THE R2R3 MYB TRANSCRIPTION FACTORS MAMYBF AND MAMYB1 REGULATE AVONOID BIOSYNTHESIS IN GRAPE HYACINTH
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The R2R3 MYB transcription factors MaMYBF and
MaMYB1 regulate flavonoid biosynthesis in grape
hyacinth
Jiangyu Wang
Northwest A&F University: Northwest Agriculture and Forestry University
Han Zhang
Northwest A&F University: Northwest Agriculture and Forestry University
Shuting Tian
Northwest A&F University: Northwest Agriculture and Forestry University
Wenhui Hao
Northwest A&F University: Northwest Agriculture and Forestry University
Kaili Chen
Southwest University
Lingjuan Du ( dulingjuan@nwafu.edu.cn )
Northwest A&F University: Northwest Agriculture and Forestry University https://orcid.org/0000-0003-
4295-3072
Research Article
Keywords: R2R3 MYB regulators, Flower color, Flavonoid, Grape hyacinth
Posted Date: April 15th, 2022
DOI: https://doi.org/10.21203/rs.3.rs-1491141/v1
License: This work is licensed under a Creative Commons Attribution 4.0 International License.
Read Full License
Page 1/26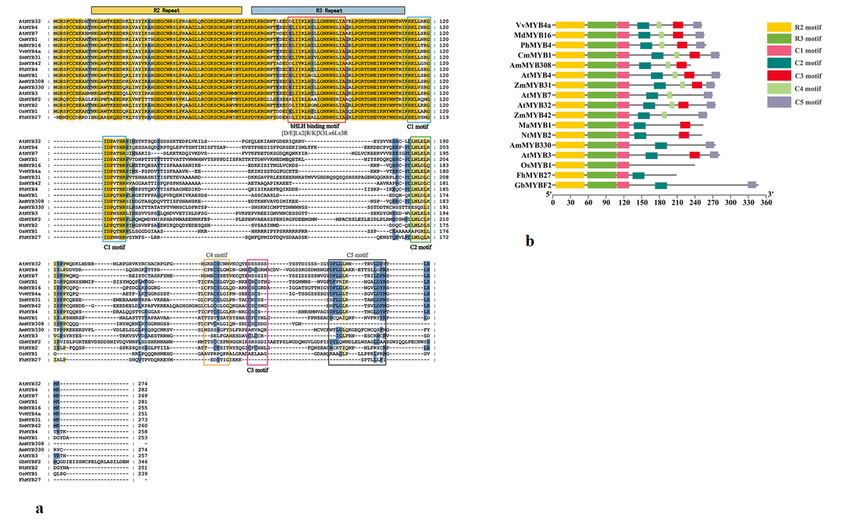
Abstract
R2R3 MYBs play vital roles in the regulation of flavonoid biosynthesis. However, the regulatory network of
R2R3 MYBs in flavonoid biosynthesis is not fully understood in grape hyacinth (Muscari spp.). Here, we
identified two R2R3 MYBs, MaMYBF and MaMYB1, as potential regulators of flavonol and anthocyanin
biosynthesis, respectively. MaMYBF and MaMYB1 expression was elevated during flower development
and was light-induced, and the expression patterns were correlated with those of the flavonoid structural
genes MaFLS and MaDFR, respectively. The BiFC assay confirmed that MaMYB1 interacts with
MabHLH1, but MaMYBF does not. A dual luciferase assay revealed that MaMYBF alone strongly
activated pMaFLS, and its activation was attenuated at reduced doses of MaMYBF in the presence of
MabHLH1, MaMybA, and MaMYB1. MaDFR transcription mediated by MaMybA and MabHLH1 was
inhibited by MaMYB1. Moreover, overexpression of MaMYBF and MaMYB1 in tobacco reduced flower
pigmentation and repressed the expression of flavonoid pathway key structure genes. Therefore,
MaMYBF regulates the flavonol pathway independently of cofactors, whereas MaMYB1 regulates
anthocyanin biosynthesis by binding to MabHLH1 and disrupting the MaMybA-bHLH complex in grape
hyacinth. Our results provide new insights into the intricate regulatory network of flavonoids in grape
hyacinth involving the regulation of both flavonol and anthocyanin.
Key Message
R2R3 MYB regulators, MaMYBF and MaMYB1 acts as flavonol-related acivator and anthocyanin-related
repressors, respectively. They are involved in the regulation of light-induced flavonoid synthesis in grape
hyacinth.
1 Introduction
Flavonoids are one of the most important groups of plant secondary metabolites and coloring
substances, exhibiting a wide range of application value. Flavonoids are divided into three main
categories: anthocyanin, flavonol, and proanthocyanidin (PA) (Williams and Grayer, 2001). Anthocyanin,
one of the most important pigments for flower coloration, is biosynthesized by a series of enzymes.
Anthocyanin contributes to the red, purple, blue–violet and blue colors in flowers and has a role in
facilitating pollination and seed dispersal. Flavonols, which are copigments for anthocyanin, affect the
hue of flowers and effectively protect plants against light, UV-B radiation and other environmental factors
(Czemmel et al. 2009; Takahashi et al. 2007). The flavonoid biosynthetic pathway isolated and
characterized to date is conserved in plants and consists of enzymatic steps leading to the different
flavonoid classes (Liu et al. 2019a, b; Shan et al. 2020). Anthocyanin and flavonol biosynthesis share
many enzymes in the flavonoid biosynthetic pathway. Studies have shown that flavonol synthase (FLS)
competes with dihydroflavonol 4-reductase (DFR) for dihydroflavonol as a substrate, competing to
regulate branches of different flavonoid biosynthesis pathways and affecting the final color of plants and
the production of flavonoids (Martens et al. 2002; Wellmann et al. 2002).
Page 2/26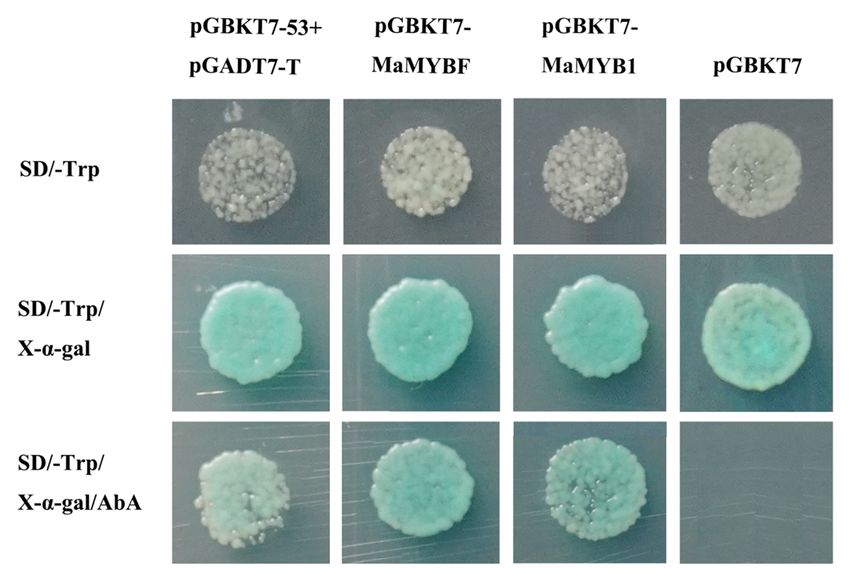
In plants, the transcription complex composed of MYB, bHLH, and WD40 (MBW) proteins regulates the
expression of multiple distinct target genes involved in various metabolic pathways (Andrew and Richard
2018; Ramsay and Glover 2005). The largest number of current MYB class transcription factors were
isolated in higher plants, of which the R2R3 MYB regulators can be categorized into at least 28
subgroups (SGs) (Stracke et al. 2001). An increasing number of flavonoid-regulating R2R3 MYBs have
been identified, and the gene homologs are generally conserved in the same regulatory pathway (Dubos
et al. 2010). SG4 MYB proteins tend to be negative regulators of phenylpropanoid-derived compound
synthesis and include the C2 repressor motif (Albert et al. 2014; Anwar et al. 2018; Dubos et al. 2010). For
example, PhMYB27 and FhMYB27 are part of the MBW complex and repress transcription through their
C2 motifs (Albert et al. 2014; Li et al. 2020). PhMYB27 also downregulates flavonoid metabolism by
interacting with the MBW complex via bHLH (Albert et al. 2014). Moreover, MdMYB16 can directly bind to
the promoters of MdANS and MdUFGT via the EAR motif to repress anthocyanin biosynthesis in Malus
domestica (Xu et al. 2017). SG5 MYBs, including AtTT2 and VvMYBPA2, are generally considered
activators of PA biosynthesis (Sharma and Dixon 2005; Terrier et al. 2009). Most SG6 MYBs are
anthocyanin activators, such as VvMYBA1/2 in grapevine (Vitis vinifera) and MaMybA and MaAN2 in
grape hyacinth, which specifically promote anthocyanin biosynthesis by regulating LBG expression (Chen
et al. 2019; Takos et al. 2006). SG7 MYB proteins are flavonol-specific regulators (Czemmel et al. 2009).
