Early Regulation of Axolotl Limb Regeneration
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
THE ANATOMICAL RECORD 295:1566–1574 (2012)
Early Regulation of Axolotl Limb
Regeneration
AKI MAKANAE1 AND AKIRA SATOH1,2*
1
Okayama University, Research Core for Interdisciplinary Sciences (RCIS),
3-1-1, Tsushima-naka, Kita-ku, Okayama 700-8530, Japan
2
Japan Science Promotion Agency (JST), PRESTO, 4-1-8 Honcho Kawguchi,
Saitama, Japan
ABSTRACT
Amphibian limb regeneration has been studied for a long time. In
amphibian limb regeneration, an undifferentiated blastema is formed
around the region damaged by amputation. The induction process of blas-
tema formation has remained largely unknown because it is difficult to
study the induction of limb regeneration. The recently developed acces-
sory limb model (ALM) allows the investigation of limb induction and
reveals early events of amphibian limb regeneration. The interaction
between nerves and wound epidermis/epithelium is an important aspect
of limb regeneration. During early limb regeneration, neurotrophic factors
act on wound epithelium, leading to development of a functional epider-
mis/epithelium called the apical epithelial cap (AEC). AEC and nerves
create a specific environment that inhibits wound healing and induces
regeneration through blastema formation. It is suggested that FGF-sig-
naling and MMP activities participate in creating a regenerative environ-
ment. To understand why urodele amphibians can create such a
regenerative environment and humans cannot, it is necessary to identify
the similarities and differences between regenerative and nonregenera-
tive animals. Here we focus on ALM to consider limb regeneration from a
new perspective and we also reported that focal adhesion kinase (FAK)–
Src signaling controlled fibroblasts migration in axolotl limb regeneration.
Anat Rec, 295:1566–1574, 2012. V C 2012 Wiley Periodicals, Inc.
Key words: limb regeneration; accessory limb model; fibroblast
growth factor; matrix metalloproteinase
The tremendous regeneration ability of urodele portant role in the induction (Wallace, 1981). AEC is an
amphibians that helps regenerate lost limbs has long epithelial structure located in the distal region of the
fascinated scientists. Once a limb is amputated, undiffer- blastema and is functionally similar to the apical
entiated cells called blastema cells arise at the ectodermal ridge (AER), which appears during limb de-
amputation site. Although the origin of blastema cells velopment in higher vertebrates (Saunders, 2002).
remains unknown, dermal fibroblasts are believed to be Therefore, gene expression patterns and functions of
one of the major sources (Gardiner et al., 1986; Muneoka
et al., 1986; Kragl et al., 2009; Hirata et al., 2010). Der- Grant sponsors: JST PRESTO; Scientific Research on
mis-derived blastema cells redifferentiate into various Innovative Areas; Grant-in-Aid for Scientific Research on
cell types during limb regeneration. Moreover, dermal Innovative Areas (#23124508).
fibroblasts regulate patterning of the regenerating limb *Correspondence to: Akira Satoh, PhD, Associate Professor,
(Tank, 1981; Maden and Mustafa, 1982; Rollman-Din- Okayama University, Research Core for Interdisciplinary
smore and Bryant, 1982). The main role of dermal Sciences (RCIS), 3-1-1, Tsushima-naka, Kita-ku, Okayama
fibroblasts in limb regeneration justifies the study of 700-8530, Japan. E-mail: satoha@cc.okayama-u.ac.jp
their behavior and function during limb regeneration. Received 1 December 2011; Accepted 26 March 2012.
While the mechanism of blastema induction from DOI 10.1002/ar.22529
mature dermal tissue remains unknown, the apical Published online 29 August 2012 in Wiley Online Library
epithelial cap (AEC) has been suggested to play an im- (wileyonlinelibrary.com).
C 2012 WILEY PERIODICALS, INC.
V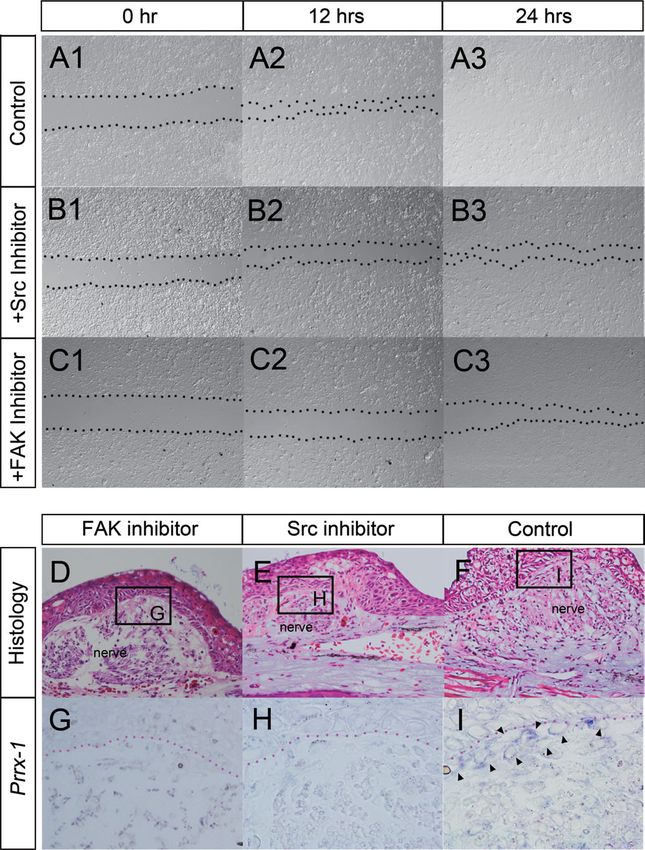
EARLY REGULATION OF AXOLOTL LIMB REGENERATION 1567
AEC have been studied and compared to those of AER. and showed cartilage differentiation ability (Satoh et al.,
Not only AEC but also blastema mesenchyme has been 2007). Thus, skin wounding and nerve deviation are suf-
likened to the limb bud mesenchyme of higher verte- ficient to induce blastema formation, and it is the
brates. In fact, an induced regenerating blastema has presence of the nerve that distinguishes wound healing
features similar to those of a developing limb bud from blastema formation.
