Homeodomain protein HLX is expressed primarily in cytotrophoblast cell types in the early pregnancy human placenta
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
CSIRO PUBLISHING
www.publish.csiro.au/journals/rfd Reproduction, Fertility and Development, 2008, 20, 357–367
Homeodomain protein HLX is expressed primarily
in cytotrophoblast cell types in the early pregnancy
human placenta
Gayathri RajaramanA , Padma MurthiA,C , Leonie QuinnB ,
Shaun P. BrenneckeA and Bill KalionisA
A Pregnancy Research Centre, Department of Perinatal Medicine, Royal Women’s Hospital and
Department of Obstetrics and Gynaecology, RWH Campus, Carlton, Vic. 3053, Australia.
B Department of Anatomy and Cell Biology, University of Melbourne, Parkville, Vic. 3052, Australia.
C Corresponding author. Email: padma@unimelb.edu.au
Abstract. Homeobox genes are a large family of transcription factors. Of these, the HLX homeobox gene (previously
known as HLX1 and HB24) is important for normal placentation. We have previously shown that HLX mRNA expression
is significantly reduced in fetal growth-restricted human placentae compared with control placentae. In this study, a rabbit
polyclonal antibody to the homeodomain protein HLX was raised and characterised. Western analysis revealed a protein
of 50 kDa. HLX protein was detected in cellular nuclei in the cytotrophoblast-derived cell lines HTR8/SVneo, SGHPL-4,
JEG-3, JAR and BeWo. Dual labelling with cytokeratin 7 was used to determine the spatial distribution of HLX in the
early placenta and fetal membranes, showing both a perinuclear and punctate nuclear distribution for HLX. In the early
pregnancy placenta HLX was localised to villous cytotrophoblast, and extravillous cytotrophoblast nuclei in the proximal
regions of the cytotrophoblast cell columns, but was not detected at significant levels in the syncytiotrophoblast. In first
trimester placental bed biopsies, HLX expression was not localised to the nucleus but instead was found in the cytoplasm.
We conclude that HLX is primarily expressed in cytotrophoblast cell types in the human placenta and propose that HLX
is involved in cytotrophoblast proliferation and downregulation of cell differentiation.
Introduction placental morphological defects (Li and Behringer 1998;
Organogenesis of the human placenta involves certain key Morasso et al. 1999; Auman et al. 2002). Homeobox genes not
events including the development of the branched villous tree, only control diverse developmental pathways but also regulate
vascularisation, syncytiotrophoblast membrane formation and specialised functions of trophoblast cells (Roberson et al. 2001;
extravillous trophoblast invasion into the maternal decidua and Peng and Payne 2002).
subsequent spiral artery remodelling. In the mouse, transcription Studies of human placental transcription factors, and home-
factors play crucial roles in organogenesis as shown by targeted obox genes in particular, are still in the early stages. We and
mutagenesis studies. For reviews on this subject see Hemberger others have isolated homeobox genes from the human placenta
and Cross (2001), Rossant and Cross (2001), Sapin et al. (2001) (Quinn et al. 1997a; Roberson et al. 2001). HLX (previously
and Cross et al. (2003). Indeed, some mouse placental mutants known as HB24 and HLX1) was isolated from a 32-week human
may provide useful models to study human pathologies (Sapin placental cDNA library (Quinn et al. 1997a). Limited in situ
et al. 2001; Cross 2003). Thus far, only the basic helix–loop– mRNA hybridisation studies revealed HLX expression in vil-
helix (bHLH) family of transcription factors has been studied lous and extravillous cytotrophoblast cells in the first trimester
intensively, providing important insights into the genetic control placenta, and expression in residual extravillous cytotrophoblast
of placentation (Westerman et al. 2003). The bHLH transcrip- cells at term (Quinn et al. 1997b). Furthermore, homozygous
tion factors are likely to interact with another important family of mouse knockouts for Hlx are found to be lethal at E15 due to
transcription factors called homeobox genes (Westerman et al. severe liver defects, causing an inability to maintain the develop-
2002, 2003). ing fetus (Hentsch et al. 1996). Finally, we have shown that there
Homeobox genes are a large family of transcriptional regu- is severe disruption of placental morphology in Hlx homozygous
lators that control embryonic development by initiating genetic mutant mice (M. Bates, P. Murthi, G. Rajaraman and B. Kalionis,
pathways of development of many cell types and tissues (Krum- unpubl. data).
lauf 1994). Proof of the importance of homeobox genes in In a previous study we used the HLX antibody to describe the
placentation comes from the mouse model system, where tar- HLX expression pattern in human term placentae by immuno-
geted disruption of Cdx2, Esx1 and Dlx3 causes severe, diverse histochemistry analysis (Murthi et al. 2006). Expression of HLX
© CSIRO 2008 10.1071/RD07159 1031-3613/08/030357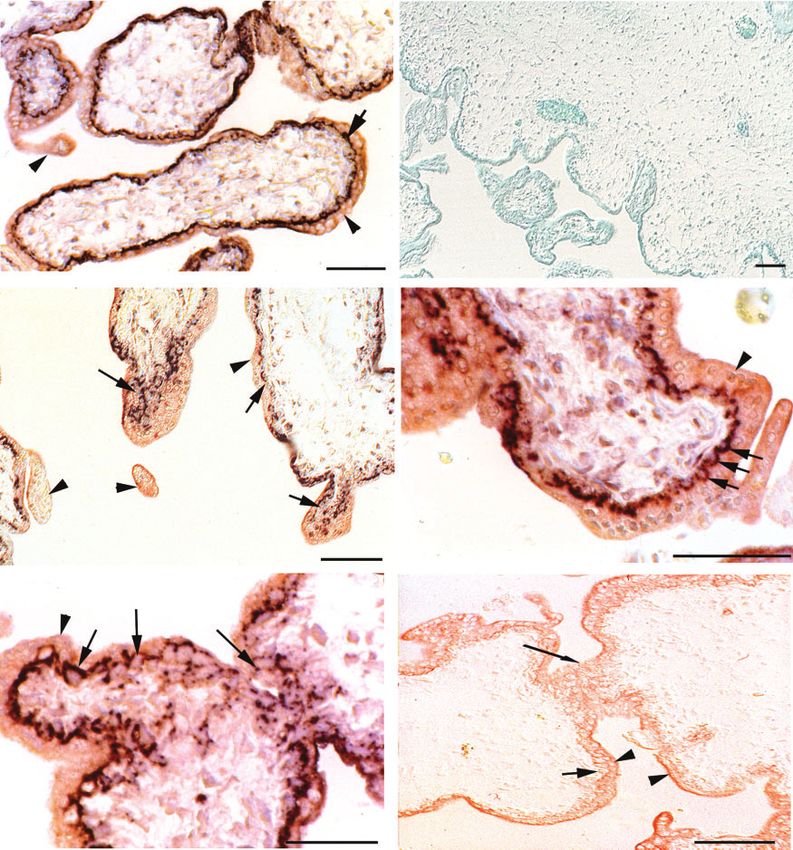
358 Reproduction, Fertility and Development G. Rajaraman et al.
was restricted to residual villous cytotrophoblast cells at term mixture (Invitrogen) and HTR-8/SVneo were grown in RPMI-
and this was consistent with mRNA studies carried out pre- 1640, supplemented with 10% FCS, 100 U mL−1 penicillin and
viously. Western analysis and immunohistochemistry showed 100 µg mL−1 streptomycin.
