Insidious adrenocortical insufficiency underlies
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The FASEB Journal • Research Communication
Insidious adrenocortical insufficiency underlies
neuroendocrine dysregulation in TIF-2 deficient mice
Alexandre V. Patchev,*,†,1 Dieter Fischer,† Siegmund S. Wolf,‡ Miles Herkenham,§
Franziska Götz,* Martine Gehin,储 Pierre Chambon,储 Vladimir K. Patchev,‡ and
Osborne F. X. Almeida†
*Institute of Experimental Endocrinology, Charité School of Medicine, Humboldt University,
Berlin, Germany; †Neuroadaptations Group, Max Planck Institute of Psychiatry, Munich, Germany;
‡
Corporate Research Gynecology and Andrology, Schering AG, Jena, Germany; §Section on
Functional Neuroanatomy, National Institute of Mental Health, National Institutes of Health,
Bethesda, Maryland, USA; 储Institute de Génétique et de Biologie Moléculaire et Céllulaire (IGBMC),
Illkirch, France
ABSTRACT The transcription-intermediary-factor-2 coid hypersecretion can eventually lead to pathological
(TIF-2) is a coactivator of the glucocorticoid receptor states (1, 2). Curtailment of the endocrine response to
(GR), and its disruption would be expected to influence stress is executed by glucocorticoid receptor (GR)-
glucocorticoid-mediated control of the hypothalamo- mediated negative feedback mechanisms in the hip-
pituitary-adrenal (HPA) axis. Here, we show that its pocampus, hypothalamus, and pituitary, manifested as
targeted deletion in mice is associated with altered reduced synthesis and secretion of pituitary ACTH and
expression of several glucocorticoid-dependent compo- its hypothalamic secretagogues, corticotropin-releasing
nents of HPA regulation (e.g., corticotropin-releasing hormone (CRH), and vasopressin (AVP) (1–3).
hormone, vasopressin, ACTH, glucocorticoid recep- GR signaling efficacy is determined by a variety of
tors), suggestive of hyperactivity under basal condi- coactivators, including the transcription intermediary
tions. At the same time, TIF-2⫺/⫺ mice display signifi- factor 2 (TIF-2/SRC-2/NCoA2/GRIP1). TIF-2 belongs
cantly lower basal corticosterone levels and a sluggish to the p160 family of cellular proteins (e.g., SRC-1 and
and blunted initial secretory response to brief emo- SRC-3), which contain a highly conserved motif (the
tional and prolonged physical stress. Subsequent anal- so-called NR-box) that facilitates interaction with the
ysis revealed this discrepancy to result from pro- ligand-dependent activation function (AF-2) of nuclear
nounced aberrations in the structure and function of receptors and amplification of target gene transcrip-
the adrenal gland, including the cytoarchitectural orga- tion (4). Interactions between TIF-2 (5), and its mouse
nization of the zona fasciculata and basal and stress- homologue GRIP1 (6), have been demonstrated for
induced expression of key elements of steroid hor- several nuclear receptors (7).
mone synthesis, such as the steroidogenic acute In the mouse, TIF-2 mRNA and protein are ex-
regulatory (StAR) protein and 3-hydroxysteroid dehy- pressed ubiquitously (5, 8, 9), but reports differ regard-
drogenase (3-HSD). In addition, altered expression ing its presence in the rat brain (10, 11). Targeted
levels of two nuclear receptors, DAX-1 and steroido- ablation of TIF-2 in the mouse is phenotypically marked
genic factor 1 (SF-1), in the adrenal cortex strengthen by impaired gonadal function (12) and altered energy
the view that TIF-2 deletion disrupts adrenocortical balance and lipid metabolism (13). Since no other
development and steroid biosynthesis. Thus, hyperac- overt functional aberrations have been so far noted in
tivity of the hypothalamo-pituitary unit is ascribed to TIF-2⫺/⫺ mice, compensatory overexpression of the
insidious adrenal insufficiency and impaired glucocor- related coactivator SRC-1, implying partial functional
ticoid feedback.—Patchev, A. V., Fischer, D., Wolf, redundancy of TIF-2 and SRC-1, has been presumed
S. S., Götz, F., Gehin, M., Chambon, P., Patchev, V. K., (14 –16).
and Almeida O. F. X. Insidious adrenocortical insuffi- Given the role of TIF-2 as a GR coactivator, we tested
ciency underlies neuroendocrine dysregulation in TIF-2 the hypothesis that disruption of TIF-2 would result in
deficient mice. FASEB J. 21, 231–238 (2007) attenuated GR signaling in the brain, measurable as
alterations in the secretory dynamics of the HPA axis
Key Words: stress 䡠 adrenal cortex 䡠 steroidogenesis 䡠 hypothal- under basal (resting) conditions and following expo-
amus 䡠 hippocampus
1
Correspondence: Institute of Experimental Endocrinol-
Stressors evoke neurohumoral responses within the ogy, Charité School of Medicine, Humboldt University, Schu-
hypothalamic-pituitary-adrenal (HPA) axis, the magni- mannstr. 20/21, Berlin 10117, Germany. E-mail: alexandre.