(Muneoka and Bryant, 1982). Most limb regeneration How does a nerve control blastema induction? Nerve
processes are believed to repeat developmental processes functions have long been the focus of limb regeneration
to restore the original structure (Gardiner et al., 2002). studies. Neurotrophic factors are believed to be involved
Indeed, the later phase of limb regeneration is called the in limb regeneration, and several factors have been iso-
redevelopment phase. Unique and key events occur dur- lated, including FGFs, GGF, transferrin, substance P,
ing the early phase of limb regeneration. In limb and anterior gradient (AG) (Brockes and Kintner, 1986;
development, undifferentiated tissues, such as the lat- Mullen et al., 1996; Wang et al., 2000; Nye et al., 2003;
eral plate mesoderm (LPM), are located adjacent to the Kumar et al., 2007; Satoh et al., 2008b, 2009a., 2010a,
limb induction site. Once AER is induced, LPM becomes 2011). However, the detailed role of these factors in blas-
a source of undifferentiated cells for the limb bud (Fer- tema induction remains unknown. ALM has revealed
nandez-Teran and Ros, 2008). In contrast, in limb that inducing AEC is one of the first nerve functions to
regeneration, a mature limb consists of mature tissues occur in limb regeneration (Satoh et al., 2008b). The
and does not have undifferentiated or embryonic tissues nerve–AEC relationship had already been implicated in
such as LPM. Blastema cells emerge from such differen- some studies (Singer, 1949; Satoh et al., 2008b; Thorn-
tiated tissues, and therefore, their induction appears to ton, 1954). ALM appears to be a suitable system for
be a regeneration specific process as compared to limb investigating early nerve function and AEC induction
development. To elucidate the superior regeneration owing to the ease with which nerve participation can be
ability of urodele amphibians, it is necessary to focus on surgically manipulated on ALM from the beginning to
the unique early phase of limb regeneration, including the end of limb regeneration.
dedifferentiation (reprogramming) of dermal fibroblasts. Comparative study of wound healing and blastema for-
The accessory limb model (ALM) was developed by mation has been undertaken to determine what
Endo et al. after reviewing classical studies to investi- distinguishes these two processes. Skin wounding without
gate the early events of limb regeneration (Bodemer, nerve deviation results in simple wound healing, whereas
1959; Lheureux, 1977; Reynolds et al., 1983; Maden and wounding with nerve deviation results in blastema induc-
Holder, 1984; Egar, 1988; Endo et al., 2004). A skin tion (Endo et al., 2004). Skin wound healing and blastema
wound, deviated nerve, and contralateral skin graft are formation can be compared easily using ALM. Amputated
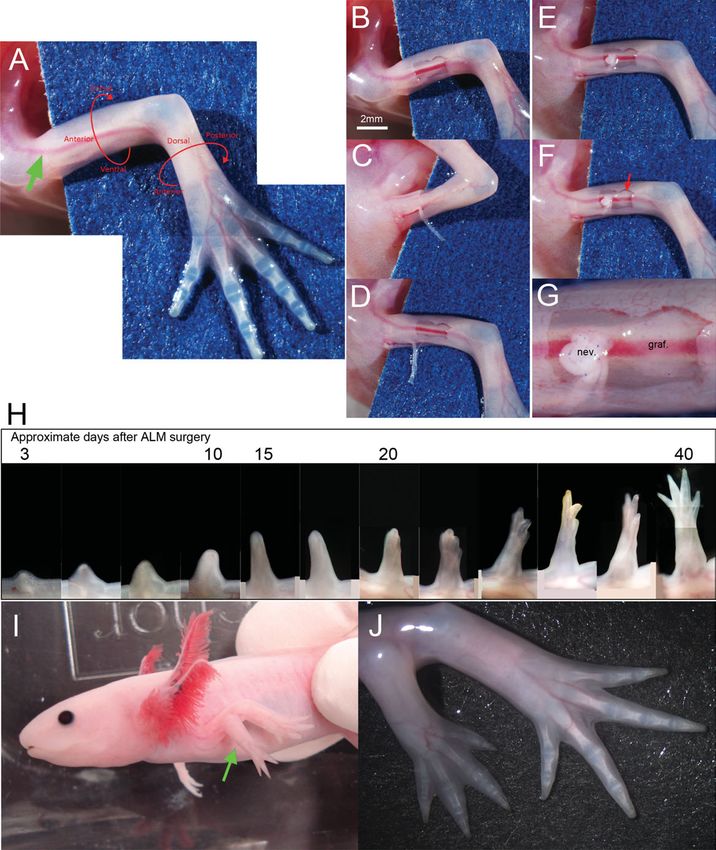
sufficient to induce accessory limb formation (Fig. 1). limbs have been used for the same purpose, but such a com-
Generally, a small square skin wound is created in an parison is difficult for several reasons. First, it is difficult to
anterior upper limb (Fig. 1B). Special attention must be induce wound healing without blastema formation.
given to the orientation of the axolotl forelimb because it Denerved limbs, which cannot grow a blastema, have been
is twisted (Fig. 1A). A relatively larger artery runs along used as the wound-healing control, but repeated denerva-
the anterior side (Fig. 1A, green arrow), providing a tion surgery is required to preserve the denerved state
good landmark for the anterior side of the forelimb. because axolotl axons regrow quickly (Schotte and Butler,
Axons are cut at the elbow/knee level and dissected out 1941; Petrosky et al., 1980). Second, it is difficult to deter-
from the ventral side (Fig. 1C). The free axon ends are mine whether or not denervation is complete at the time
then placed on the anterior wound (Fig. 1D,E); these are samples are collected. The success of denervation can be
sufficient to induce a blastema, though the induced blas- determined only several days after surgery. Third, multiple
tema regresses at the end, as mentioned later. To induce tissues are present in a limb, and amputation damages all
regeneration of a perfectly patterned limb, a small piece of them. Each tissue has its own healing process independ-
of skin from the contralateral side (posterior) should be ent of epimorphic regeneration, leading to higher
grafted by the deviated nerve (Fig. 1F,G). If the skin background in a comparative study. Fourth, limb regenera-
graft is not prepared from the exact contralateral region, tion induced by amputation is likely to be driven by two
a limb is not induced or is malformed, usually with distinct regeneration systems (Satoh et al., 2010a). One is
fewer digits than normal. A blastema starts forming on AEC-dependent and regenerates distal structure(s), while
top of the deviated nerve and continues to grow as the other is AEC-independent and regenerates proximal
observed during normal limb regeneration (Fig. 1H). structures (Bryant and Iten, 1977). Amphibian limb regen-
Although it remains difficult to obtain a perfectly pat- eration has generally been studied with respect to AEC
terned limb (one possessing all skeletal elements), formation, and in this approach the AEC-independent
regeneration of a limb structure can be induced at a mechanism adds noise to the analysis. ALM is a superior
high rate (>73%) by means of the skin graft (Fig. 1I,J; model for focusing on the AEC-dependent mechanism
Endo et al., 2004). As mentioned above, the skin graft because this is the only mechanism used in ectopic
from the contralateral side is not necessary to induce an limb regeneration. Thus, ALM provides an ideal model for a
ectopic blastema. An ectopic blastema can be induced by study comparing wound healing with blastema formation
skin wounding and nerve deviation without skin graft- and is an advanced model to investigate limb regeneration.
