Extracellular Vesicles in Cancer Metastasis: Potential as Therapeutic Targets and Materials - MDPI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
International Journal of
Molecular Sciences
Review
Extracellular Vesicles in Cancer Metastasis:
Potential as Therapeutic Targets and Materials
Akiko Kogure, Yusuke Yoshioka * and Takahiro Ochiya
Department of Molecular and Cellular Medicine, Institute of Medical Science, Tokyo Medical University,
Shinjuku-ku, Tokyo 1600023, Japan; akogure@tokyo-med.ac.jp (A.K.); tochiya@tokyo-med.ac.jp (T.O.)
* Correspondence: yyoshiok@tokyo-med.ac.jp; Tel.: +81-3-3342-6111
Received: 19 May 2020; Accepted: 22 June 2020; Published: 23 June 2020
Abstract: The vast majority of cancer-related deaths are due to metastasis of the primary tumor
that develops years to decades after apparent cures. However, it is difficult to effectively prevent or
treat cancer metastasis. Recent studies have shown that communication between cancer cells and
surrounding cells enables cancer progression and metastasis. The comprehensive term “extracellular
vesicles” (EVs) describes lipid bilayer vesicles that are secreted to outside cells; EVs are well-established
mediators of cell-to-cell communication. EVs participate in cancer progression and metastasis by
transferring bioactive molecules, such as proteins and RNAs, including microRNAs (miRNAs),
between cancer and various cells in local and distant microenvironments. Clinically, EVs functioning
as diagnostic biomarkers, therapeutic targets, or even as anticancer drug-delivery vehicles have been
emphasized as a result of their unique biological and pathophysiological characteristics. The potential
therapeutic effects of EVs in cancer treatment are rapidly emerging and represent a new and important
area of research. This review focuses on the therapeutic potential of EVs and discusses their utility for
the inhibition of cancer progression, including metastasis.
Keywords: extracellular vesicles; cancer metastasis; cancer therapy; cancer biomarker
1. Introduction
Metastasis is the primary cause of cancer mortality. It is responsible for 90% of all cancer-related
deaths. The metastatic process involves invasion of tumor cells from the primary tumor, intravasation,
arrest and extravasation from the circulatory system, followed by angiogenesis and growth at a distant
site. Many of the molecular pathways that promote tumorigenesis also promote metastasis and are
important in the treatment of both aspects of cancer progression. Although cancer therapies, including
chemoradiotherapy, immunotherapy, and molecularly targeted treatment, have been developed,
there has been a lack of satisfactory clinical outcomes for patients with cancer metastasis [1,2]. Most
therapeutic developments are based on anticancer activity against tumorigenesis and primary growth
rather than cancer metastasis. Further clinical therapy applications of agents and preclinical evidence
for the effectiveness of these agents against cancer metastasis are still lacking. Therefore, it is important
to develop new antimetastatic therapeutics for clinical application.
Cancer metastasis refers to the process by which primary tumor cells invade other sites of the body,
proliferate, and finally form new tumors. In the complex tumor microenvironment, cancer cells and
surrounding cells (e.g., endothelial cells, fibroblasts, and macrophages) communicate with each other,
which creates a metastatic niche and promotes cancer metastasis [3,4]. Recently, “extracellular vesicles”
(EVs) have attracted more attention as a novel mode of cell-cell communication. EVs are cell-derived
vesicles that contain lipids, proteins, mRNAs, and small RNAs, such as microRNAs (miRNAs). EVs
were previously seen as “garbage bags” for the elimination of cellular components. However, in 1996,
Raposo et al. showed that EVs derived from B cells induced antigen-specific major histocompatibility
Int. J. Mol. Sci. 2020, 21, 4463; doi:10.3390/ijms21124463 www.mdpi.com/journal/ijms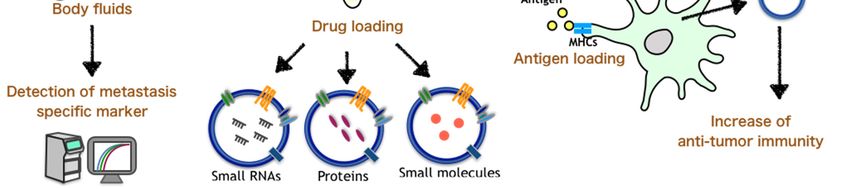
Int. J. Mol. Sci. 2020, 21, 4463 2 of 16
complex (MHC) class II-restricted T cell responses [5]. In 2007, Valadi et al. reported that the mRNAs
contained in EVs could be translated in recipient cells [6]. Moreover, several groups demonstrated that
EV-associated miRNAs were also transferred to recipient cells and led to gene silencing in recipient
cells [7–9]. These findings suggested that EVs could contribute to the transfer of functional contents,
and EVs have rapidly attracted attention as mediators of cell-to-cell communication. Many important
biological functions of EVs have been revealed, including functions in cancer, in recent years. Recent
work also showed that EVs secreted from various types of cells transport and transfer bioactive
molecules, and EVs have attracted broad attention for their ability to regulate metastasis [10–13]. In this
review, we summarize the current knowledge of EVs in cancer metastasis. Furthermore, we discuss
the potential use of EVs in clinical treatment of metastasis.
2. Extracellular Vesicles (EVs)
2.1. Classification and Biogenesis of EVs
EVs are generally classified into at least three types based on their size and biogenesis: exosomes,
apoptotic bodies, and microvesicles. Exosomes are approximately 100-nm extracellular vesicles that are
generated by the fusion of multivesicular bodies (MVBs) to the plasma membrane [14–17]. The fusion
of the MVB with the plasma membrane is in part regulated by neutral sphingomyelinase 2 (nSMase2),
endosomal sorting complex required for transport (ESCRT), syntenin, ALIX, tetraspanins, Rab proteins,
and phospholipase D2 [17,18]. Due to their endosomal origin, all exosomes contain membrane transport
and fusion proteins (GTPases, Annexins, and flotillin), tetraspanins (CD9, CD63, CD81, and CD82),
heat shock proteins (Hsc70, Hsp90, Hsp60m, and Hsp20), proteins involved in multivesicular body
biogenesis (Alix and TSG101), and lipid-related proteins and phospholipases [15,17]. Microvesicles
are generated by outward budding and fission of the plasma membrane and the subsequent release
of these vesicles into the extracellular space. They are usually approximately 1 µm in diameter [19].
Microvesicles are enriched in some lipids and phosphatidylserine [20]. Apoptotic bodies are generally
larger than exosomes and microvesicles (µm size) and are produced by dying cells. They contain
several intracellular fragments, cellular organelles, membranes, and cytosolic contents [21]. However,
it is still difficult to accurately distinguish between different types of vesicles. In this review, we use
the term EVs for all types of vesicles present in the extracellular space.
2.2. Isolation Methods of EVs
The most popular method of EV isolation remains differential ultracentrifugation (UC) [22].
In the first step, contaminating material is removed through a series of low-speed centrifugation
steps. Following this, at higher speeds, the EV fraction is generated. Although UC is a low-cost and
simple procedure, making it an ideal means of EV isolation in many labs, nonvesicular proteins are
present as contaminants [23]. The combination of UC with other purification methods, such as density
gradient centrifugation or ultrafiltration, can increase the purity of the EV fraction [22,24]. Density
gradient-based isolation can provide high purification of the EV fraction. However, the method is
time-consuming and cannot separate EVs from contaminants of similar density. There are several
ways to isolate EVs without using UC. For example, size exclusion chromatography (SEC) is another
isolation approach for EVs. The SEC method separates solution molecules based on their size using gel
filtration. Several commercial products have been used for EV isolation, such as qEV (IZON) [25]. This
method is easy and results in high purity from a small amount of sample, although it is not suitable
for a large amount of sample [26]. Because of the presence of many proteins and receptors in the
membrane of EVs, immunoaffinity capture can be used to isolate EVs by relying on immunoaffinity
interactions between proteins, such as CD63 and CD9, and their antibodies [27]. The combination of
immunoaffinity and microfluidic system approaches has been developed for the detection of EV-based
diagnostic biomarkers. As immunoaffinity capture can trap specific markers of EVs, the degree of
purification could be higher than that of other methods, as mentioned above. However, becauseInt. J. Mol. Sci. 2020, 21, 4463 3 of 16
Int. J. Mol. Sci. 2020, 21, x FOR PEER REVIEW 3 of 16
the method is unsuitable for EV isolation on a large scale, EVs would need to be preconcentrated
94 preconcentrated
with another method.with another method.
