Sol Sherry Lecture in Thrombosis - Molecular Events in Acute Inflammation
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Sol Sherry Lecture in Thrombosis
Molecular Events in Acute Inflammation
Stephen M. Prescott, Thomas M. McIntyre, Guy A. Zimmerman, Diana M. Stafforini
Abstract—The inflammatory response is characterized by a multistep molecular interaction between “signaling” cells, such
as endothelial cells, and “responding” cells, such as neutrophils and monocytes. In the first step, selectins produced by
signaling cells mediate the tethering of responding cells at sites of inflammation. Subsequently, an additional mediator
expressed by signaling cells activates the tethered responding cells. Under pathological conditions, the same mechanism
is invoked in inappropriate ways: (1) by prolonged presentation of selectins on the cell surface and (2) by the unregulated
production of oxidized phospholipids that mimic the normal secondary signaling molecule, platelet-activating factor
(PAF). The enzyme PAF acetylhydrolase (PAF-AH) inactivates PAF and oxidized phospholipids and constitutes an
“off” switch that suppresses inflammation. Inhibition of normal PAF-AH function or inactivating mutations of the
PAF-AH gene can lead to increased susceptibility to inflammatory disease. These studies have relevance to
atherosclerosis and thrombosis, because inflammation is a central feature of both. (Arterioscler Thromb Vasc Biol.
2002;22:727-733.)
Key Words: inflammation 䡲 selectins 䡲 platelet-activating factor 䡲 chemokines 䡲 oxidized phospholipids
T his article is a summary of the Sol Sherry lecture of the
Council on Arteriosclerosis, Thrombosis, and Vascular
Biology, which was presented at the 73rd Scientific Sessions
these cases, the signaling cell communicates with the re-
sponding target cell in a multistep fashion. First, it tethers the
target cell at the site of inflammation, and then it sends a
of the American Heart Association, New Orleans, La, No- signal to it through a molecule recognized by a receptor on
vember 12–15, 2000. It highlights work from our laboratory the target cell, often in a juxtacrine fashion. The target cell
addressing the molecular basis of inflammation, particularly responds by eliciting new cellular behaviors, including, in
early events at the vascular wall. For this lecture, we have some cases, altered gene transcription.
focused on our own work and have not attempted to provide In the case of the immediate inflammatory response be-
a general review of this topic. tween PMNs and endothelial cells, the initial adhesion is
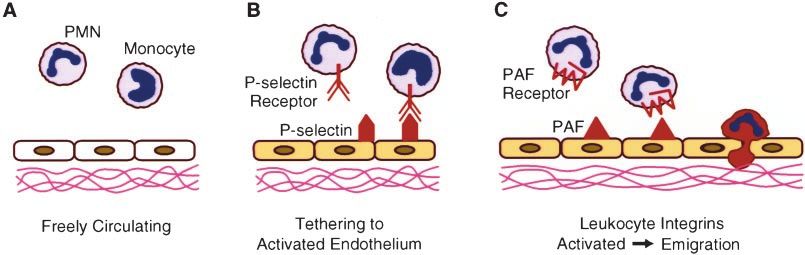
In response to infection and other tissue damage, leuko- mediated by P-selectin. On appropriate stimulation by ago-
cytes emigrate from the bloodstream to the tissues. The nists, P-selectin stored within endothelial cells is rapidly
adhesion of leukocytes to endothelial cells is a crucial first translocated to the cell surface. As a result, the surface of
step in this process. When appropriately regulated, the in- endothelial cells is altered from a nonadhesive and nonthrom-
flammatory response is physiological and homeostatic. How- bogenic character to a proadhesive surface. P-selectin binds
ever, when the stimulus is spatially, temporally, or quantita- to counterreceptors, such as P-selectin glycoprotein ligand-1
tively inappropriate, adhesion and activation of leukocytes (PSGL-1) on leukocytes, tethering leukocytes to the activated
can have pathological effects (Figure 1). We have focused on endothelial cells. Quantitative analyses show that P-selectin is
the molecular changes responsible for these early steps. We only transiently expressed on the surface of activated endo-
have described a mechanism by which cells react to inflam- thelial cells2 and that the bulk is reinternalized with 30
matory signals to permit a spatially specific response and a minutes of agonist exposure.3
high-fidelity reaction (Figure 2). Several different types of A subsequent step consists of activation of the tethered
circulating cells use this same molecular mechanism. leukocytes by an additional signal from the activated endo-
“Signaling” cell/“responding” cell pairs of a number of thelial cells. This is accomplished by the production of
different types are involved in inflammatory events, eg, platelet-activating factor (PAF) by the endothelial cell and its
endothelial cells and polymorphonuclear neutrophils (PMNs), expression on the cell surface. Leukocytes express a receptor
endothelial cells and monocytes, and platelets and mono- for PAF that potently induces a program of adhesion and
cytes. We and others (see review1) have shown that in all of -integrin–mediated migration by the leukocyte. PAF also
Received November 27, 2001; revision accepted March 11, 2002.
From the Huntsman Cancer Institute (S.M.P., D.M.S.), Program in Human Molecular Biology and Genetics (T.M.M., G.A.Z.), and the Departments
of Internal Medicine (S.M.P., T.M.M., G.A.Z., D.M.S.), Oncological Sciences (S.M.P.), and Pathology (T.M.M.), University of Utah, Salt Lake City.