reduced expression levels of HLX in placentae affected with
fetal growth restriction compared with gestation age-matched Immunocytochemistry on cultured cells
control placentae (Murthi et al. 2006).
Antibody staining on cultured cells employed four-chamber slide
In the present study we characterise HLX expression in early
flasks (Medos, Unley, South Australia, Australia). Cells were
pregnancy placental tissues, placental bed biopsies and placental
cultured at 37◦ C in RPMI-1640 medium containing 10% FCS,
cell lines representative of extravillous trophoblast.
100 U mL−1 penicillin and 100 µg mL−1 streptomycin. Cells
were fixed for 20 min in 10% formaldehyde/PBS and perme-
Materials and methods abilised in methanol for 3 min followed by acetone for 1 min.
The following research protocols have been approved by the Immunostaining was performed as described below.
Research and Ethics committee of the Royal Women’s Hospi-
tal, Victoria, Australia and the Research Ethics committee of Antibody preparation
Flinders Medical Centre, Flinders University of South Australia,
A 432 bp fragment of HLX cDNA (nucleotides 1436–
Australia.
1908 GenBank Accession number BC007294) was amplified
with PCR primers incorporating BamHI/EcoRI cloning sites
Sample collection (forward primer –5 GCGGATCCGGCACTCCAAGGAGGC3
Early pregnancy placental tissues were obtained at 12–14 reverse primer –5 GCGAATTCTATAAGCAGCCAAGCG3 ),
weeks gestation following therapeutic terminations of preg- subcloned into pGEX3 (Oettinger et al. 1992) and transformed
nancy. Informed patient consent for sampling was obtained from into the E. coli DH5-α strain. The cDNA region used corre-
all donors of early pregnancy placental tissue in accordance with sponded to the C-terminal region of the HLX coding sequence,
the Committee on Clinical Investigation, Flinders Medical Cen- did not include the homeodomain and showed no significant
tre, South Australia. All data presented are representative of at homology to other cDNA sequences in database searches (data
least n = 5 samples. All experiments were carried out at least in not shown). HLX-pGEX3 fusion protein was prepared by induc-
triplicate. ing fusion protein with 0.1 mm IPTG; cells were harvested by
centrifugation (2300g for 5 min at room temperature) and lysed
Placental bed biopsies in protein lysis buffer (50 mm NaCl, 1 mm EDTA, 1% (v/v)Triton
X-100). The sample was sonicated and cell debris was removed
Human placental bed biopsy tissue was obtained from thera-
by centrifugation (5750g for 10 min at 4◦ C). The supernatant
peutic pregnancy terminations. The material was collected by
was incubated with glutathione–Sepharose 4B beads (Pharma-
cervical dilatation and curettage of the decidua and obtained
cia (Pfizer), West Ryde, New South Wales, Australia) for 30 min
from Dr Miriam Hanssens (Department of Obstetrics and Gynae-
at room temperature. The beads were washed, eluted with elution
cology, University of Leuven, Belgium). Gestation was in the
buffer (10 mm reduced glutathione, 50 mm TRIS-HCl, pH 8.0)
range of 12–14 weeks. Tissue blocks were fixed at room temper-
and analysed by PAGE.
ature in 5% formalin, paraffin embedded, sectioned and stained
For primary immunisations, 1 mL of Freund’s Complete
with hematoxylin and eosin. Sections were assessed by an expe-
Adjuvant and 1 mL (0.5–1 mg) of the affinity-purified fusion
rienced pathologist (M. Hansenns). Samples were confirmed to
protein were mixed to give an emulsion for injection into two
be placental bed by morphological criteria and to contain at
separate rabbits. For secondary and tertiary immunisations,
least one spiral artery either in the decidual or myometrial layer.
given between 4 and 6 weeks after the primary injection, 1 mL
Immunohistochemistry on representative sections is as described
of Freund’s Incomplete Adjuvant and 1 mL (0.5–1 mg) of the
below.
affinity-purified fusion protein were made into an emulsion for
injection as described above. Two weeks after the secondary and
Cell culture tertiary immunisations, a terminal bleed was carried out and the
Trophoblast cell lines JEG-3, JAR, and BeWo were obtained serum recovered. Western blot analysis was used to test serial
from the American Type Culture Collection (ATCC, Rockville, dilutions of the sera for reactivity with the HLX-pGEX3 fusion
MD, USA). HTR8/SVneo cells were obtained from Dr C. protein and GST protein alone.