In addition, theIninteraction
addition, the interaction
between between
EVs and theirEVs and their
antibodies is antibodies
not easily
95 is
reversible [22,24,28]. Therefore, the method is limited to downstream experiments. For EVs toFor
not easily reversible [22,24,28]. Therefore, the method is limited to downstream experiments. be
96 EVs to be
applied in applied in clinical
clinical settings, settings,
they must bethey must beatproduced
produced at a Recently,
a large scale. large scale. Recently, a hollow-fiber
a commercial commercial
97 hollow-fiber
bioreactor systembioreactor system
has been has been
utilized utilized
effectively foreffectively
large-scaleforEVlarge-scale
production EV[29,30].
production [29,30]. The
The hollow-fiber
98 hollow-fiber bioreactor
bioreactor is based is basedthe
on culturing on cells
culturing the cells
in hollow and in hollow and semipermeable
semipermeable fibers, which canfibers, which
attain can
a higher
99 attain a higher cell yield in a short incubation time. By scaling up the cell culture,
cell yield in a short incubation time. By scaling up the cell culture, the EV yield could be increased.the EV yield could
100 be increased.
Thus, although various isolation techniques have been developed for clinical applications, it is
101 Thus, although
still difficult to obtainvarious
both high isolation techniques
efficiency and high have been
purity developed
when isolatingforEVs.
clinical applications, it is
102 still difficult to obtain both high efficiency and high purity when isolating EVs.
3. Function of EVs in Cancer Metastasis
103 3. Function of EVs in Cancer Metastasis
Crosstalk between primary cancer cells and microenvironmental cells is a key step in cancer
104 Crosstalk
metastasis, between primary
as described cancer cells and
in the Introduction microenvironmental
section. Recently, severalcells is a key
studies step in
revealed cancer
that EVs
105 metastasis,
contribute toasdistinct
described
stepsin of the Introduction
cancer metastasis,section. Recently, several
such as promoting studiesmodulating
angiogenesis, revealed that EVs
immune
106 contribute to distinct steps of cancer metastasis, such as promoting angiogenesis,
systems, and conditioning the premetastatic niche (Figure 1). In this section, we describe cell-to-cell modulating
107 immune systems,
communication and conditioning
between tumor cells the andpremetastatic nichethrough
surrounding cells (FigureEVs.
1). In this section, we describe
108 cell-to-cell communication between tumor cells and surrounding cells through EVs.
Cancer-derived EVs
CAF or MSC-derived EVs
Mesenchymal stem cells Metastatic sites
Fibroblasts Induction of EMT
Tumor cells Pre-metastatic niche formation
CAF-transition Epithelial cells
Induction of EMT
Recruitment
Inhibition of activity
Angiogenesis Disruption T cells Neutrophils
of tight junction
NK cells
Endothelial cells
Expansion
Regulatory T cells
109
110 Figure 1.
Figure Cell-to-cell communication
1. Cell-to-cell communication between
between tumor
tumor cells
cells and
and surrounding
surrounding cells
cells via
via “extracellular
“extracellular
vesicles” (EVs) in metastasis. Cancer-derived EVs contribute to cancer metastasis by educating
111 vesicles” (EVs) in metastasis. Cancer-derived EVs contribute to cancer metastasis by educating
primary and distant tumor environmental cells. In the metastatic process, cancer-derived EVs induce
112 primary and distant tumor environmental cells. In the metastatic process, cancer-derived EVs induce
angiogenesis or disrupt tight junctions in ECs, induce cancer-associated fibroblast (CAF) transition,
113 angiogenesis or disrupt tight junctions in ECs, induce cancer-associated fibroblast (CAF) transition,
suppress the host immune system, and create a premetastatic niche. Moreover, EVs-derived from
114 suppress the host immune system, and create a premetastatic niche. Moreover, EVs-derived from
cancer surrounding cells, such as CAFs and mesenchymal stem cells (MSCs), also contribute to
115 cancer surrounding cells, such as CAFs and mesenchymal stem cells (MSCs), also contribute to cancer
cancer metastasis.
116 metastasis.
3.1. Function of Cancer-Derived EVs in Angiogenesis
117 3.1. Function of Cancer-Derived EVs in Angiogenesis
Angiogenesis is a dynamic and tightly regulated process involving angiogenic factors, extracellular
118 matrix components,isand
Angiogenesis a endothelial
dynamic andcellstightly regulated
(ECs) [31]. process
Recently, involving
the role angiogenic
of cancer-derived EVsfactors,
in the
119 extracellular matrix components, and endothelial cells (ECs) [31]. Recently, the role of cancer-derived
process of angiogenesis in primary tumors and distant metastatic sites has attracted attention [32].
120 EVs in al.
Maji et theshowed
processthatof metastatic
angiogenesis in cancer-derived
breast primary tumors and
EVs, distant
which metastatic
highly expressedsites
Anxhas attracted
II, promoted
121 attention
angiogenesis [32].inMaji et al. showed
a Matrigel thatinmetastatic
plug assay vivo [33].breast
Anothercancer-derived
group showed EVs,
thatwhich highly from
EVs derived expressed
head
122 Anx II, promoted angiogenesis in a Matrigel plug assay in vivo [33]. Another group showed that EVs
123 derived from head and neck squamous cell carcinoma regulated in vitro and in vivo tumor
124 angiogenesis through ephrin-B reverse signaling [34]. Furthermore, Wang et al. demonstrated thatInt. J. Mol. Sci. 2020, 21, 4463 4 of 16
and neck squamous cell carcinoma regulated in vitro and in vivo tumor angiogenesis through ephrin-B
reverse signaling [34]. Furthermore, Wang et al. demonstrated that yes-associated protein (YAP),
which plays an important role in angiogenesis, was transferred to HUVECs via lung adenocarcinoma
cell-derived EVs [35]. Tight junctions between endothelial cells in brain capillaries are also critical to
the metastatic process of cancer cells [36]. Two groups reported that EVs secreted from metastatic
breast cancer cells disrupted the formation of tight junctions [37,38]. Thus, metastatic cancer-derived
EVs play a role in the promotion of angiogenesis and the destruction of vascular endothelial barriers,
which may lead to the promotion of cancer metastasis.
3.2. Effect of Cancer-Derived EVs on Host Immune Systems
During the metastatic cascade, tumor cells are exposed to the immune system, which can
recognize them and restrict their growth [1]. Recent studies have shown that cancer-derived EVs
mediate immunosuppression through various mechanisms [39]. Several reports have revealed that
cancer-derived EVs promote the expansion of regulatory T-cells (Tregs). Yen et al. showed that the
expression of transforming growth factor (TGF)-β1 in EVs derived from gastric cancer (GC) patients
and the ratio of Tregs were both significantly associated with the pathological stages and lymph node
metastasis of GC [40]. Their results suggested that immune surveillance can be modulated in GC
patients through the induction of Tregs. Another study reported that primary Lewis lung carcinoma
(LLC)-derived EVs activated toll-like receptor 3 (TLR3) in lung epithelial cells. Activated TLR3 in lung
epithelial cells recruits neutrophils and forms a premetastatic niche in the lung [41]. Cancer-derived EVs
also promote myeloid-derived suppressor cell (MDSC) activity, which assists cancer cell dissemination.
Huber et al. showed that melanoma-derived EVs mediated MDSC generation from monocytes [42].
Then, the authors incubated EVs purified from conditioned medium from a patient melanoma cell
line with monocytes isolated from the peripheral blood mononuclear cells of the same patient and
showed that the monocytes inhibited autologous T cell proliferation. In addition, it is well known that
cancer-derived EVs suppress natural killer (NK) cell and T cell activity to enhance immune evasion.
EVs derived from highly metastatic pancreatic cancer cells [43] and acute myeloid leukemia (AML) [44]
interfere with the antitumor activity of NK cells. Ricklefs et al. revealed that cancer-derived EVs
suppress T cell activity [45]. Moreover, EV-mediated T cell suppression is due to PD-L1, which is
expressed on the surface of cancer-derived EVs [46,47].
These studies suggest that EVs from cancer cells mediate the activation of the immune system to
promote cancer metastasis.
3.3. Crosstalk between Cancer Cells and Fibroblasts through EVs
Cancer cells educate fibroblasts through EVs, which leads to the progression of metastasis [48].