Grapevine VvMYBF1 (Czemmel et al. 2009), apple MdMYB22 (Wang et al. 2017), and Freesia hybrida
FhMYBFs (Shan et al. 2020) target the flavonol-specific pathway gene FLS to regulate flavonol
biosynthesis. Moreover, studies have found that VvMYBF1 regulates flavonol synthesis in response to UV-
B (Czemmel et al. 2009). Unlike R2R3 MYBs, R3 MYB repressors involved in flavonoid biosynthesis, such
as CAPRICE-like (CPC-like), lack repression motifs in the C-termini. The CPC-like homologs PhMYBx
(Albert et al. 2014), FhMYBx (Li et al. 2020) and MaMYBx (Zhang et al. 2020) were found to repress
anthocyanin biosynthesis by competing with the R2R3 MYB activators for binding to the bHLH cofactor.
However, the overall orchestration of these MYBs in the coordination of different branches of flavonoid
biosynthesis is not completely understood, especially in ornamental plants, which usually exhibit a
complex feedback regulation network. Thus, further studies on the MYB transcriptional regulators of
flavonoid biosynthesis in ornamental plants are crucial.
Grape hyacinth (Muscari spp.) is widely used in flower breeding and garden landscaping due to its unique
blue–purple flowers, and flavonoids are critical substances in its flower color. The grape hyacinth flower
coloring process is a genetically programmed and highly coordinated process that leads to structural and
biochemical changes, such as changes in the contents of anthocyanins, proanthocyanins, flavonols,
glycosylation, and acylation. Recently, functional analysis of MaFLS and MaDFR was performed, and
heterologous expression of MaFLS and MaDFR in tobacco (N. tabacum) resulted in increased flavonol
and anthocyanin accumulation, respectively, suggesting that MaFLS and MaDFR were involved in flower
coloration (Liu et al. 2019a, b). Thus far, we identified two anthocyanin-related R2R3 MYB activators
(MaAN2 and MaMybA) and an R3 MYB repressor (MaMYBx) in grape hyacinth (Chen et al. 2017, 2019;
Zhang et al. 2020). Ectopic expression of MaAN2, MaMybA, and MaMYBx in Nicotiana tabacum (N.
tabacum) resulted in obviously darker and lighter flower colors, respectively (Chen et al. 2017, 2019;
Page 3/26
Zhang et al. 2020). However, the feedback regulation mechanism of the R2R3 MYB transcription factor in
grape hyacinth flavonoid biosynthesis and the response adjustment mechanism of the environment
remain incomplete.
In this study, we characterized two R2R3 MYBs, the flavonol-specific TF MaMYBF and the anthocyanin
repressor MaMYB1 from grape hyacinth. We further elucidate the mechanism by which MYB TFs regulate
flavonoid synthesis in grape hyacinth and provide a theoretical basis to further enrich the understanding
of the complex flavonoid regulatory network in ornamental plants.
2 Materials And Methods
2.1 Plant materials and growth conditions
A grape hyacinth cultivar (M. aucheri ‘Dark Eyes’) was field cultivated at Northwest A&F University in
Yangling District, Shaanxi Province, China. The shading treatment of the inflorescence was completed
using bags. Unbagged inflorescences were used as controls. Five floral developmental stages (S1–S5)
were sampled as described by Lou et al. (2017): S1, closed buds, no pigmentation; S2, closed buds, initial
tepal pigmentation; S3, closed buds just before blooming; S4, open flowers; and S5, senescent flowers.
Fresh buds or flowers from each stage treated in the same condition were combined into one biological
replicate, and analyses were completed for at least three biological replicates. The collected samples
were immediately frozen in liquid nitrogen and stored at -80°C.
Tobacco plants (N. tabacum ‘SR-1’) were aseptically grown for approximately 4 weeks and prepared for
stable genetic transformation. Tobacco transformation was conducted using a previously described
protocol (Horsch et al. 1985). Wild-type (WT) and transgenic tobacco plants were transferred to a
greenhouse for approximately one month with a 16 h light:8 h dark photoperiod. The MaMYBF-
overexpressing (OE-MaMYBF) and MaMYB1-overexpressing (OE-MaMYB1) transgenic lines exhibiting
obvious color changes in flowers were used for further analysis
2.2 Gene and promoter isolation, sequence alignment, and
phylogenetic analysis
To identify flavonoid-related R2R3 MYBs in grape hyacinth, AtMYB12 and AtMYB4 were downloaded
from the National Center for Biotechnology Information (NCBI) GenBank database. Using the
homologous protein local BLASTP and gene function annotation, we obtained two R2R3 MYB unigenes
designated “MaMYBF” and “MaMYB1”. The Information of MaMYBF and MaMYB1 gene were described
in Table S1. The full-length MaMYBF and MaMYB1 sequences were PCR-amplified from genomic DNA
(gDNA) and cDNA from M. aucheri ‘Dark Eyes’, as described by Chen et al. (2017). The primers are listed
in Supplementary Table S2. The cDNA and gDNA sequences of MaMYBF and MaMYB1 were submitted
to the NCBI GenBank database (accession numbers: OM171266, OM171269, OM171267, and OM171268,
respectively). Amino acid sequences were submitted to CLUSTAL OMEGA
(http://www.ebi.ac.uk/Tools/msa/clustalo/) for multiple sequence alignment. For phylogenetic analysis,
Page 4/26
the alignments were further processed by MEGA 6.0 (Tamura et al. 2013). The maximum likelihood
method and 1000 bootstrap replicates were used to evaluate the reliability of the phylogenetic analysis.
2.3 Real-time quantitative PCR (qRT–PCR)
Total RNA extraction from the flowers, bulbs, and leaves of M. aucheri ‘Dark Eyes’ as well as from the
leaves and flowers of tobacco, underwent reverse transcription to synthesize cDNA, and qRT–PCR assays
were performed using a previously described protocol (Chen et al. 2017). MaActin and NtTubA1 were
used as internal control genes in each grape hyacinth and tobacco sample, respectively. Analysis was
performed using three biological and technical replicates. The qRT–PCR primers used are listed in
Supplementary Table S2.
2.4 Subcellular localization and bimolecular fluorescence
complementation (BiFC) assay
The 35S:MaMYBF:GFP and 35S:MaMYB1:GFP constructs were used to identify the subcellular
localization of MaMYBF and MaMYB1. Samples transformed with 35S:GFP were used as controls. To
verify whether MaMYBF and MaMYB1 interact with MabHLH1 in vivo, a BiFC assay was performed as
described by Zhang et al. (2020). The primers are listed in Supplementary Table S2. The green and yellow
fluorescent protein (GFP/YFP) fluorescence was visualized 18 h after transformation using a confocal
microscope (TCS SP8; Leica, Wetzlar, Germany), and images were analyzed using LAS X software.