Correspondence to Stephen M. Prescott, MD, Huntsman Cancer Institute, 2000 Circle of Hope, Salt Lake City, UT 84112. E-mail
steve.prescott@hci.utah.edu
© 2002 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol. is available at http://www.atvbaha.org DOI: 10.1161/01.ATV.0000016153.47693.B2
727
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015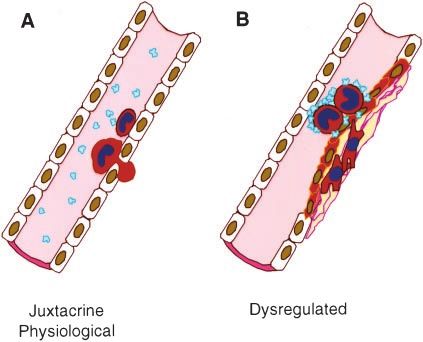
728 Arterioscler Thromb Vasc Biol. May 2002
account for the observed temporal differences between im-
mediate and delayed inflammatory processes.
Several features differentiate delayed from immediate re-
sponses. In more delayed adhesive responses, activated en-
dothelial cells use E-selectin as a tether, and the secondary
activating molecule is a chemokine, such as interleukin-8
(IL-8). In this case, E-selectin is not previously stored in
unstimulated endothelial cells. Instead, agonists such as
tumor necrosis factor (TNF) transcriptionally activate the
E-selectin gene in a highly regulated manner, a process that
results in E-selectin accumulation within a few hours. Surface
expression of E-selectin reaches a maximum within 12 hours
and typically returns to basal levels by 24 hours. The second
signaling molecule in the stepwise process is also synthesized
in response to inflammatory stimuli. Thus, the expressions of
Figure 1. Physiological (A) and dysregulated (B) inflammation tethering and activating molecules occur in parallel. The time
are mediated by leukocyte adhesion to the vascular wall in
response to a variety of stimuli. required for the expression and accumulation of each signal-
ing protein affects the temporal readout of delayed inflam-
primes leukocytes for agonists that they will encounter after matory responses. An additional molecular difference is that
the leukocytes migrate out of the vasculature. PAF is cell-associated, whereas IL-8 is released from acti-
The multistep process has several advantages. First, the vated endothelial cells as a soluble molecule. This could
fact that the initiating signal is expressed on the surface of the theoretically result in widespread inflammation. However,
endothelial cell means that the signal is fixed in space. One surface receptors expressed by endothelial cells specifically
implication of this model is that the local concentration of the bind IL-8, which serves to localize the response to discrete
activating mediator could be very high, despite a low sys- areas of the vasculature.
temic concentration. As a result, leukocytes are recruited
precisely where they are needed. Second, the initial “tethering Monocyte Cytokine Secretion Is Regulated by
without activating” step permits an editing function by the P-Selectin Tethering and PAF Signaling
PMN. Binding to P-selectin brings the PMN into proximity Cell-cell interactions at vascular interfaces may establish
with the endothelial cell, permitting the PMN to scan for an levels or patterns of immediate-early gene expression. In fact,
activating signal. If such a signal is present, the PMN is it is known that contact between monocytes and stimulated
activated and expresses integrins to augment the adhesive endothelial cells is a critical control point in inflammation,
response. If no activator is present, the leukocyte is released particularly in atherosclerosis. In our studies of cell-cell
and reenters the circulation. Tethering is the molecular basis interactions that influence signaling by cytokines, we showed
for the observation that leukocytes roll on the endothelium in that monocytes adherent to either purified P-selectin or to
vivo. P-selectin–transfected Chinese hamster ovary cells secrete
Various types of molecules participate as mediators of cytokines, such as monocyte chemotactic protein (MCP)-1
immediate versus delayed (ie, requiring ⬎1 hour to become and TNF-␣, in response to stimulation by PAF.
evident) inflammatory responses. However, the general prin- To explore the mechanism by which adhesion to P-selectin
ciple of the response is similar. In both cases, endothelial influences cytokine gene expression in monocytes, we fo-
cells are stimulated to express members of the selectin family cused on the effect of adhesion on nuclear factor-B (NF-
of adhesion molecules that then bind constitutively expressed B), a transcription factor.4 NF-B is a key regulator of
counterreceptors on circulating leukocytes. The specific mol- immediate-early genes in monocytes5 and is required for
ecules and the manner in which each species is expressed MCP-16 and TNF-␣ expression. We found that the adhesion
Figure 2. P-selectin and PAF expressed by endothelial cells provide a physiological mechanism for temporally and spatially regulated
leukocyte recruitment.