Cancer-derived EVs are involved in the reprogramming of normal stromal fibroblasts into activated
cancer-associated fibroblasts (CAFs) in chronic lymphocytic leukemia [49], hepatocellular carcinoma
(HCC) [50], and melanoma [51]. These studies revealed that the CAF transition leads to metastatic
niche formation and promotes cancer metastasis. Moreover, several reports have shown that EVs
derived from highly metastatic cancer cells are more likely to convert normal fibroblasts into CAFs
compared with those derived from non-metastatic cancer cells [50,52].
EVs derived from CAFs also play an important role in cancer metastasis. Kong et al. showed the
roles of CAF-derived EVs in premetastatic niche formation [53]. Moreover, Baroni et al. revealed that
miR-9 was important for the CAF transition in EVs derived from breast cancer cells [54]. The authors
also showed that EVs derived from miR-9-overexpressing normal fibroblasts were taken up by cancer
cells, in which they induced tumor cell aggressiveness by modulating genes involved in cell motility
and extracellular matrix (ECM) remodeling. These findings suggested that cancer cells and fibroblasts
engage in cross talk with each other via EVs to create a metastatic niche.Int. J. Mol. Sci. 2020, 21, 4463 5 of 16
3.4. Premetastatic Niche Formation by Cancer-Derived EVs
A premetastatic niche is a microenvironment prepared for the colonization of circulating tumor
cells in specific organs. It is known that premetastatic niche formation is promoted by angiogenesis,
CAF transition of fibroblasts, ECM remodeling, and immunosuppression, as mentioned above.
Cancer-derived EVs also influence premetastatic niche formation at distant sites. For example, Jung
et al. showed that EVs derived from CD44 variant isoform (CD44v6)-positive pancreatic cancer
cells contributed to the formation of a premetastatic niche in lymph nodes and lungs [55]. Similarly,
another study showed the effect of cancer-derived EVs on lung premetastatic niche formation. Grange
et al. reported that treatment with the intravenous injection of EVs derived from CD105-positive
renal cell carcinoma modified the lung microenvironment by enhancing the expression of vascular
endothelial growth factor (VEGF) and matrix metalloproteinase 2 (MMP2) in lung endothelial cells
and that of MMP9 in whole lung tissue [56]. In addition, Peinado et al. also showed the function of
cancer-derived EVs in lung premetastatic formation in mice. Intravenously injected EVs derived from
highly metastatic mouse melanoma cells were mainly distributed to the interstitium of the lung and bone
marrow [57]. Moreover, they found that EVs enhanced lung endothelial permeability and educated
bone marrow-derived cells, leading to the creation of a metastatic niche for lung metastasis. These
studies indicate that cancer-derived EVs may contribute to metastatic organotropism. Hoshino et al.
demonstrated the involvement of EVs as mediators of organotropism of metastasis. EVs derived
from a breast cancer cell subline that colonize the lung and brain could home into the corresponding
target organs after EV injection. Moreover, EVs significantly increased the lung metastatic capacity
of nonmetastatic breast cancer cells in the lung [58]. They also revealed that distant metastases were
associated with integrins on the EV surface. In addition, integrin expression patterns play a crucial role
in determining the metastatic site. For example, α6β4 and α6β1 are associated with lung metastasis,
while integrin αvβ5 is linked to liver metastasis [58]. Thus, cancer-derived EVs have the potential to
generate premetastatic niches and regulate metastatic organotropism.
3.5. Mesenchymal Stem Cell (MSC)-Derived EVs Contribute to Cancer Metastasis
EVs derived from cancer stromal cells have also been implicated in different stages of cancer
metastasis, especially epithelial-mesenchymal transition (EMT). EMT is a key process involved in
cancer cell metastasis, during which epithelial cells acquire mesenchymal characteristics and the
enhancement of cell motility and migration. The importance of MSCs present in the cancer stroma in
EMT induction has been well studied [59–62]. These studies demonstrated that treatment with MSC
culture medium or MSC coculture promoted EMT in breast or gastric cancer cells. Recently, several
studies have shown that MSC-derived EVs contain factors that stimulate EMT induction and promote
EMT and cancer metastasis. For example, Lin et al. investigated the effect of EVs derived from adipose
tissue-derived MSCs on breast cancer cells and demonstrated that EVs promoted the migration of
cancer cells via activation of the Wnt signaling pathway [63]. Similarly, Zhou et al. showed that
MSC-derived EVs promoted EMT in breast cancer cells. In this study, human umbilical cord-derived
MSCs promoted the invasion and migration of breast cancer cells and enhanced the induction of
EMT through the ERK pathway [64]. In addition, Zhao et al. found that the EMT-promoting effect
in lung cancer was mediated by EVs secreted from human umbilical cord-derived MSCs [65]. The
authors also showed that the knockdown of TGF-β in MSCs inhibited the EMT induction ability of
MSC-derived EVs. These reports suggest that MSCs present in the tumor stroma can promote EMT
induction through the secretion of EVs.
Collectively, these findings on the function of EVs derived from both cancer cells and stromal
cells, such as MSCs, in metastasis provide new insights into the potential clinical application of EVs in
the treatment of cancer metastasis.Int. J. Mol. Sci. 2020, 21, 4463 6 of 16
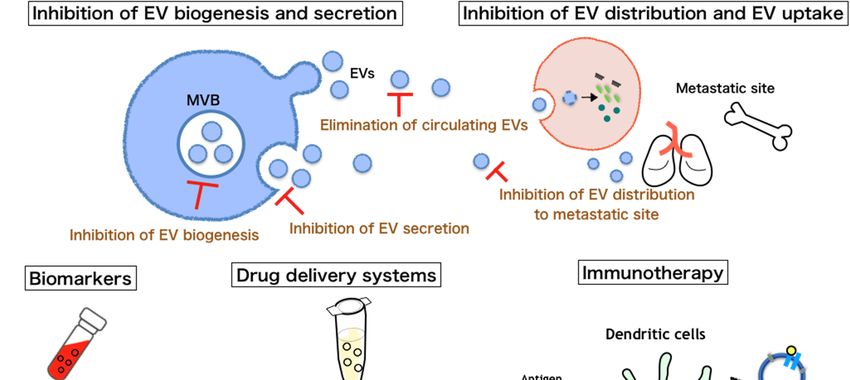
4. Therapeutic Implications of EVs
4.1. Inhibition of EV Biogenesis and Secretion
As mentioned above, EVs secreted by cancer cells play a pivotal role in promoting cancer metastasis.
Therefore, impairing the biogenesis and secretion of EVs by cancer cells might constitute a potential
strategy for cancer therapy.
Rab proteins have been shown to be involved in the production of EVs in both normal cells and
cancer cells [14,66,67]. Several Rabs have been reported to be involved in cancer development and
progression. Bobrie et al. showed that knockdown of Rab27b reduced the numbers of EVs in the culture
medium and inhibited the lung metastasis of 4T1 breast cancer cells in mice [68]. The formation and
secretion of EVs may be modulated through several approaches. GW4869, a nSMase2 inhibitor, inhibits
EV secretion and decreases the metastatic rate [69–71]. Fabbri et al. showed that the intravenous
injection of GW4869 significantly decreased the number of lung metastases in tumor-bearing mice [72].
This effect was partially rescued when LLC-derived EVs were injected into GW4869-treated mice [72].
Savina et al. showed that EV secretion is induced by an intracellular increase in calcium (Ca2+) [73].
To decrease the level of tumor-derived EV release, Chalmin et al. used dimethyl amiloride, which
blocks H+/Na+ and Na+/Ca2+ channels, in tumor-bearing mice. They showed that dimethyl amiloride
decreased the secretion of EVs from cancer cells by blocking the channels, which subsequently delayed
the growth of cancer cells [74]. Thus, targeting EV biogenesis and secretion may have potential
clinical implications for cancer metastatic therapy. However, to date, the mechanisms underlying
the biogenesis of different EV subtypes are extremely difficult to define. To target EV biogenesis and
secretion for clinical applications, more studies that allow the discrimination of EV subpopulations
are needed.
4.2. Inhibition of EV Uptake by Recipient Cells
Another option for cancer treatment is to capture and remove circulating cancer-derived EVs.
Nishida-Aoki et al. tried to capture circulating EVs derived from cancer cells by using antibodies
against human CD9 and CD63 [75]. The treatment of mice with anti-CD9 or anti-CD63 antibodies
stimulated EV removal by macrophages. Although this treatment had no effect on the primary tumor,
tumor metastasis was significantly reduced. Thus, the removal of cancer-derived EVs could be a
novel strategy for therapy for cancer metastasis. Targeting surface proteins on EVs can also inhibit the
distribution of EVs to distant sites, leading to blockade of the creation of a premetastatic niche. As we
mentioned before, Hoshino et al. showed a correlation between the specific integrins expressed on the
EV surface and metastatic tropism [58]. Therapeutic strategies targeting distinct integrin expression
patterns may significantly minimize EV uptake, which could inhibit cancer metastasis.