tude and duration of which reliably describe the organ- patchev@charite.de
ism’s adaptive potential. Chronic stress and glucocorti- doi: 10.1096/fj.06-6952com
0892-6638/07/0021-0231 © FASEB 231sure to stressful stimuli. We expected that TIF-2 defi- Hormone assays
ciency would become manifest as a “weakening of the
grip” of GR-mediated restraint of the HPA axis, result- Serum CORT was measured by RIA (MP Biomedicals, Or-
ing in pituitary-adrenal hyperactivity under quiescent angeburg, NY, USA). Pituitary ACTH contents were deter-
conditions, exaggerated and prolonged secretory re- mined by RIA (DSL, Webster, TX, USA) in 0.1 N acetic acid
extracts, and results were normalized to tissue protein levels;
sponse to stress, and increased expression of neuropep- the antibody (Ab) had 100% cross-reaction for the species-
tides that trigger HPA activation, e.g., CRH and AVP. As invariable, bioactive amino acid fragment ACTH (1–24).
reported here, our starting hypothesis was supported in Coefficients of variation of the CORT and ACTH assays were
several respects; however one unexpected but impor- 12 and 10%, respectively.
tant finding was that TIF-2⫺/⫺ mice demonstrate symp-
toms of adrenal insufficiency. The latter result suggests Nucleic acid probes and primers
that impaired HPA axis regulation in TIF-2 deficient
mice may result from insidious adrenocortical hypo- AVP transcripts were detected with a 48-mer antisense probe
function rather than failure to amplify GR signaling. (17) complementary to bases 1493–1540 of the murine vaso-
pressin gene (M_88345). The 40-mer probe for 3-HSD
mRNA corresponded to bases 531–570 of the murine gene
sequence (M_77015), with one mismatch to a rat-specific
MATERIALS AND METHODS probe (18). Oligonucleotides were labeled with [35S]-dATP
(NEN, Boston, MA, USA) using terminal deoxynucleotidyl
Animals transferase (TdT) (Invitrogen, Karlsruhe, Germany). Labeled
ribonucleotide probes for the detection of CRH-, GR-, SRC-1,
Animals were generated from a TIF-2⫹/⫺ male on a and StAR protein-encoding transcripts were produced from
C57BL/6N background (Charles River, Germany); five first- linearized plasmids using in vitro-transcription kits with T7,
generation pairs of TIF-2⫹/⫺ mice were used for crossbreed- T3, and Sp6 RNA polymerases (Promega, Madison, WI, USA)
ing. Randomly cycling females, aged 6 –7 mo, from the and [35S]-dUTP. The CRH cDNA template was a 760 bp
second generation of null mutants were used in this study, fragment comprising bases 260 –1020 of the mouse CRH gene
with wild-type (WT) C57BL/6N age-matched females serving (AY_128673). The GR template included bases 81–528 of the
as controls. Mutant mice were genotyped as described previ- rat GR mRNA sequence (M_14053). A 622 bp fragment
ously (12). Polymerase chain reaction (PCR) was done using corresponding to bases 1121–1743 of the human SRC-1
the primers P1 [CTG CAC GGT GCC automatic gain control (U_90661) was used for SRC-1 mRNA detection. The probe
(AGC) AAA GC), P2 (GAC CAG GGC TTG CTC AGA AC) for StAR protein transcripts was amplified by PCR from the
and P3 (CCC CTG GAT TGT TCC AAA GG]. The WT and clone IRAKp961L18177Q (RZPD, Berlin, Germany) using the
⫺/⫺ alleles resulted in products of 512 and 298 bp, respec- primers 5⬘-ATGTTCCTCGCTACGTTCAAG-3⬘ and 5⬘-
tively. CCAAGCGAAACACCTTGCC-3⬘. The resulting fragment of
Animals were housed under standard conditions (24°C; 364 bp, corresponding to bases 57– 420 of the murine StAR
60 –70% relative humidity, 12h light/dark with lights on at protein sequence (NM_011485), was cloned into pTOPO
06:00; ad libitum access to chow pellets and water). All and, on sequence verification and linearization with HindIII
procedures complied with NIH guidelines on laboratory and XbaI, used for the generation of antisense and sense
animal use and were approved by the local ethical board. probes, respectively. The plasmid encoding 246 bp (bases
1914 –2160) of the 3⬘UTR of the mouse SF-1 gene
Stress protocol (NM_139051) was linearized with HindIII and EcoRI, for
transcription of antisense and sense probe, respectively.
Peak and nadir basal CORT levels were determined in tail
vein blood (collected within 30 s of removal from home cage) In situ hybridization histochemistry
at 18:00 and 06:00 h.
The first experiment, which examined the endocrine re- Transcripts encoding AVP and CRH were quantified in the
sponse to short-term acute stress, started immediately after rostral extension of the PVN (bregma – 0.75 to – 0.85); those
the morning blood collection. Stress procedures encom- encoding GR were measured in the dorsal hippocampus
passed transfer to the procedure room, placement into a glass (bregma –1.70 to –1.90) and the PVN (19). Expression of
container (diameter 15 cm), and exposure to low-frequency SRC-1 mRNA was examined in the hippocampus. Coronal
vibration (ca. 25 Hz, delivered by a Vortex) and loud music 10-m-thick brain cryosections were hybridized with oligo-
(80 –90 dB) for 1 min, before returning to the home cage. and ribonucleotide probes according to established protocols
Blood samples were taken to measure stress-induced (30 min) (20, 21). Autoradiographs were generated by film exposure
and recovery-phase (120 min) levels of CORT. One week (BioMax MR; Kodak, Rochester, NY, USA).