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015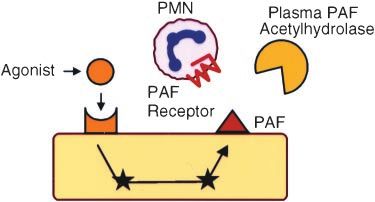
Prescott et al Molecular Events in Acute Inflammation 729
of monocytes to purified immobilized P-selectin caused a To establish whether the binding of P-selectin to PSGL-1 is
small but consistent translocation of NF-B to the nucleus. sufficient for activated platelets to induce chemokine secre-
This nuclear translocation was enhanced dramatically when tion by monocytes, we used a similar system, ie, endothelial
the cells were stimulated with PAF. This suggests that cells and PMNs. These experiments showed that platelets
facilitated transfer of NF-B may be a mechanism by which must express an additional component for platelet-monocyte
adhesion to P-selectin amplifies MCP-1 and TNF-␣ secretion signaling to take place. The activating molecule was identi-
by stimulated monocytes. Thus, monocyte adhesion to fied as RANTES, a C-C chemokine originally detected in
P-selectin presented by stimulated endothelial cells or by activated T lymphocytes that is released by activated platelets
platelets can potentially influence cytokine generation early and regulates MCP-1 secretion by monocytes. RANTES is
in inflammatory processes. Furthermore, the concerted ex- stored and then secreted from platelets stimulated with
pression of PAF can dramatically augment this response. thrombin; a cell surface receptor for RANTES is found on
monocytic cells. Thus, signaling that arises from platelet-
Molecules Required for Activated Platelets to monocyte interactions is similar to that following endothelial
Adhere to and Signal Human Monocytes cell–neutrophil adhesion in that both require expression of
To understand the inflammatory responses associated with P-selectin by the signaling cell. In neither case, however, is
the simultaneous recruitment of platelets and leukocytes, we this sufficient. Expression of a second molecule, PAF or
identified the functional alterations induced when monocytes RANTES, for endothelial cells and platelets, respectively, is
bind to the surface of activated platelets. The activation of necessary for the completion of the signal.
platelets by thrombin, PAF, oxidized phospholipids (OxPLs), Studies of the role of RANTES as a platelet-derived signal
and other agonists leads to the adhesion of monocytes to the for monocytes have shown that this chemokine is present in
platelets, intercellular signaling, altered gene expression, and substantial amounts in the supernatants from activated plate-
synthesis of mediators. In turn, monocytes engage adhesion lets and that MCP-1 release in platelet-monocyte suspensions
molecules that tether these cells and that induce intracellular is inhibited by an antibody against RANTES. Because the
signals delivered through surface receptors. antibody did not block the formation of platelet-monocyte
When they are activated in vivo, endothelial cells and aggregates, RANTES-induced signaling apparently occurs
platelets translocate P-selectin to the surface from intracellu- after the binding event. However, induction of MCP-1 in-
lar storage granules. Monocytes adhere to both cell types.7–9 volved the expression of P-selectin and also of RANTES,
Endothelial cells express P-selectin transiently under most inasmuch as monocytes cultured on albumin rather than
physiological conditions. In contrast, platelets express P-selectin showed no induction of MCP-1 secretion when
P-selectin in a relatively stable manner and can sustain treated with RANTES.
platelet-monocyte contact for hours. We asked whether the MCP-1 was not the only monocyte chemokine induced by
binding of monocyte PSGL-1 to P-selectin presented on exposure to activated platelets. IL-8, a C-X-C chemokine that
activated human platelets affects nuclear signaling or chemo- is a potent neutrophil chemoattractant, was released by
kine generation by the leukocytes.10 This issue is important monocytes incubated with activated platelets but not by
because monocytes and platelets adhere to one another in monocytes cultured alone or with unstimulated platelets. We
numerous inflammatory and thrombotic disorders.11–14 As found that IL-8 secretion was induced when monocytes that
previously shown, when monocytes were mixed with had adhered to purified immobilized P-selectin were stimu-
thrombin-activated platelets, adhesion between the 2 types of lated with RANTES but was not induced on control surfaces
cells resulted. In addition, activated platelets formed “ro- coated with albumin or fibronectin. Thus, the formation of
settes” around several monocytes. To determine whether the platelet-monocyte aggregates results in the production of a
adherent platelets induced chemokine release by the mono- variety of potent chemokine effectors.
cytes, we coincubated the 2 cell types and monitored release
of the chemokine MCP-1. Platelets activated by thrombin Pathological Activation
induced significant release of MCP-1 compared with control Oxidants and free radicals are deleterious in many ways, and
levels. Chemokine secretion was dependent on P-selectin organisms use numerous approaches to block their production
expression, inasmuch as neutralization of P-selectin with the and limit their damage. Many oxidative chemical reactions
use of a specific blocking antibody significantly attenuated are not constrained by enzymes and may show exponential
MCP-1 secretion and inhibited rosette formation in platelet- reaction rates. Some products of oxidative attacks, including
monocyte mixtures. those of lipid oxidation, are highly reactive species that
Complementary experiments have shown that chemokine modify proteins and DNA or activate components of the
secretion is dependent on the presence of PSGL-1. P-selectin immune and inflammatory systems. Such changes are thought
preferentially binds to PSGL-1, a sialomucin that is consti- to be important in the pathogenesis of atherosclerosis and
tutively present on monocytes and other myeloid leuko- other diseases.