2.5 Transcriptional activation ability of MaMYBF and
MaMYB1
To determine the transcriptional activation ability of MaMYBF and MaMYB1, a yeast expression vector
was constructed by fusing the MaMYBF and MaMYB1 ORF products into a pGBKT7 vector and
introduced into the yeast strain Y2Hgold. Yeast transformation and autoactivation testing were
performed as described by Chen et al. (2017).
2.6 Dual luciferase assay
For the dual luciferase assay, the full-length sequences MaMYBF and MaMYB1 and the promoters
pMaFLS and pMaDFR were introduced into the pGreenII 62-SK and pGreenII 0800-LUC vectors (Hellens et
al. 2005), respectively. The primers are listed in Supplementary Table S2. All constructs transformation
and the activities measurement of these promoters, luciferase (LUC) to 35S: Renilla (REN), were
performed as described by Zhang et al. (2020). Three biological replicates were evaluated per assay.
2.7 Measurement of flavonol and anthocyanin contents
Total flavonol and anthocyanin samples were extracted from five floral developmental stages (S1–S5) of
grape hyacinth and flowers of transgenic tobacco lines and quantified as previously described (Huang et
al. 2015; Shan et al. 2019). Flavonol and anthocyanin were separated, identified and quantified according
to the method described by Chen et al. (2017). Quantitative analysis of anthocyanins and flavonols was
Page 5/26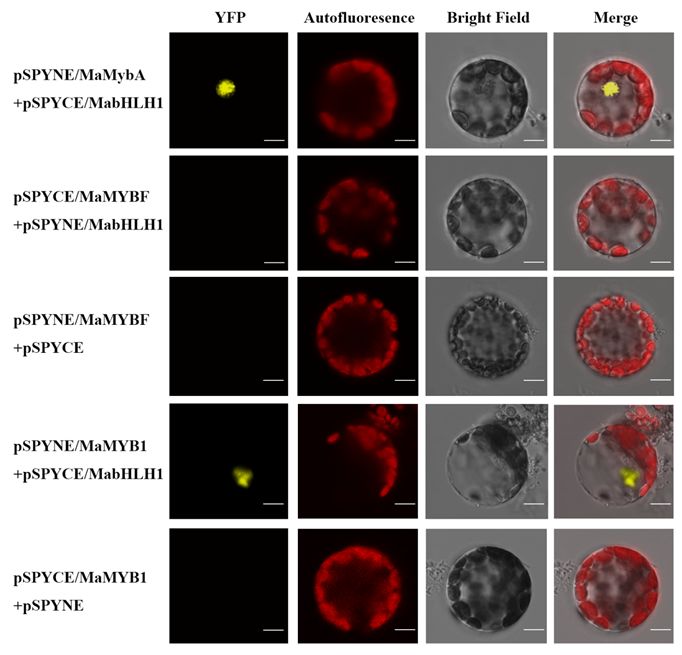
performed using the external standard curve calibration of cyanidin, quercetin, and kaempferol. The
anthocyanin content was quantified using the following formula:
[A530 − (0.25 × A657)] × Volume
TotalantℎocyanincontentArabidopsis =
FresℎWeigℎt
All samples were analyzed in three biological replicates, and three independent experiments were
performed separately.
2.8 Statistical analysis
Statistical analysis of the data was conducted using SPSS 20.0 software (SPSS, Chicago, IL, USA).
Significant differences were identified using Tukey's HSD test at P < 0.05.
3 Results
3.1 MaMYBF and MaMYB1 encode different R2R3 MYB
transcription factors
In this study, combining transcriptome-related data (Lou et al. 2014) from grape hyacinth flowers in the
early stage, through the homologous protein local BLASTP and gene function annotation, we obtained
two R2R3 MYB unigenes. They were cloned from M. aucheri 'Dark Eyes' flowers and named MaMYBF and
MaMYB1. MaMYBF had an ORF of 927 bp and encoded a predicted R2R3 MYB protein with 308 amino
acid residues (GenBank accession number: OM171266), while MaMYB1 had an ORF of 762 bp and
encoded a predicted R2R3 MYB protein with 253 amino acid residues (GenBank accession number:
OM171267).
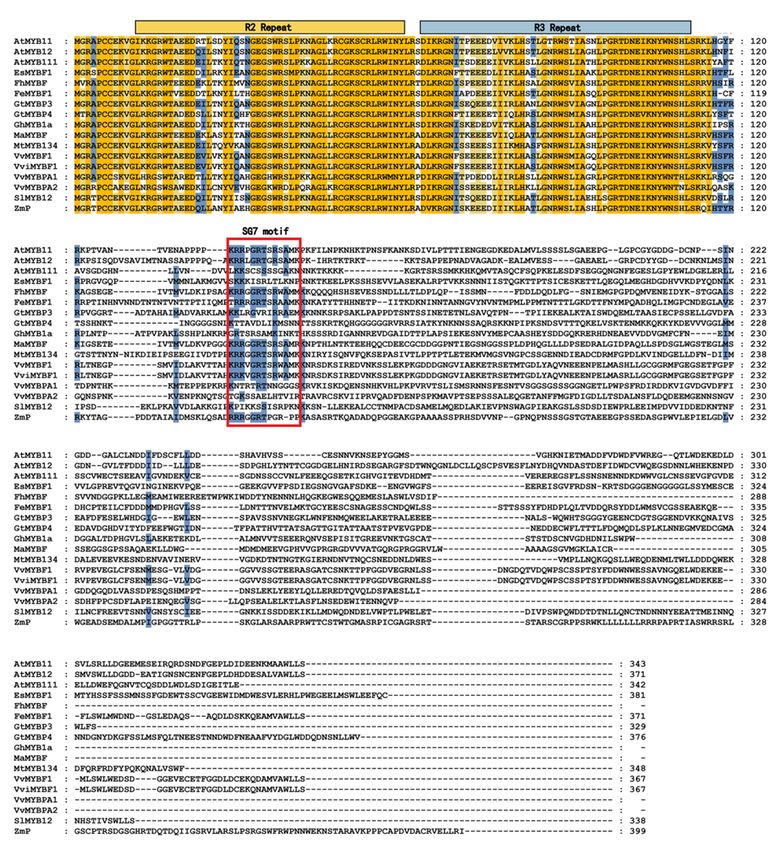
Sequence alignment analysis revealed that MaMYBF was defined by an N-terminal R2R3 MYB domain
and a signature motif SG7 ([K/R][R/x][R/K]xGRT[S/x][R/G]xx[M/x]K) (Fig. 1). Additionally, the R3 domain
of MaMYBF lacks the [D/E]Lx2[R/K]x3Lx6Lx3R motif necessary for interaction with the bHLH protein.
Another signature motif, SG7-2 ([W/x][L/x]LS), was detected at the C-terminus of AtMYB12, AtMYB11,
AtMYB111, and VvMYBF1, which was also absent in MaMYBF (Fig. 1). In contrast, MaMYB1 had the
[D/E]Lx2[R/K]x3Lx6Lx3R motif necessary for interaction with bHLH factors. The MaMYB1 C-terminus
contained additional conserved motifs, such as the C1-like motif (LlsrGIDPxT/SHRxI/L), C2-like motif
(pdLNLD/ELxiG/S), C3 ZnF (zink-finger) motif (CX1-2CX7-12CX2C), and LxLxL-type EAR repressor
domain (Fig. 2a, b). These conserved motifs belong to subgroup 4 (SG4) of R2R3 MYB transcription
factors (Jin et al. 2000, Sateesh and Kevin 2011). However, MaMYB1 lacks the C4 motif, unlike other
members of subgroup 4 R2R3 MYB proteins (Fig. 2b).