later, animals were randomly assigned to subgroups for tissue Semiquantitative densitometric analysis of brain autora-
collection. Between 08:00 and 11:00 h one subgroup was diograhs was performed using the NIH software Scion Image
killed immediately on removal from the home cage, and the Beta 4.2.0. AVP transcript signal intensity was measured in
other 30 min after exposure to the above-described stress fixed-size squares within the ventromedial compartment of
procedure. Tissue specimens were either snap-frozen or im- the PVN, centered over its parvocellular division. CRH tran-
mersion-fixed in PBS/p-formaldehyde for subsequent analy- script densities were evaluated within the entire PVN. GR
sis. hybridization signals were measured in the pyramidal cell
A different set of animals from the third cross-bred gener- layer of the CA1–2 fields of the hippocampus and in the PVN.
ation was used for examination of responses to prolonged SRC-1 mRNA transcript intensities were compared in the
physical stress. The mice were immobilized in plastic restrain- granule cell layer of dentate gyrus (22).
ers for 4 h between 08:00 and 12:00 h. Blood from tail vein Bilateral optical density (OD) measurements were made in
incisions was collected for CORT determination 15, 120, and two sections per animal. Individual averaged transmittance
240 min after the beginning of immobilization. levels were converted to specific radioactivity by third-order
232 Vol. 21 January 2007 The FASEB Journal PATCHEV ET AL.polynomial equations generated from coexposed 14C stan- significantly smaller (8.5⫾0.4 mg) than those of WT
dards (ARC; St. Louis, MO, USA). and heterozygous TIF-2⫹/⫺ (12.2⫾0.8 and 11.0⫾0.8
Detection of transcripts encoding SF-1, StAR protein, and mg, respectively). Adrenal and uterine weights in TIF-
3-HSD in the adrenal cortex was performed in 5 m sections
through the equatorial plane of paraffin-embedded glands. 2⫺/⫺ mice were lower but not significantly different
Following deparaffinization (xylene), sections were perme- from those observed in WT mice.
abilized, delipidated, hybridized, and stringently washed (40,
41). Slides were exposed to photographic emulsion (NTB; TIF-2 deletion is associated with increased neural
Kodak, Rochester, NY, USA) for 3 d and lightly counter- drive on the HPA axis
stained with cresyl violet.
The software package Image Pro Plus (Media Cybrnetics,
Silver Spring, MD, USA) was used for the quantification of Expression levels of hypothalamic CRH and AVP, and
transcripts in adrenal sections. In each section, two rectangu- pituitary ACTH content in TIF-2 deficient animals
lar frames of preset size were placed within the microscopic indicated hyperactivation of the hypothalamo-pituitary
image of zona fasciculata at a magnification of ⫻400. Identical unit. Under basal (stress-free) conditions, the levels of
illumination conditions and size thresholds for inclusion/ the mRNAs encoding CRH and AVP in the hypotha-
exclusion of silver grains were applied to all sections. The
number of signal-positive particles was subsequently related
lamic paraventricular nucleus (PVN) were significantly
to the cell counts measured in each frame. Four measure- elevated in TIF-2⫺/⫺ mice, as compared to WT and
ments were averaged to produce individual means. TIF-2⫹/⫺ (Fig. 1; F2,10⫽12.19 and 10.66; P⫽0.004 and
0.003, respectively). CRH and AVP mRNA transcript
Reverse-transcriptase polymerase chain reaction (RT-PCR) numbers were rapidly up-regulated by stress in WT and
detection of DAX-1 TIF-2⫹/⫺, but not TIF-2⫺/⫺, mice (Fig. 1; F2,11⫽5.71 and
4.48; P⫽0.02 and 0.035, respectively). Pituitary ACTH
RNA was isolated using the RNeasy Plus Mini Kit (Qiagen, content in TIF-2⫺/⫺ mice was significantly higher than
Hilden, Germany); integrity was confirmed with the Agilent that measured in WT and TIF-2⫹/⫺ animals (Fig. 2;
System (Agilent Technologies, Palo Alto, CA, USA), accord- F2,10⫽8.55; P⫽0.007). Stress exposure failed to signifi-
ing to the manufacturer’s protocol. From each individual
extract, 500 ng were reverse-transcribed into cDNA using the cantly change pituitary ACTH content in any genotype.
Superscript III Platinum Kit (Invitrogen, Karlsruhe, Ger- Genotype-associated changes in the expression of GR
many). For real-time PCR, cDNA (50 ng) was amplified using in the brain complemented the symptoms of HPA
the TaqMan Universal PCR Master Mix (Applied Biosystems, hyperactivity. Significantly decreased GR mRNA levels
Foster City, CA, USA) according to the protocol of the were observed in the CA1–2 subfield of the hippocam-
supplier. The oligonucleotide probe sets for mouse DAX-1 pus of homo- and heterozygous TIF-2 deficient mice
(Mm 00431729_m1) and -actin (Mm 00607939_s1) were
(Fig. 3; F2,8⫽58.47; P⬍0.001), and a similar, albeit
purchased from Applied Biosystems. Normalization to -actin
and calculation of relative DAX-1 expression was done by the nonsignificant, trend was seen in the hypothalamic
⌬Ct method (Applied Biosystems). PVN. Expression of SRC-1 mRNA was seen throughout
the hippocampal formation, without any significant
Adrenocortical morphometry genotype-related differences (Fig. 3).