cytes.15,16 We used a monoclonal antibody that recognizes We have studied the characteristics and actions of OxPLs
PSGL-1 and blocks its binding to P-selectin, and we found that are involved in inflammatory stimulation. To approach
that blocking this interaction inhibits platelet-monocyte ro- this question, we subjected human endothelial cells to oxida-
sette grouping and MCP-1 secretion by monocytes. Thus, the tive stress by exposing them to low concentrations (100 to
expression of P-selectin and also of PSGL-1 is required for 200 mol/L) of oxidants, such as H 2 O 2 and tert-
these responses to occur. butylhydroperoxide. We found that PMNs adhered to treated
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015730 Arterioscler Thromb Vasc Biol. May 2002
endothelial cells and that this adhesion was blocked entirely polyunsaturated fatty acids, yielding a large series of phos-
by antibodies to P-selectin.17 We observed that the time phatidylcholines with shortened sn-2 residues. Some of these
course of P-selectin expression by cells subjected to oxidative oxidation products, termed “PAF-like” phospholipids, are
stress was different from that by cells treated with physio- able to activate cells expressing the PAF receptor,19,23,24
logical stimuli. P-selectin remained on the surface of oxidized including monocytes,25 leukocytes,19 and platelets.23 Because
endothelial cells for hours compared with its transient appear- oxidation of phosphatidylcholines generates a plethora of
ance on cells treated with thrombin or histamine. This lack of chemically related molecules and because only some of these
regulation of P-selectin expression by endothelial cells ex- stimulate the PAF receptor, identification of the specific
posed to oxidants is likely to have important implications in biologically active species has been complicated. We have
pathological inflammation. found that oxidation of very rare phosphatidylcholine species
In addition to our findings in P-selectin expression, we in LDL generates all of the bioactive PAF receptor agonists
observed a dramatic alteration of the morphology of endo- derived from LDL oxidation.20,26
thelial cells exposed to oxidants. A time-dependent vesicula- In summary, we have found that oxidant-exposed endothe-
tion, or “blebbing,” of the plasma membrane occurred, and lium sheds vesicles that contain potent activators of neutro-
many vesicles were shed into the supernatant.18 In contrast to phil function. The active compounds are phospholipids that
buffer-treated controls, supernatants from oxidant-treated en- have been oxidatively fragmented to yield structural ana-
dothelial cells caused PMNs to adhere to gelatin-coated logues of PAF. In contrast to PAF synthesis, which is
plates, a response that required integrin activation. The carefully controlled, the formation of potent mimetics after
vesicles contained little, if any, P-selectin, as determined by oxidation of synthetic phosphatidylcholines,19,23 isolated
immunoassay. LDLs,27–29 and foodstuffs is unregulated. Therefore, these
We explored further the identity of the oxidant-induced oxidative reactions have the potential to produce high con-
activating agent. We had found previously that synthetic centrations of potent inflammatory agents. Endothelial cells
phosphatidylcholine is fragmented by oxidation, generating transiently synthesize PAF in response to physiological ago-
analogues of PAF.19 In related experiments, we demonstrated nists but do not release it. In contrast, oxidant-treated endo-
that oxidatively fragmented phosphatidylcholines activated thelial cells release PAF-like phospholipids, which may
neutrophils, a response that was inhibited by specific antag- spread tissue damage from the immediate area to other
onists of the PAF receptor. We hypothesized that the expo- organs. The production of oxidatively fragmented phospho-
sure of endothelial cells to strong oxidants might generate lipids is not regulated by the usual constraints on synthesis.
oxidatively fragmented phospholipids with PAF-like biolog- Thus, degradation represents a crucial mechanism necessary
ical activity.18 In subsequent experiments, we showed this to to suppress the actions of these toxic products. Indirect
be the case and identified the active component as a phos- evidence supporting the notion that PAF-like lipids have
pholipid that caused adhesion of neutrophils by functional pathological effects includes the fact that a deficiency of
upregulation of CD11/CD18 integrins.18 Competitive antag- PAF-AH is correlated with an increased risk of stroke30 and
onists of the PAF receptor completely blocked neutrophil increased levels of circulating PAF-like lipids.31
adhesion induced by these OxPLs. We examined the possi-
bility that the vesicles contained authentic PAF but, in an In Vivo Tests of OxPLs as Inflammatory Agents
extensive set of experiments, found little or none. Because the oxidative reactions that create PAF-like lipids
To physically characterize the bioactive phospholipid, we were all performed in a laboratory setting, it was not known
separated vesicular lipids by high-performance liquid chro- whether sufficiently oxidizing conditions exist in vivo to
matography.20 Fractions were assayed for their ability to create the inflammatory PAF mimetics that we had studied. It
stimulate neutrophil adhesion. The stimulatory material that was also not known whether such mediators could accumu-
we identified did not comigrate with authentic PAF and was late to effective levels in the face of plasma antioxidants and
identical to fractions that we had previously shown to contain endogenous defense mechanisms, such as the constitutively
the free radical– catalyzed oxidation products of 1-palmitoyl- active PAF-AH. On the basis of these considerations, we and
2-arachidonoyl-sn-glycero-3-phosphocholine. Like PAF, the others32–35 sought to determine whether cigarette smoke could
phospholipids we identified were hydrolyzed by PAF acetyl- elicit the accumulation of PAF-like lipids in an intact organ-
hydrolase (PAF-AH), a phospholipase A2 that attacks only ism and whether these compounds constitute the circulating
phospholipids with fragmented and/or oxidized residues at mediators responsible for inflammatory changes observed
the sn-2 position.21 We found that exposure of the active after cigarette smoke exposure. If this were the case, simple
fractions to PAF-AH completely destroyed their ability to antioxidant protection might suppress the accumulation of
activate neutrophils. toxic oxidation products, thereby suppressing the proinflam-
PAF is recognized by a single receptor that is a member of matory effects of smoking.
a family of 7-transmembrane–spanning G-protein–linked re- To address these issues, we first isolated polar lipids from
ceptors.22 The PAF receptor recognizes an intact phospho- the plasma of hamsters exposed to the smoke of 1 reference
lipid, a feature that is characteristic of the PAF receptor cigarette and tested the ability of these lipids to promote
exclusively. The PAF receptor has 2 additional important neutrophil adhesion. These experiments showed that lipids
recognition requirements: (1) for a specific head group and from treated, but not control, plasma exerted a marked
(2) for a specific atypical sn-2 residue. Oxidation of alkyl adhesion-promoting effect.25 The stimulatory material was
phosphatidylcholine results in the fragmentation of sn-2 eluted in fractionation experiments with the same profile as
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015Prescott et al Molecular Events in Acute Inflammation 731
OxPLs with PAF-like activity. The biological activity of the
smoke-induced inflammatory agonists was equivalent to 10⫺7
mol/L PAF per milliliter plasma. The effect of these lipids
could be blocked with a specific competitive antagonist to the
PAF receptor. Thus, in vivo stimulation of OxPL production
by exposure to cigarette smoke was effective in inducing
inflammatory cell adhesion.