Graham (Sunnybrook Health Science Centre, Canada). The Immunoaffinity purification was carried out by coupling
HTR-8/SVneo cell line used in this study was a long-term cell 8 mg of purified HLX-pGEX3 fusion protein to CNBr-activated
line that senesces after ∼30 passages. The SGHPL-4 cell line Sepharose 4B, or GST protein to Sepharose 4B beads, following
was a kind gift from Dr Claire Roberts (Department of Obstetrics the manufacturer’s instructions (Pharmacia). Immune serum was
and Gynaecology, University of Adelaide, South Australia, Aus- added to GST-Sepharose 4B beads and incubated overnight at
tralia). JEG-3, JAR, and BeWo cells were grown in RPMI-1640 4◦ C. The supernatant was added to HLX-pGEX3 fusion protein-
(Invitrogen, Mount Waverly, Victoria, Australia) culture medium Sepharose 4B beads. Beads were washed thoroughly with 50 mm
supplemented with 10–20% heat-inactivated fetal calf serum TRIS-HCl, pH 7.4. Elution was carried out 4 times with 0.2 m
(FCS), 100 U mL−1 penicillin and 100 µg mL−1 streptomycin glycine, 1 mm EGTA, pH 2.5, before storage. The eluants were
(Invitrogen). SGHPL-4 cells were grown in Ham’s F10 nutrient adjusted to 0.3 m TRIS-HCl, pH 8.7.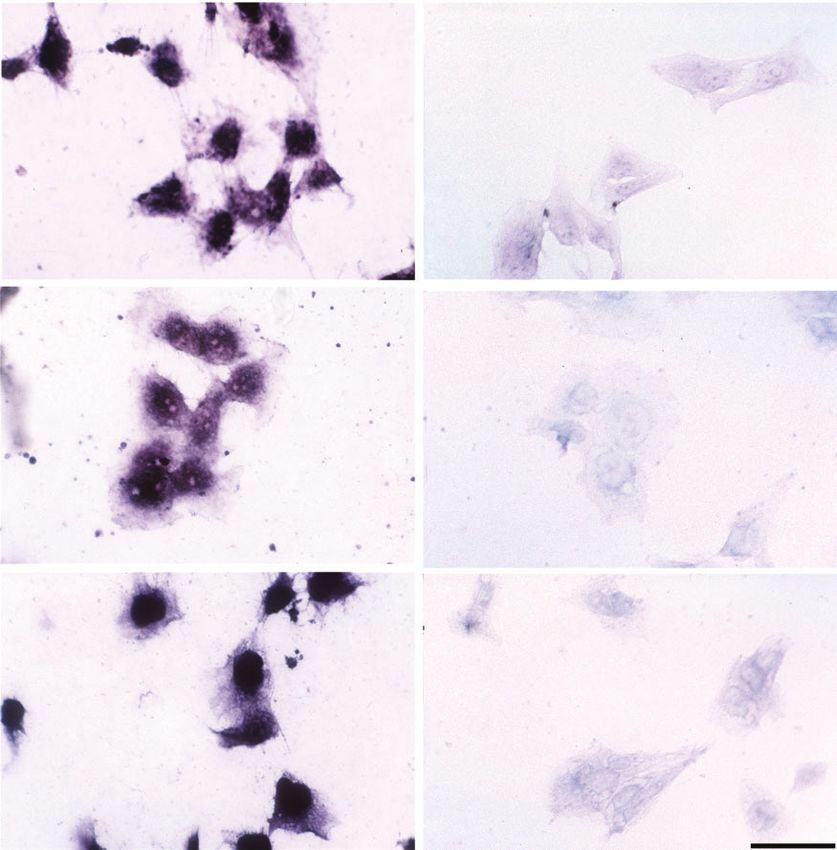
HLX expression in the placenta Reproduction, Fertility and Development 359
Western blot analysis of cell line and placental tissue Amplification (Perkin-Elmer) was then carried out following
homogenates the manufacturer’s instructions. All sections were incubated with
JEG3, JAR, BeWo and HTR8/SVneo cell lines were grown to alkaline phosphatase for 30 min at room temperature. For dual
near confluence, collected by centrifugation (300g for 5 min at staining, sections were treated with 3% H2 O2 /PBS and incubated
room temperature) and resuspended in 0.5 mL of 10 mm TRIS- with a monoclonal anti-cytokeratin 7 (CK-7, Dako, Carpinte-
HCl (pH 7.5) containing 1.5 mm EDTA, 10 nm sodium moly- ria, CA, USA) for 30 min at room temperature followed by
bdate, 1 mm DTT, 10% glycerol, 0.1 mm PMSF and a complete streptavidin-HRP (1 : 1000 dilution, 1 mg mL−1 , Silenus) for
protease inhibitor cocktail (Roche, Indianapolis, IN, USA). Cel- 30 min at room temperature. Chromogenic detection of alka-
lular protein was extracted and denatured by boiling for 5 min. line phosphatase was carried out with NBT and BCIP (Roche
Tissue homogenates of first-trimester placental tissues were pre- Molecular Biochemicals) or detection of streptavidin-HRP with
pared by homogenisation using an Ultra-Turex homogeniser with AEC (3-amino-9-ethylcarbazole kit, Zymed (Invitrogen), Carls-
subsequent addition of an equal amount homogenisation buffer bad, CA, USA). Sections were counterstained with 0.5% methyl
(0.5% CHAPS, 0.05% NaN3, 1 mm PMSF, 5 mm EDTA, 20 mm green and mounted with 80% glycerol. Control slides included
TRIS-HCl, pH 8.0). Insoluble debris from cellular and tissue pro- incubation with pre-immune serum as negative control and sec-
tein extracts were removed by centrifugation at 2500g for 15 min tions that were treated with HLX-GST fusion protein to show the
at room temperature. The supernatant was analysed immediately specificity of the HLX antibody.
for protein concentration using Pierce Protein Assay reagent
(Pierce (Quantum Scientific), Murarrie, Queensland, Australia) Results
using bovine serum albumin as standard.
Approximately 25 µg of each protein sample was elec- HLX antibody of tissue and cell culture homogenates
trophoresed on 10% PAGE/0.1% SDS gels and transferred Immunoblotting of trophoblast cell protein extracts and placental
electrophoretically to Immobilon P membrane (NEN (Perkin protein extracts with affinity purified polyclonal antibody HLX
Elmer), Waltham, MA, USA). The membrane was blocked with is shown in Fig. 1.There was an equal load of total protein (25 µg)
10% non-fat milk powder/TBST (0.14 m NaCl, 2.7 mm KCl, in each track. A predominant protein of 50 kDa was detected in
0.025 m TRIS-HCl, pH 7.4)/0.05% Tween-20 for 4 h. Incuba- first trimester placenta protein homogenate and a band of the
tion with the HLX immunoaffinity-purified antibody was carried same size was detected in the cell lines tested (lanes 3–5, 7–8).
out for 16 h at 4◦ C. The membrane was then washed in TBST The specificity of the antibody for the 50 kDa protein species
and incubated with biotinylated anti-rabbit antibody in imida- was demonstrated by loss of the signal when the HLX antibody
zole buffer (20 mm imidazole, 150 mm NaCl, 0.05% Thimerosal, was competed with the HLX-GST fusion protein, which was the
pH 7.0)/2% non-fat milk powder for 1 h at room temperature. peptide antigen used to raise the antibody (lane 6). The signal
Streptavidin-HRP in 2% non-fat milk powder/TBST buffer was was not lost when an unrelated fusion protein, DLX4-GST, was
added and incubation was carried out at room temperature for 1 h. used (data not shown).