To better define MaMYBF and MaMYB1, phylogenetic analysis with respect to other MYB regulators was
performed. The phylogeny implied that a number of clades were resolved, implicating their different
functions (Fig. 3). MaMYBF grouped with the previously characterized flavonol regulator Freesia hybrida
FhMYBF and grape VvMYBF1, which belong to subgroup 7 (SG7). MaMYB1 clustered with other SG4
Page 6/26
R2R3 MYB transcription factors, such as PhMYB4, NtMYB2 and AtMYB32, which are involved in the
suppression of the flavonoid and lignin biosynthetic pathways (Fig. 3) (Sateesh and Kevin 2011). These
results indicate that MaMYBF and MaMYB1 might be candidate regulators of flavonoid biosynthesis.
3.2 MaMYBF and MaMYB1 are transcription factors
To verify the localization of the MaMYBF and MaMYB1 proteins, the 35S:MaMYBF:GFP and
35S:MaMYB1:GFP plasmids were transformed into Arabidopsis mesophyll protoplasts. GFP signals were
detected in the whole control transgenic cells expressing 35S:GFP. However, strong fluorescence from
35S:MaMYBF:GFP and 35S:MaMYB1:GFP was detected in the nucleus (Fig. 4), indicating that MaMYBF
and MaMYB1 are localized to the nucleus. To determine whether MaMYBF and MaMYB1 exhibit
transcriptional activity, we conducted a transactivation assay in yeast. Yeasts transformed with pGBKT7-
MaMYBF and vector pGBKT7-MaMYB1 and the positive control pGBKT7-53 plus pGADT7-T exhibited
blue yeast plaques, while the negative control pGBKT7 did not grow in SD/-Trp medium plus 60 µg/ml X-
α-Gal and 200 ng/ml AbA (Fig. 5). These results suggest that MaMYBF and MaMYB1 might function as
transcription factors.
3.3 The spatiotemporal and light-responsive expression of
MaMYBF and MaMYB1
The bulbs, leaves and inflorescences of different flower developmental stages (S1–S5) of the blue
cultivar M. aucheri ‘Dark Eyes’ are shown in Fig. 6a. We observed that the flower color gradually became
darker during the five flowering stages, reaching a peak at stage 4 (Fig. 6a). qRT–PCR analysis revealed
that MaMYBF was highly expressed in young leaves and flowers and that MaMYB1 was highly expressed
in bulbs, old leaves and flowers (Fig. 6b). MaDFR, MaFLS, MaMybA, and MaMYBx were highly expressed
in flowers (Fig. 6b). The expression patterns of these genes in flowers at five developmental stages, from
closed buds (S1) to flower fading (S5), were examined. The results showed that the transcription of
MaMYBF and MaMYB1 peaked at stages 2 and 3, respectively, after which their transcription gradually
declined (Fig. 6a, c). qRT–PCR analysis showed that the expression patterns of MaMYBF and MaMYB1
were correlated with those of the flavonoid structural genes MaFLS and MaDFR, respectively (Fig. 6c).
Therefore, MaMYBF and MaMYB1 might be associated with flavonoid biosynthesis.
Several lines of evidence indicate that MYB regulators of the flavonoid biosynthesis pathway are
promoted by light-induced plant flavonoid accumulation (Gu et al. 2019; Zhang et al. 2019). Thus, we
cloned the promoter fragments of pMaMYBF (GenBank accession number: OM171269, 960 bp) and
pMaMYB1 (GenBank accession number: OM171268, 1288 bp) from M. aucheri 'Dark Eyes'. Analysis of
PlantCARE regulatory elements revealed that the promoter contained predicted light-related elements,
such as AE-box (AGAAACAA), GT1 motif (GTGTGTGAA), TCCC motif (TCTCCCT), and TCT motif
(TCTTAC) (Fig. 6d). These light-responsive elements suggest that the MaMYBF and MaMYB1 genes are
regulated by light.
Page 7/26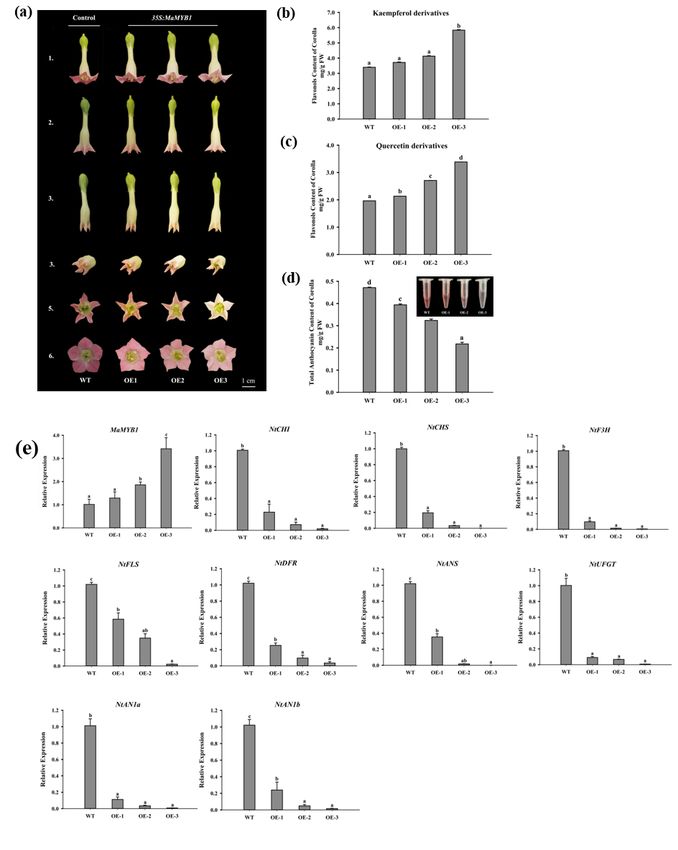
Next, we assessed their potential role in the light-induced flavonoid pathway in grape hyacinth. Our
results revealed that the shading treatment affected flower coloration in grape hyacinth, resulting in
etiolated phenotypes (Fig. 6e). HPLC analysis also showed that the total anthocyanin and flavonol
(kaempferol and quercetin) contents of the five developmental stages were significantly reduced (Fig. 6f).
Thus, we suggest that light plays an indispensable role in flower coloration of grape hyacinth. Moreover,
qRT–PCR analysis revealed that the expression of MaMYBF and MaMYB1 was downregulated from S2 to
S5 under shading treatment and significantly decreased at the S2 and S3 periods, respectively (Fig. 6g).
Moreover, the expression of flavonoid synthesis pathway structures (MaFLS, MaDFR) and regulatory
genes (MaMybA, MaMYBx) also significantly decreased under shading treatment (Fig. 6g). Overall,
MaMYBF and MaMYB1 may be involved in the regulation of light-induced flavonoid synthesis in grape
hyacinth.
3.4 MaMYB1 interacts with bHLH1 but MaMYBF does not
A bimolecular fluorescence complementation (BiFC) assay was conducted to test whether MaMYBF and
MaMYB1 binds to MabHLH1 in vivo. YFP fluorescence was observed in the nuclei of A. thaliana
protoplasts when pSPYCE/MabHLH1 was coexpressed with pSPYNE/MaMYB1 (Fig. 7). This result was
similar to that in the positive control group pSPYNE/MaMaMybA with pSPYCE/MabHLH1. However, YFP
fluorescence was not observed in the pSPYNE/MabHLH1 and pSPYCE/MaMYBF coexpression groups or
in any of the negative control groups, including pSPYCE/MaMYB1 with pSPYNE and pSPYCE with
pSPYNE/MaMYBF. Therefore, the BiFC assay confirmed that MaMYB1 interacts with MabHLH1 but
MaMYBF does not.