Cellular densities in the adrenocortical zona fasciculata were
determined in hematoxylin-eosin-stained paraffin sections
through the equatorial plane. Cell nuclei were counted using
the Image Pro Plus software within fixed-size rectangular
frames of 120 ⫻ 120 m, positioned in two diametrically
opposed sites of zona fasciculata. The threshold discrimination
tool was uniformly applied to ensure recognition of only
nuclear staining as positive signal. For each animal, counts
from four frames placed at the opposite poles two sections
were averaged.
Statistics
Numerical data (shown as mean ⫾ sem) were analyzed by
1-way ANOVA and Tukey’s test, where appropriate. Key
ANOVA parameters (F-values, degrees of freedom) are given
in the description of results. The level of significance was
preset at P ⬍ 0.05.
Figure 1. Altered expression of HPA axis-related neuropep-
RESULTS tides in TIF-2 mutant mice. Note that although CRH and AVP
expression was increased in the hypothalamic PVN of TIF-2
null mutants, the expression of the mRNAs encoding these
No conspicuous gross phenotypic features were ob- peptides was not influenced by exposure to stress. Asterisks
served in TIF-2⫺/⫺ mice (up to 1 yr old), except that the indicate significant differences as compared to WT mice;
males have easily discernible reduced testicular vol- daggers denote significant changes 30 min after stress; n ⫽
umes. At autopsy, the ovaries of TIF-2⫺/⫺ mice were 3–5 mice/group.
TIF-2 DISRUPTION AND PITUITARY-ADRENAL FUNCTION 233Figure 2. TIF-2 deletion results in increased ACTH produc- Figure 4. Corticosteroid secretion is impaired in TIF-2 mu-
tion. Pituitary ACTH content is elevated in TIF-2⫺/⫺ animals; tants. TIF-2 deficiency is associated with decreased basal
stress-induced changes were not observed in any genotype. corticosterone levels and attenuated early secretory responses
Asterisks mark significant differences to WT mice; n ⫽ 3–5 to both short-lasting (n⫽7 mice/group) and prolonged (n⫽6
mice/group. mice/group) stress. Asterisks indicate significant differences
from levels measured in WT mice at the corresponding time
point.
Impaired adrenocortical output in TIF2ⴚ/ⴚ mice
Surprisingly, basal (morning) serum corticosterone significantly attenuated in TIF-2⫺/⫺ animals, as com-
(CORT) levels in TIF-2⫺/⫺ mice were significantly lower pared to that in TIF-2⫹/⫺ and WT mice (Fig. 4;
than those in TIF-2⫹/⫺ and WT animals (Fig. 4; F2,22⫽17.35; P⬍0.001). Although similar trends were
F2,22⫽5.82, P⫽0.01). At the diurnal zenith of adreno- observed in CORT levels measured after 120 min, they
cortical activity CORT secretion was similar in all were not statistically significant.
groups (140⫾22, 139⫾21 and 137⫾20 ng/ml). Expo- An independent experiment corroborated the obser-
sure to a brief emotional stress significantly increased vation of decreased adrenocortical output in TIF-2-
CORT levels over baseline in all genotypes; however, deficiency during the initial phase of the stress re-
the secretory response measured 30 min post-stress was sponse. CORT levels, measured during protracted
immobilization stress for 4 h, were significantly lower in
TIF-2 ⫺/⫺ and heterozygous animals at 15 min (Fig. 4;
F2,12⫽8.90; P⬍0.004), but these differences between
genotypes became less pronounced with increasing
stress duration. As before, resting (prestress) CORT
levels were significantly lower in TIF-2 deficient mice
(F2,12⫽4.35; P⬍0.04).
Cellular basis for diminished adrenocortical activity
in TIF2ⴚ/ⴚ mice
The discrepancy between signs of increased hypo-
thalamo-pituitary stimulation and modest adrenocorti-
cal secretory output prompted us to focus on the
adrenal gland. Morphological examination revealed
that homo- and heterozygous TIF-2 deficient mice
display reduced cell densities in the zona fasciculata of
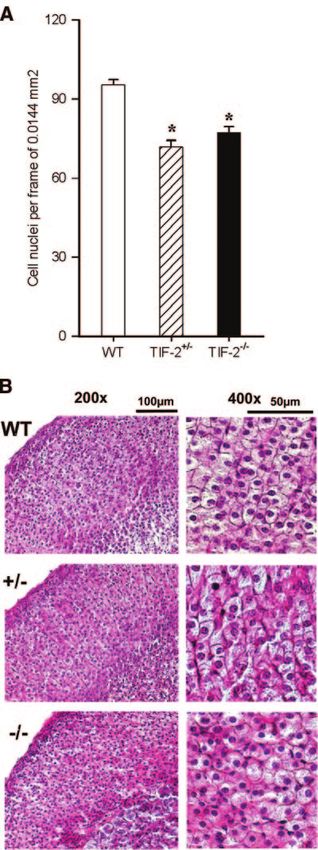
the cortex (Fig. 5; F2,24⫽30.09, P⬍0.001); these animals
Figure 3. Reduced expression of glucocorticoid receptors in showed a loss of the usual columnar organization of
the brain of TIF-2 mutants. Basal levels of GR mRNA in the cells and an abundance of hypertrophic cells and cells
hippocampal area CA1–2 are decreased in TIF2⫹/⫺ and with pyknotic nuclei.