4.3. Immunotherapy
An extensive investigation has been carried out to determine the roles of EVs in immunological
reactions after Zitvogel et al. reported that EVs are loaded with tumor antigens [76]. Dendritic cell
(DC)-derived EVs are enriched in membrane proteins involved in antigen presentation, including MHC
class I and II molecules, MHC class I-like molecule (CD1), and costimulatory molecules (CD80 and
CD86), which induce T-cell activation [77,78]. Therefore, their incorporation by cancer cells could turn
them into immunogenic targets. In fact, DC-derived EVs have been used in phase I trials in melanoma,
colorectal cancer (CRC), and non-small-cell lung cancer (NSCLC) patients [79–82]. These trials
demonstrated the safety of the use of DC-derived EVs in antitumor treatments in patients. Besse et al.
showed the capacity of DC-derived EVs to boost NK cell antitumor immunity in patients with advanced
NSCLC in a phase II clinical trial [79]. Thus, the activation of antitumor-specific T cell responses by
DC-derived EVs has been proposed to play a key role in the suppression of established tumor growth.
Recently, immunotherapy has shown effectiveness in patients with metastatic cancers [83]. Therefore,
EV-based immunotherapy could also effectively prevent or inhibit cancer metastasis.Int. J. Mol. Sci. 2020, 21, 4463 7 of 16
4.4. Encapsulation
EVs have emerged as delivery vesicles for small RNAs (siRNAs and miRNAs) for therapeutic
applications due to their natural role in intercellular RNA transport. In addition, as cancer-derived EVs
have a property of organ tropism in relation to metastasis [58], EVs may efficiently reach metastatic
sites. Therefore, it is important to develop techniques for the encapsulation of drugs that inhibit the
formation of the metastatic niche by EVs and for the delivery of EVs to the metastatic site. Several
studies succeeded in encapsulating small molecules, such as siRNAs, and showed that the siRNAs
could be delivered to recipient cells. Alvarez-Erviti et al. used EVs from self-derived DCs that express
Lamp2, a membrane protein in EVs, fused to the neuron-specific rabies virus glycoprotein (RVG)
peptide for siRNA delivery. The intravenously injected EVs delivered glyceraldehyde 3-phosphate
dehydrogenase (GAPDH) siRNA specifically to neurons, microglia, and oligodendrocytes in the brain,
resulting in specific gene knockdown [84]. They also showed that siRNA against beta-secretase 1
(BACE1) encapsulated in EVs was successfully delivered to neurons and downregulated BACE1
expression [84]. Similarly, Wahlgren et al. tried to use EVs originating from the peripheral blood
of healthy donors as carriers of genetic material to target cells. EVs loaded with siRNAs against
mitogen-activated protein kinase (MAPK) were able to efficiently knock down the target gene upon
their delivery into in vitro monocytes and lymphocytes [85]. In another study, Shtam et al. targeted
RAD51 genes with specific siRNAs delivered via EVs derived from HeLa cells and ascitic fluids because
they previously revealed that the decrease in the RAD51 protein level induced massive reproductive
cell death in most cancer cell types [86]. They showed that EVs effectively delivered siRNAs against
RAD51 into target cells, causing selective gene silencing and leading to reproductive cancer cell death
by knockdown of RAD51 [87]. These studies used siRNAs encapsulated by the electroporation method.
However, electroporation has been reported to show inefficiency in loading EVs with siRNAs due to
protein aggregation [88]. Therefore, the optimization of the electroporation method has been performed
recently [89,90]. Recently, some research reported that EVs can be used for chemical antitumor drug
delivery. Pascucci showed that EVs containing paclitaxel inhibited in vitro tumor cell growth [91].
Another report showed that engineered EVs were injected intravenously into mice and delivered
doxorubicin to tumors, resulting in the inhibition of tumor growth [92].
Thus, EVs loaded with small RNAs and anticancer drugs could be a useful material for drug
delivery. Research on EVs for clinical use is in the early stage of development. Therefore, much
effort needs to be made to guarantee their safe and effective application for therapeutic use. Based on
these studies, targeting the metastatic niche using EV-based drug delivery systems could be a new
therapeutic approach to combat cancer metastasis.
4.5. Biomarkers of Cancer Metastasis
EVs can be easily detected in a variety of body fluids, including blood [93,94], urine [95], and
saliva [96,97]. EVs released into body fluids may provide an excellent biomarker for monitoring
the emergence, progression, and prognosis of cancer, as well as predicting the efficacy of treatment
regimens. In fact, there are many reports regarding the use of EVs as cancer biomarkers [98–101].
Recently, a correlation between cancer metastasis and cargos in circulating EVs was reported. Several
studies have shown the potential of specific EV-associated miRNAs as biomarkers for metastatic
cancer patients. Bryant et al. found an association between miR-141 and miR-375 levels in EVs
and metastatic prostate cancer [102]. Tokuhisa et al. investigated the miRNA profiles of EVs
derived from peritoneal lavage fluid (PLF) samples from GC patients with or without peritoneal
dissemination. miR-21 and miR-1225-5p levels in the PLF-derived EVs of GC patients with the highest
incidence of peritoneal metastasis were significantly increased compared to those of patients without
metastasis [103]. In another study focusing on EVs in PLF, the upregulation of EV-associated miRs
(miR-21-5p, miR-92a-3p, miR-223-3p, and miR-342-3p) was observed in GC patients with peritoneal
metastasis [104]. miR-21 expression in plasma-derived EVs was also positively correlated with liver
metastasis in CRC patients [105]. Tang et al. showed that miR-320d could be a useful biomarker forInt. J. Mol. Sci. 2020, 21, 4463 8 of 16
distinguishing metastatic from nonmetastatic CRC. In addition, the expression levels of miR-320d
were not associated with background factors, such as age, sex, drinking status, and histological
type [106]. Other small RNAs in EVs also reflect metastatic conditions. Chen et al. compared urothelial
carcinoma of the bladder (UCB) tissue and matched nontumor tissue samples by using circular RNA
(circRNA) microarrays to identify UCB-specific circRNAs. Then, they identified a novel small RNA,
which was termed circPRMT5. Moreover, upregulated expression of circPRMT5 in the tumor tissue
was more likely to occur in patients with lymph node metastasis than in patients without metastasis
in EVs derived from serum and urine [107]. Liu et al. showed that the increased expression of
the EV-associated lncRNA CRNDE-h was significantly correlated with CRC regional lymph node
metastasis and distant metastasis [108]. Few studies have reported that proteins on the EV surface also
have potential as biomarkers for cancer metastasis. For example, Wang et al. found that, compared
with patients with nonmetastatic NSCLC and healthy donors, patients with metastatic NSCLC showed
higher expression levels of lipopolysaccharide-binding proteins [109].
Thus, EV-associated RNAs and proteins show potential as biomarkers for cancer metastasis
(Table 1). Compared with traditional biomarkers, EVs have some advantages as liquid biomarkers.
First, as EVs can be detected from patient body fluids, detection could be noninvasive. Second, EVs are
protected from degradation by protein nucleases in the phospholipid bilayer.
Table 1. Biomarkers for cancer metastasis in EVs.
Isolation Marker
Primary Cancer Metastatic Site EV Source Biomarker Reference No.
Method Detection
1.2 µm miR-141 and
Prostate cancer Not mentioned Plasma qRT-PCR [102]
filtration miR-375
Peritoneal Peritoneal miR-21 and
Gastric cancer UC qRT-PCR [103]
dissemination lavage fluid miR-1225-5p
miR-21-5p,
miR-92a-3p,
Peritoneal
Gastric cancer Peritoneal metastasis UC miR-223-3p, qRT-PCR [104]
lavage fluid
and
miR-342-3p
Colorectal cancer Liver metastasis Plasma UC miR-21 tCLN [105]
Colorectal cancer Distant metastasis Serum UC miR-320d qRT-PCR [106]
Urothelial carcinoma Lymph node Serum and
UC circPRMT5 qRT-PCR [107]
of the bladder metastasis urine
Lymph node
Colorectal cancer metastasis Serum ExoQuick CRNDE-h qRT-PCR [108]
Distant metastasis
tCLN Tethered Cationic Lipoplex Nanoparticles, UC Ultracentrifugation.