3.5 MaMYBF and MaMYB1 are involved in the flavonoid
pathway
To investigate the effect of MaMYBF and MaMYB1 on the transcription of key anthocyanin structural
genes, the flavonol-specific branching key gene FLS and the anthocyanin pathway late gene DFR were
chosen as potential targets of MaMYBF and MaMYB1 transcriptional activation. The cis-acting elements
of pMaFLS (Su 2020) and pMaDFR (Liu et al. 2019a) were analyzed using the online PlantCARE
database. As shown in Fig. 8a, pMaFLS and pMaDFR contained predicted MYB-binding elements: a MYB
core element (CNGTTR) and Myb (NAACNN). These structures suggest that MaMYBF and MaMYB1 are
regulated by MYBs.
The underlying regulatory relationships were further examined using a dual luciferase assay in N.
benthamiana leaves (Fig. 8b). The results showed that MaMYBF alone strongly activated pMaFLS, and
its activation was attenuated at reduced doses of MaMYBF in the presence of MabHLH1, MaMybA, and
MaMYB1 (Fig. 8c). However, MaMYBF did not exhibit the ability to activate pMaDFR either alone or in
coinfiltrated cells (Fig. 8d). Additionally, when MaMybA and MabHLH1 were coinfiltrated with MaMYB1,
pMaDFR activation was repressed, whereas MaMYB1 infiltration alone did not result in pMaFLS and
pMaDFR activation (Fig. 8c, d). Taken together, these results suggest that MaMYBF, a potential positive
Page 8/26
regulator of flavonol synthesis, regulates the flavonol pathway gene MaFLS and regulates the flavonol
pathway independently of MaMybA and MabHLH1 cofactors. In contrast, MaMYB1 is a potential
repressor of anthocyanin biosynthesis that binds to MabHLH1, disrupts the binding of the MaMybA-
MabHLH1 complex to pMaDFR, and activates its transcription in grape hyacinth.
3.6 Ectopic expression of MaMYBF and MaMYB1 regulates
flavonoid accumulation in tobacco
In MaMYBF- and MaMYB1-transgenic tobacco, the petals of OE-MaMYBF lines presented as white or light
pink color, whereas those of OE-MaMYB1 lines presented as pink color (Fig. 9a and Fig. 10a). A marked
difference was observed between OE-MaMYBF, OE-MaMYB1, and the WT lines (Fig. 9b and Fig. 10b).
HPLC analysis showed that the flavonol (kaempferol and quercetin) contents were significantly increased
in OE-MaMYBF and OE-MaMYB1 lines (Fig. 9c, d and 10c, d), whereas the total anthocyanin contents
were decreased in both OE-MaMYBF and OE-MaMYB1 tobacco petals (Fig. 9e and 10e). The
spatiotemporal expression analysis revealed that a synchronous relationship exists between the tissue-
specific expression patterns of MaMYBF and MaFLS (Fig. 6b). qRT–PCR assays demonstrated that the
expression of NtFLS was significantly upregulated in OE-MaMYBF lines compared to WT, while genes
involved in the flavonoid pathway, including NtCHI, NtCHS, NtF3H, NtDFR, NtANS, NtUFGT and bHLH
endogenous genes NtAN1a and NtAN1b, were significantly downregulated (Fig. 9f). However, NtCHI,
NtCHS, NtF3H, NtFLS, NtDFR, NtANS, NtUFGT, NtAN1a and NtAN1b in OE-MaMYB1 lines were
downregulated (Fig. 10f). Thus, these results indicate that MaMYBF and MaMYB1 may regulate
flavonoid biosynthesis in tobacco plants and produce shallower phenotypes (Figs. 9 and 10).
4 Discussion
MYB TFs play key roles in plant flavonoid biosynthesis. In grape hyacinth, MaAN2 (Chen et al. 2017),
MaMybA (Chen et al. 2019), and MaMYBx (Zhang et al. 2020) have been shown to regulate anthocyanin
accumulation. However, whether other MYBs are involved in grape hyacinth flavonoid metabolism and
flower coloration is unclear. Here, we identified a flavonol regulator and an anthocyanin repressor that
have essential roles in the grape hyacinth flavonoid pathway.
Phylogenetic analysis revealed that MaMYBF belongs to SG7 MYBs (Fig. 3). Notably, the MaMYBF
protein contained the flavonol-specific regulator SG7 motif and displayed high sequence similarity to
other MYB TF members of the SG7 flavonol clade of the R2R3 MYB family. The conserved DNA-binding
domain (R2 and R3 repeats) in the N-terminal and the two SG7 motifs have been used as a tool to
identify additional putative flavonol-specific transcriptional regulators of plant species.
However, the signature motif SG7-2 ([W/x][L/x]LS) detected at the C-terminus of VvMYBF1 (Czemmel et
al. 2009), AtMYB11 (Stracke et al. 2007), AtMYB12 (Mehrtens et al. 2005), and VviMYBF1 (Czemmel et
al. 2017) was absent in MaMYBF (Fig. 1). This is consistent with other SG7 MYB proteins, including
AtMYB111 (Stracke et al. 2007), LhMYB12 (Yamagishi et al. 2012), and MtMYB134 (Naik et al. 2021).
Moreover, the SG7-2 domain is less conserved in Zea mays ZmP and apple MdMYB22 (Wang et al.
Page 9/26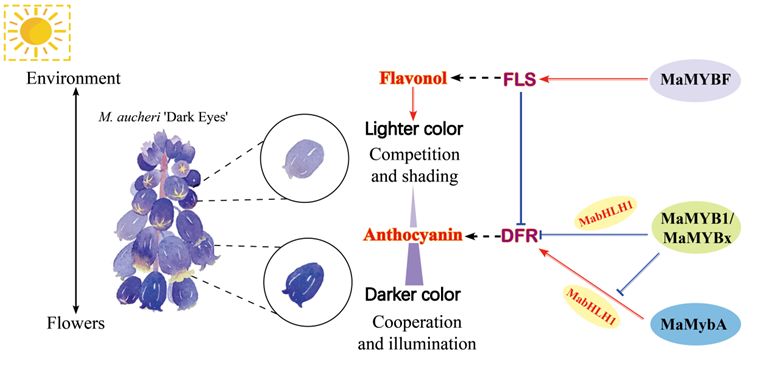
2017). Therefore, both the SG7 and SG7-2 motifs may be part of specific functional domains outside the
DNA-binding region of flavonol regulators and may play redundant roles (Czemmel et al. 2009).
Studies have shown that MYB TFs promote flavonoid synthesis in response to light induction (Gu et al.
2019). The promoter fragments of MaMYBF and MaMYB1 contain predicted light-related elements and
are primarily expressed during flower development and light induction (Fig. 6b, d). Furthermore, shading
treatment resulted in etiolated phenotypes of grape hyacinth, and the content of flavonoids and the
expression of related genes were significantly downregulated (Fig. 6e, f). These results are in line with our
previous gene expression level studies in other horticultural fruit crops showing the correlation of
MaMYBF and MaMYB1 expression with light-induced flavonoid synthesis (Aharoni et al. 2001; Ferreyra et
al. 2010).
In the current study, overexpression of MaMYBF in tobacco upregulated NtFLS expression. The flavonoid-
related genes NtCHI, NtCHS, NtF3H, NtDFR, NtANS, and NtUFGT and the bHLH endogenous genes NtAN1a
and NtAN1b were significantly downregulated in OE-MaMYBF (Fig. 9e). Dual luciferase assays
confirmed that MaMYBF activated the promoter of MaFLS. MaMYBF alone activated more
strongly than coinfection, which may indicate that MaMYBF regulates the flavonol pathway
independently of cofactors (Fig. 8). Based on the overall data from our study, we propose that MaMYBF
contributes to grape hyacinth flavonol biosynthesis and results in light phenotypes.
