Physiologically relevant divalent cations modulate citrate recognition by the McpS chemoreceptor
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Research Article
Received: 18 May 2010, Revised: 6 September 2010, Accepted: 20 September 2010, Published online in Wiley Online Library: 2011
(wileyonlinelibrary.com) DOI:10.1002/jmr.1101
Physiologically relevant divalent cations
modulate citrate recognition by the McpS
chemoreceptor
Jesús Lacala, Cristina Garcı́a-Fontanaa, Carla Callejo-Garcı́aa,
Juan-Luis Ramosa and Tino Krella *
The McpS chemoreceptor of Pseudomonas putida KT2440 recognizes six different tricarboxylic acid (TCA) cycle
intermediates. However, the magnitude of the chemotactic response towards these compounds differs largely, which
has led to distinguish between strong attractants (malate, succinate, fumarate, oxaloacetate) and weak attractants
(citrate, isocitrate). Citrate is abundantly present in plant tissues and root exudates and can serve as the only carbon
source for growth. Citrate is known to form complexes with divalent cations which are also abundantly present in
natural habitats of this bacterium. We have used isothermal titration calorimetry to study the formation of
citrate–metal ion complexes. In all cases binding was entropy driven but significant differences in affinity were
observed ranging from KD ¼ 157 mM (for Mg2R) to 3 mM (for Ni2R). Complex formation occurred over a range of pH and
ionic strength. The ligand binding domain of McpS (McpS-LBD) was found to bind free citrate, but not complexes with
physiologically relevant Mg2R and Ca2R. In contrast, complexes with divalent cations which are present as trace
elements (Co2R, Cd2R and Ni2R) were recognized by McpS-LBD. This discrimination differs from other citrate sensing
proteins. These results are discussed in the context of the three dimensional structure of free citrate and its complex
with Mg2R. Chemotaxis assays using P. putida revealed that taxis towards the strong attractant malate is strongly
reduced in the presence of free citrate. However, this reduction is much less important in the presence of citrate–Mg2R
complexes. The physiological relevance of these findings is discussed. Copyright ß 2011 John Wiley & Sons, Ltd.
Keywords: chemoreceptor; chemotaxis; isothermal titration calorimetry; TCA cycle; citrate
INTRODUCTION have been described for the catabolism of TCA cycle intermedi-
ates (Bott, 1997).
Chemotaxis is a complex spatial-temporal response of microor- Different studies report that TCA cycle intermediates are
ganisms to environmental cues which modulates the activity of present at high concentrations in root exudates and dry plant
the flagellar motor enabling the bacterium to move towards or mass (Lipton et al., 1987; Kamilova et al., 2006) and soil bacteria
away from stimulatory signals. The major physiological purpose can thus use TCA cycle intermediates from these sources for
of chemotaxis consists in optimizing niche colonization through growth. Pseudomonas putida KT2440 is able to efficiently colonize
sensing of different signals, i.e. O2, pH, salt concentration or roots and seeds (Molina et al., 2000), and was found to use
availability of carbon and nitrogen sources. The chemotaxis signal organic acids and amino acids present in root exudates as the
transduction pathway has been comprehensively studied in primary carbon source during rhizosphere colonization (Vı́lchez
Escherichia coli (Bourret and Stock, 2002; Wadhams and Armitage, et al., 2000; Lugtenberg et al., 2001; Revelles et al., 2007).
2004; Hazelbauer et al., 2008) and a number of studies have A chemotactic movement towards TCA cycle intermediates has
shown that the main components of the chemotaxis signal been observed for a wide range of bacteria including P. putida
transduction pathway are relatively well conserved in the (Parales et al., 2000), P. aeruginosa (Alvarez-Ortega and Harwood,
bacterial kingdom. The key element of the chemotaxis signalling
apparatus is a ternary complex, which is composed of the
methyl-accepting chemotaxis receptor protein (MCP), the CheA
sensor kinase and the adaptor protein CheW. Signal recognition * Correspondence to: T. Krell, Estación Experimental del Zaidı́n, Consejo Superior
by the chemoreceptor modulates CheA autophosphorylation de Investigaciones Cientı́ficas, C/Profesor Albareda 1, Granada 18008, Spain.