ing (Endo et al., 2004; Satoh et al., 2007). Without skin
grafting the blastema does not continue growing and, in MATRIX METALLOPROTEINASE (MMP)
most cases, it starts regressing (Endo et al., 2004). How-
ever, although the induced blastema cannot continue
ACTIVATION
growing, it shows blastema features (Satoh et al., 2007). The very beginning of limb regeneration shares a com-
These blastema cells expressed blastema marker genes mon mechanism with wound healing in which MMPs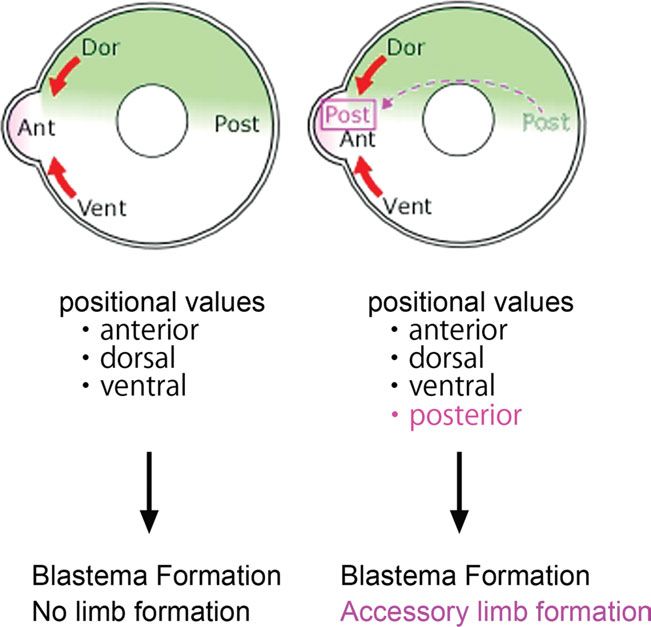
1568 MAKANAE AND SATOH Fig. 1. Accessory limb induction. Nerve deviation and a skin graft elbow level and then dissected out. (E) The nerve ends were placed are sufficient to induce an accessory limb. (A–G) procedure of induc- on the skin wound. (F) A skin graft (red arrow) was prepared from the tion of an accessory limb on an upper arm. (A) Normal axolotl forearm. contralateral side of the limb (posterior) and placed adjacent to the When an axolotl is laid on a plastic dish, the orientation of the forelimb deviated nerves. (G) Higher magnification of F. nev ¼ nerve. graf. ¼ must be noted. Limb orientation is directed by red arrows and letters. skin graft. (H) Time course of accessory limb growth. Numbers at top Green arrow indicates the anterior artery running along the anterior of panels indicate the approximate days after limb accessory surgery. side. (B) Skin wounding was performed prior to the nerve deviation. (I) Accessory limb on the upper arm (green arrow). (J) Higher-magnifi- (C, D) Nerves running along ventral blood vessels were cut at the cation view of the accessory limb. are involved (Gill and Parks, 2008; Parks, 2008; Satoh limb regeneration (Yang and Bryant, 1994; Miyazaki et al., 2011). MMPs are enzymes that digest the extrac- et al., 1996; Yang et al., 1999; Vinarsky et al., 2005). ellular matrix (ECM) such as collagens. The number of GM6001, a broad MMP inhibitor, inhibits limb regenera- MMP genes in an axolotl remains uncertain, but some tion (Vinarsky et al., 2005); however, the mechanism MMPs appear to be upregulated from the early phase of remains unclear. Immediately after limb amputation,
EARLY REGULATION OF AXOLOTL LIMB REGENERATION 1569
the surrounding epidermis starts migrating to cover the et al., 2000; Han et al., 2001). FGF-8 is expressed in AEC
amputation surface. MMPs are expressed in migrating during amphibian limb regeneration (Endo et al., 2000;
epidermis/epithelial cells (Mullen et al., 1996; Satoh Han et al., 2001). The basal layer of blastema epithelium,
et al., 2008b, 2009b) and MMP-9 is strongly expressed in where Sp9 is expressed, expresses FGF-8. Considering
the basal layer of these cells. Not only the migrating epi- the FGF gene expression pattern in AER of higher verte-
dermis but also leucocytes appear to show high MMP brates (Niswander et al., 1993; Savage et al., 1993;
activity by histological observation (Satoh et al., 2008b). Mahmood et al., 1995; Moon et al., 2000; Niswander,
Specific leucocyte types cannot be identified because no 2003), it is probable that other FGF genes are expressed
axolotl leucocyte antibodies are available. However, from in AEC, in view of the apparent equivalence of the AEC
our knowledge of higher vertebrates and of the histology and AER structures. AEC promotes mitotic activity of
of the regenerating axolotl limb, we expect that at least blastema cells (Boilly and Albert, 1990); this supports the
neutrophils and microphages accumulate around an idea that AEC is another FGF source because many
amputation surface and express MMPs (Fishman and FGFs show mitotic activity. Characteristically, FGF-8 can
Hay, 1962; Tsonis, 1996; Gill and Parks, 2008). It is rea- be detected in axolotl blastema mesenchyme and in AEC
sonable to expect an accumulation of leucocytes after (Han et al., 2001), although FGF-8 is expressed specifi-
amputation as an anti-inflammatory response. MMP cally in AER in higher vertebrates (Mahmood et al.,
activities increase in the damaged region because of the 1995). However, blastema cells appear after AEC induc-
migrating epidermis and leucocytes and might be nerve- tion. Therefore, the initial FGF source could be AEC and
independent. However, it has been suggested that MMP the nerve. Coincidentally, fibronectin, which is related to
activity is at least partially controlled by nerves during FGF signaling, is expressed in AEC and distal blastema
later phases of regeneration (Yang et al., 1999; Satoh mesenchyme (Nace and Tassava, 1995). It is likely that
et al., 2011). In higher vertebrates, digestion of ECM by FGFs are enriched in the distal region of blastema
MMPs triggers cellular activation, resulting in activation because FGFs can bind fibronectin (Martino and Hubbell,
of Prrx1, a blastema cell marker gene expressed during 2010). Thus, FGFs secreted by nerves and AEC might be
amphibian limb regeneration (Jones et al., 1999, 2001; enriched by binding to fibronectin in the distal region of
McKean et al., 2003; Suzuki et al., 2005; Satoh et al., the blastema. Thus, FGF-dominant conditions are created
2007). In case of axolotls, digestion of ECM also acti- in the distal region of the regenerating limb.