TIF-2⫺/⫺ animals, with a similar pattern being observed in the We next studied several functional descriptors of the
hypothalamic PVN. Lack of significant differences in SRC-1
mRNA in the dentate gyrus of the hippocampus failed to
adrenal cortex. Adrenal DAX-1 mRNA levels (mea-
support the notion of compensatory overexpression in TIF- sured by semiquantitative RT-PCR) were significantly
2⫺/⫺ mice. Asterisks denote significant differences vs. WT higher in TIF-2⫺/⫺ vs. WT animals (Fig. 6; P⬍0.009). In
mice; n ⫽ 3–5 mice/group. contrast, steady-state levels of mRNA coding for the
234 Vol. 21 January 2007 The FASEB Journal PATCHEV ET AL.Figure 6. TIF-2 deletion is associated with altered adrenal
expression of DAX-1 and SF-1. Increased expression of DAX-1
and decreased abundance of SF-1 in the adrenals of mice
homozygous for TIF-2 deficiency. Asterisks denote significant
differences to WT; n ⫽ 3–5 per group.
over, these animals showed a markedly attenuated
increase in this parameter following exposure to
prolonged immobilization stress (Fig. 7; F5,15⫽22.24;
P⬍0.001). Similarly, the basal and stress-induced
expression of 3-hydroxysteroid dehydrogenase
(3-HSD) in TIF-2⫺/⫺ mice was significantly lower
than that in WT mice (Fig. 7; F5,18⫽28.06; P⬍0.001).
DISCUSSION
TIF-2 has been shown to amplify GR signaling in a
variety of cellular models and contexts (8), but its
physiological relevance in GR-mediated regulation of
Figure 5. Aberrant adrenocortical histology in TIF-2 ablated
mice. Disruption of TIF-2 is associated with decreased cell
density and severe cytoachitectural alterations in the zona
fasciculata of the adrenal cortex. Asterisks indicate significant
differences to WT; n ⫽ 4 –5 per group.
orphan nuclear receptor SF-1 were slightly but signifi-
cantly decreased in TIF-2⫺/⫺ vs. WT mice (Fig. 6;
P⬍0.04). Neither one of these parameters was altered
30 min after stress (data not shown).
Basal mRNA levels of steroidogenic acute regulatory Figure 7. Expression of StAR protein and 3-HSD is compro-
mised by TIF-2 ablation. Induction of StAR protein and
(StAR) protein in the adrenal cortex were similar in 3-HSD expression in the adrenal zona fasciculata is attenu-
WT and TIF-2⫺/⫺ genotypes. Notably, however, TIF- ated in TIF-2 deficient mice following either brief or pro-
2⫺/⫺ mice failed to show an up-regulation of StAR longed stress. Asterisks show significant differences to WT at
protein mRNA levels 30 min after acute stress; more- corresponding timepoints; n ⫽ 3– 6 per time point.
TIF-2 DISRUPTION AND PITUITARY-ADRENAL FUNCTION 235the HPA axis has not been addressed. Thus, experi- 16), despite recent evidence that SRC-1 is involved in ments were designed to test the hypothesis that abla- HPA axis regulation (25). Thus, as the importance of tion of TIF-2 would disrupt the efficacy of GR-mediated coregulators in the control of nuclear receptor expres- restraint on basal and stress-induced activity at various sion has not been comprehensively elucidated, we regulatory levels. considered it prudent to assume that TIF-2 may be While TIF-2⫹/⫺ mice showed profiles that largely involved in the control of GR expression through resembled those found in WT mice, gene dosage mechanisms that are independent of its role as a proved an important factor in several respects. Under nuclear receptor coregulator. stress-free conditions, hypothalamic levels of the mR- Deficits in the endocrine response to stress could also NAs encoding CRH and AVP, the two major cortico- originate from impaired hypothalamo-pituitary neuro- tropin secretagogues, were elevated in TIF-2⫺/⫺ mice. humoral communication. Unlike WT and TIF-2⫹/⫺ Predictably, pituitary reserves of ACTH were also signif- mice, CRH and AVP expression in TIF-2 null mutants icantly increased in TIF-2⫺/⫺ animals. Given that the was refractory to brief stressful stimuli. Accordingly, the CRH and AVP genes, as well as the POMC gene (whose inability of TIF-2⫺/⫺ mice to up-regulate CRH and AVP posttranslational products give rise to ACTH), are all expression in response to a stressor may be ascribed to subject to negative regulation by GR activation, these either limited synthetic capacity of the (already hyper- results indicate that deletion of TIF-2 leads to disrup- active) hypothalamic neurons or autologous desensiti- tion of GR negative feedback at the hypothalamic and zation of pituitary CRH and AVP receptors (26, 27), pituitary levels and, ultimately, hyperactivity of the ultimately resulting in impaired translation of the pep- central compartment of the HPA axis. Reduced GR tidergic stimuli into adequate pituitary-adrenal secre- mRNA expression in the hippocampus and PVN, both tory responses. This aspect of the neuroendocrine of which are crucially involved in the central control of response of TIF-2⫺/⫺ mice resembles the situation the HPA axis, would also be expected to contribute to observed in adrenalectomized animals; the latter dis- reduced restraint of HPA activity in TIF-2⫺/⫺ mice. play increased basal and blunted stress-induced expres- Increased basal adrenocortical activity