To further understand the mechanism of action of PAF-like
lipids, we visualized leukocyte interaction with the microvas-
cular endothelium in response to cigarette smoke. We found Figure 3. The biological action of PAF (and PAF-like lipids) is
that smoke-induced leukocyte adhesion to arteriolar and controlled in a variety of ways, including tightly regulated syn-
venular endothelium was significantly inhibited by pretreat- thetic pathways for PAF, localization of receptors on specific
target cells, juxtacrine activation of target cells by membrane-
ing the animals with a highly selective PAF receptor antag- associated PAF (spatial regulation of signaling), and degradation
onist or by blocking the PAF receptor before smoke exposure. of PAF and PAF-like lipids by cellular and plasma PAF-AHs.
In addition to leukocyte– endothelial cell interactions, we
observed rapid formation of circulating leukocyte-platelet pathways for PAF synthesis, regulation of the presentation of
aggregates in hamsters exposed to cigarette smoke. Experi- PAF by juxtacrine signaling, and efficient degradative en-
ments showed that PAF-like lipids extracted from the plasma
zymes, the PAF-AHs. There are several forms of PAF-AH
of smoke-exposed hamsters were responsible for this aggre-
activities, both intracellular and secreted. These enzymes are
gate formation. A characteristic feature of monocyte-platelet
constitutively active and are calcium independent (Figure 3).
aggregates that is not shown by individual monocytes4 is their
The plasma form of PAF-AH plays a key role as a
enhanced ability to synthesize and secrete inflammatory
scavenger of PAF and OxPLs. We have characterized the
cytokines that attract and activate monocytes (C-C family, eg,
MCP-1) and neutrophils (C-X-C family, eg, IL-8). We found human plasma PAF-AH and cloned its cDNA.37 The enzyme
that in the presence of platelets, PAF-like lipids induce is inactive against long-chain phospholipids, a property that
substantial synthesis of MCP-1 and IL-8 by monocyte-plate- explains its ability to circulate in active form without acting
let aggregates compared with controls.25 These data suggest on lipids that constitute blood cell membranes and lipopro-
that PAF-like lipids amplify the inflammatory response, teins. In addition to PAF, a second group of compounds
inasmuch as they induce the synthesis of inflammatory hydrolyzed by PAF-AH, the oxidatively fragmented phos-
cytokines by circulating platelet-monocyte aggregates. pholipids,18,28 also have short and/or oxidized acyl groups at
We next sought to determine whether antioxidants prevent the sn-2 position of glycerol.
the accumulation of PAF-like lipids that mediate the inflam- In contrast to the synthesis of PAF, which is highly
matory reaction induced by cigarette smoke. We fed the regulated,38 OxPLs are produced in an uncontrolled manner.
antioxidant vitamin C to hamsters and then extracted and The extent to which products of phospholipid oxidation
purified PAF-like lipids from the animals after exposure to accumulate depends heavily on the rate at which they are
smoke. We measured the amount of stimulatory activity in catabolized. Inasmuch as PAF-AHs are maximally active in
these extracts relative to maximally effective levels of PAF. the basal state and do not require calcium for activity, they
We found that exposure to cigarette smoke results in the provide an immediate defense mechanism against toxic ef-
accumulation of significant levels of circulating PAF-like fects mediated by fragmented phospholipids.
lipids and that vitamin C pretreatment reduces the amount of PAF provokes inflammation in part by its ability to activate
these lipids to background levels. PMNs, resulting in changes required for the directional
The work mentioned above demonstrated for the first time migration of cells.39 We found that preincubation of PAF with
that PAF-like lipids are formed and accumulate in vivo. recombinant PAF-AH caused PAF to lose its ability to
Characterization of these modified phospholipids shows that activate cell polarization and cell spreading in vitro. PAF is
the biological activity falls within an order of magnitude of
also a potent stimulus for increased permeability of the
that of PAF. Thus, the results from our in vitro and in vivo
vasculature, a prominent component of severe inflammation.
approaches have demonstrated that PAF-like lipids generated
Using the well-characterized paw edema model, we found
in the plasma of hamsters exposed to cigarette smoke act as
that pretreatment of the footpads with recombinant PAF-AH
the primary mediators of subsequent microvascular events.