vated Prrx1 in skin cultures (Satoh et al., 2011). This
may be reasonable because fibroblasts adhere to ECM in CELL ACCUMULATION AT THE SITE
a normal (unamputated) limb, but they must be free
from ECM to migrate toward a damaged region during
WHERE A BLASTEMA FORMS
limb regeneration. Fibroblasts, which are free from In ALM, a blastema is induced on top of a deviated
ECM, may be called preblastema cells. Suzuki et al. nerve, suggesting that the nerve attracts blastema cells
(2005) originally proposed fibroblast activation before from the surrounding tissue. Migration of dermal fibro-
blastema cell induction. Induction of fibroblast activation blasts was observed previously in axolotl limb
occurs through the integrin/focal adhesion kinase (FAK) regeneration (Gardiner et al., 1986). Gardiner et al. used
signaling pathway (McKean et al., 2003; Satoh et al., a diploid/triploid cell marker in the axolotl to investigate
2011). Although this activation mechanism is conserved the movement of cells from the dermis into the early
among species, the induction of full blastema cells from limb blastema. Cells of dermal origin began to migrate
activated fibroblasts has not been reported. beneath the WE at about 5 days postamputation, and by
10 days they were widely distributed across the amputa-
FIBROBLAST GROWTH FACTOR tion surface. By 15 days, a dense accumulation of
(FGF)-DOMINANT ENVIRONMENT blastema cells was detected beneath the apical cap, and
these cells were oriented preferentially in a circumferen-
CREATED BY AEC AND NERVES tial direction. In ALM, dermal fibroblast migration from
FGFs have been suggested as the neurotrophic factors a dermis to a blastema was traced by injecting red fluo-
involved in limb regeneration. Damaged axons are specu- rescent dye into a dermis. In ALM, as in regular limb
lated to release FGFs (Poulin et al., 1993; Satoh et al., regeneration, dermal fibroblasts migrated into a blas-
2008b, 2011). Furthermore, some FGFs are expressed in tema (Endo et al., 2004). However, although FGF
dorsal root ganglia, from which sensory nerves are pro- signaling may play a role in the migration mechanisms
jected to limbs (Li et al., 2002; Satoh et al., 2008b, 2011). of fibroblasts toward a blastema, the mechanism
Basic FGF (FGF-2), which is expressed in nerves, rescues remains unclear. FGF has been suggested as a fibroblast
the denervation effect during limb regeneration (Mullen chemoattractant in a higher vertebrate (Li and
et al., 1996). Studies with ALM have revealed that FGF Muneoka, 1999). FGF-4, which was expressed in an
signaling induces expression of Sp9, an AEC marker AER, showed chemotactic activity for limb bud cells.
gene, in an overlying wound epithelium (Satoh et al., Although FGF-4 expression has not been observed in a
2008b). The deviated nerves are covered by a migrating regenerating blastema (Christensen et al., 2002), it has
epidermis/epithelium called wound epithelium (WE) been reported that ectopic AEC transplantation resulted
within several hours. Then direct interaction between the in additional blastema growth (Thornton, 1960). This
nerve and WE occurs. Sp9 is initially induced broadly implies that the ectopic AEC accumulates blastema cells
throughout the layers of the overlying WE, after which and directs the growth of an additional limb. Given that
expression begins to be restricted to the basal layer of FGFs showed mutual functional redundancy, other
WE. Within 5 days of ALM surgery, Sp9 is completely re- FGFs would function as chemoattractants. Elucidation
stricted to the basal layer, suggesting that AEC is of the migration mechanism is important for under-
induced. AEC also functions as an FGF source (Endo standing cell movement in limb regeneration as well as1570 MAKANAE AND SATOH
chemotaxis; however, it has received little attention. To sufficient to induce a blastema, but the induced blas-
focus on this issue, cell migration was investigated fo- tema regresses; such a regressing blastema develops a
cusing on FAK/Src signaling. A focus on integrin/FAK/ cartilage, which eventually disappears. The cartilage in
Src signaling was suggested, since this signaling system a regressing blastema is usually round and single (Endo
regulates cell migration in fibroblasts of higher verte- et al., 2004), suggesting that there is no developmental
brates and is activated in axolotl limb regeneration axis. This patternless structure has similarity to a
(Jones et al., 1999, 2001; Parsons and Parsons, 2004; regenerating Xenopus froglet limb (Satoh et al., 2005).
Mitra et al., 2005; Satoh et al., 2011). The ECM diges- Xenopus can regrow a structure distally after amputa-
tion leads to an alternation of integrin gene expression. tion, but the structure does not have a definite pattern
As mentioned above, ECM degradation can be expected and is called a ‘‘spike.’’ The cartilage in the spike is cone-
in the early phase of limb regeneration. Indeed, integrin shaped, but no patterning occurs along the anterior–pos-
switching from integrin b1 to b3 occurs in this phase terior (AP) axis (Yakushiji et al., 2009). This defect is at
(Tsonis et al., 1997; Satoh et al., 2011). FAK phosphoryl- least partially because of the loss of Shh expression
ation follows integrin switching (Satoh et al., 2011). Src (Yakushiji et al., 2009). Inhibiting Shh signaling results
is reported as a downstream gene of FAK that controls in the same patternless structure in axolotls (Roy and
cell migration in higher vertebrates (Sieg et al., 1999). Gardiner, 2002). Thus, it would be worthwhile to investi-
Accordingly, we investigated FAK/Src signaling in cell gate Shh expression in ALM blastema. Although Shh
migration. Axolotl dermal fibroblasts were isolated enzy- expression may underlie the second determination fac-
matically and cultured in a plastic dish. A scratch assay tor, the factor remains unknown. An accessory limb
was performed to investigate cell migration (Fig. 2). Axo- forms when contralateral skin is placed adjacent to a
lotl dermal fibroblasts were confluently cultured on a deviated nerve, suggesting that all positional informa-
plate and a gap was made with a pipette tip (Fig. 2A1). tion is present and necessary for a patterned limb (Fig.
Cells migrated and filled the gap within 24 h (Fig. 3). When a blastema is created on the anterior side with-
2A2,A3). As expected, treatment with FAK and Src out the skin graft, presumably three of the four
inhibitors delayed cell migration (Fig. 2B1–3,C1–3). This positional values—such as anterior, ventral, and dor-
FAK/Src-dependent migration was also confirmed in vivo sal—can be expected in the wound (Fig. 3, left).