Recently, several commercially available tools, such as ExoDx Prostate kit (IntelliScore), have
been used as noninvasive risk assessment tools for the detection of high-grade cancer. The kit assays
the gene expression of urine EVs by measuring a combination of ETS-related gene (ERG), prostate
cancer antigen (PCA)3, and SAM pointed domain containing ETS transcription factor (SPDEF), which
detects and stratifies by risk cases of prostate cancer. Moreover, several clinical trials of EV-based
biomarkers in cancer diagnosis have been reported [110–112] and registered (clinicaltrial.gov, accessed
on June 17, 2020). Although the clinical usage of EVs as biomarkers has not yet been fully validated
and explored, EVs derived from body fluids could be a good source of biomarkers for the detection of
cancer metastasis in the future.
5. Conclusions and Perspectives
Over the past few years, many studies have revealed that cancer-derived EVs play a crucial role
in cancer metastasis, as mentioned above (Figure 2). In fact, inhibition of EV biogenesis, secretion,
distribution, and uptake significantly reduces cancer metastasis in vivo [58,68,69,75]. In addition, the
treatment of DC-derived EVs boosts the antitumor immunity of NK cells in cancer patients, though notInt. J. Mol. Sci. 2020, 21, x FOR PEER REVIEW 9 of 16
349 Int. J. Mol. Sci.
expected to 2020, 21, 4463
be good delivery materials for therapeutic cargo, such as small RNAs and antitumor 9 of 16
350 drugs, for cancer treatment. However, an efficient EV purification technique and an effective method
351 for
with loading specific
metastasis [79].cargos into EVs are required
As immunotherapy for in
is effective thepatients
use of with
EVs as delivery cancers,
metastatic tools. InDC-derived
the future,
352 these problems need to be solved.
EV-based immunotherapy is expected to treat cancer metastasis. Moreover, many studies show that
353 Moreover, EVs can function as potential biomarkers in the diagnosis and prognosis of cancer
EVs can efficiently deliver different kinds of cargo to the target cell. Therefore, EVs are expected to be
354 metastasis. In particular, EV-associated miRNAs are well documented to reflect the presence of
good delivery materials for therapeutic cargo, such as small RNAs and antitumor drugs, for cancer
355 cancer metastasis [98,113,114]. However, for clinical usage, screens of more specific markers and
treatment. However, an efficient EV purification technique and an effective method for loading specific
356 prospective studies are needed. Although, currently, many clinical trials utilizing EV-based
cargos into EVs are required for the use of EVs as delivery tools. In the future, these problems need to
357 technologies in cancer diagnostics are underway compared to those involving cancer therapy, EVs
be solved.
358 are very promising tools for the diagnosis and also the treatment of cancer metastasis in the future.
359
360 Figure 2.
Figure PossibleEV
2. Possible EVengineering
engineeringfor
forcancer
cancertherapy.
therapy. EVs
EVssecreted
secretedby
bycancer
cancer cells
cells have
have aa pivotal
pivotal role
role
in promoting cancer metastasis. Therefore, these EVs could be useful as delivery tools or biomarkers
361 in promoting cancer metastasis. Therefore, these EVs could be useful as delivery tools or biomarkers
for patients with cancer metastasis.
362 for patients with cancer metastasis.
Moreover, EVs can function as potential biomarkers in the diagnosis and prognosis of cancer
363 Author Contributions: A.K., Y.Y., and T.O. drafted the manuscript. Y.Y. and T.O. reviewed the manuscript, and
364 metastasis.
Y.Y. Inthe
approved particular,
submittedEV-associated
manuscript. AllmiRNAs are well
authors read and documented to reflect
approved the final the presence of cancer
manuscript.
metastasis [98,113,114]. However, for clinical usage, screens of more specific markers and prospective
365 Funding: This work was supported by a Grant-in-Aid for Early Career Scientists from the Japan Society for the
studies are needed. Although, currently, many clinical trials utilizing EV-based technologies in cancer
366 Promotion of Science (No. 19K16848) and CREST from the Japan Science and Technology Agency (No.
diagnostics are underway compared to those involving cancer therapy, EVs are very promising tools
367 JPMJCR19H1).
for the diagnosis and also the treatment of cancer metastasis in the future.
368 Conflicts of Interest: The authors declare no conflict of interest.
Author Contributions: A.K., Y.Y., and T.O. drafted the manuscript. Y.Y. and T.O. reviewed the manuscript, and Y.Y.
369 approved the submitted manuscript. All authors have read and agreed to the published version of the manuscript.
Abbreviations
Funding: This work was supported by a Grant-in-Aid for Early Career Scientists from the Japan Society for
EVsPromotion
the extracellular
of Science vesicles
(No. 19K16848) and CREST from the Japan Science and Technology Agency (No.
miRNAs
JPMJCR19H1).microRNAs
MHC of Interest:
Conflicts The authors declare
major histocompatibility no conflict of interest.
complex
MVBs multivesicular bodies
nSMase2 neutral sphingomyelinase 2
ESCRT endosomal sorting complex required for transport
UC ultracentrifugationInt. J. Mol. Sci. 2020, 21, 4463 10 of 16
Abbreviations
EVs extracellular vesicles
miRNAs microRNAs
MHC major histocompatibility complex
MVBs multivesicular bodies
nSMase2 neutral sphingomyelinase 2
ESCRT endosomal sorting complex required for transport
UC ultracentrifugation
SEC size exclusion chromatography
ECs endothelial cells
YAP Yes-associated protein
Tregs regulatory T-cells
TGF transforming growth factor
GC gastric cancer
LLC Lewis lung carcinoma
TLR toll-like receptor
MDSC myeloid-derived suppressor cell
NK natural killer
AML acute myeloid leukemia
CAFs cancer-associated fibroblasts
HCC hepatocellular carcinoma
ECM extracellular matrix
VEGF vascular endothelial growth factor
MMP matrix metalloproteinase
MSC mesenchymal stem cell
EMT epithelial-mesenchymal transition
DC dendritic cell
CRC colorectal cancer
NSCLC non-small-cell lung cancer
RVG rabies virus glycoprotein
GAPDH glyceraldehyde 3-phosphate dehydrogenase
BACE1 beta-secretase 1
MAPK mitogen-activated protein kinase
PLF peritoneal lavage fluid
UCB urothelial carcinoma of the bladder
circRNA circular RNA
ERG ETS-related gene
PCA prostate cancer antigen
SAM pointed domain containing ETS transcription
SPDEF
factor
References
1. Steeg, P.S. Targeting metastasis. Nat. Rev. Cancer 2016, 16, 201–218. [CrossRef] [PubMed]
2. Anderson, R.L.; Balasas, T.; Callaghan, J.; Coombes, R.C.; Evans, J.; Hall, J.A.; Kinrade, S.; Jones, D.; Jones, P.S.;
Jones, R.; et al. A framework for the development of effective anti-metastatic agents. Nat. Rev. Clin. Oncol
2019, 16, 185–204. [CrossRef] [PubMed]
3. Hanahan, D.; Weinberg, R.A. Hallmarks of cancer: The next generation. Cell 2011, 144, 646–674. [CrossRef]
4. Joyce, J.A.; Pollard, J.W. Microenvironmental regulation of metastasis. Nat. Rev. Cancer 2009, 9, 239–252.
[CrossRef]
5. Raposo, G.; Nijman, H.W.; Stoorvogel, W.; Liejendekker, R.; Harding, C.V.; Melief, C.J.; Geuze, H.J. B
lymphocytes secrete antigen-presenting vesicles. J. Exp. Med. 1996, 183, 1161–1172. [CrossRef]Int. J. Mol. Sci. 2020, 21, 4463 11 of 16
6. Valadi, H.; Ekstrom, K.; Bossios, A.; Sjostrand, M.; Lee, J.J.; Lotvall, J.O. Exosome-mediated transfer of mRNAs and
microRNAs is a novel mechanism of genetic exchange between cells. Nat. Cell Biol 2007, 9, 654–659. [CrossRef]
[PubMed]
7. Kosaka, N.; Iguchi, H.; Yoshioka, Y.; Takeshita, F.; Matsuki, Y.; Ochiya, T. Secretory mechanisms and
intercellular transfer of microRNAs in living cells. J. Biol Chem 2010, 285, 17442–17452. [CrossRef] [PubMed]
8. Pegtel, D.M.; Cosmopoulos, K.; Thorley-Lawson, D.A.; van Eijndhoven, M.A.; Hopmans, E.S.; Lindenberg, J.L.;
de Gruijl, T.D.; Wurdinger, T.; Middeldorp, J.M. Functional delivery of viral miRNAs via exosomes. Proc.