blocked PAF-induced edema by ⬎85%.37 Similarly, intrave-
Regulation of PAF-Mediated Effects nous pretreatment with recombinant PAF-AH blocked PAF-
PAF mediates a wide range of immune and allergic reac- induced footpad edema by ⬎80%. We also tested PAF-AH in
tions.36 If the mechanism for inactivation of PAF is impaired, a pleurisy model characterized by vascular leakage into the
effector cells may be recruited to sites of injury for inappro- pleural space in response to PAF. Rats were pretreated with
priately long times, resulting in prolonged inflammation. either recombinant PAF-AH or control buffer, and then PAF
There are several powerful mechanisms that, individually or was injected into the pleural space. One hour later, the degree
together, can control the biological activities of PAF under of vascular leakage was determined. Compared with control
physiological conditions. These include regulation of the animals, the rats that received PAF-AH had a ⬎80% reduc-
expression and activity of the receptor, tightly controlled tion of vascular leakage. Collectively, the in vivo and in vitro
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015732 Arterioscler Thromb Vasc Biol. May 2002
results demonstrate that PAF-AH abolishes the inflammatory 3. Subramaniam M, Koedam JA, Wagner DD. Divergent fates of P- and
effects of PAF on leukocytes and the vasculature. E-selectins after their expression on the plasma membrane. Mol Cell Biol.
1993;4:791– 801.
Many studies have examined the possibility that deficiency 4. Weyrich AS, McIntyre TM, McEver RP, Prescott SM, Zimmerman GA.
of the plasma form of PAF-AH is associated with inflamma- Monocyte tethering by P-selectin regulates monocyte chemotactic
tory disease. Inasmuch as the secreted enzyme can control the protein-1 and tumor necrosis factor-alpha secretion: signal integration and
NF-kappaB translocation. J Clin Invest. 1995;95:2297–2303.
magnitude and duration of PAF-mediated signaling, a de- 5. Juliano RL, Haskill S. Signal transduction from the extracellular matrix.
creased ability to degrade PAF would be predicted to result in J Cell Biol. 1993;120:577–585.
pathological responses. Studies from other laboratories have 6. Ueda AK, Okuda S, Ohno A, Shirasi T, Iagarashi K, Matsunaga J,
Fukushima S, Kawamoto Y, Ishigatsubo Y, Okubo T. NF-kb and Sp1
shown that the activity of plasma PAF-AH is decreased in
regulate transcription of the human monocyte chemoattractant protein-1
patients with asthma,40 systemic lupus erythematosus,41 and gene. J Immunol. 1994;153:2052–2063.
septic shock.42 Our results have shown that the administration 7. Larsen E, Celi A, Gilbert GE, Furie BC, Erban JK, Bonfanti R, Wagner
of supplemental exogenous PAF-AH suppresses inflamma- DD, Furie B. PADGEM protein: a receptor that mediates the interaction
of activated platelets with neutrophils and monocytes. Cell. 1989;59:
tion in vivo. Thus, in diseases in which there is a deficiency 305–312.
of enzyme activity, PAF or related compounds may accumu- 8. McEver RP. Selectins. Curr Opin Immunol. 1994;6:75– 84.
late, and supplemental PAF-AH may reverse the pathological 9. Elstad MR, McIntyre TM, Prescott SM, Zimmerman GA. The interaction
of leukocytes with platelets in blood coagulation. Curr Opin Hematol.
response. 1995;2:47–54.
It has been known since 1988 that ⬇4% of the Japanese 10. Weyrich AS, Elstad MR, McEver RP, McIntyre TM, Moore KL, Morrissey
population is deficient in plasma PAF-AH activity.40 The vast JH, Prescott SM, Zimmerman GA. Activated platelets signal chemokine
synthesis by human monocytes. J Clin Invest. 1996;97:1525–1534.
majority of these subjects carry a PAF-AH mutation that
11. Munro JM, Cotran RS. Biology of disease: the pathogenesis of athero-
results in the substitution of Val279 with Phe.43 This muta- sclerosis: atherogenesis and inflammation. Lab Invest. 1988;58:249 –261.
tion, when present as a homozygous trait, results in undetect- 12. Lehr HA, Olofsson AM, Carew TE, Vajkoczy P, von Andrian UH,
able levels of PAF-AH activity in the plasma. Several groups Hubner C, Berndt MC, Steinberg D, Messmer K, Arfors KE. P-selectin
mediates the interaction of circulating leukocytes with platelets and
have found that this mutation increases susceptibility to microvascular endothelium in response to oxidized lipoprotein in vivo.
inflammatory and allergic diseases,44,45 demonstrating that Lab Invest. 1994;71:380 –386.
PAF-AH functions as an anti-inflammatory safety net. 13. Lehr HA, Frei B, Arfors KE. Vitamin C prevents cigarette smoke-induced
leukocyte aggregation and adhesion to endothelium in vivo. Proc Natl
Acad Sci U S A. 1994;91:7688 –7692.
Conclusions 14. Palabrica T, Lobb R, Furie BC, Aronovitz M, Benjamin C, Hsu Y-M,
Sajer SA, Furie B. Leukocyte accumulation promoting fibrin deposition is
A multistep molecular interaction occurs when cells are
mediated in vivo by P-selectin on adherent platelets. Nature. 1992;359:
activated by inflammatory agents. Activated cells tether 848 – 851.
responding cells through the expression of selectins. Activa- 15. Sako D, Chang X-J, Barone KM, Vachino G, White HM, Shaw G,
tion of responding cells occurs by means of a second type of Veldman GM, Bean KM, Ahem TJ, Furie B, et al. Expression cloning of
a functional glycoprotein ligand for P-selectin. Cell. 1993;75:1179 –1186.
molecule expressed by the signaling cell. In endothelial 16. Moore KL, Patel KD, Bruehl RE, Fugang L, Johnson DA, Lichenstein
cell–PMN interactions, P-selectin is the tether, and the lipid HS, Cummings RD, Bainton DF, McEver RP. P-selectin glycoprotein
PAF is the activating molecule. Under pathological condi- ligand-1 mediates rolling of human neutrophils on P-selectin. J Cell Biol.