(Fig. 2D–I). Animals that underwent accessory limb sur- Anterior–dorsal and anterior–ventral dermal fibroblasts
gery were administered inhibitors for 5 days. The can accumulate in the nerve’s deviated region (Fig. 3,
blastemas were harvested and sectioned for in situ red arrows). However, posterior fibroblasts cannot accu-
hybridization. In the control (no inhibitor), Prrx1-posi- mulate in the nerve’s deviated region because they are
tive cells could be detected between the overlying located at a great distance for this region. A skin graft
epithelium and the deviated nerve (Fig. 2F,I, arrow- from the contralateral side provides the missing value
heads). In contrast, Prrx1-positive cells were not (Fig. 3, right). The skin graft contains many posterior
observed at the site where a nerve was rerouted when a dermal fibroblasts placed adjacent to the deviated nerve
FAK or Src inhibitor was administered (Fig. 2D,E,G,H), (Fig. 1G). Therefore, posterior dermal fibroblasts can
suggesting that FAK/Src signaling plays a role in axolotl participate in a blastema as well as in dermal fibroblasts
fibroblast migration similar to as in higher vertebrates. from other three regions (Fig. 3, right). These mixed
It is possible that the inhibitors interrupt cell prolifera- positional values may constitute a force for limb pattern-
tion because of which cell migration appeared slow. ing. However, the importance of positional values in
However, because cultured axolotl cells showed very amphibian limb development and regeneration is not yet
slow cell proliferation (Boilly and Albert, 1988), it is well explained at the molecular level. There are at least
unlikely that cell proliferation contributed appreciably to three axes during limb development and regeneration,
migration. Fibroblasts accumulating by FGF-dependent including the AP, dorsal–ventral (DV), and proximal–dis-
chemotaxis and a FAK/Src-dependent migration mecha- tal (PD). The PD axis has been most studied during limb
nism may thus be responsible for blastema formation on regeneration. HoxA genes, which are regulated along
top of a deviated nerve. the PD axis, have been reported (Savard et al., 1988;
Simon and Tabin, 1993; Beauchemin et al., 1994;
STEPWISE LIMB REGENERATION Gardiner et al., 1995; Endo et al., 2000; Christen et al.,
2003; Ohgo et al., 2010). However, the AP and DV axes
REGULATION are not well studied at the molecular level, and only a
ALM studies are based on the stepwise model pro- few studies have been published on Xenopus (Takaba-
posed by Endo et al. (2004). The names of ‘‘ALM’’ and take et al., 2000; Matsuda et al., 2001). Even the gene
‘‘stepwise model’’ would be a little confusing. But ALM is expression patterns related to the DV and AP axes have
an experimental system to probe a new concept of limb not been reported in axolotls. To investigate whether the
regeneration named stepwise model. In the stepwise same position-specific genes as in higher vertebrates
model, at least three steps and two determination fac- were expressed in axolotl blastema, we performed in situ
tors are necessary for amphibian limb regeneration. In hybridization (Fig. 4). Lmx1b is expressed in a dorsal
the first step, wound healing takes place after skin region of a developing limb bud in higher vertebrates
wounding. When nerves are supplied to the wound, (Vogel et al., 1995; Loomis et al., 1998) and in the dorsal
instead of wound healing, a blastema grows at the region of the axolotl blastema (Fig. 4A). Tbx3 is used as
wound site (second step). As mentioned previously, it is an AP marker gene in higher vertebrates because of its
the availability of a nerve that distinguishes the first expression in the anterior and posterior edge of a limb
and second steps. The third step is limb patterning, bud (Bamshad et al., 1997). Tbx3 was similarly
which is determined presumably by mixed positional in- expressed in AP regions of the axolotl blastema
formation. Interestingly, skin wounding and nerves are (Fig. 4B). The expression pattern of these genes wouldEARLY REGULATION OF AXOLOTL LIMB REGENERATION 1571
Fig. 2. FAK/Src signaling regulates axolotl fibroblast migration. A on FAK- (D, G) or Src inhibitor-containing (E, H) water. (D–F) Hematox-
scratch assay was performed to investigate the regulatory mechanism ylin and eosin staining. (G–I) Prrx-1 expression pattern by in situ
of axolotl fibroblast migration. (A) Axolotl fibroblasts filled the gap hybridization in the boxed region of D–F. (G, H) Prrx1-positive blas-
within 24 hr. (B) The Src inhibitor SU6656 delayed cell migration com- tema cells were undetectable at the induction site 5 days after sur-
pared with that in the control. The gap was observable even 24 hr af- gery. (I) Prrx1-positive cells were observable 5 days after surgery
ter scratching (B3). (C) A FAK inhibitor also delayed cell migration. (arrowheads) in the control. These results suggest that migration of
Dotted lines in A–C indicate the gap (acellular region). (D–I) Ectopic Prrx1-positive cells toward the blastema induction region was con-
blastema was induced with or without inhibitors. Transverse sections trolled by FAK/Src signaling. Red dotted lines in G–I indicate the
of an induced blastema in ALM. Blastema induction procedure boundary of the blastema epidermis.
(wounding þ nerve deviation) was performed, and animals were raised
be helpful for studying the role of positional values in POSSIBILITY OF APPLICATIONS TO
limb regeneration, although the expression pattern of
position-specific genes does not represent positional
HIGHER VERTEBRATES
value itself. While we can analyze limb regeneration in Many processes are common to limb regeneration in
a stepwise fashion with the ALM system, molecular data urodele amphibians and limb development in higher ver-
will be required for further progress. tebrates. The FGF-signaling pathway plays a role in1572 MAKANAE AND SATOH
Fig. 3. Presumptive positional values in ALM. Presumptive aggrega-
tion of positional values in ALM. Illustrations are transverse sections of
a limb. (Left) Three positional values can be expected when a blas-
tema is induced on the anterior side, but no posterior value is gener-
ated. Blastema formation takes place despite the absence of one Fig. 4. Lmx1b and Tbx3 expression patterns in a late bud blas-
positional value. (Right) A contralateral graft supplies the missing, that tema. (A) Lmx1b expression in a late bud blastema. Dorsal blastema
is, posterior, value. In this case, a patterned limb forms in the pres- cells were Lmx1b positive (arrowheads). (B) Tbx3 expression in a late
ence of all four values. bud blastema. Anterior and posterior blastema cells were Tbx3 posi-
tive (arrowheads).
inducing a limb bud from a LPM during limb develop-
ment (Tanaka and Gann, 1995). Based on ALM studies, it tivate most of their developmental genetic networks
is likely that FGF signaling also functions during the after birth. The question why higher vertebrates cannot
induction phase of limb regeneration (Satoh et al., induce a blastema after fibroblast activation needs to be
2008a,b, 2011). Moreover, AER or AER factor (FGF-2 or answered. We hope that a combination of ALM and
FGF-4) grafting onto an amputated limb bud in chick recently developed high-throughput analysis will provide
embryos induced a regeneration response (Hayamizu the answer.