E-mail: tino.krell@eez.csic.es
activity and in turn alters transphosphorylation of the CheY
response regulator, which acts on the flagellar motor. a J. Lacal, C. Garcı́a-Fontana, C. Callejo-Garcı́a, J.-L. Ramos, T. Krell
Many bacteria can use tricarboxylic acid (TCA) cycle Department of Environmental Protection, Estación Experimental del Zaidı́n,
intermediates as carbon and energy source. Bacteria possessing Consejo Superior de Investigaciones Cientı́ficas, C/Profesor Albareda, 1,
a complete TCA cycle require only an uptake system for the Granada 18008, Spain
utilization of these compounds (Warner and Lolkema, 2002; Abbreviations: Pb, polybuffer; LBD, ligand binding domain; TCA, tricarboxylic
Yurgel and Kahn, 2004) and in addition several anaerobic routes acid.
378
J. Mol. Recognit. 2011; 24: 378–385 Copyright ß 2011 John Wiley & Sons, Ltd.CITRATE RECOGNITION BY McpS
2007), P. fluorescens (de Weert et al., 2002), Rhizobium meliloti MATERIALS AND METHODS
(Robinson and Bauer, 1993), Campylobacter jejeuni (Hugdahl et al.,
1988) or Azospirillum spp. (Reinhold et al., 1985). Three Overexpression and purification of McpS-LBD
chemoreceptors for TCA cycle intermediates have been identified
in the past, which are Tcp of S. typhimurium (Yamamoto and Imae, Plasmid pETMcpS encodes the sequence of the McpS LBD
1993) that mediates taxis towards citrate, the malate-specific (Gly47-Ser283) fused to an N-terminal poly-histidine tag. The
receptor PA2652 of P. aeruginosa (Alvarez-Ortega and Harwood, construction of this plasmid as well as protein expression and
2007) and the McpS receptor of Pseudomonas putida KT2440 purification have been described previously (Lacal et al., 2010a).
which has recently been identified in our laboratory (Lacal et al., Briefly, E. coli BL21(DE3) containing pETMcpS was grown until the
2010a). culture reached an OD600 of 0.6 at which point IPTG was added.
The recombinant ligand binding domain (LBD) of McpS Growth was continued at 168C overnight. Cells were harvested by
contains around 250 amino acids and belongs to cluster II centrifugation and subsequently resuspended in buffer and
domains in the classification system proposed by Lacal et al. broken by French Press. Following a centrifugation step, the
(2010b). The particular interest of McpS consists in the fact that it supernatant was loaded onto a HisTrapHP column (GE
mediates chemotaxis towards more than a single TCA cycle Healthcare) and eluted with an imidazole gradient. Protein-
intermediate. In fact, McpS-LBD recognizes specifically seven containing fractions were pooled, concentrated to 5 ml, dialyzed
naturally occurring compounds which are the TCA cycle against 50 mM Tris/HCl, 0.5 M NaCl, pH 8.0 and loaded onto a
intermediates: succinate, fumarate, malate, oxaloacetate, citrate, HiPrepTM 26/60 SephacrylTM S200 gel filtration column (GE
isocitrate and in addition butyrate. However, McpS mediates a Healthcare). Protein was eluted isocratically (1 ml/min). Coomas-
strong chemotactic response only towards succinate, fumarate, sie stained SDS-PAGE gels containing 30 mg of pure McpS-LBD did
malate and oxaloacetate, which were termed strong chemot- not show any additional bands.
tractants (Lacal et al., 2010a). The remaining three compounds,
citrate, isocitrate and butyrate, cause only a marginal chemotactic Isothermal titration calorimetry
response and are referred to as weak chemoattractants. We were
Measurements were done on a VP-microcalorimeter (MicroCal,
able to demonstrate that the capacity of the strong attractants to
Amherst, MA, USA) at 208C. All measurements were done in
stabilize the dimeric form of the McpS-LBD is the molecular
Polybuffer (5 mM Tris, 5 mM MES, 5 mM PIPES, adjusted to pH
feature which determines the magnitude of the chemotactic
6.0–9.0 by the addition of concentrated HCl or NaOH). For protein
response. The binding of weak attractants to McpS-LBD did not
binding studies McpS-LBD was dialyzed overnight against
stabilize the dimeric form of this domain and consequently only a
polybuffer, pH 6. The protein sample was then adjusted to a
very modest chemotactic response is observed. Most impor-
concentration of 34–38 mM, filtered, degassed and then
tantly, weak and strong chemoattractants were found by in vitro
introduced into the instrument. Ligand solutions were made
and in vivo experimentation to compete for binding at McpS
up with dialysis buffer. For organic acid–cation binding studies,
(Lacal et al., 2010a). We were able to show that the presence of
ligand solutions were made up in the corresponding buffer,
equimolar concentrations of citrate significantly reduced the
filtered, degassed and introduced into the instrument. Titration
chemotactic response towards the strong attractants malate and
raw data were corrected for dilution heats and concentration-
succinate. Therefore, weak attractants have an antagonistic
normalized. Data analysis was carried out using the ‘One binding
action with respect to chemotaxis mediated by strong
site’ model of the MicroCal version of ORIGIN, leaving all
chemoattractants.
parameters floating.