Natl. Acad. Sci. USA 2010, 107, 6328–6333. [CrossRef] [PubMed]
9. Zhang, Y.; Liu, D.; Chen, X.; Li, J.; Li, L.; Bian, Z.; Sun, F.; Lu, J.; Yin, Y.; Cai, X.; et al. Secreted monocytic
miR-150 enhances targeted endothelial cell migration. Mol. Cell 2010, 39, 133–144. [CrossRef]
10. Azmi, A.S.; Bao, B.; Sarkar, F.H. Exosomes in cancer development, metastasis, and drug resistance: A
comprehensive review. Cancer Metastasis Rev. 2013, 32, 623–642. [CrossRef] [PubMed]
11. Becker, A.; Thakur, B.K.; Weiss, J.M.; Kim, H.S.; Peinado, H.; Lyden, D. Extracellular Vesicles in Cancer:
Cell-to-Cell Mediators of Metastasis. Cancer Cell 2016, 30, 836–848. [CrossRef] [PubMed]
12. Esposito, M.; Kang, Y. Targeting tumor-stromal interactions in bone metastasis. Pharmacol Ther 2014, 141,
222–233. [CrossRef]
13. Zhang, C.; Ji, Q.; Yang, Y.; Li, Q.; Wang, Z. Exosome: Function and Role in Cancer Metastasis and Drug
Resistance. Technol. Cancer Res. Treat. 2018, 17, 1533033818763450. [CrossRef] [PubMed]
14. Raposo, G.; Stoorvogel, W. Extracellular vesicles: Exosomes, microvesicles, and friends. J. Cell. Biol. 2013,
200, 373–383. [CrossRef] [PubMed]
15. Thery, C.; Zitvogel, L.; Amigorena, S. Exosomes: Composition, biogenesis and function. Nat. Rev. Immunol.
2002, 2, 569–579. [CrossRef]
16. Witwer, K.W.; Buzas, E.I.; Bemis, L.T.; Bora, A.; Lasser, C.; Lotvall, J.; Nolte-’t Hoen, E.N.; Piper, M.G.;
Sivaraman, S.; Skog, J.; et al. Standardization of sample collection, isolation and analysis methods in
extracellular vesicle research. J. Extracell. Vesicles 2013, 2. [CrossRef]
17. Yanez-Mo, M.; Siljander, P.R.; Andreu, Z.; Zavec, A.B.; Borras, F.E.; Buzas, E.I.; Buzas, K.; Casal, E.; Cappello, F.;
Carvalho, J.; et al. Biological properties of extracellular vesicles and their physiological functions. J. Extracell.
Vesicles 2015, 4, 27066. [CrossRef]
18. Hurley, J.H. ESCRT complexes and the biogenesis of multivesicular bodies. Curr. Opin. Cell. Biol. 2008, 20, 4–11.
[CrossRef]
19. Heijnen, H.F.; Schiel, A.E.; Fijnheer, R.; Geuze, H.J.; Sixma, J.J. Activated platelets release two types
of membrane vesicles: Microvesicles by surface shedding and exosomes derived from exocytosis of
multivesicular bodies and alpha-granules. Blood 1999, 94, 3791–3799. [CrossRef]
20. Muralidharan-Chari, V.; Clancy, J.W.; Sedgwick, A.; D’Souza-Schorey, C. Microvesicles: Mediators of
extracellular communication during cancer progression. J. Cell. Sci. 2010, 123, 1603–1611. [CrossRef]
21. Thery, C.; Boussac, M.; Veron, P.; Ricciardi-Castagnoli, P.; Raposo, G.; Garin, J.; Amigorena, S. Proteomic
analysis of dendritic cell-derived exosomes: A secreted subcellular compartment distinct from apoptotic
vesicles. J. Immunol. 2001, 166, 7309–7318. [CrossRef] [PubMed]
22. Thery, C.; Amigorena, S.; Raposo, G.; Clayton, A. Isolation and characterization of exosomes from cell culture
supernatants and biological fluids. Curr. Protoc. Cell. Biol. 2006, 30, 3.22.1–3.22.29. [CrossRef]
23. Gardiner, C.; Di Vizio, D.; Sahoo, S.; Thery, C.; Witwer, K.W.; Wauben, M.; Hill, A.F. Techniques used for the
isolation and characterization of extracellular vesicles: Results of a worldwide survey. J. Extracell. Vesicles
2016, 5, 32945. [CrossRef] [PubMed]
24. Heath, N.; Grant, L.; De Oliveira, T.M.; Rowlinson, R.; Osteikoetxea, X.; Dekker, N.; Overman, R. Rapid
isolation and enrichment of extracellular vesicle preparations using anion exchange chromatography. Sci.
Rep. 2018, 8, 5730. [CrossRef] [PubMed]
25. An, M.; Wu, J.; Zhu, J.; Lubman, D.M. Comparison of an Optimized Ultracentrifugation Method versus Size-Exclusion
Chromatography for Isolation of Exosomes from Human Serum. J. Proteome Res. 2018, 17, 3599–3605. [CrossRef]
[PubMed]
26. Lobb, R.J.; Becker, M.; Wen, S.W.; Wong, C.S.; Wiegmans, A.P.; Leimgruber, A.; Moller, A. Optimized
exosome isolation protocol for cell culture supernatant and human plasma. J. Extracell. Vesicles 2015, 4, 27031.
[CrossRef]Int. J. Mol. Sci. 2020, 21, 4463 12 of 16
27. Zarovni, N.; Corrado, A.; Guazzi, P.; Zocco, D.; Lari, E.; Radano, G.; Muhhina, J.; Fondelli, C.; Gavrilova, J.;
Chiesi, A. Integrated isolation and quantitative analysis of exosome shuttled proteins and nucleic acids using
immunocapture approaches. Methods 2015, 87, 46–58. [CrossRef]
28. Ramirez, M.I.; Amorim, M.G.; Gadelha, C.; Milic, I.; Welsh, J.A.; Freitas, V.M.; Nawaz, M.; Akbar, N.; Couch, Y.;
Makin, L.; et al. Technical challenges of working with extracellular vesicles. Nanoscale 2018, 10, 881–906. [CrossRef]
29. Phan, J.; Kumar, P.; Hao, D.; Gao, K.; Farmer, D.; Wang, A. Engineering mesenchymal stem cells to improve
their exosome efficacy and yield for cell-free therapy. J. Extracell. Vesicles 2018, 7, 1522236. [CrossRef]
30. Yan, I.K.; Shukla, N.; Borrelli, D.A.; Patel, T. Use of a Hollow Fiber Bioreactor to Collect Extracellular Vesicles
from Cells in Culture. Methods Mol. Biol. 2018, 1740, 35–41.
31. Carmeliet, P.; Jain, R.K. Angiogenesis in cancer and other diseases. Nature 2000, 407, 249–257. [CrossRef]
[PubMed]
32. Kikuchi, S.; Yoshioka, Y.; Prieto-Vila, M.; Ochiya, T. Involvement of Extracellular Vesicles in Vascular-Related
Functions in Cancer Progression and Metastasis. Int J. Mol. Sci. 2019, 20, 2584. [CrossRef] [PubMed]
33. Maji, S.; Chaudhary, P.; Akopova, I.; Nguyen, P.M.; Hare, R.J.; Gryczynski, I.; Vishwanatha, J.K. Exosomal
Annexin II Promotes Angiogenesis and Breast Cancer Metastasis. Mol. Cancer Res. 2017, 15, 93–105.