with flattened sion of ACTH, CRH, and AVP (28 –30). diurnal oscillations, and exaggerated and prolonged In light of the various above-mentioned findings that response to stress would be the expected manifestations suggested deficits at the distal end of the HPA cascade, of impaired glucocorticoid negative feedback in the we investigated some key aspects of adrenocortical brain and pituitary of TIF-2⫺/⫺ mice. However, none of morphology and function. Previous studies described these predictions was fulfilled: despite compelling evi- anomalous gonadal developmental and impaired re- dence for hyperactivation of the hypothalamo-pituitary productive function in TIF-2 deficient mice (12, 15). In unit, in TIF-2 deficient mice: i) the amplitude of the the present study, we observed that TIF-2 null mutants nocturnal increase in CORT was commensurate with tend to have smaller adrenal glands, hinting at possible that seen in WT animals; ii) glucocorticoid levels at defects in adrenal organogenesis. Both TIF-2⫺/⫺ and their diurnal nadir were significantly lower than in TIF-2⫹/⫺ mice were found to have structural abnormal- other genotypes; and iii) unlike the situation in WT and ities in the glucorticoid-synthesizing zona fasciculata of heterozygous TIF-2 mutants, the CORT response to the adrenal cortex, namely, reduced cell densities and stress in TIF-2⫺/⫺ mice was markedly attenuated during loss of the columnar organization. These characteris- the initial phase and generally showed sluggish dynam- tics, together with the sporadic occurrence of pyknotic ics. Thus, the most striking feature of aberrant HPA cells in the zona fasciculata, are reminiscent of the axis regulation in TIF-2 deficient animals is the disso- histological changes described in congenital adrenal ciation between hyperactivity of the hypothalamo-pitu- hypoplasia (31). The view that adrenocortical develop- itary compartment and symptoms of inadequate adre- ment and function may be disrupted in TIF-2 null nocortical secretory output. Accordingly, alterations in mutants was bolstered by our observations of high levels the central mechanisms of HPA axis regulation, al- of DAX-1 expression in the adrenal cortex and, con- though indicative of decreased glucocorticoid restraint, comitantly, reduced abundance of SF-1. These two cannot be solely ascribed to the lack of TIF-2, a major transcription factors have been shown to act in a amplifier of GR transcriptional signaling (5–9). reciprocal manner in the coordination of adrenocorti- The patterns of GR expression in the hippocampus cal growth, maturation and steroidogenesis (32–34); of TIF-2⫺/⫺ mice illustrate the confounds associated their expression profiles in TIF-2⫺/⫺ mice indicate that with interpretation of our studies in terms of TIF-2 as a aberrant adrenocortical development belies the re- GR coactivator only. Based on the prevailing view (23, duced steroidogenic capacity in these animals. 24), homologous down-regulation by CORT excess Since TIF-2 can coactivate SF-1 (35, 36), it seems would be the most plausible explanation of decreased plausible that disruption of TIF-2 affects SF-1 depen- GR expression in the hippocampus. However, this is dent gene transcription which, in turn, impairs cortico- hardly tenable in view of the absence of symptoms of steroidogenesis. Indeed, StAR protein and 3-HSD, hypercorticalism in TIF-2 deficient mice under either which are positively regulated by SF-1 and crucially basal or stressful conditions. Our data also do not involved in glucocorticoid synthesis (37, 38), were support previous suggestions that another GR coactiva- poorly induced in the TIF-2 null mutants. Interestingly, tor, SRC-1, may compensate for TIF-2 deficiency (14 – TIF-2 null mutants and SF-1-haploinsufficient mice 236 Vol. 21 January 2007 The FASEB Journal PATCHEV ET AL.
show highly similar phenotypes in terms of adrenal teracting protein 1 in murine tissues using two novel polyclonal
histology and stress responses (39). antibodies. Eur. J. Endocrinol. 145, 323–333
10. Meijer, O.C., Steenbergen, P. J., and de Kloet, E. R. (2000)
In summary, our data demonstrate that signs of a Differential expression and regional distribution of steroid
“loss of grip” of glucocorticoid feedback in the HPA receptor coactivators SRC-1 and SRC-2 in brain and pituitary.
axis in TIF-2 deficient mice are not likely to be solely Endocrinology 141, 2192–2199
11. Martinez de Arrieta, C., Koibuchi, N., and Chin, W. W. (2000)
ascribed to disrupted GR signal amplification. Prelimi- Coactivator and corepressor gene expression in the rat cerebel-
nary data from ongoing experiments addressing the lum during postnatal development and the effect of altered
responsiveness of TIF-2 deficient mice to exogenous thyroid status. Endocrinology 141, 1693–1698
glucocorticoids are not indicative of glucocorticoid 12. Gehin, M., Mark, M., Dennefeld, C., Dierich, A., Gronemeyer,
H., and Chambon, P. (2002) The function of TIF2/GRIP1 in
resistance in these animals. Symptoms of structural and mouse reproduction is distinct from those of SRC-1 and p/CIP.