Tyramide Signal Amplification (TSA, Perkin-Elmer) was then
carried out following the manufacturer’s instructions. Detection Cellular distribution of HLX in trophoblast cell lines
of alkaline phosphatase was carried out overnight with NBT The staining pattern for HLX to the trophoblast cell lines is
and BCIP (Roche Molecular Biochemicals). Competition exper- shown in Fig. 2a, c, e. The signal for HLX protein was predom-
iments were carried out by prior incubation of HLX antibody inantly nuclear but there was detectable weaker expression in
with an excess of HLX-GST fusion protein. the cytoplasm. The signal was uniform over the entire nucleus.
When the antibody was competed with HLX-GST fusion pro-
Immunohistochemistry tein, the signal was removed to reveal a low level of non-specific
For immunohistochemical analyses, fixed and paraffin- background staining (Fig. 2b, d, f ).
embedded tissues were cut into 5 µm thick sections and trans-
ferred onto Superfrost Plus microscope slides (Medos). Paraffin Cellular distribution of HLX during early pregnancy
sections were dewaxed by heating at 37◦ C for 1 h and fur- Dual label immunohistochemistry was used to show the stain-
ther treatment with xylene for 10 min. Dewaxed sections were ing pattern of HLX in first trimester villi (Fig. 3). Cytokeratin
rehydrated in graded ethanol and washed in distilled water. 7 staining (AEC, red) was detected in all trophoblast cell types
Slides were blocked for endogenous peroxidase activity using in the villi (villous cytotrophoblast, syncytiotrophoblast, syn-
1% H2 O2 /PBS. The sections were treated with proteinase K cytial sprouts, new villi). HLX protein expression (detected by
(50 µg mL−1 ) at 37◦ C for 10 min. NBT/BCIP, purple) was seen in villous cytotrophoblasts (VCT),
Fixed cells and paraffin tissue sections were blocked with but not in syncytiotrophoblast (S) or in syncytial sprouts (SP)
10% non-fat milk powder/TBST for 2 h. Immunoaffinity- (Fig. 3a). Figure 3c showed that HLX staining was present in
purified HLX antibody was added and incubated overnight at the VCT and in cytotrophoblast cells within a new villous (NV)
4◦ C. The sections were briefly washed in TBST and incu- branch. No HLX staining was observed in (SP) or in (S) that
bated with biotinylated anti-rabbit secondary antibody (1:200 stained with cytokeratin. Figure 3d shows a high magnification
dilution, 100 µg mL−1 , Silenus/Chemicon, North Ryde, New view, where perinuclear staining of HLX on VCT nuclei can
South Wales, Australia) for 30 min at room temperature fol- be seen. Figure 3e shows a junction between two villi (J) and
lowed by incubation with streptavidin-HRP. Tyramide Signal perinuclear HLX staining in the cytotrophoblast cells within the360 Reproduction, Fertility and Development G. Rajaraman et al.
1 2 3 4 5 6 7 8
50 kDa
Fig. 1. Western immunoblot analysis for HLX protein expression in early pregnancy placenta and trophoblast cell lines. Representative immunoblot
for HLX protein expression in the early pregnancy placenta and in the trophoblast homogenates is shown. As described in the methods section, 25 µg total
protein was loaded onto each well of 10% SDS–PAGE, immunoblotting performed using rabbit polyclonal HLX. A 50 kDa band corresponding to endogenous
HLX protein is marked with an arrow. Lane 1 – protein standards; lane 2 – early pregnancy placenta; lane 3 – JEG-3 choriocarcinoma cell line; lane 4 –
JAR choriocarcinoma cell line; lane 5 – BeWo choriocarcinoma cell line; lane 6 – negative control (competition of the HLX antibody with HLX-GST fusion
protein before Western analysis of the placental homogenate); lane 7 – SGHPL-4 human extravillous trophoblast cell line; lane 8 – HTR-8/SVneo human
extravillous trophoblast cell line.
junction and in VCT. Figure 3f is an adjacent section to that HLX expression in early pregnancy placental bed biopsies
shown in Fig. 3e and was stained with cytokeratin only and shows HLX signals in the placental bed biopsies were consistently
staining of both the VCT and S. weaker than those seen in the placenta and increased proteinase K
Therefore, HLX expression was not found in the syncy- treatment permeabilisation times were needed to detect the sig-
tiotrophoblast layer or the syncytial nuclei comprising syncytial nals. HLX protein was detected in endovascular trophoblast that
sprouts that were attached to the villi or free in the intervil- colonise the vessel walls in transformed spiral arteries (Fig. 6a,
lous space. In contrast to the uniform nuclear staining pattern c) and in endovascular trophoblast cells that plug the lumen of
on cultured cells (Fig. 2a, c, e, g, i), nuclear staining of the the spiral artery (Fig. 6a). Interstitial trophoblast, that develop
cytotrophoblast cells was punctate and often appeared perinu- from invasive extravillous cytotrophoblast derived from the dis-
clear (Fig. 3d, e). HLX protein was also detected in the nuclei of tal region of the cell column, were also positive for cytoplasmic
cytotrophoblast cells that penetrate syncytial sprouts in the pro- HLX expression (Fig. 6c, e). Panel B, D and F show correspond-
cess of forming a new villous branch (Fig. 3c). The specificity of ing cytokeratin staining of adjacent sections to confirm that HLX
the HLX antibody is demonstrated in Fig. 3b, where the signal signals were derived from trophoblast cells. There was a corre-
was removed by competing the HLX antibody with HLX-GST spondence in signals with those cells expressing HLX (compare
fusion protein. Fig. 6c, d and Fig. 6e, f ). It was evident that some cells in the
A variety of different fixation conditions were used but all endometrial stroma stain for HLX but are not positive for cyto-
generated punctate nuclear and/or perinuclear staining (data not keratin 7 (compare Fig. 6c, d). Trophoblast giant cells did not
shown). The punctate–perinuclear staining was not due to TSA show significant reactivity with HLX (data not shown). In all
amplification, as the same pattern appeared when TSA ampli- of the above cell types in the placental bed biopsies, the expres-
fication was omitted (see Fig. 4a). Similar protein localisation sion of HLX was cytoplasmic and did not show the characteristic
patterns were obtained on frozen sections (see Fig. 5a). punctate–perinuclear staining seen in the villous cytotrophoblast
Without TSA amplification, HLX protein was detected only or extravillous cytotrophoblast in the proximal regions of the cell
in the villous cytotrophoblast cells (Fig. 4a). With amplifica- columns.