[CrossRef] [PubMed]
34. Sato, S.; Vasaikar, S.; Eskaros, A.; Kim, Y.; Lewis, J.S.; Zhang, B.; Zijlstra, A.; Weaver, A.M. EPHB2 carried on
small extracellular vesicles induces tumor angiogenesis via activation of ephrin reverse signaling. JCI Insight
2019, 4, e132447. [CrossRef]
35. Wang, Y.; Dong, L.; Zhong, H.; Yang, L.; Li, Q.; Su, C.; Gu, W.; Qian, Y. Extracellular Vesicles (EVs) from Lung
Adenocarcinoma Cells Promote Human Umbilical Vein Endothelial Cell (HUVEC) Angiogenesis through
Yes Kinase-associated Protein (YAP) Transport. Int J. Biol. Sci. 2019, 15, 2110–2118. [CrossRef]
36. Ballabh, P.; Braun, A.; Nedergaard, M. The blood-brain barrier: An overview: Structure, regulation, and
clinical implications. Neurobiol. Dis. 2004, 16, 1–13. [CrossRef] [PubMed]
37. Zhou, W.; Fong, M.Y.; Min, Y.; Somlo, G.; Liu, L.; Palomares, M.R.; Yu, Y.; Chow, A.; O’Connor, S.T.; Chin, A.R.;
et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell 2014,
25, 501–515. [CrossRef] [PubMed]
38. Tominaga, N.; Kosaka, N.; Ono, M.; Katsuda, T.; Yoshioka, Y.; Tamura, K.; Lotvall, J.; Nakagama, H.; Ochiya, T.
Brain metastatic cancer cells release microRNA-181c-containing extracellular vesicles capable of destructing
blood-brain barrier. Nat. Commun. 2015, 6, 6716. [CrossRef]
39. Jella, K.K.; Nasti, T.H.; Li, Z.; Malla, S.R.; Buchwald, Z.S.; Khan, M.K. Exosomes, Their Biogenesis and Role
in Inter-Cellular Communication, Tumor Microenvironment and Cancer Immunotherapy. Vaccines 2018, 6,
69. [CrossRef] [PubMed]
40. Yen, E.-Y.; Miaw, S.-C.; Yu, J.-S.; Lai, I.-R. Exosomal TGF-β1 is correlated with lymphatic metastasis of gastric
cancers. Am. J. Cancer Res. 2017, 7, 2199–2208.
41. Liu, Y.; Gu, Y.; Han, Y.; Zhang, Q.; Jiang, Z.; Zhang, X.; Huang, B.; Xu, X.; Zheng, J.; Cao, X. Tumor Exosomal
RNAs Promote Lung Pre-metastatic Niche Formation by Activating Alveolar Epithelial TLR3 to Recruit
Neutrophils. Cancer Cell 2016, 30, 243–256. [CrossRef]
42. Huber, V.; Vallacchi, V.; Fleming, V.; Hu, X.; Cova, A.; Dugo, M.; Shahaj, E.; Sulsenti, R.; Vergani, E.;
Filipazzi, P.; et al. Tumor-derived microRNAs induce myeloid suppressor cells and predict immunotherapy
resistance in melanoma. J. Clin. Investig. 2018, 128, 5505–5516. [CrossRef] [PubMed]
43. Zhao, J.; Schlosser, H.A.; Wang, Z.; Qin, J.; Li, J.; Popp, F.; Popp, M.C.; Alakus, H.; Chon, S.H.; Hansen, H.P.; et al.
Tumor-Derived Extracellular Vesicles Inhibit Natural Killer Cell Function in Pancreatic Cancer. Cancers 2019,
11, 874. [CrossRef] [PubMed]
44. Hong, C.S.; Sharma, P.; Yerneni, S.S.; Simms, P.; Jackson, E.K.; Whiteside, T.L.; Boyiadzis, M. Circulating
exosomes carrying an immunosuppressive cargo interfere with cellular immunotherapy in acute myeloid
leukemia. Sci. Rep. 2017, 7, 14684. [CrossRef] [PubMed]
45. Ricklefs, F.L.; Alayo, Q.; Krenzlin, H.; Mahmoud, A.B.; Speranza, M.C.; Nakashima, H.; Hayes, J.L.; Lee, K.;
Balaj, L.; Passaro, C.; et al. Immune evasion mediated by PD-L1 on glioblastoma-derived extracellular
vesicles. Sci. Adv. 2018, 4, eaar2766. [CrossRef] [PubMed]
46. Poggio, M.; Hu, T.; Pai, C.C.; Chu, B.; Belair, C.D.; Chang, A.; Montabana, E.; Lang, U.E.; Fu, Q.; Fong, L.; et al.
Suppression of Exosomal PD-L1 Induces Systemic Anti-tumor Immunity and Memory. Cell 2019, 177, 414–427.e413.
[CrossRef] [PubMed]Int. J. Mol. Sci. 2020, 21, 4463 13 of 16
47. Chen, G.; Huang, A.C.; Zhang, W.; Zhang, G.; Wu, M.; Xu, W.; Yu, Z.; Yang, J.; Wang, B.; Sun, H.; et al.
Exosomal PD-L1 contributes to immunosuppression and is associated with anti-PD-1 response. Nature 2018,
560, 382–386. [CrossRef] [PubMed]
48. De Wever, O.; Van Bockstal, M.; Mareel, M.; Hendrix, A.; Bracke, M. Carcinoma-associated fibroblasts provide
operational flexibility in metastasis. Semin. Cancer Biol. 2014, 25, 33–46. [CrossRef]
49. Paggetti, J.; Haderk, F.; Seiffert, M.; Janji, B.; Distler, U.; Ammerlaan, W.; Kim, Y.J.; Adam, J.; Lichter, P.;
Solary, E.; et al. Exosomes released by chronic lymphocytic leukemia cells induce the transition of stromal
cells into cancer-associated fibroblasts. Blood 2015, 126, 1106–1117. [CrossRef]
50. Fang, T.; Lv, H.; Lv, G.; Li, T.; Wang, C.; Han, Q.; Yu, L.; Su, B.; Guo, L.; Huang, S.; et al. Tumor-derived
exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer.
Nat. Commun. 2018, 9, 191. [CrossRef]
51. Gener Lahav, T.; Adler, O.; Zait, Y.; Shani, O.; Amer, M.; Doron, H.; Abramovitz, L.; Yofe, I.; Cohen, N.;
Erez, N. Melanoma-derived extracellular vesicles instigate proinflammatory signaling in the metastatic
microenvironment. Int. J. Cancer 2019, 145, 2521–2534. [CrossRef] [PubMed]
52. Naito, Y.; Yamamoto, Y.; Sakamoto, N.; Shimomura, I.; Kogure, A.; Kumazaki, M.; Yokoi, A.; Yashiro, M.;
Kiyono, T.; Yanagihara, K.; et al. Cancer extracellular vesicles contribute to stromal heterogeneity by inducing
chemokines in cancer-associated fibroblasts. Oncogene 2019, 38, 5566–5579. [CrossRef] [PubMed]
53. Kong, J.; Tian, H.; Zhang, F.; Zhang, Z.; Li, J.; Liu, X.; Li, X.; Liu, J.; Li, X.; Jin, D.; et al. Extracellular vesicles
of carcinoma-associated fibroblasts creates a pre-metastatic niche in the lung through activating fibroblasts.
Mol. Cancer 2019, 18, 175. [CrossRef] [PubMed]
54. Baroni, S.; Romero-Cordoba, S.; Plantamura, I.; Dugo, M.; D’Ippolito, E.; Cataldo, A.; Cosentino, G.;
Angeloni, V.; Rossini, A.; Daidone, M.G.; et al. Exosome-mediated delivery of miR-9 induces cancer-associated
fibroblast-like properties in human breast fibroblasts. Cell Death Dis. 2016, 7, e2312. [CrossRef]
55. Jung, T.; Castellana, D.; Klingbeil, P.; Cuesta Hernandez, I.; Vitacolonna, M.; Orlicky, D.J.; Roffler, S.R.;
Brodt, P.; Zoller, M. CD44v6 dependence of premetastatic niche preparation by exosomes. Neoplasia 2009, 11,
1093–1105. [CrossRef]
56. Grange, C.; Tapparo, M.; Collino, F.; Vitillo, L.; Damasco, C.; Deregibus, M.C.; Tetta, C.; Bussolati, B.;
Camussi, G. Microvesicles released from human renal cancer stem cells stimulate angiogenesis and formation
of lung premetastatic niche. Cancer Res. 2011, 71, 5346–5356. [CrossRef]
57. Peinado, H.; Aleckovic, M.; Lavotshkin, S.; Matei, I.; Costa-Silva, B.; Moreno-Bueno, G.;
Hergueta-Redondo, M.; Williams, C.; Garcia-Santos, G.; Ghajar, C.; et al. Melanoma exosomes educate bone
marrow progenitor cells toward a pro-metastatic phenotype through MET. Nat. Med. 2012, 18, 883–891.