1995;128:661– 671.
tions, this same stepwise mechanism is invoked in inappro- 17. Patel KD, Zimmerman GA, Prescott SM, McEver RP, McIntyre TM.
priate ways, first by prolonged expression of P-selectin and Oxygen radicals induce human endothelial cells to express GMP-140 and
then by the unregulated production of OxPLs that mimic the bind neutrophils. J Cell Biol. 1991;112:749 –759.
18. Patel KD, Zimmerman GA, Prescott SM, McIntyre TM. Novel leukocyte
actions of PAF. PAF-AH acts as an off signal for PAF and
agonists are released by endothelial cells exposed to peroxide. J Biol
also for OxPL because it inactivates these lipids, thereby Chem. 1992;267:15168 –15175.
suppressing inflammation. Inactivation of PAF-AH by ge- 19. Smiley PL, Stremler KE, Prescott SM, Zimmerman GA, McIntyre TM.
netic or environmental causes leads to an increased suscep- Oxidatively fragmented phosphatidylcholines activate human neutrophils
through the receptor for platelet-activating factor. J Biol Chem. 1991;
tibility to inflammatory disease. These studies have important 266:11104 –11110.
implications in the pathogenesis of disease with components 20. Marathe GK, Davies SS, Harrison KA, Silva AR, Murphy RC, Castro-
of inflammation, such as atherosclerosis and other vascular Faria-Neto H, Prescott SM, Zimmerman GA, McIntyre TM. Inflammatory
PAF-like lipids in oxidized low density lipoproteins are fragmented alkyl
syndromes. phosphatidylcholines. J Biol Chem. 1999;274:28395–28404.
21. Stafforini DM, Prescott SM, McIntyre TM. Human plasma platelet-
activating factor acetylhydrolase: purification and properties. J Biol
Acknowledgments Chem. 1987;262:4223– 4230.
National Institutes of Health grants HL-35828, HL-44525, HL- 22. Shimizu T, Honda Z, Nakamura M, Bito H, Izumi T. Platelet-activating
44513, and HL-50153 supported this work. We thank Ellen T. factor receptor and signal transduction. Biochem Pharmacol. 1992;44:
Wilson, PhD, for preparation of the manuscript and Diana Lim for 1001–1008.
preparation of the figures. 23. Tanaka T, Iimori M, Tsukatani H, Tokumura A. Platelet-aggregating
effects of platelet-activating factor-like phospholipids formed by oxi-
dation of phosphatidylcholines containing an sn-2-polyunsaturated fatty
References acyl group. Biochim Biophys Acta. 1994;1210:202–208.
1. Zimmerman GA, McIntyre TM, Prescott SM. Adhesion and signaling in 24. Kern H, Volk T, Knauer-Schiefer S, Mieth T, Rustow B, Kox WJ,
vascular cell-cell interactions. J Clin Invest. 1996;98:1699 –1702. Schlame M. Stimulation of monocytes and platelets by short-chain phos-
2. Hattori R, Hamilton KK, Fugate RD, McEver RP, Sims PJ. Stimulated phatidylcholines with and without terminal carboxyl group. Biochim
secretion of endothelial von Willebrand factor is accompanied by rapid Biophys Acta. 1998;1394:33– 42.
redistribution to the cell surface of the intracellular granule membrane 25. Lehr H-A, Weyrich AS, Saetzler RK, Jurek A, Arfors KE, Zimmerman
protein GMP-140. J Biol Chem. 1989;264:7768 –7771. GA, Prescott SM, McIntyre TM. Vitamin C blocks inflammatory plate-
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015Prescott et al Molecular Events in Acute Inflammation 733
let-activating factor mimetics created by cigarette smoking. J Clin Invest. 36. Prescott SM, Zimmerman GA, Stafforini DM, McIntyre TM. Platelet-
1997;99:2358 –2364. activating factor and related lipid mediators. Annu Rev Biochem. 2000;
26. Tsoukatos DC, Arborati M, Liapikos T, Clay KL, Murphy RC, Chapman 69:419 – 445.
MJ, Ninio E. Copper-catalyzed oxidation mediates PAF formation in 37. Tjoelker LW, Wilder C, Eberhardt C, Stafforini DM, Dietsch G, Schimpf
human LDL subspecies: protective role of PAF:acetylhydrolase in dense B, Hooper S, Le Trong H, Cousens LS, Zimmerman GA, et al. Anti-
LDL. Arterioscler Thromb Vasc Biol. 1997;17:3505–3512. inflammatory properties of a platelet-activating factor acetylhydrolase.
27. Tokumura A, Toujima M, Yoshioka Y, Fukuzawa K. Lipid peroxidation Nature. 1995;374:549 –553.
in low density lipoproteins from human plasma and egg yolk promotes 38. Zimmerman GA, Prescott SM, McIntyre TM. Platelet-activating factor: a
accumulation of 1-acyl analogues of platelet-activating factor-like lipids. fluid-phase and cell-mediator of inflammation. In: Gallin JI, Goldstein
Lipids. 1996;31:1251–1258. IM, eds. Inflammation: Basic Principles and Clinical Correlates. 2nd ed.