tion, HLX expression was detected more strongly in the villous
cytotrophoblast in a punctate–perinuclear pattern. In addition,
lower levels of HLX expression were detected in endothelial cells Discussion
that comprise the fetal capillary vessel walls and in perivascular We have evaluated the HLX protein expression pattern in cul-
cells around the vessel wall (Fig. 4b, c), by staining. HLX was tured trophoblast cells, in early pregnancy placental tissues
not detected strongly in the fibroblasts or macrophages of the and placental bed biopsies. In general, we have shown that
villous stroma. trophoblasts that express HLX go on to differentiate into syn-
HLX expression was detected in the extravillous cytotro- cytiotrophoblasts that do not express HLX. A rabbit polyclonal
phoblast cells that are found at the tips of anchoring villi. antibody to the human HLX protein was raised. Western analy-
Both frozen sections (Fig. 5a) and paraformaldehyde fixed sec- sis revealed the affinity-purified antibody raised against HLX
tions showed punctate–perinuclear HLX staining in the proximal detected a discrete 45 kDa molecular mass species in early preg-
regions of the cell columns (Fig. 5a–d). Weak HLX expres- nancy placental homogenates and in the cultured cells. This
sion could be detected in the distal regions of the columns and size is similar to that predicted by conceptual translation of the
appeared to be cytoplasmic expression rather than nuclear (see HLX cDNA sequence (50 kDa). The HLX molecular weight also
also Fig. 6). matches that determined in a related study on HLX expression inHLX expression in the placenta Reproduction, Fertility and Development 361
(a) (b)
(c) (d )
(e) (f )
Fig. 2. Immunocytochemical detection of HLX expression in trophoblast cell lines. Immunocytochemistry for HLX expression
was performed in confluent cultures of HTR8/SVneo, JEG-3 and JAR, BeWo and SGHPL-4, as described in the methods section.
(a) HLX on HTR8/SVneo; (b) competition experiment for HLX on HTR8/SVneo; (c) HLX on JEG-3; (d) competition experiment
for HLX on JEG-3; (e) HLX on JAR; ( f ) competition experiment for HLX on JAR. Magnification is 400×, scale bar is 10 µm.
the late pregnancy placenta (Murthi et al. 2006) and a study on stained uniformly in cell cultures. Expression of HLX in cell
HLX expression in malignant and benign human colorectal tis- lines representative of extravillous trophoblast is consistent with
sues (Hollington et al. 2004). Prior binding of the HLX antibody HLX detection in the extravillous cytotrophoblast cell columns
with the HLX-GST fusion protein substantially decreased the (Fig. 5).
immunoreactivity, whereas the use of an unrelated homeodomain In early pregnancy placental sections, HLX was detected
fusion protein (DLX4-GST ) did not decrease the immunoreac- in the villous cytotrophoblast, and extravillous cytotrophoblast
tivity (data not shown). These data verify the specificity of the cells that form the cytotrophoblast cell columns, which is con-
antibody for HLX protein. sistent with our previous studies of early human pregnancy
The transformed human trophoblast cell line HTR8/SVneo placental mRNA expression patterns (Quinn et al. 1997b). The
and human choriocarcinoma cell lines (JEG-3 and JAR) showed expression pattern of HLX in human placental cell types is
predominant expression of HLX in the nucleus. Nuclei were summarised in Fig. 7.362 Reproduction, Fertility and Development G. Rajaraman et al.
(a) (b)
VCT
SP
S
(c) (d )
S
NV S
VCT
SP VCT
SP NV
(e) (f )
S
VCT
J
J
S
VCT
Fig. 3. Immunohistochemical detection of HLX in the early pregnancy placenta. Immunohistochemical localisation of HLX
protein was performed on chorionic villi to show various trophoblast-derived structures, using NBT/BCIP detection to give a dark
purple stain and staining for cytokeratin, which was detected by AEC red stain as described in the methods section (Panels (a), (c)–( f )).
Panel (b) shows a competition experiment with the HLX-GST fusion protein and the section was counterstained with methyl green.
The section in Panel (b) was not stained for cytokeratin. Sections contain villous cytotrophoblasts (VCT), syncytiotrophoblast (S),
syncytial sprouts (SP), new branches of the chorionic villi (NV) and junctions between villi (J). The scale bar is 50 µm in each panel.
In the early pregnancy tissues, HLX was not detected in syn- 1997b, 1998, 2000), HLX shows no expression in terminally
cytial nuclei from the outer layer of the villi or in syncytial differentiated syncytiotrophoblast cells.
buds and sprouts. These data are consistent with the recently- We also detected HLX expression at a lower level in the
published pattern of HLX expression in the term placenta where, microvascular endothelial cells that line the fetal capillaries,
again, no expression was detected in the syncytiotrophoblast in perivascular cells and in scattered stromal cells. Endothelial
(Murthi et al. 2006). Unlike other placental homeobox genes cell expression was consistent with our recent studies showing
we have investigated in the villous compartment of the pla- that HLX is expressed in these cell types in the term placen-
centa i.e. DLX4, MSX2, MOX2 (also called GAX ) (Quinn et al. tae (Murthi et al. 2006), in enriched primary microvascularHLX expression in the placenta Reproduction, Fertility and Development 363
(a) (b) (c)
Fig. 4. HLX staining in first trimester placental tissue sections without TSA amplification. Panel (a) shows HLX antibody staining without TSA
amplification. Punctate–perinuclear signal was detected in villous cytotrophoblast (arrow) but not in the syncytiotrophoblast nuclei or the syncytial cytoplasm
(arrowhead). Panel (b) is a higher magnification view of HLX staining following TSA amplification. The colour reaction development time was extended
to allow detection of the weaker HLX staining in endothelial and perivascular cells. The section was counterstained with methyl green to show the nuclei.
Panel (b) shows perinuclear staining of HLX in villous cytotrophoblast nuclei (arrow) but not in syncytiotrophoblast nuclei (arrowhead). Panel (c) is a high
magnification view around the vessel seen in Panel (b). A methyl green-stained syncytial nucleus is shown (arrow). Endothelial staining (concave arrows) is
shown around the vessel. Perinuclear cells showing HLX staining are shown by the concave arrowhead. The scale bars in Panels (a) and (b) are 50 µm and in
Panel (c) the bar is 10 µm.
(a) (b)
(c) (d )
Fig. 5. HLX staining of extravillous trophoblast cells. Panels (a) and (b) show low magnification images of frozen and paraformaldehyde-fixed sections
stained with HLX antibody. HLX staining was detected in the villous cytotrophoblast (arrow) and in the proximal region of the extravillous cytotrophoblast
column extending from the tip of some villi (arrowhead). Panel (c) is a higher magnification view of a villus showing staining of HLX ; signal was detected
in the same cell types as Panel (b). Panel (d) shows a high magnification view of HLX staining in the proximal regions of the extravillous cytotrophoblast
cell column (arrowhead). Staining of HLX was perinuclear–punctate in this region and arrows show representative nuclei. The weaker cytoplasmic staining
of HLX in the distal regions of the column cannot be seen without further permeabilisation of the cells (see Fig. 6). The scale bars in (a)–(c) are 50 mm and
in (d) the bar is 10 µm.364 Reproduction, Fertility and Development G. Rajaraman et al.