et al., 1994; Taylor et al., 1994; Kostakopoulou et al.,
1996; Satoh et al., 2010b). Hence, FGF signaling likely
plays a role in the early phase of limb development and
MATERIALS AND METHODS
regeneration. In addition, as mentioned above, Prrx1 acti- Axolotl dermal fibroblasts and blastema cells were
vation through the integrin/FAK-signaling pathway also prepared as described previously (Satoh et al., 2007). Ba-
appears to be conserved in both human and axolotl fibro- sic culture medium contained 80% L15, 1% fetal calf
blasts (McKean et al., 2003; Satoh et al., 2011). Notably, serum (FCS), and 200 lg mL1 gentamycin. Mouse FGF-
Prrx1 is a direct upstream regulator of tenascin-C in 2 (100 ng mL1; R&D Systems) and human FGF-8 (50
human fibroblasts, and tenascin-C is a classical blastema ng mL1; Peprotech) were supplied for experimental
marker gene (Onda et al., 1990). Thus, it is likely that purposes. Axolotl dermal fibroblasts were cultured in a
higher vertebrates maintain a basic gene network(s) of penicillin cup to obtain confluent cells for the scratch
regeneration even though some elements may be missing. assay. After a 24-hr incubation, a scratch was made with
Research into limb regeneration in higher vertebrates a 2-lL pipette tip and then inhibitors were added. FAK
has already started and it is becoming possible to supply inhibitor 14 (20 lM; Sigma) and Src inhibitor (3 lM,
missing factor(s) (Han et al., 2003, 2005, 2008; Masaki SU6656; Calbiochem) were used to study their effects on
and Ide, 2007; Muneoka et al., 2008; Satoh et al., 2010b; limb regeneration. Cell migration was observed under a
Yu et al., 2010; Fernando et al., 2011). These advances microscope (Olympus, SZX16).
encourage further attempts to describe the extent of con- The axolotls were raised in tap water. We used 10–12-
servation among amphibians and higher vertebrates. cm length axolotl. The ALM surgical procedure has been
Finally, new experimental techniques employing the described previously (Endo et al., 2004). They were kept
ALM afford new opportunities to understand blastema for several hours to heal the surgical damage and then
induction mechanisms in animals that can regenerate. immersed in tap water with or without inhibitors. Con-
ALM studies suggest that even the initiation mecha- centrations of inhibitors were the same as for the
nisms of limb regeneration, which has been considered scratch assay. The inhibitor-containing water was
to be specific to regenerative animals, are conserved in refreshed at Days 0, 2, and 4. Samples were harvested 5
higher vertebrates. However, human beings cannot reac- days after the ALM surgery.EARLY REGULATION OF AXOLOTL LIMB REGENERATION 1573
The in situ hybridization procedure was the same as Han MJ, Yang X, Lee J, Allan CH, Muneoka K. 2008. Development
that used in a previous study (Satoh et al., 2007). and regeneration of the neonatal digit tip in mice. Dev Biol 315:
Lmx1b and Tbx3 were isolated by RT-PCR as previously 125–135.
Han MJ, Yang X, Taylor G, Burdsal CA, Anderson RA, Muneoka K.
described (Satoh et al., 2007). Their gene sequences
2005. Limb regeneration in higher vertebrates: developing a road-
have been deposited in Sal-Site (http://www.ambystoma. map. Anat Rec B New Anat 287:14–24.
org/). Hayamizu TF, Wanek N, Taylor G, et al. 1994. Regeneration of
HoxD expression domains during pattern regulation in chick
ACKNOWLEDGEMENTS wing buds. Dev Biol 161:504–512.
Hirata A, Gardiner DM, Satoh A. 2010. Dermal fibroblasts contrib-
The authors thank Dr. D. M. Gardiner and Dr. T. Endo ute to multiple tissues in the accessory limb model. Dev Growth
for constructive comments. Differ 52:343–350.
Jones FS, Meech R, Edelman DB, Oakey RJ, Jones PL. 2001. Prx1
controls vascular smooth muscle cell proliferation and tenascin-C
LITERATURE CITED expression and is upregulated with Prx2 in pulmonary vascular
disease. Circ Res 89:31–38.
Bamshad M, Lin RC, Law DJ, et al. 1997. Mutations in human Jones PL, Jones FS, Zhou B, Rabinovitch M. 1999. Induction of vas-
TBX3 alter limb, apocrine and genital development in ulnar- cular smooth muscle cell tenascin-C gene expression by denatured
mammary syndrome. Nat Genet 16:311–315. type I collagen is dependent upon a beta3 integrin-mediated mito-
Beauchemin M, Noiseux N, Tremblay M, Savard P. 1994. Expres- gen-activated protein kinase pathway and a 122-base pair pro-
sion of Hox A11 in the limb and the regeneration blastema of moter element. J Cell Sci 112 (Part 4):435–445.
adult newt. Int J Dev Biol 38:641–649. Kostakopoulou K, Vogel A, Brickell P, Tickle C. 1996. ‘‘Regeneration’’
Bodemer CW. 1959. Observations on the mechanism of induction of of wing bud stumps of chick embryos and reactivation of Msx-1
supernumerary limbs in adult Triturus viridescens. J Exp Zool and Shh expression in response to FGF-4 and ridge signals. Mech
140:79–99. Dev 55:119–131.
Boilly B, Albert P. 1988. Blastema cell proliferation in vitro: effects Kragl M, Knapp D, Nacu E, et al. 2009. Cells keep a memory of their
of limb amputation on the mitogenic activity of spinal cord tissue origin during axolotl limb regeneration. Nature 460:60–65.
extracts. Biol Cell 62:183–187. Kumar A, Godwin JW, Gates PB, Garza-Garcia AA, Brockes JP.
Boilly B, Albert P. 1990. In vitro control of blastema cell prolifera- 2007. Molecular basis for the nerve dependence of limb regenera-
tion by extracts from epidermal cap and mesenchyme of regener- tion in an adult vertebrate. Science 318:772–777.
ating limbs of axolotls. Roux’s Arch Dev Biol 198:443–447. Lheureux E. 1977. Importance of limb tissue associations in the de-
Brockes JP, Kintner CR. 1986. Glial growth factor and nerve-de- velopment of nerve-induced supernumerary limbs in the newt
pendent proliferation in the regeneration blastema of Urodele Pleurodeles waltlii Michah (author’s transl). J Embryol Exp Mor-
amphibians. Cell 45:301–306. phol 38:151–173.
Bryant SV, Iten LE. 1977. Intercalary and supernumerary regener- Li GD, Wo Y, Zhong MF, et al. 2002. Expression of fibroblast growth
ation in regenerating the mature limbs of Notophthalmus virides- factors in rat dorsal root ganglion neurons and regulation after
cens. J Exp Zool 202:1–16. peripheral nerve injury. Neuroreport 13:1903–1907.
Christen B, Beck CW, Lombardo A, Slack JM. 2003. Regeneration- Li S, Muneoka K. 1999. Cell migration and chick limb development:
specific expression pattern of three posterior Hox genes. Dev Dyn chemotactic action of FGF-4 and the AER. Dev Biol 211:335–347.
226:349–355. Loomis CA, Kimmel RA, Tong CX, Michaud J, Joyner AL. 1998.