There are several studies which report the composition of root
and seed exudates from various plants (Lipton et al., 1987; Lucas
Garcı́a et al., 2001; Kamilova et al., 2006; Liao et al., 2006). In all Optimization of cell motility
cases the antagonist citrate was present at high concentrations, Prior to chemotaxis assays the motility of P. putida KT2440 was
which is exemplified by data available on cucumber that show optimized using 1:10 diluted LB agar plates. Cells were placed
that citrate amounts to around 75% of the total organic acid into the centre of the plate and after growth for 24 h, cells present
fraction of seed and root exudates, whereas the strong on the fringe of the growth zone were again transferred to the
attractants, malate, fumarate and succinate, are present at much centre of a fresh plate. The procedure was repeated twice.
lower concentrations (Kamilova et al., 2006).
On the other hand, root exudates were also found to contain a
significant amount of divalent cations such as Ca2þ and Mg2þ Agarose plug chemotaxis assays
(Lipton et al., 1987; Qin et al., 2007). These metal ions are known Assays were carried out as described previously (Parales et al.,
to form complexes with citrate, which implies that in 2000). Bacteria were grown in M9 minimal medium supple-
physiologically relevant niches, such as the rhizosphere citrate, mented with 15 mM succinate. Plugs containing chemotaxis
is present in its free form but also as a complex with these cations. buffer (KH2PO4/K2HPO4, 0.05% (v/v) glycerol, 10 mM EDTA, pH
As stated above, citrate reduces the chemotaxis towards the 7.0) or 5 mM toluene were used as negative and positive controls,
strong attractants such as malate. The objective of this study is respectively. Cells were harvested when the OD600 was between
thus to evaluate the influence of citrate–metal complexation on 0.15 and 0.35, washed once in chemotaxis buffer and
the molecular recognition by McpS-LBD. We were able to resuspended to an OD600 of 0.7.
demonstrate that McpS-LBD binds free citrate and complexes
with cations which are present as trace elements. No interaction
Quantitative capillary assays
was observed with complexes with physiologically relevant
cations Mg2þ and Ca2þ. The physiological relevance of these Assays were carried out as described previously (Parales et al.,
findings is discussed. 2000). Cultures of P. putida KT2440 were grown in M9 minimal
379
J. Mol. Recognit. 2011; 24: 378–385 Copyright ß 2011 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmrJ. LACAL ET AL.
medium containing 15 mM succinate, harvested when the OD600 entropy changes (TDS ¼ 4.1 kcal/mol). The corresponding
was between 0.25 and 0.35, washed once in chemotaxis buffer dissociation constant was 109 mM.
and resuspended to an OD600 of 0.1. Capillaries contained 2% Subsequently, this experiment was repeated in the presence of
low-melting temperature agarose in chemotaxis buffer with or 1 mM MgCl2, which was added to both, the protein and citrate
without added attractant. solution. As shown in Figure 1B, the heat signals are very small
and almost identical to the control which consisted in a titration
of MgCl2 with the citrate–MgCl2 solution (not shown). To exclude
that the absence of binding heats is due to the compensation of
RESULTS enthalpic and entropic contributions at 208C, the experiment was
repeated at 108C, which however gave the same result. The
McpS-LBD does not recognize citrate/Mg2R complexes
ensemble of these data, thus, indicates that citrate does not bind
The Tcp chemoreceptor of Salmonella was shown to bind citrate to McpS in the presence of MgCl2.
both in its free form and in complex with magnesium ions. To verify whether the inhibitory action of MgCl2 is due to Mg2þ
Binding at Tcp occurred at the same site but in two distinct or to Cl, McpS-LBD was titrated with citrate in the presence of
manners involving different sets of amino acids (Iwama et al., NaCl instead of MgCl2 and binding was observed similar to that in
2006). Root exudates contain a significant amount of bivalent the absence of MgCl2. To determine whether the inhibitory action
cations, such as Ca2þ and Mg2þ (Lipton et al., 1987; Qin et al., of Mg2þ ions is due to its binding to the protein or citrate,
2007), which form a complex with citrate. The initial set of McpS-LBD and citrate were titrated with MgCl2. No interaction
experiments was aimed at evaluating the influence of Mg2þ on between McpS-LBD and MgCl2 was observed (not shown)
the molecular recognition of citrate by the LBD of McpS, whereas Mg2þ was found to bind to citrate (Figure 2A). Binding
McpS-LBD. To this end, we have used isothermal titration was driven by favourable entropy changes and counterbalanced
calorimetry (Krell, 2008) which can be used to derive the by unfavourable enthalpy changes (Table 1).