We also found that overexpression of MaMYB1 in tobacco resulted in the accumulation of large amounts
of flavonol and significantly downregulated the expression of flavonoid pathway structural genes,
resulting in lighter phenotypes (Fig. 10). This indicates the recruitment of MaMYB1, which strongly
represses flavonoid pathway structural genes, represses anthocyanin biosynthesis. Of note, flavonol and
anthocyanin share the same biosynthetic pathway from phenylalanine to dihydroflavonol, and the key
gene FLS of the flavonol synthesis pathway and the key gene DFR of the anthocyanin synthesis
pathway compete from common precursors. Decoupling FLS and DFR regulation should theoretically
result in an increase in precursor flow to the flavonol branch over the anthocyanin branch, when
MaMYB1 represses the expression of MaDFR, thereby enhancing flux in the flavonol
pathway. Furthermore, we found that MaMybA alone slightly activated pMaFLS and that MaMybA
enhanced activation in the presence of MabHLH1 (Fig. 8c, d). We found that the pMaFLS promoter
contained putative MYB-binding elements (Fig. 8a). Interestingly, the apple R2R3 MYB activator MYB10
exerted a significant effect on the promoter activity of FLS in addition to UFGT and ANS when interacting
with bHLH3 (Wang et al. 2017). As described previously, the Arabidopsis activator PAP1 mutant pap1-D is
characterized by strongly increased concentrations of flavonols in comparison with wild-type Col-0
(Borevitz et al. 2000). This suggests that anthocyanin R2R3 MYB activators may also display a degree
of subfunctionalization for regulating flavonol biosynthesis.
In conclusion, our study reveals a new type of regulatory network coregulated by MaMYBF, MaMYB1 and
other MYBs that regulates flavonoid metabolism and flower coloration in grape hyacinth (Fig. 11).
MaMYBF independently affects the transcriptional activation of MaFLS and promotes flavonol
Page 10/26accumulation. MaMYB1 and MaMYBx act as anthocyanin-related repressors that can bind to MabHLH1,
affecting the transcriptional activation of MaDFR by disturbing the interaction between MaMybA and
MabHLH1. Notably, expression of these MYB TFs may be induced by light signals and affect the
pigmentation of grape hyacinth flowers through their influence on structural gene expression levels
related to flavonoid biosynthesis. In the study of petunia, a feedback mechanism exists in R2R3 MYB
TFs, with the R3 MYB repressor PhMYBx being expressed under high light, while the R2R3 repressor
PhMYB27 is highly expressed during noninductive shade treatments (Albert et al. 2011). Visible light and
UV-B strongly affect the expression of grapevine R2R3 MYB TFs VviMYBF1 and its target VviFLS1 to
regulate flavonoid synthesis (Czemmel et al. 2017). However, to date, the deep molecular mechanism of
light-induced anthocyanin biosynthesis remains elusive, and many challenges remain unclear regarding
phenotypic change and corresponding gene regulation in ornamental plants. Therefore, future research
should focus on the impact of environmental factors, such as light, on plant flavonoid biosynthesis and
further establish a feedback regulatory network for flavonoid biosynthesis in grape hyacinth.
Declarations
Acknowledgments
The authors would like to thank the anonymous reviewers for their comments. This study was supported
by the National Natural Science Foundation of China (grant nos. 32171863 and 31700625), the Shaanxi
Province Key Research and Development Program (grant no. 2021NY-071) and the Fundamental
Research Funds for the Central Universities (grant no. SWU118099).
Author contribution statement
DL conceived and designed the research. WJ, ZH, TS, HW, and CK conducted the experiments and
analyzed the data. WJ wrote the manuscript. DL and ZH modified the manuscript. All authors read and
approved the manuscript.
Compliance with Ethical Standards
Conflict of Interest
The authors declare that they have no conflicts of interest.
References
1. Aharoni A, De Vos CHR, Wein M, Sun Z, Greco R, Kroon A, Mol JNM, O'Connell AP (2001) The
strawberry FaMYB1 transcription factor suppresses anthocyanin and flavonol accumulation in
transgenic tobacco. Plant J 28:319–332
2. Albert NW, Davies KM, Lewis DH, Zhang H, Montefiori M et al (2014) A conserved network of
transcriptional activators and repressors regulates anthocyanin pigmentation in eudicots. Plant Cell
Page 11/2626:962–980
3. Albert NW, Lewis DH, Zhang H, Schwinn KE, Jameson PE, Davies KM (2011) Members of an R2R3-
MYB transcription factor family in Petunia are developmentally and environmentally regulated to
control complex floral and vegetative pigmentation patterning. Plant J 65:771–784
4. Andrew CA, Richard VE (2018) MYBs drive novel consumer traits in fruits and vegetables. trends in
plant sci 23:693–705
5. Anwar M, Wang GQ, Wu JC et al (2018) Ectopic overexpression of the novel R2R3 MYB NtMYB2 from
Chinese narcissus represses anthocyanin biosynthesis in tobacco. Molecules 23(4):781
6. Borevitz JO, Xia YJ, Blount J, Dixon RA, Lamb C (2000) Activation tagging identifies a conserved
MYB regulator of phenylpropanoid biosynthesis. Plant Cell 12:2383–2393
7. Chen K, Du L, Liu H, Liu Y (2019) A novel R2R3 MYB from grape hyacinth MaMybA, which is different
from MaAN2, confers intense and magenta anthocyanin pigmentation in tobacco. BMC Plant Biol
19:390
8. Chen K, Liu H, Lou Q, Liu Y (2017) Ectopic expression of the grape hyacinth (Muscari armeniacum)
R2R3 MYB transcription factor gene MaAN2 induces anthocyanin accumulation in tobacco. Front
Plant Sci 8:965
9. Czemmel S, Höll J, Loyola R et al (2017) Transcriptome-wide identification of novel UV-B- and light
modulated flavonol pathway genes controlled by VviMYBF1. Front Plant Sci 8:1084
10. Czemmel S, Stracke R, Weisshaar B, Cordon N, Harris NN, Walker AR et al (2009) The grapevine R2R3
MYB transcription factor VvMYBF1 regulates flavonol synthesis in developing grape berries. Plant
Physiol 151:1513–1530
11. Dubos C, Stracke R, Grotewold E, Weisshaar B, Martin C, Lepiniec L (2010) MYB transcription factors
in Arabidopsis. Trends Plant Sci 15:573–581
12. Ferreyra MLF, Rius S, Emiliani J, Pourcel L, Feller A, Morohashi K, Casati P, Grotewold E (2010)
Cloning and characterization of a UV-B-inducible maize flavonol synthase. Plant J 62:77–91
13. Gu KD, Wang CK, Hu DG, Hao YJ (2019) How do anthocyanins paint our horticultural products? Sci
Hortic-Amsterdam 249:257–262
14. Hellens RP, Allan AC, Friel EN, Bolitho K, Grafton K, Templeton MD et al (2005) Transient expression
vectors for functional genomics, quantification of promoter activity and RNA silencing in plants.
