Hattori, N., Frosch, M.P.,
The day before transfection, COS-1 cells were seeded on glass Trockenbacher, A., Schneider, R., Mizuno, Y., Kosik, K.S. and Selkoe, D.J.
coverslips. Transfections were performed with Lipofectamine (2001) Ubiquitination of a new form of alpha-synuclein by parkin from
human brain: implications for Parkinson’s disease. Science, 293,
Plus reagent (Invitrogen) and 0.4 mg of DNA as described by 263–269.
the manufacturer. Twenty-four hours after transfection, cells 17. Nagakubo, D., Taira, T., Kitaura, H., Ikeda, M., Tamai, K.,
were rinsed with PBS, fixed in 4% paraformaldehyde and Iguchi-Ariga, S.M. and Ariga, H. (1997) DJ-1, a novel oncogene which
permeabilized with methanol. DJ-1 was detected by using transforms mouse NIH3T3 cells in cooperation with ras. Biochem. Biophys.
Res. Commun., 231, 509–513.
SN1132 (1 : 100), or mouse monoclonal DJ-1 (1 : 100), 18. Hod, Y., Pentyala, S.N., Whyard, T.C. and El-Maghrabi, M.R. (1999)
antibodies and TRITC-conjugated anti-rabbit IgG or FITC- Identification and characterization of a novel protein that regulates
conjugated anti-mouse IgG secondary antibodies (1 : 500). RNA–protein interaction. J. Cell. Biochem., 72, 435–444.
Cells were rinsed and mounted on a glass slide in Vectashield 19. Niki, T., Takahashi-Niki, K., Taira, T., Iguchi-Ariga, S.M. and Ariga, H.
mounting medium (Vector) containing DAPI. Images were (2003) DJBP: a novel DJ-1-binding protein, negatively regulates the
androgen receptor by recruiting histone deacetylase complex, and DJ-1
captured using the Power Gene FISH system on a Leica antagonizes this inhibition by abrogation of this complex. Mol. Cancer
DMRXA microscope at 1000 amplification. Res., 1, 247–261.
20. Jackson, P.K. (2001) A new RING for SUMO: wrestling transcriptional
responses into nuclear bodies with PIAS family E3 SUMO ligases. Genes
Dev., 15, 3053–3058.
ACKNOWLEDGEMENTS 21. Mitsumoto, A. and Nakagawa, Y. (2001) DJ-1 is an indicator for
endogenous reactive oxygen species elicited by endotoxin. Free Radic.
We thank Dr J.J. Palvimo for kindly providing the FLAG- Res., 35, 885–893.
PIASxa/ARIP3 constructs, L. van Unen for technical expertise 22. Mitsumoto, A., Nakagawa, Y., Takeuchi, A., Okawa, K., Iwamatsu, A. and
and T. de Vries Lentsch and R. Koppenol for art work. This Takanezawa, Y. (2001) Oxidized forms of peroxiredoxins and DJ-1 on
work was supported by the Michael J. Fox Foundation. two-dimensional gels increased in response to sublethal levels of paraquat.
Free Radic. Res., 35, 301–310.
23. Du, X., Choi, I.G., Kim, R., Wang, W., Jancarik, J., Yokota, H. and
Kim, S.H. (2000) Crystal structure of an intracellular protease from
Pyrococcus horikoshii at 2-A resolution. Proc. Natl Acad. Sci. USA, 97,
REFERENCES 14079–14084.
1. Lang, A.E. and Lozano, A.M. (1998) Parkinson’s disease. First of two parts. 24. Takahashi, K., Taira, T., Niki, T., Seino, C., Iguchi-Ariga, S.M. and
New Engl. J. Med., 339, 1044–1053. Ariga, H. (2001) DJ-1 positively regulates the androgen receptor by
2. Trojanowski, J.Q., Goedert, M., Iwatsubo, T. and Lee, V.M. (1998) Fatal impairing the binding of PIASx alpha to the receptor. J. Biol. Chem., 276,
attractions: abnormal protein aggregation and neuron death in Parkinson’s 37556–37563.
disease and Lewy body dementia. Cell Death Differ., 5, 832–837. 25. Rizzu, P., Hinkle, D.A., Zhukareva, V., Bonifati, V., Severijnen, L.-A.,
3. Gasser, T. (2001) Genetics of Parkinson’s disease. J. Neurol., 248, 833–840. Martinez, D., Ravid, R., Kamphorst, W., Eberwine, J.H., Lee, V. M.-Y. et al.
4. Giasson, B.I. and Lee, V.M. (2001) Parkin and the molecular pathways of (2003) DJ-1 colocalizes with pathological tau inclusions: A possible link
Parkinson’s disease. Neuron, 31, 885–888. between Parkinsonism and dementia. Ann. Neurol. (in press).