Christensen RN, Weinstein M, Tassava RA. 2002. Expression of Analysis of the genetic pathway leading to formation of ectopic
fibroblast growth factors 4, 8, and 10 in limbs, flanks, and blaste- apical ectodermal ridges in mouse Engrailed-1 mutant limbs. De-
mas of Ambystoma. Dev Dyn 223:193–203. velopment 125:1137–1148.
Egar MW. 1988. Accessory limb production by nerve-induced cell Maden M, Holder N. 1984. Axial characteristics of nerve induced
proliferation. Anat Rec 221:550–564. supernumerary limbs in the axolotl. Roux’s Arch Dev Biol 193:
Endo T, Bryant SV, Gardiner DM. 2004. A stepwise model system 394–401.
for limb regeneration. Dev Biol 270:135–145. Maden M, Mustafa K. 1982. Axial organization of the regenerating
Endo T, Tamura K, Ide H. 2000. Analysis of gene expressions dur- limb: asymmetrical behavior following skin transplantation. J
ing Xenopus forelimb regeneration. Dev Biol 220:296–306. Embryol Exp Morphol 70:197–213.
Fernandez-Teran M, Ros MA. 2008. The apical ectodermal ridge: Mahmood R, Bresnick J, Hornbruch A, et al. 1995. A role for FGF-8
morphological aspects and signaling pathways. Int J Dev Biol 52: in the initiation and maintenance of vertebrate limb bud out-
857–871. growth. Curr Biol 5:797–806.
Fernando WA, Leininger E, Simkin J, et al. 2011. Wound healing Martino MM, Hubbell JA. 2010. The 12th-14th type III repeats of
and blastema formation in regenerating digit tips of adult mice. fibronectin function as a highly promiscuous growth factor-bind-
Dev Biol 350:301–310. ing domain. FASEB J 24:4711–4721.
Fishman DA, Hay ED. 1962. Origin of osteoclasts from mononuclear Masaki H, Ide H. 2007. Regeneration potency of mouse limbs. Dev
leucocytes in regenerating newt limbs. Anat Rec 143:329–337. Growth Differ 49:89–98.
Gardiner DM, Blumberg B, Komine Y, Bryant SV. 1995. Regulation Matsuda H, Yokoyama H, Endo T, Tamura K, Ide H. 2001. An epi-
of HoxA expression in developing and regenerating axolotl limbs. dermal signal regulates Lmx-1 expression and dorsal-ventral pat-
Development 121:1731–1741. tern during Xenopus limb regeneration. Dev Biol 229:351–362.
Gardiner DM, Endo T, Bryant SV. 2002. The molecular basis of am- McKean DM, Sisbarro L, Ilic D, et al. 2003. FAK induces expression
phibian limb regeneration: integrating the old with the new. of Prx1 to promote tenascin-C-dependent fibroblast migration. J
Semin Cell Dev Biol 13:345–352. Cell Biol 161:393–402.
Gardiner DM, Muneoka K, Bryant SV. 1986. The migration of dermal Mitra SK, Hanson DA, Schlaepfer DD. 2005. Focal adhesion kinase: in
cells during blastema formation in axolotls. Dev Biol 118:488–493. command and control of cell motility. Nat Rev Mol Cell Biol 6:56–68.
Gill SE, Parks WC. 2008. Metalloproteinases and their inhibitors: Miyazaki K, Uchiyama K, Imokawa Y, Yoshizato K. 1996. Cloning
regulators of wound healing. Int J Biochem Cell Biol 40:1334–1347. and characterization of cDNAs for matrix metalloproteinases of
Han MJ, An JY, Kim WS. 2001. Expression patterns of Fgf-8 during de- regenerating newt limbs. Proc Natl Acad Sci USA 93:6819–6824.
velopment and limb regeneration of the axolotl. Dev Dyn 220:40–48. Moon AM, Boulet AM, Capecchi MR. 2000. Normal limb develop-
Han MJ, Yang X, Farrington JE, Muneoka K. 2003. Digit regenera- ment in conditional mutants of Fgf4. Development 127:989–996.
tion is regulated by Msx1 and BMP4 in fetal mice. Development Mullen LM, Bryant SV, Torok MA, Blumberg B, Gardiner DM.
130:5123–5132. 1996. Nerve dependency of regeneration: the role of Distal-less1574 MAKANAE AND SATOH and FGF signaling in amphibian limb regeneration. Development Satoh A, Makanae A, Wada N. 2010b. The apical ectodermal ridge 122:3487–3497. (AER) can be re-induced by wounding, wnt-2b, and fgf-10 in the Muneoka K, Allan CH, Yang X, Lee J, Han M. 2008. Mammalian chicken limb bud. Dev Biol 342:157–168. regeneration and regenerative medicine. Birth Defects Res C Satoh A, Suzuki M, Amano T, Tamura K, Ide H. 2005. Joint devel- Embryo Today 84:265–280. opment in Xenopus laevis and induction of segmentations in Muneoka K, Bryant SV. 1982. Evidence that patterning mecha- regenerating froglet limb (spike). Dev Dyn 233:1444–1453. nisms in developing and regenerating limbs are the same. Nature Saunders JW, Jr. 2002. Is the progress zone model a victim of pro- 298:369–371. gress? Cell 110:541–543. Muneoka K, Fox WF, Bryant SV. 1986. Cellular contribution from Savage MP, Hart CE, Riley BB, Sasse J, Olwin BB, Fallon JF. 1993. dermis and cartilage to the regenerating limb blastema in axo- Distribution of FGF-2 suggests it has a role in chick limb bud lotls. Dev Biol 116:256–260. growth. Dev Dyn 198:159–170. Nace JD, Tassava RA. 1995. Examination of fibronectin distribution Savard P, Gates PB, Brockes JP. 1988 Position dependent expres- and its sources in the regenerating newt limb by immunocyto- sion of a homeobox gene transcript in relation to amphibian limb chemistry and in situ hybridization. Dev Dyn 202:153–164. regeneration. EMBO J 7:4275–4282. Niswander L. 2003. Pattern formation: old models out on a limb. Schotte OE, Butler EG. 1941. Morphological effects of denervation Nat Rev Genet 4:133–143. and amputation of limbs in urodele larvae. J Exp Zool 87:279–322. Niswander L, Tickle C, Vogel A, Booth I, Martin GR. 1993. FGF-4 Sieg DJ, Hauck CR, Schlaepfer DD. 1999. Required role of focal ad- replaces the apical ectodermal ridge and directs outgrowth and hesion kinase (FAK) for integrin-stimulated cell migration. J Cell patterning of the limb. Cell 75:579–587. Sci 112 (Ptar 16):2677–2691. Nye HL, Cameron JA, Chernoff EA, Stocum DL. 2003. Regeneration Simon HG, Tabin CJ. 1993. Analysis of Hox-4.5 and Hox-3.6 expres- of