28. Heery JM, Kozak M, Stafforini DM, Jones DA, Zimmerman GA, New York, NY: Raven Press; 1992:149 –176.
McIntyre TM, Prescott SM. Oxidatively modified LDL contains phos- 39. Lorant DE, Topham MK, Whatley RE, McEver RP, McIntyre TM,
pholipids with PAF-like activity and stimulates the growth of smooth Prescott SM, Zimmerman GA. Inflammatory roles of P-selectin. J Clin
muscle cells. J Clin Invest. 1995;96:2322–2330. Invest. 1993;92:559 –570.
29. Frostegard J, Huang YH, Ronnelid J, Schafer-Elinder L. Platelet-
40. Miwa M, Miyake T, Yamanaka T, Sugatani J, Suzuki Y, Sakata S, Araki
activating factor and oxidized LDL induce immune activation by a
Y, Matsumoto M. Characterization of serum platelet-activating factor
common mechanism. Arterioscler Thromb Vasc Biol. 1997;17:963–968.
(PAF) acetylhydrolase: correlation between deficiency of serum PAF
30. Hiramoto M, Yoshida H, Imaizumi T, Yoshimizu N, Satoh K. A mutation
acetylhydrolase and respiratory symptoms in asthmatic children. J Clin
in plasma platelet-activating factor acetylhydrolase (Val2793 Phe) is a
Invest. 1988;82:1983–1991.
genetic risk factor for stroke. Stroke. 1997;28:2417–2420.
31. Satoh K, Imaizumi T, Yoshida H, Hiramoto M, Takamatsu S. Increased 41. Tetta C, Bussolino F, Modena V, Montrucchio G, Segoloni G, Pescarmona
levels of blood platelet-activating factor (PAF) and PAF-like lipids in G, Camussi G. Release of platelet-activating factor in systemic lupus erythe-
patients with ischemic stroke. Acta Neurol Scand. 1992;85:122–127. matosus. Int Arch Allergy Appl Immunol. 1990;91:244–256.
32. Harats D, Ben-Naim M, Dabach Y, Hollander G, Stein O, Stein Y. Cigarette 42. Graham RM, Stephens CJ, Silvester W, Leong LLL, Strurm MJ, Taylor
smoking renders LDL susceptible to peroxidative modification and enhanced RR. Plasma degradation of platelet-activating factor in severely ill
metabolism by macrophages. Atherosclerosis. 1989;79:245–252. patients with clinical sepsis. Crit Care Med. 1994;22:204 –212.
33. Lehr HA, Frei B, Arfors KE. Vitamin C prevents cigarette smoke-induced 43. Stafforini DM, Satoh K, Atkinson DL, Tjoelker LW, Eberhardt C,
leukocyte aggregation and adhesion to endothelium in vivo. Proc Natl Yoshida H, Imaizumi T, Takamatsu S, Zimmerman GA, McIntyre TM,
Acad Sci U S A. 1994;91:7688 –7692. Gray PW, Prescott SM. Platelet-activating factor acetylhydrolase defi-
34. Lehr HA, Kress E, Menger MD, Friedl HP, Hubner C, Arfors KE, Messmer ciency. J Clin Invest. 1996;97:2784 –2791.
K. Cigarette smoke elicits leukocyte adhesion to endothelium in hamsters: 44. Tjoelker LW, Stafforini DM. Platelet-activating factor acetylhydrolases
inhibition by CuZn-SOD. Free Radic Biol Med. 1993;14:573–581. in health and disease. Biochim Biophys Acta. 2000;1488:102–123.
35. Lehr HA. Adhesion-promoting effects of cigarette smoke on leukocytes 45. Stafforini DM. PAF acetylhydrolase gene polymorphisms and asthma
and endothelial cells. Ann N Y Acad Sci. 1993;686:112–119. severity. Pharmacogenomics. 2001;2:163–175.
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015Sol Sherry Lecture in Thrombosis: Molecular Events in Acute Inflammation
Stephen M. Prescott, Thomas M. McIntyre, Guy A. Zimmerman and Diana M. Stafforini
Arterioscler Thromb Vasc Biol. 2002;22:727-733; originally published online March 21, 2002;
doi: 10.1161/01.ATV.0000016153.47693.B2
Arteriosclerosis, Thrombosis, and Vascular Biology is published by the American Heart Association, 7272
Greenville Avenue, Dallas, TX 75231
Copyright © 2002 American Heart Association, Inc. All rights reserved.
Print ISSN: 1079-5642. Online ISSN: 1524-4636
The online version of this article, along with updated information and services, is located on the
World Wide Web at:
http://atvb.ahajournals.org/content/22/5/727
Permissions: Requests for permissions to reproduce figures, tables, or portions of articles originally published
in Arteriosclerosis, Thrombosis, and Vascular Biology can be obtained via RightsLink, a service of the
Copyright Clearance Center, not the Editorial Office. Once the online version of the published article for which
permission is being requested is located, click Request Permissions in the middle column of the Web page
under Services. Further information about this process is available in thePermissions and Rights Question and
Answer document.
Reprints: Information about reprints can be found online at:
http://www.lww.com/reprints
Subscriptions: Information about subscribing to Arteriosclerosis, Thrombosis, and Vascular Biology is online
at:
http://atvb.ahajournals.org//subscriptions/
Downloaded from http://atvb.ahajournals.org/ by guest on March 17, 2015You can also read

















