(a) (b)
(c) (d )
(e) (f )
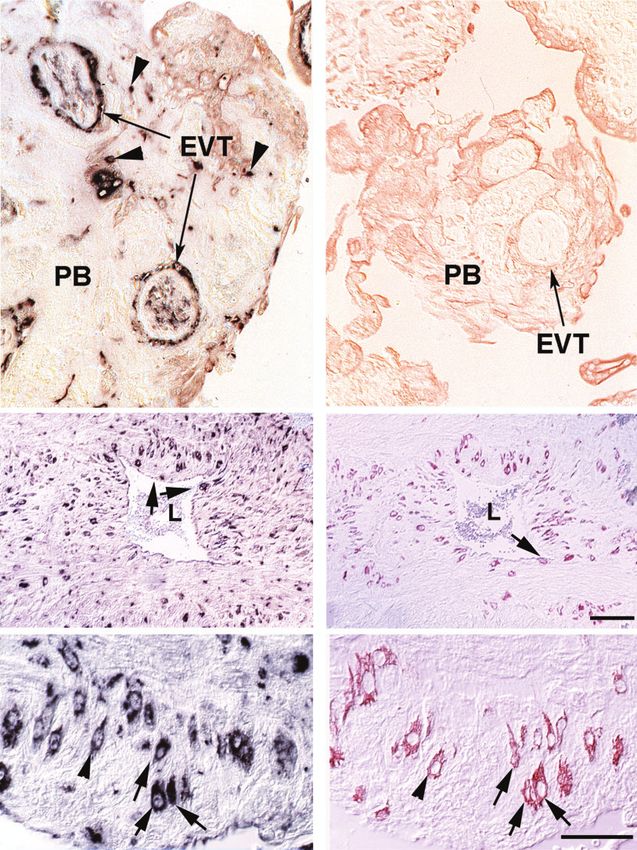
Fig. 6. Immunohistochemical detection of HLX in early pregnancy placental bed biopsies. Panel (a) shows the distribution
of HLX in the placental bed (PB). HLX is detected in endovascular trophoblast (EVT) that line transformed spiral artery walls and
in endovascular cells that plug the artery (concave arrowheads). Arrowheads show scattered interstitial trophoblast cells. Panel (b)
is a cytokeratin 7 staining of an adjacent section showing that cells of trophoblast origin are present in the vessel walls. Panel (c)
shows HLX distribution surrounding a partially-transformed spiral artery. HLX is detected in a few endovascular trophoblast cells
that have invaded the vessel wall (arrows). Interstitial trophoblast cells expressing HLX are scattered around the vessel. Panel (d) is
a cytokeratin staining of the section adjacent to that shown in Panel (c). In many positions, the cytokeratin staining corresponds with
the HLX staining. Panel (e) is a higher magnification of (c) showing predominantly uniform cytoplasmic staining of HLX. Panel ( f )
is the adjacent section stained with cytokeratin. Matching cells in (e) and ( f ) are shown with arrowheads and arrows. The scale bars
for (c)–( f ) are 50 µm.HLX expression in the placenta Reproduction, Fertility and Development 365
-VCT
syn-
Proliferation HLX expression
potential
High Nuclear
-CC
-ITC
-ENV
Low Cytoplasmic
-GC
Fig. 7. Summary of HLX expression in the early pregnancy human placenta. HLX expression changes
from nuclear in the villous cytotrophoblast cells (VCT) and the highly proliferative extravillous trophoblasts
in the proximal region of the cell column (CC), to cytoplasmic expression in the distal region of the
cell column, in the interstitial trophoblasts (ITC) and in the endovascular trophoblasts (ENV), where the
proliferation potential is low. HLX is not expressed in the giant cells (GC).
endothelial cells and in endothelial cell lines (Murthi et al. HLX showed punctate nuclear–perinuclear localisation in
2007). proliferating extravillous trophoblast nuclei in the proximal
The subcellular localisation of HLX in placental villous cells regions of the cell columns in early pregnancy tissue. The proxi-
was predominantly nuclear, but nuclei did not stain uniformly. mal regions of cell columns contain highly proliferative extravil-
Both punctate and perinuclear staining was observed. The reason lous trophoblast cells that are detected by specific markers such
for this variation is not known and it appears to be tissue- and cell as Ki67 (Genbacev et al. 2000). Expression of HLX in these pro-
type-dependent since the same antibody showed uniform nuclear liferative cells is consistent with studies in other cell types where
staining in cultured cell lines (Fig. 2) and in colorectal tissues HLX acts to control the proliferation of haematopoietic cells
(Hollington et al. 2004). The same punctate nuclear–perinuclear (Deguchi et al. 1992; Deguchi and Kehrl 1993). One of the pro-
pattern of HLX expression in the placental villi was found in posed direct or indirect targets of HLX in haematopoietic cells is
fixed and frozen tissues, with a variety of signal detection meth- the important cell cycle regulator p34cdc2 (cdk1) (Deguchi et al.
ods and with various fixation and permeabilisation methods. 1992). The protein expression pattern of CDK1 in first trimester366 Reproduction, Fertility and Development G. Rajaraman et al.
and term placenta (Genbacev et al. 2000) closely parallels that development and we propose that reduced levels of HLX are
shown for HLX in the present study, suggesting that CDK1 may required for cytotrophoblast differentiation.
also be a target for HLX regulation in cytotrophoblast cells.