5. Cookson, M.R. (2003) Pathways to Parkinsonism. Neuron, 37, 7–10. 26. Kotaja, N., Karvonen, U., Janne, O.A. and Palvimo, J.J. (2002) PIAS
6. Dawson, T.M. and Dawson, V.L. (2003) Rare genetic mutations shed light proteins modulate transcription factors by functioning as SUMO-1 ligases.
on the pathogenesis of Parkinson disease. J. Clin. Invest., 111, 145–151. Mol. Cell. Biol., 22, 5222–5234.
7. Bonifati, V., Rizzu, P., van Baren, M.J., Schaap, O., Breedveld, G.J., 27. Xu, J., Kao, S.Y., Lee, F.J., Song, W., Jin, L.W. and Yankner, B.A.
Krieger, E., Dekker, M.C., Squitieri, F., Ibanez, P., Joosse, M. et al. (2003) (2002) Dopamine-dependent neurotoxicity of alpha-synuclein:
Mutations in the DJ-1 gene associated with autosomal recessive early-onset a mechanism for selective neurodegeneration in Parkinson disease.
parkinsonism. Science, 299, 256–259. Nat. Med., 8, 600–606.2816 Human Molecular Genetics, 2003, Vol. 12, No. 21
28. Tao, X. and Tong, L. (2003) Crystal structure of human DJ-1, a protein population, as was the case for 625G!A, together conferring susceptibility
associated with early-onset Parkinson’s diseasec. J. Biol. Chem., 278, to ethylmalonic aciduria. Hum. Mol. Genet., 7, 619–627.
31372–31379. 34. Bross, P., Corydon, T.J., Andresen, B.S., Jorgensen, M.M., Bolund, L. and
29. Honbou, K., Suzuki, N.N., Horiuchi, M., Niki, T., Taira, T., Ariga, H. and Gregersen, N. (1999) Protein misfolding and degradation in genetic
Inagaki, F. (2003) The crystal structure of DJ-1, a protein related to male diseases. Hum. Mutat., 14, 186–198.
fertility and Parkinson’s disease. J. Biol. Chem., 8, 8. 35. Gregersen, N., Bross, P., Jorgensen, M.M., Corydon, T.J. and
30. Trotter, E.W., Kao, C.M., Berenfeld, L., Botstein, D., Petsko, G.A. and Gray, J.V. Andresen, B.S. (2000) Defective folding and rapid degradation of mutant
(2002) Misfolded proteins are competent to mediate a subset of the responses to proteins is a common disease mechanism in genetic disorders. J. Inherit.
heat shock in Saccharomyces cerevisiae. J. Biol. Chem., 277, 44817–44825. Metab. Dis., 23, 441–447.
31. Ward, C.L. and Kopito, R.R. (1994) Intracellular turnover of cystic fibrosis 36. Waters, P.J. (2001) Degradation of mutant proteins, underlying ‘loss of
transmembrane conductance regulator. Inefficient processing and rapid function’ phenotypes, plays a major role in genetic disease. Curr. Iss.
degradation of wild-type and mutant proteins. J. Biol. Chem., 269, Mol. Biol., 3, 57–65.
25710–25718. 37. Miller, D.W., Ahmad, R., Hague, S., Baptista, M.J., Canet-Aviles, R.,
32. Wu, Y., Whitman, I., Molmenti, E., Moore, K., Hippenmeyer, P. and McLendon, C., Carter, D.M., Zhu, P.P., Stadler, J., Chandran, J.
Perlmutter, D.H. (1994) A lag in intracellular degradation of mutant alpha et al. (2003) L166P mutant DJ-1, causative for recessive
1-antitrypsin correlates with the liver disease phenotype in homozygous PiZZ Parkinson’s disease, is degraded through the ubiquitin-proteasome
alpha 1-antitrypsin deficiency. Proc. Natl Acad. Sci. USA, 91, 9014–9018. system. J. Biol. Chem., 278, 36588–36595.
33. Gregersen, N., Winter, V.S., Corydon, M.J., Corydon, T.J., Rinaldo, P., 38. Rizzu, P., Joosse, M., Ravid, R., Hoogeveen, A., Kamphorst, W.,
Ribes, A., Martinez, G., Bennett, M.J., Vianey-Saban, C., Bhala, A. et al. Van Swieten, J.C., Willemsen, R. and Heutink, P. (2000) Mutation-
(1998) Identification of four new mutations in the short-chain acyl-CoA dependent aggregation of tau protein and its selective depletion from the
dehydrogenase (SCAD) gene in two patients: one of the variant alleles, soluble fraction in brain of P301L FTDP-17 patients. Hum. Mol. Genet.,
511C!T, is present at an unexpectedly high frequency in the general 9, 3075–3082.
Downloaded from http://hmg.oxfordjournals.org/ by guest on November 10, 2015You can also read

















