Plant Methods 1:13–26
15. Horsch R, Fry JE, Hoffmann NL, Eichholtz DZ, Rogers SG, Fraley RT (1985) A simple and general
method for transferring genes into plants. Science 227:1229–1232
16. Huang W, Zeng S, Gong X, Wei G, Liao S, Chen J, Wei S, Lv H, Ying W (2015) Elucidating the
biosynthetic and regulatory mechanisms of flavonoid-derived bioactive components in Epimedium
sagittatum. Front Plant Sci 6:689
17. Jin HL, Cominelli E, Bailey P, Parr A, Mehrtens F, Jones J, Tonelli C, Weisshaar B, Martin C (2000)
Transcriptional repression by AtMYB4 controls production of UV-protecting sunscreens in
Page 12/26Arabidopsis. EMBO J 19:6150–6161
18. Li Y, Shan X, Gao R et al (2020) MYB repressors and MBW activation complex collaborate to fine-tune
flower coloration in Freesia hybrida. Commun Biol 3:396
19. Liu H, Lou Q et al (2019a) Cloning and functional characterization of dihydroflavonol 4-reductase
Gene Involved in anthocyanidin biosynthesis of grape hyacinth. Int J Mol Sci 20(19):4743
20. Liu H, Su B, Zhang H et al (2019b) Identification and functional analysis of a flavonol synthase gene
from grape hyacinth. Molecules 24(8):1579
21. Lou Q, Liu Y, Qi Y, Jiao S, Tian F, Jiang L et al (2014) Transcriptome sequencing and metabolite
analysis reveals the role of delphinidin metabolism in flower colour in grape hyacinth. J Exp Bot
65:3157–3164
22. Lou Q, Wang L, Liu HL, Liu YL (2017) Anthocyanin profiles in flowers of grape hyacinth. Molecules
22:688
23. Martens S, Teeri T, Forkmann G (2002) Heterologous expression of dihydroflavonol 4-reductases
from various plants. FEBS Lett 531:453–458
24. Mehrtens F, Kranz H, Bednarek P, Weisshaar B (2005) The Arabidopsis transcription factor MYB12 is
a flavonol-specific regulator of phenylpropanoid biosynthesis. Plant Physiol 138:1083–1096
25. Naik J, Rajput R, Pucker B, Stracke R, Pandey A (2021) The R2R3-MYB transcription factor
MtMYB134 orchestrates flavonol biosynthesis in Medicago truncatula. Plant Mol Biol 106(1):157–
172
26. Ramsay NA, Glover BJ (2005) MYB-bHLH-WD40 protein complex and the evolution of cellular
diversity. Trends Plant Sci 10:63–70
27. Sateesh K, Kevin R (2011) Ear motif-mediated transcriptional repression in plants: An underlying
mechanism for epigenetic regulation of gene expression. Epigenetics 6:141–146
28. Shan X, Li Y, Yang S, Gao R, Zhou L, Bao T, Han T, Wang S, Gao X, Wang L (2019) A functional
homolog of Arabidopsis TTG1 from Freesia interacts with bHLH proteins to regulate anthocyanin
and proanthocyanidin biosynthesis in both Freesia hybrida and Arabidopsis thaliana. Plant Physiol
Bioch 141:60–72
29. Shan X, Li Y, Yang S, Yang Z, Qiu M, Gao R, Han T, Meng X, Xu Z, Wang L, Gao X (2020) The spatio-
temporal biosynthesis of floral flavonols is controlled by differential phylogenetic MYB regulators in
Freesia hybrida. New Phytol 228:1864–1879
30. Sharma SB, Dixon RA (2005) Metabolic engineering of proanthocyanidins by ectopic expression of
transcription factors in Arabidopsis thaliana. Plant J 44:62–75
31. Stracke R, Ishihara H et al (2007) Differential regulation of closely related R2R3 MYB transcription
factors controls flavonol accumulation in different parts of the Arabidopsis thaliana seedling. Plant J
50:660–677
32. Stracke R, Werber M, Weisshaar B (2001) The R2R3-MYB, gene family in Arabidopsis thaliana. Curr
Opin Plant Biol 4:447–456
Page 13/2633. Su BB (2020) Cloning and functional study of grape hyacinth MaFLS Gene Promoter. Dissertation,
Northwest A&F University
34. Takahashi R, Githiri SM, Hatayama K, Dubouzet EG (2007) A single-base deletion in soybean flavonol
synthase gene is associated with magenta flower color. Plant Mol Biol 63:125–135
35. Takos AM, Jaffe FW, Jacob SR, Bogs J, Robinson SP, Walker AR (2006) Light-induced expression of a
MYB gene regulates anthocyanin biosynthesis in red apples. Plant Physiol 142:1216–1232
36. Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) MEGA6: molecular evolutionary genetics
analysis version 6.0,Mol Biol Evol30
37. Terrier N, Torregrosa L, Ageorges A et al (2009) Ectopic expression of VvMybPA2 promotes
proanthocyanidin biosynthesis in grapevine and suggests additional targets in the pathway. Plant
Physiol 149:1028–1041
38. Wang N, Xu H, Jiang S, Zhang Z, Lu N, Qiu H, Qu C, Wang Y, Wu S, Chen X (2017) MYB12 and MYB22
play essential roles in proanthocyanidin and flavonol synthesis in red-fleshed apple (Malus sieversii
f. niedzwetzkyana). Plant J 90:276–292
39. Wellmann F, Lukacin R, Moriguchi T, Britsch L, Schiltz E, Matern U (2002) Functional expression and
mutational analysis of flavonol synthase from Citrus unshiu. Eur J Biochem 269:4134–4142
40. Williams CA, Grayer RJ (2001) Anthocyanins and other flavonoids. Nat Prod Rep 21:539–573
41. Xu H, Wang N, Liu J, Qu C, Wang Y, Jiang S, Lu N, Wang D, Zhang Z, Chen X (2017) The molecular
mechanism underlying anthocyanin metabolism in apple using the MdMYB16 and MdbHLH33
genes. Plant Mol Biol 94:149–165
42. Yamagishi M, Yoshida Y, Nakayama M (2012) The transcription factor LhMYB12 determines
anthocyanin pigmentation in the tepals of Asiatic hybrid lilies (Lilium spp.) and regulates pigment
quantity. Mol Breed 30(2):913–925
43. Zhang H, Gong JX, Chen KL, Yao WK, Zhang BX, Wang JY, Tian ST et al (2020) A novel R3 MYB
transcriptional repressor, MaMYBx, finely regulates anthocyanin biosynthesis in grape hyacinth.
Plant Sci 298:110588
44. Zhang YJ, Li Y, Li WP, Hu ZL, Yu XH, Tu Y, Zhang M, Huang JY, Chen GP (2019) Metabolic and
molecular analysis of nonuniform anthocyanin pigmentation in tomato fruit under high light. Hortic
Res 6:56
Figures
Page 14/26Figure 1
Alignment of the protein sequence of MaMYBF with selected homologs. The alignment was performed
using MEGA 6 and Genedoc software. Amino acids with 100% identity are shown with an orange
background, and those with 80% and 60% identity are shown in blue and yellow. The characteristic R2/R3
domains and SG7 motifs are shown with orange lines, blue lines and red frames, respectively.
Page 15/26Figure 2
Alignment of the protein sequence of MaMYB1 with selected homologs.a The alignment was performed
using MEGA 6 and Genedoc software. Amino acids with 100% identity are shown with an orange
background, and those with 80% and 60% identity are shown in blue and yellow, respectively. The
characteristic R2/R3 domains are shown with orange and blue lines, respectively. The characteristic C1-
C5 domains are shown with blue, green, pink, orange and black frames, respectively. b Amino acid
sequences were submitted to MEME Suite (http://meme-suite.org/), and conserved domain analysis was
used to annotate the locations of R2/R3 domains and the C1-C5 motif.
Page 16/26Figure 3
Phylogenetic analysis of MaMYBF, MaMYB1 and other MYB regulators related to flavonoid biosynthesis.
The phylogenetic tree was constructed using MEGA 6. The numbers at each interior branch indicate the
bootstrap values of 1000 replicates. MaMYBF and MaMYB1 are marked in blue and red, respectively.