functional impairment in the adrenal cortex of TIF-2 Mol. Cell. Biology. 22, 5923–5937
mutants suggest that alterations in the neural control of 13. Picard, F., Gehin, M., Annicotte, J., Rocchi, S., Champy, M.F.,
O’Malley, B.W., Chambon, P., and Auwerx, J. (2002) SRC-1 and
the HPA axis occur secondarily to insidious adrenocor- TIF2 control energy balance between white and brown adipose
tical insufficiency. In view of earlier and recent demon- tissues. Cell 111, 931–941
strations of GR expression in the fetal (40) and adult 14. Xu, J., Qiu, Y., DeMayo, F.J., Tsai, S.Y., Tsai, M. J., and O’Malley,
adrenal cortex (41), and potentiation of corticotropin B. W. (1998) Partial hormone resistance in mice with disruption
of the steroid receptor coactivator-1 (SRC-1) gene. Science 279,
effects in adrenocortical cells by glucocorticoids (42), 1922–1925
the intriguing question opened by these observations is 15. Xu, J., and Li, Q. (2003) Review of the in vivo functions of the
whether the lack of TIF-2, which results in poor ampli- p160 steroid receptor coactivator family. Mol. Endocrinol. 17,
fication of trophic GR signaling, may be responsible for 1681–1692
16. Mark, M., Yoshida-Komiya, H., Gehin, M., Liao, L., Tsai, M.J.,
compromised adrenocortical development and steroi- O’Malley, B.W., Chambon, P., and Xu, J. (2004) Partially
dogenic capacity. Meanwhile, the association between redundant functions of SRC-1 and TIF2 in postnatal survival and
TIF-2 deletion and adrenal dysfunction (this study) and male reproduction. Proc. Natl. Acad. Sci. U. S. A. 101, 4453– 4458
17. Nishi, R., Ueta, Y., Serino, R., Nomura, M., Yamamoto, Y.,
hypogonadism (12, 15) strongly suggest a novel role for Shibuya, I., Koizumi, K., and Yamashita, H. (1999) Increase of
this nuclear receptor coregulator in the morphogenesis vasopressin mRNA in the hypothalamus of inbred polydipsic
and function of steroid-producing glands. mice. Brain Res. Bull. 50, 47–51
18. Coirini, H., Gouézou, M., Liere, P., Delespierre, B., Pianos, A.,
Eychenne, B., Schumacher, M., and Guennoun, R. (2002)
This study was supported by an EC (FP6) collaborative
3-Hydroxasteroid dehydrogenase expression in rat spinal cord.
grant (QLG3-CT-2000 – 00844). We appreciate the technical Neuroscience 113, 883– 891
assistance of A. Fishbach, H. Lück, S. Thalheim, S. Herrmann, 19. Franklin, K.B.J., and Paxinos, G. (1997) The Mouse Brain in
and J. Waldherr. The generous provision of expression plas- Stereotaxic Coordinates. Academic Press, San Diego, CA, U. S. A.
mids by Drs. J. Herman (Cincinnati, OH, USA), K. Yamamoto 20. Brady, L.S., Smith, M., Gold, P. W., and Herkenham, M. (1990)
(San Francisco, CA, USA), M.-J. Tsai (Houston, TX, USA) and Altered expression of hypothalamic neuropeptide mRNA in
K. Parker (Dallas, TX, USA) is gratefully acknowledged. food-restricted and food-deprived rats. Neuroendocrinology 52,
441– 447
21. Whitfield, H.J. jr., Brady, L.S., Smith, M.A., Mamalaki, E., Fox,
R., and Herkenham, M. (1990) Optimization of cRNA probe in
situ hybridization methodology for localization of glucocorti-
REFERENCES coid receptor mRNA in rat brain: A detailed protocol. Cell. Mol.
Neurobiol. 10, 145–157
1. Chrousos, G. P., and Gold, P. W. (1992) The concept of stress 22. Mitev, Y.A., Wolf, S.S., Almeida, O. F. X., and Patchev, V. K.
and stress system disorders. Overview of physiological and (2003) Developmental expression profiles and distinct regional
behavioral homeostasis. JAMA 267, 1244 –1252 estrogen responsiveness suggest a novel role for the steroid
2. Sapolsky, R.M., Romero, L. M., and Munck, A. U. (2000) How receptor coactivator SRC-1 as discriminative amplifier of estro-
do glucocorticoids influence stress responses? Integrating per- gen signaling in the rat brain. FASEB J. 17, 518 –519
missive, suppressive, stimulatory and preparative actions. Endocr. 23. Patchev, V.K., Brady, L.S., Karl, M., and Chrousos, G. P. (1994)
Rev. 21, 55– 89 Regulation of HSP90 and corticosteroid receptor mRNA by
3. De Kloet, E.R., Joels, M., and Holsboer, F. (2005) Stress and the corticosterone levels in vivo. Mol. Cell. Endocrinol. 103, 57– 64
brain: from adaptation to disease. Nat. Rev. Neurosci. 6, 463– 475 24. Herman, J. P., and Spencer, R. (1998) Regulation of hippocam-
4. Glass, C. K., and Rosenfeld, M. G. (2000) The coregulator pal glucocorticoid receptor gene transcription and protein
exchange in transcriptional functions of nuclear receptors. expression in vivo. J. Neurosci. 18, 7462–7473
Genes Dev. 14, 121–141 25. Winnay, J.N., Xu, J., O’Malley, B. W., and Hammer, G. D. (2006)
5. Voegel, J.J., Heine, M.J., Zechel, C., Chambon, P., and Grone- Steroid receptor coactivator-1-deficient mice exhibit altered
meyer, H. (1996) TIF2, a 160 kDa transcriptional mediator for hypothalamic-pituitary-adrenal axis function. Endocrinology 147,
the ligand-dependent activation function AF-2 of nuclear recep- 1322–1332
tors. EMBO J. 15, 3667–3675 26. Aguilera, G., and Rabadan-Diehl, C. (2000) Regulation of
6. Hong, H., Kohli, K., Trivedi, A., Johnson, D. L., and Stallcup, vasopressin V1b receptors in the anterior pituitary gland of the
M. R. (1996) GRIP1, a novel mouse protein that serves as a rat. Exp. Physiol. 85, 19S–26S
transcriptional coactivator in yeast for the hormone binding 27. Aguilera, G., Rabadan-Diehl, C., and Nikodemova, M. (2001)
domains of steroid receptors. Proc. Natl. Acad. Sci. U. S. A. 93, Regulation of pituitary corticotropin releasing hormone recep-
4948 – 4952 tors. Peptides 22, 769 –774
7. Leo, C., and Chen, J. D. (2000) The SRC family of nuclear 28. Akana, S. F., and Dallman, M. F. (1997) Chronic cold in
receptor coactivators. Gene 245, 1–11 adrenalectomized, corticosterone (B)-treated rats: Facilitated
8. Hong, H., Kohli, K., Garabedian, M. J., and Stallcup, M. R. corticotropin responses to acute restraint emerge as B increases.