the urodele limb: a review. Dev Dyn 226:280–294. sion during newt limb regeneration: differential regulation of Ohgo S, Itoh A, Suzuki M, Satoh A, Yokoyama H, Tamura K. 2010. paralogous Hox genes suggest different roles for members of dif- Analysis of hoxa11 and hoxa13 expression during patternless ferent Hox clusters. Development 117:1397–1407. limb regeneration in Xenopus. Dev Biol 338:148–157. Singer M. 1949. The invasion of the epidermis of the regenerating Onda H, Goldhamer DJ, Tassava RA. 1990. An extracellular matrix forelimb of the urodele, Triturus, by nerve fibers. J Exp Zool 111: molecule of newt and axolotl regenerating limb blastemas and 189–209. embryonic limb buds: immunological relationship of MT1 antigen Suzuki M, Satoh A, Ide H, Tamura K. 2005. Nerve-dependent and - with tenascin. Development 108:657–668. independent events in blastema formation during Xenopus froglet Parks WC. 2008. Matrix metalloproteinases: introduction. Semin limb regeneration. Dev Biol 286:361–375. Cell Dev Biol 19:1. Takabatake Y, Takabatake T, Takeshima K. 2000. Conserved and Parsons SJ, Parsons JT. 2004. Src family kinases, key regulators of divergent expression of T-box genes Tbx2-Tbx5 in Xenopus. Mech signal transduction. Oncogene 23:7906–7909. Dev 91:433–437. Petrosky N, Tassava RA, Olsen CL. 1980. Cellular events in dener- Tanaka EM, Gann AF. 1995 Limb development. The budding role of vated limb stumps of Ambystoma larvae during reinnervation FGF. Curr Biol 5:594–597. and subsequent regeneration. Experientia 36:601–603. Tank PW. 1981. The ability of localized implants of whole or minced Poulin ML, Patrie KM, Botelho MJ, Tassava RA, Chiu IM. 1993. dermis to disrupt pattern formation in the regenerating forelimb Heterogeneity in the expression of fibroblast growth factor recep- of the axolotl. Am J Anat 162:315–326. tors during limb regeneration in newts (Notophthalmus virides- Taylor GP, Anderson R, Reginelli AD, Muneoka K. 1994. FGF-2 cens). Development 119:353–361. induces regeneration of the chick limb bud. Dev Biol 163: Reynolds S, Holder N, Fernandes M. 1983. The form and structure 282–284. of supernumerary hindlimbs formed following skin grafting and Thornton CS. 1954. The relation of epidermal innervation to limb nerve deviation in the newt Triturus cristatus. J Embryol Exp regeneration in Ambystoma larvae. J Exp Zool 127:577–601. Morphol 77:221–241. Thornton CS. 1960. Influence of an eccentric epidermal cap on limb Rollman-Dinsmore C, Bryant SV. 1982. Pattern regulation between regeneration in Amblystoma larvae. Dev Biol 2:551–569. hind- and forelimbs after blastema exchanges and skin grafts in Tsonis PA. 1996. Limb regeneration. Cambridge: Cambridge Univer- Notophthalmus viridescens. J Exp Zool 223:51–56. sity Press. Roy S, Gardiner DM. 2002. Cyclopamine induces digit loss in regen- Tsonis PA, Doane K, Del Rio-Tsonis K. 1997. Expression of integrins erating axolotl limbs. J Exp Zool 293:186–190. during axolotl limb regeneration. Dev Growth Differ 39:9–14. Satoh A, Bryant SV, Gardiner DM. 2008a. Regulation of dermal fibro- Vinarsky V, Atkinson DL, Stevenson TJ, Keating MT, Odelberg SJ. blast dedifferentiation and redifferentiation during wound healing 2005. Normal newt limb regeneration requires matrix metallopro- and limb regeneration in the Axolotl. Dev Growth Differ 50:743–754. teinase function. Dev Biol 279:86–98. Satoh A, Cummings GM, Bryant SV, Gardiner DM. 2009a. Neuro- Vogel A, Rodriguez C, Warnken W, Izpisua Belmonte JC. 1995. Dor- trophic regulation of fibroblast dedifferentiation during limb skel- sal cell fate specified by chick Lmx1 during vertebrate limb devel- etal regeneration in the axolotl (Ambystoma mexicanum). Dev opment. Nature 378:716–720. Biol 337:444–457. Wallace H. 1981. Vertebrate limb regeneration. New York: Wiley. Satoh A, Cummings GM, Bryant SV, Gardiner DM. 2010a. Regulation Wang L, Marchionni MA, Tassava RA. 2000. Cloning and neuronal of proximal-distal intercalation during limb regeneration in the axo- expression of a type III newt neuregulin and rescue of dener- lotl (Ambystoma mexicanum). Dev Growth Differ 52:785–798. vated, nerve-dependent newt limb blastemas by rhGGF2. J Neu- Satoh A, Gardiner DM, Bryant SV, Endo T. 2007. Nerve-induced ec- robiol 43:150–158. topic limb blastemas in the Axolotl are equivalent to amputation- Yakushiji N, Suzuki M, Satoh A, Ide H, Tamura K. 2009. Effects of induced blastemas. Dev Biol 312:231–244. activation of hedgehog signaling on patterning, growth, and dif- Satoh A, Graham GM, Bryant SV, Gardiner DM. 2008b. Neurotro- ferentiation in Xenopus froglet limb regeneration. Dev Dyn 238: phic regulation of epidermal dedifferentiation during wound heal- 1887–1896. ing and limb regeneration in the axolotl (Ambystoma Yang EV, Bryant SV. 1994. Developmental regulation of a matrix mexicanum). Dev Biol 319:321–335. metalloproteinase during regeneration of axolotl appendages. Dev Satoh A, James MA, Gardiner DM. 2009b. The role of nerve signal- Biol 166:696–703. ing in limb genesis and agenesis during axolotl limb regeneration. Yang EV, Gardiner DM, Carlson MR, Nugas CA, Bryant SV. 1999. J Bone Joint Surg Am 91 (Suppl 4):90–98. Expression of Mmp-9 and related matrix metalloproteinase genes Satoh A, Makanae A, Hirata A, Satou Y. 2011. Blastema induction during axolotl limb regeneration. Dev Dyn 216:2–9. in aneurogenic state and Prrx-1 regulation by MMPs and FGFs Yu L, Han M, Yan M, Lee EC, Lee J, Muneoka K. 2010. BMP sig- in Ambystoma mexicanum limb regeneration. Dev Biol 355: naling induces digit regeneration in neonatal mice. Development 263–274. 137:551–559.
You can also read

















