[CrossRef]
58. Hoshino, A.; Costa-Silva, B.; Shen, T.L.; Rodrigues, G.; Hashimoto, A.; Tesic Mark, M.; Molina, H.; Kohsaka, S.;
Di Giannatale, A.; Ceder, S.; et al. Tumour exosome integrins determine organotropic metastasis. Nature
2015, 527, 329–335. [CrossRef]
59. Xu, Q.; Wang, L.; Li, H.; Han, Q.; Li, J.; Qu, X.; Huang, S.; Zhao, R.C. Mesenchymal stem cells play a potential
role in regulating the establishment and maintenance of epithelial-mesenchymal transition in MCF7 human
breast cancer cells by paracrine and induced autocrine TGF-beta. Int. J. Oncol. 2012, 41, 959–968. [CrossRef]
60. Martin, F.T.; Dwyer, R.M.; Kelly, J.; Khan, S.; Murphy, J.M.; Curran, C.; Miller, N.; Hennessy, E.; Dockery, P.;
Barry, F.P.; et al. Potential role of mesenchymal stem cells (MSCs) in the breast tumour microenvironment:
Stimulation of epithelial to mesenchymal transition (EMT). Breast Cancer Res. Treat. 2010, 124, 317–326.
[CrossRef] [PubMed]
61. Xue, Z.; Wu, X.; Chen, X.; Liu, Y.; Wang, X.; Wu, K.; Nie, Y.; Fan, D. Mesenchymal stem cells promote epithelial
to mesenchymal transition and metastasis in gastric cancer though paracrine cues and close physical contact.
J. Cell Biochem. 2015, 116, 618–627. [CrossRef]
62. Kletukhina, S.; Neustroeva, O.; James, V.; Rizvanov, A.; Gomzikova, M. Role of Mesenchymal Stem
Cell-Derived Extracellular Vesicles in Epithelial-Mesenchymal Transition. Int. J. Mol. Sci. 2019, 20, 4813.
[CrossRef] [PubMed]
63. Lin, R.; Wang, S.; Zhao, R.C. Exosomes from human adipose-derived mesenchymal stem cells promote
migration through Wnt signaling pathway in a breast cancer cell model. Mol. Cell Biochem. 2013, 383, 13–20.
[CrossRef] [PubMed]Int. J. Mol. Sci. 2020, 21, 4463 14 of 16
64. Zhou, X.; Li, T.; Chen, Y.; Zhang, N.; Wang, P.; Liang, Y.; Long, M.; Liu, H.; Mao, J.; Liu, Q.; et al. Mesenchymal
stem cellderived extracellular vesicles promote the in vitro proliferation and migration of breast cancer cells
through the activation of the ERK pathway. Int. J. Oncol. 2019, 54, 1843–1852.
65. Zhao, X.; Wu, X.; Qian, M.; Song, Y.; Wu, D.; Zhang, W. Knockdown of TGF-beta1 expression in human
umbilical cord mesenchymal stem cells reverts their exosome-mediated EMT promoting effect on lung cancer
cells. Cancer Lett. 2018, 428, 34–44. [CrossRef] [PubMed]
66. Hendrix, A.; De Wever, O. Rab27 GTPases distribute extracellular nanomaps for invasive growth and
metastasis: Implications for prognosis and treatment. Int. J. Mol. Sci. 2013, 14, 9883–9892. [CrossRef]
67. Li, Z.; Fang, R.; Fang, J.; He, S.; Liu, T. Functional implications of Rab27 GTPases in Cancer. Cell Commun.
Signal. 2018, 16, 44. [CrossRef]
68. Bobrie, A.; Krumeich, S.; Reyal, F.; Recchi, C.; Moita, L.F.; Seabra, M.C.; Ostrowski, M.; Thery, C. Rab27a
supports exosome-dependent and -independent mechanisms that modify the tumor microenvironment and
can promote tumor progression. Cancer Res. 2012, 72, 4920–4930. [CrossRef]
69. Kosaka, N.; Iguchi, H.; Hagiwara, K.; Yoshioka, Y.; Takeshita, F.; Ochiya, T. Neutral sphingomyelinase 2
(nSMase2)-dependent exosomal transfer of angiogenic microRNAs regulate cancer cell metastasis. J. Biol.
Chem. 2013, 288, 10849–10859. [CrossRef]
70. Nakamura, K.; Sawada, K.; Kinose, Y.; Yoshimura, A.; Toda, A.; Nakatsuka, E.; Hashimoto, K.; Mabuchi, S.;
Morishige, K.I.; Kurachi, H.; et al. Exosomes Promote Ovarian Cancer Cell Invasion through Transfer of
CD44 to Peritoneal Mesothelial Cells. Mol. Cancer Res. 2017, 15, 78–92. [CrossRef]
71. Singh, R.; Pochampally, R.; Watabe, K.; Lu, Z.; Mo, Y.Y. Exosome-mediated transfer of miR-10b promotes cell
invasion in breast cancer. Mol. Cancer 2014, 13, 256. [CrossRef] [PubMed]
72. Fabbri, M.; Paone, A.; Calore, F.; Galli, R.; Gaudio, E.; Santhanam, R.; Lovat, F.; Fadda, P.; Mao, C.; Nuovo, G.J.;
et al. MicroRNAs bind to Toll-like receptors to induce prometastatic inflammatory response. Proc. Natl.
Acad. Sci. USA 2012, 109, E2110–E2116. [CrossRef] [PubMed]
73. Savina, A.; Furlan, M.; Vidal, M.; Colombo, M.I. Exosome release is regulated by a calcium-dependent
mechanism in K562 cells. J. Biol. Chem. 2003, 278, 20083–20090. [CrossRef]
74. Chalmin, F.; Ladoire, S.; Mignot, G.; Vincent, J.; Bruchard, M.; Remy-Martin, J.P.; Boireau, W.; Rouleau, A.;
Simon, B.; Lanneau, D.; et al. Membrane-associated Hsp72 from tumor-derived exosomes mediates
STAT3-dependent immunosuppressive function of mouse and human myeloid-derived suppressor cells. J.
Clin. Investig. 2010, 120, 457–471. [CrossRef]
75. Nishida-Aoki, N.; Tominaga, N.; Takeshita, F.; Sonoda, H.; Yoshioka, Y.; Ochiya, T. Disruption of Circulating
Extracellular Vesicles as a Novel Therapeutic Strategy against Cancer Metastasis. Mol. Ther. 2017, 25, 181–191.
[CrossRef] [PubMed]
76. Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.;
Amigorena, S. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived
exosomes. Nat. Med. 1998, 4, 594–600. [CrossRef]
77. Romagnoli, G.G.; Zelante, B.B.; Toniolo, P.A.; Migliori, I.K.; Barbuto, J.A. Dendritic Cell-Derived Exosomes
may be a Tool for Cancer Immunotherapy by Converting Tumor Cells into Immunogenic Targets. Front.
Immunol. 2014, 5, 692. [CrossRef]
78. Admyre, C.; Johansson, S.M.; Paulie, S.; Gabrielsson, S. Direct exosome stimulation of peripheral human T
cells detected by ELISPOT. Eur. J. Immunol. 2006, 36, 1772–1781. [CrossRef]
79. Besse, B.; Charrier, M.; Lapierre, V.; Dansin, E.; Lantz, O.; Planchard, D.; Le Chevalier, T.; Livartoski, A.;
Barlesi, F.; Laplanche, A.; et al. Dendritic cell-derived exosomes as maintenance immunotherapy after first
line chemotherapy in NSCLC. Oncoimmunology 2016, 5, e1071008. [CrossRef]
80. Dai, S.; Wei, D.; Wu, Z.; Zhou, X.; Wei, X.; Huang, H.; Li, G. Phase I clinical trial of autologous ascites-derived
exosomes combined with GM-CSF for colorectal cancer. Mol. Ther. 2008, 16, 782–790. [CrossRef]
81. Morse, M.A.; Garst, J.; Osada, T.; Khan, S.; Hobeika, A.; Clay, T.M.; Valente, N.; Shreeniwas, R.; Sutton, M.A.;
Delcayre, A.; et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell
lung cancer. J. Transl. Med. 2005, 3, 9. [CrossRef] [PubMed]
82. Escudier, B.; Dorval, T.; Chaput, N.; Andre, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.;
Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC)
derived-exosomes: Results of thefirst phase I clinical trial. J. Transl. Med. 2005, 3, 10. [CrossRef] [PubMed]You can also read

















