In placental bed biopsies, only cytoplasmic HLX expres- Acknowledgements
sion was detected in interstitial trophoblast and endovascular
The authors wish to gratefully acknowledge the patients and the clinical
trophoblasts. This change in staining from the nucleus to the
and nursing staff of the hospitals participating in this study for the sup-
cytoplasm is seen in other cell types in reproductive tissues. ply of samples. We thank Dr Miriam Hanssens for supplying placental bed
Homeobox gene HOXA7 also shows a change in expression biopsy samples for this study. The Early Career Researcher Scheme, Uni-
from nuclear to cytoplasmic as cells make the transition from versity of Melbourne, supported P. Murthi. A grant from Equity Trustees on
proliferative to differentiated cells during ovarian folliculoge- behalf of the Lynne Quayle Charitable Trust supported the work. The Uni-
nesis (Ota et al. 2006). Differential subcellular localisation of versity of Melbourne, Felix Myer postgraduate scholarship award, supported
homeobox gene products between the nucleus and cytoplasm is G. Rajaraman.
also seen in carcinomas (D’Elia et al. 2002). We note similari-
ties to several known nuclear localisation signals (NLS) found References
in homeobox genes. The RRMKWRH sequence of HLX has a 5 Auman, H. J., Nottoli, T., Lakiza, O., Winger, Q., Donaldson, S., and
out of 7 amino acid match to the RRMKWKK NLS of the PDX- Williams, T. (2002). Transcription factor AP-2gamma is essential in
1 homeodomain (Moede et al. 1999) and the RRMKWKR NLS the extra-embryonic lineages for early postimplantation development.
of the HB9 homeodomain (Kosaka et al. 2004). Furthermore, Development 129, 2733–2747.
a putative NLS RKRSWSRAVFSNLQRKGLEK (consensus Cokol, M., Nair, R., and Rost, B. (2000). Finding nuclear localization signals.
sequence (RK)[3,5]×[11,18](RK)K×[2,3]K) was found by the EMBO Rep. 1, 411–415. doi:10.1093/EMBO-REPORTS/KVD092
PredictNLS software program at sequence position 274 in the Cross, J. C. (2003). The genetics of pre-eclampsia: a feto-placental
or maternal problem? Clin. Genet. 64, 96–103. doi:10.1034/J.1399-
region N-terminal to helix 3 (Quinn et al. 1997a; Cokol et al.
0004.2003.00127.X
2000). Finally, there is sequence similarity to the leucine-rich
Cross, J. C., Baczyk, D., Dobric, N., Hemberger, M., Hughes, M.,
nuclear export sequence found in the engrailed homeodomain Simmons, D. G., Yamamoto, H., and Kingdom, J. C. (2003). Genes,
protein (Maizel et al. 1999). Thus, HLX shows features consis- development and evolution of the placenta. Placenta 24, 123–130.
tent with a protein that is regulated in its entry into the nucleus doi:10.1053/PLAC.2002.0887
and may be important in controlling trophoblast proliferation. Deguchi, Y., and Kehrl, J. H. (1993). High level expression of the homeobox
The expression pattern of HLX is consistent with a role in gene HB24 in a human T-cell line confers the ability to form tumors in
proliferation since HLX is not expressed in terminally differ- nude mice. Cancer Res. 53, 373–377.
entiated trophoblast cell types (syncytiotrophoblast, placental Deguchi, Y., Kirschenbaum, A., and Kehrl, J. H. (1992). A diverged homeo-
bed giant cells). HLX has been shown to regulate cell prolifera- box gene is involved in the proliferation and lineage commitment
of human hematopoietic progenitors and highly expressed in acute
tion in haematopoietic cell lineages (Deguchi and Kehrl 1993).
myelogenous leukemia. Blood 79, 2841–2848.
Using siRNA-based gene inactivation methods we have recently
D’Elia, A. V., Tell, G., Russo, D., Arturi, F., Puglisi, F., et al. (2002). Expres-
shown that decreasing HLX levels in cultured trophoblast cell sion and localization of the homeodomain-containing protein HEX
line results in substantially reduced cell proliferation (Rajaraman in human thyroid tumors. J. Clin. Endocrinol. Metab. 87, 1376–1383.
et al. 2007). doi:10.1210/JC.87.3.1376
HLX may also act as a downregulator of cytotrophoblast dif- Genbacev, O., McMaster, M. T., and Fisher, S. J. (2000). A repertoire
ferentiation or fusion to form multinucleate trophoblast in both of cell cycle regulators whose expression is coordinated with human
the villi and the placental bed. In this respect the role of HLX cytotrophoblast differentiation. Am. J. Pathol. 157, 1337–1351.
in the trophoblast lineage may be similar to that in haematopoi- Hemberger, M., and Cross, J. C. (2001). Genes governing placental devel-
etic progenitor cells, where downregulation of HLX expression opment. Trends Endocrinol. Metab. 12, 162–168. doi:10.1016/S1043-
2760(01)00375-7
appears to be required for differentiation (Deguchi et al. 1992).
Hentsch, B., Lyons, I., Li, R., Hartley, L., Lints, T. J., Adams, J. M., and
Abnormal homeobox gene expression can result in aberrant
Harvey, R. P. (1996). Hlx homeobox gene is essential for an inductive
cellular differentiation and is often associated with a transformed tissue interaction that drives expansion of embryonic liver and gut. Genes
phenotype (Kongsuwan et al. 1989; Kamps et al. 1990, 1991; Dev. 10, 70–79. doi:10.1101/GAD.10.1.70
Perkins et al. 1990; Lu et al. 1991). Ectopic expression of Hollington, P., Neufing, P., Kalionis, B., Waring, P., Bentel, J., Wattchow, D.,
HLX in nude mice resulted in the formation of tumours, sug- and Tilley, W. D. (2004). Expression and localization of homeodomain
gesting that HLX plays a role in cell differentiation (Deguchi proteins DLX4, HB9 and HB24 in malignant and benign human
and Kehrl 1993). Evidence for a role for HLX in cell differ- colorectal tissues. Anticancer Res. 24, 955–962.
entiation was also provided in collaborative studies (Hollington Kamps, M. P., Murre, C., Sun, X. H., and Baltimore, D. (1990). A new
et al. 2004) where nuclear epithelial immunoreactivity of HLX homeobox gene contributes the DNA binding domain of the t(1;19)
translocation protein in pre-BALL. Cell 60, 547–555. doi:10.1016/0092-
decreased in moderately-differentiated tumours compared with
8674(90)90658-2
the corresponding matched non-malignant mucosa. Therefore,
Kamps, M. P., Look, A. T., and Baltimore, D. (1991). The human t(1;19)
disregulation of HLX may result in aberrant cytotrophoblast translocation in pre-B ALL produces multiple nuclear E2A-Pbx1 fusion
proliferation and differentiation and contribute to placental proteins with differing transforming potentials. Genes Dev. 5, 358–368.
pathologies such as choriocarcinoma. doi:10.1101/GAD.5.3.358
In conclusion, the HLX homeobox gene is expressed primar- Kongsuwan, K., Allen, J., and Adams, J. M. (1989). Expression of
ily in proliferating cytotrophoblast cell types in early placental Hox-2.4 homeobox gene directed by proviral insertion in a myeloidHLX expression in the placenta Reproduction, Fertility and Development 367
leukemia. Nucleic Acids Res. 17, 1881–1892. doi:10.1093/NAR/ Perkins, A., Kongsuwan, K., Visvader, J., Adams, J. M., and Cory, S. (1990).