Clades are indicated in color as follows: yellow, subgroup 4 transcriptional repressors for flavonoid
biosynthesis and lignin; blue, subgroup 5 for proanthocyanidin synthesis; pink, subgroup 6 for
anthocyanin biosynthesis; gray, subgroup 7 for flavonol biosynthesis; green, CPC-like transcriptional
repressors. GenBank accession numbers for all proteins are listed in Supplementary Table S3.
Page 17/26Figure 4
Subcellular localization of MaMYBF and MaMYB1. The 35S:GFP:MaMYBF and 35S:GFP:MaMYB1 fusion
proteins in Arabidopsis thaliana mesophyll protoplasts. Free GFP served as a control. MaMYBF and
MaMYB1 are located in the nucleus. Autofluorescence: chloroplast autofluorescence; GFP: GFP
fluorescence; Merge: merged images of chloroplast autofluorescence, GFP fluorescence, and bright-field
microscopy. Scale bars, 10 mm.
Page 18/26Figure 5
Transcriptional activation ability of MaMYBF and MaMYB1 in Y2Hgold yeast. The positive control was
pGBKT7-53 plus pGADT7-T, and the negative control was pGBKT7. Yeasts transformed with the positive
and negative controls, pGBKT7-MaMYBF and pGBKT-MaMYB1 vectors were cultivated in SD/-Trp
medium, SD/-Trp medium with 60 μg/ml X-α-Gal, and SD/-Trp medium plus 60 μg/ml X-α-Gal and 200
ng/ml AbA, respectively. The positive control and pGBKT7-MaAN2 exhibited blue yeast plaques, while the
negative control did not grow in SD/-Trp medium supplemented with 60 μg/ml X-α-Gal and 200 ng/ml
AbA.
Page 19/26Figure 6
The spatiotemporal and light-responsive expression of MaMYBF and MaMYB1. a Bulbs, young leaves,
old leaves, and petals of five flower developmental stages of M. aucheri ‘Dark Eyes’. Scale bars, 1 cm. b
The expression profiles of MaMYBF, MaMYB1, MaMybA, MaMYBx, MaFLS, and MaDFR in each tissue of
M. aucheri ‘Dark Eyes’. c The expression profiles of MaMYBF, MaMYB1, MaFLS, and MaDFR at different
flower developmental stages in M. aucheri ‘Dark Eyes’. d Schematic of the promoters of pMaMYBF and
Page 20/26pMaMYB1. The lengths and light-responsive elemental sites of the pMaMYBF and pMaMYB1 promoters
are indicated by colorful ellipses. e The inflorescence phenotypes of M. aucheri ‘Dark Eyes’ in five
flowering stages (S1-S5) under light and dark treatment. f The anthocyanin and flavonol contents of
petals at the five flower developmental stages. DW: dry weight. Different lowercase letters represent
significant differences calculated using least-significant difference (LSD) analysis at the level of P < 0.05.
g The expression profiles of MaMYBF, MaMYB1, MaFLS, MaDFR, MaMybA, and MaMYBx in flowers
during the five developmental stages (S1–S5) of M. aucheri ‘Dark Eyes’ under light and dark treatment.
MaActin was used as the reference gene to normalize the expression of these genes. Each column
represents the mean ± SD from three independent experiments.
Figure 7
Page 21/26Bimolecular fluorescence complementation of MaMybA, MaMYBF, MaMYB1 and MabHLH1 interactions
in A. thaliana mesophyll protoplasts. YFP: fluorescence of YFP; Merge is the digital image merged with
bright field and fluorescent images. Scale bars, 10 μm.
Figure 8
MaMYBF and MaMYB1 are involved in the flavonoid pathway. a Distribution of MYB-binding elements in
the promoters of MaFLS and MaDFR. b Schematic representation of the constructs used for the dual-
luciferase assay. The reporter construct contains firefly luciferase (LUC) driven by the promoter of MaFLS
or MaDFR and Renilla luciferase (REN) driven by the CaMV 35S promoter. The effector constructs contain
MaMYBF, MaMYB1, MaMybA or MabHLH1 driven by the CaMV 35S promoter. c Dual luciferase assays
of the effects of MaMybA, MaMYB1, MaMYBF, MaMybA/MaMYBF, MaMYBF/MaMYB1,
MaMybA/MabHLH1, MaMYB1/MabHLH1, MaMYB1/MaMybA/MaMYBF, and
MaMybA/MabHLH1/MaMYB1 on the transcriptional regulation of the promoters of key anthocyanin
Page 22/26biosynthetic genes (MaFLS and MaDFR) in tobacco leaves. Data are shown as the mean ± SD. Different
letters above the bars indicate significantly different values calculated by Tukey’s HSD tests (P < 0.05).
Figure 9
Overexpression of MaMYBF in tobacco plants promotes flavonol accumulation and reduces anthocyanin
accumulation in the corolla. a Pigmentation phenotypes of flowers of wild-type (WT) and three MaMYBF-
Page 23/26overexpressing tobacco lines (OE1, OE2, and OE3) at three flowering stages (1–3). b, c HPLC analysis of
anthocyanin and flavonol in tobacco corollas in mg/g fresh weight (FW) of WT, OE1, OE2, and OE3. A
standard curve of cyanidin 3-rutinoside, quercetin, and kaempferol content was used to calculate
anthocyanin and flavonol contents. Data are shown as the mean±SD. d, e Expression profiles of
MaMYBF, flavonol and anthocyanin biosynthesis (NtCHS, NtCHI, NtF3H, NtFLS, NtDFR, NtANS, NtUFGT,
NtAN1a, and NtAN1b) in corollas of WT, OE1, OE2, and OE3 tobacco. NtTubA1 was used as an internal
control. Data are shown as the mean ± SD. Different letters above the bars indicate significantly different
values calculated by Tukey’s HSD tests (P < 0.05).
Page 24/26Figure 10
Overexpression of MaMYB1 in tobacco plants reduces anthocyanin accumulation in the corolla. a
Pigmentation phenotypes of flowers of wild-type (WT) and three MaMYB1-overexpressing tobacco lines
(OE1, OE2, and OE3) at three flowering stages (1–3). b, c HPLC analysis of anthocyanin and flavonol in
tobacco corollas in mg/g fresh weight (FW) of WT, OE1, OE2, and OE3. A standard curve of cyanidin 3-
rutinoside, quercetin, and kaempferol content was used to calculate anthocyanin and flavonol contents.
Page 25/26Data are shown as the mean±SD. d, e Expression profiles of MaMYB1, flavonol and anthocyanin
biosynthesis (NtCHS, NtCHI, NtF3H, NtFLS, NtDFR, NtANS, NtUFGT, NtAN1a, and NtAN1b) in corollas of
WT, OE1, OE2, and OE3 tobacco. NtTubA1 was used as an internal control. Data are shown as the mean ±
SD. Different letters above the bars indicate significantly different values calculated by Tukey’s HSD tests
(P < 0.05).
Figure 11
Suggested model of action of MaMYBF, MaMYB1 and other MYBs in flavonoid biosynthesis in grape
hyacinth. MaMYBF represents an SG7 group activator of flavonol biosynthesis, which affects the
transcriptional activation of MaFLS. MaMYBF promotes flavonol accumulation and reduces anthocyanin
accumulation. MaMybA represents an R2R3 MYB activator of anthocyanin biosynthesis. MaMybA and
MabHLH1 form the anthocyanin activator complex. MaMYB1 and MaMYBx act as anthocyanin-related
repressors that can bind to MabHLH1, thereby affecting the transcriptional activation of MaDFR by
disturbing the interaction between MaMybA and MabHLH1. The arrows and blunt-ended lines indicate
positive or negative regulation, respectively.
Supplementary Files
This is a list of supplementary files associated with this preprint. Click to download.
ElectronicSupplymentMaterial.docx
Page 26/26You can also read

















