(1997) GRIP1, a transcriptional coactivator for the AF-2 trans- Endocrinology 138, 3249 –3258
activation domain of steroid, thyroid, retinoid and vitamin D 29. Marti, O., Harbuz, M.S., Andrés, R., Lightman, S. L., and
receptors. Mol. Cell. Biology. 17, 2735–2744 Armario, A. (1999) Activation of the hypothalamic-pituitary axis
9. Puustinen, R., Sarvilinna, N., Manninen, T., Tuohimaa, P., and in adrenalectomised rats: potentiation by chronic stress. Brain
Ylikomi, T. (2001) Localization of glucocorticoid receptor in- Res. 821, 1–7
TIF-2 DISRUPTION AND PITUITARY-ADRENAL FUNCTION 23730. Weidenfeld, J., and Feldman, S. (2000) Effects of adrenalectomy 37. Parker, K. L., and Schimmer, B. P. (1997) Steroidogenic factor
and corticosterone replacement on the hypothalamic-pituitary 1: a key determinant of endocrine development and function.
response to neural stimuli. Brain Res. 877, 73–78 Endocr. Rev. 18, 361–377
31. Weiss, L., and Mellinger, R. C. (1970) Congenital adrenal 38. Sandhoff, T.W., Hales, D.B., Hales, K. H., and McLean, M. P.
hypoplasia—an X-linked disease. J. Med. Genet. 7, 27–32 (1998) Transcriptional regulation of the rat steroidogenic acute
32. Babu, P.S., Bavers, D.L., Beuschlein, F., Shah, S., Jeffs, B., regulatory protein gene by steroidogenic factor 1. Endocrinology
Jameson, J. L., and Hammer, G. D. (2002) Interaction between 139, 4820 – 4831
Dax-1 and steroidogenic factor-1 in vivo: increased adrenal 39. Bland, M., Jamieson, C.A.M., Akana, S.F., Bornstein, S.R., Eisen-
responsiveness to ACTH in the absence of Dax-1. Endocrinology hofer, G., Dallman, M. F., and Ingraham, H. A. (2000) Haplo-
143, 665– 673 insufficiency of steroidogenic factor-1 in mice disrupts adrenal
33. Lalli, E., and Sassone-Corsi, P. (2003) DAX-1, an unusual development leading to an impaired stress response. Proc. Natl.
orphan receptor at the crossroads of steroidogenic function and Acad. Sci. U. S. A. 97, 14488 –14493
sexual differentiation. Mol. Endocrinol. 17, 1445–1453 40. Condon, J., Gosden, C., Gardener, D., Nickson, P., Hewison, M.,
34. Hammer, G.D., Parker, K. L., and Schimmer, B. P. (2005) Howie, A. J., and Stewart, P. M. (1998) Expression of type 2
Transcriptional regulation of adrenocortical development. En- 11beta-hydroxysteroid dehydrogenase and corticosteroid hor-
docrinology 146, 1018 –1024 mone receptors in early human fetal life. J. Clin. Endocrinol.
35. Hammer, G.D., Krylova, I., Zhang, Y., Darimont, B.D., Simpson, Metab. 83, 4490 – 4497
K., Weigel, N. L., and Ingraham, H. A. (1999) Phosphorylation 41. Paust, H.J., Loeper, S., Else, T., Bamberger, A.M., Papadopou-
of the nuclear receptor SF-1 modulates cofactor recruitment: los, G., Pankoke, D., Saeger, W., and Bamberger, C. M. (2006)
Integration of hormone signaling in reproduction and stress. Expression of glucocorticoid receptor in the human adrenal
Molecular Cell. 3, 521–526 cortex. Exp. Clin. Endocrinol. Diabetes. 114, 6 –10
36. Børud, B., Hoang, T., Bakke, M., Jacob, A.L., Lund, J., and 42. Picard-Hagen, N., Penhoat, A., Hue, D., Jaillard, C., and Durand, P.
Mellgren, G. (2002) The nuclear receptor coactivators p300/ (1997) Glucocorticoids enhance corticotropin receptor mRNA
CBP/cointegrator-associated protein (p/CIP) and transcription levels in ovine adrenocortical cells. J. Mol. Endocrinol. 19, 29 –36
intermediary factor 2 (TIF2) differentially regulate PKA-stimu-
lated transcriptional activity of steroidogenic factor 1. Mol. Received for publication July 24, 2006.
Endocrinol. 16, 757–773 Accepted for publication August 21, 2006.
238 Vol. 21 January 2007 The FASEB Journal PATCHEV ET AL.You can also read

















