17.5.1881 Homeobox gene expression plus autocrine growth factor production
Kosaka, Y., Akimoto, Y., Yokozawa, K., Obinata, A., and Hirano, H. (2004). elicits myeloid leukemia. Proc. Natl. Acad. Sci. USA 87, 8398–8402.
Localization of HB9 homeodomain protein and characterization of its doi:10.1073/PNAS.87.21.8398
nuclear localization signal during chick embryonic skin development. Quinn, L. M., Johnson, B. V., Nicholl, J., Sutherland, G. R., and Kalionis, B.
Histochem. Cell Biol. 122, 237–247. doi:10.1007/S00418-004-0698-5 (1997a). Isolation and identification of homeobox genes from the human
Krumlauf, R. (1994). Hox genes in vertebrate development. Cell 78, 191– placenta including a novel member of the Distal-less family, DLX4. Gene
201. doi:10.1016/0092-8674(94)90290-9 187, 55–61. doi:10.1016/S0378-1119(96)00706-8
Li, Y., and Behringer, R. R. (1998). Esx1 is an X-chromosome-imprinted Quinn, L. M., Latham, S. E., and Kalionis, B. (1997b). Homeobox gene
regulator of placental development and fetal growth. Nat. Genet. 20, HB24, a regulator of haematopoiesis, is a candidate for regulating differ-
309–311. doi:10.1038/3129 entiation of the extra-embryonic trophoblast cell lineage. Reprod. Fertil.
Lu, M., Gong, Z. Y., Shen, W. F., and Ho, A. D. (1991). The tcl-3 proto- Dev. 9, 617–623. doi:10.1071/R97025
oncogene altered by chromosomal translocation in T-cell leukemia codes Quinn, L. M., Kilpatrick, L. M., Latham, S. E., and Kalionis, B. (1998).
for a homeobox protein. EMBO J. 10, 2905–2910. Homeobox genes DLX4 and HB24 are expressed in regions of epithelial-
Maizel, A., Bensaude, O., Prochiantz, A., and Joliot, A. (1999). A short mesenchymal cell interaction in the adult human endometrium. Mol.
region of its homeodomain is necessary for engrailed nuclear export and Hum. Reprod. 4, 497–501. doi:10.1093/MOLEHR/4.5.497
secretion. Development 126, 3183–3190. Quinn, L. M., Latham, S. E., and Kalionis, B. (2000). The homeobox genes
Moede, T., Leibiger, B., Pour, H. G., Berggren, P., and Leibiger, I. B. MSX2 and MOX2 are candidates for regulating epithelial-mesenchymal
(1999). Identification of a nuclear localization signal, RRMKWKK, in cell interactions in the human placenta. Placenta 21(Suppl.), S50–S54.
the homeodomain transcription factor PDX-1. FEBS Lett. 461, 229–234. doi:10.1053/PLAC.1999.0514
doi:10.1016/S0014-5793(99)01446-5 Rajaraman, G., Murthi, P., Leo, B., Brennecke, S. P., and Kalionis, B. (2007).
Morasso, M. I., Grinberg, A., Robinson, G., Sargent, T. D., and Mahon, K. A. Homeobox gene HLX1 is a regulator of colony stimulating factor 1-
(1999). Placental failure in mice lacking the homeobox gene Dlx3. Proc. dependent trophoblast cell proliferation. Placenta, in press.
Natl. Acad. Sci. USA 96, 162–167. doi:10.1073/PNAS.96.1.162 Roberson, M. S., Meermann, S., Morasso, M. I., Mulvaney-Musa, J. M.,
Murthi, P., Doherty, V., Said, J., Donath, S., Brennecke, S. P., and Kalionis, B. and Zhang, T. (2001). A role for the homeobox protein Distal-less 3
(2006). Homeobox gene HLX1 expression is decreased in idio- in the activation of the glycoprotein hormone alpha subunit gene in
pathic human fetal growth restriction. Am. J. Pathol. 168, 511–518. choriocarcinoma cells. J. Biol. Chem. 276, 10016–10024. doi:10.1074/
doi:10.2353/AJPATH.2006.050637 JBC.M007481200
Murthi, P., So, M., Gude, N. M., Doherty, V. L., Brennecke, S. P., and Rossant, J., and Cross, J. C. (2001). Placental development: lessons from
Kalionis, B. (2007). Homeobox genes are differentially expressed in mouse mutants. Nat. Rev. Genet. 2, 538–548. doi:10.1038/35080570
macrovascular human umbilical vein endothelial cells and microvas- Sapin, V., Blanchon, L., Serre, A. F., Lemery, D., Dastugue, B., and Ward, S. J.
cular placental endothelial cells. Placenta 28, 219–223. doi:10.1016/ (2001). Use of transgenic mice model for understanding the placen-
J.PLACENTA.2006.02.012 tation: towards clinical applications in human obstetrical pathologies?
Oettinger, H. F., Pasqualini, R., and Bernfield, M. (1992). Recombinant pep- Transgenic Res. 10, 377–398. doi:10.1023/A:1012085713898
tides as immunogens: a comparison of protocols for antisera production Westerman, B. A., Poutsma, A., Maruyama, K., Schrijnemakers, H. F., van
using the pGEX system. Biotechniques 12, 544–549. Wijk, I. J., and Oudejans, C. B. (2002). The proneural genes NEUROD1
Ota, T., Choi, K. B., Gilks, B., Leung, P., and Auersperg, N. (2006). Cell and NEUROD2 are expressed during human trophoblast invasion. Mech.
type- and stage-specific changes in HOXA7 protein expression in human Dev. 113, 85–90. doi:10.1016/S0925-4773(01)00665-7
ovarian folliculogenesis: possible role of GDF-9. Differentiation 74, 1. Westerman, B. A., Murre, C., and Oudejans, C. B. (2003). The cellular Pax-
doi:10.1111/J.1432-0436.2006.00053.X Hox-helix connection. Biochim. Biophys. Acta 1629, 1–7.
Peng, L., and Payne, A. H. (2002). AP-2 gamma and the homeodomain
protein distal-less 3 are required for placental-specific expression of the
murine 3 beta-hydroxysteroid dehydrogenase VI gene, Hsd3b6. J. Biol.
Chem. 277, 7945–7954. doi:10.1074/JBC.M106765200 Manuscript received 14 September 2007, accepted 16 December 2007
http://www.publish.csiro.au/journals/rfdYou can also read

















