complete set of thermodynamic binding parameters (dis-
sociation constant, changes in enthalpy, entropy and free energy)
Citrate–Mg2R interaction occurs over a range of pH and
in a single experiment. Figure 1A shows the microcalorimetric
ionic strength
titration of McpS-LBD with free citrate. Data analysis reveals that
citrate binding is driven by favourable enthalpy changes A literature research revealed that there are a large number of
(DH ¼ 9.4 kcal/mol) which is counterbalanced by unfavourable studies of the interaction of citrate/ion complexes with proteins
but information on the actual citrate–metal complex formation is
Figure 1. Isothermal titration of the recombinant ligand binding
domain of McpS (McpS-LBD) in the absence and presence of MgCl2.
(A) Injection of 1.6 ml and a series of 12.8 ml aliquots of 1 mM citrate into 36
mM of McpS. (B) Injection of 1.6 ml and a series of 12.8 ml aliquots of 1 mM Figure 2. ITC studies of the binding of divalent cations to citrate. Shown
citrate containing 1 mM MgCl2 into 36 mM of McpS containing 1 mM are in the upper panel titrations of 90 mM citrate with 3 mM solutions of
MgCl2. Ligands were in polybuffer (5 mM Tris, 5 mM MES, 5 mM PIPES), pH MgCl2 (A) NiCl2 (B) and CoCl2 (C). The injection volume was in all cases
6.0. Experiments were carried out at 208C. Upper panel: Raw titration data. 6.4 ml. Lower panel: Integrated, dilution-corrected and concentration-
Lower panel: Integrated and dilution corrected peak areas of raw data. normalized peak areas of titration raw data. Shown are the fits with
Data were fitted using the ‘One binding site model’ of the MicroCal the ‘One binding site model’ of the MicroCal version of ORIGIN: MgCl2 (D),
version of ORIGIN. NiCl2 (*), CoCl2 ( ).
380
wileyonlinelibrary.com/journal/jmr Copyright ß 2011 John Wiley & Sons, Ltd. J. Mol. Recognit. 2011; 24: 378–385CITRATE RECOGNITION BY McpS
Table 1. Binding parameters derived from isothermal titration calorimetry experiments. All experiments were conducted at 208C.
Ligand 1 was placed into the sample cell and ligand 2 was present in the syringe. The composition of polybuffer is 5 mM Tris, 5 mM
MES, 5 mM PIPES adjusted to the pH indicated by the addition of concentrated HCl or NaOH
Ligand 1 Ligand 2 Buffer KD (mM) DH (kcal/mol) TDS (kcal/mol)
McpS-LBD Citrate Polybuffer (Pb) pH 6.0 109 7 9.4 0.3 4.01 0.3
McpS-LBD Citrate Pb, pH 6.0, þ 1 mM MgCl2 No binding
McpS-LBD Citrate Pb, pH 6.0, þ 1 mM CaCl2 No binding
McpS-LBD Citrate Pb, pH 6.0, þ 1 mM CdCl2 251 24 6.6 0.4 1.73 0.4
McpS-LBD Citrate Pb, pH 6.0, þ 1 mM CoCl2 330 50 6.12 0.9 1.44 0.9
McpS-LBD Citrate Pb, pH 6.0, þ 1 mM NiCl2 168 12 1.02 0.1 3.95 0.1
McpS-LBD MgCl2 Pb, pH 6.0 No binding
Citrate MgCl2 Pb, pH 6.0 157 3 1.38 0.01 6.35 0.05
Citrate MgCl2 Pb, pH 7.0 93 4 2.05 0.06 7.32 0.05
Citrate MgCl2 Pb, pH 8.0 85 5 2.05 0.08 7.38 0.09
Citrate MgCl2 Pb, pH 9.0 64 3 1.53 0.04 7.03 0.06
Citrate MgCl2 Pb, pH 8.0, 30 mM NaCl 218 14 2.56 0.4 7.35 0.38
Citrate MgCl2 Pb, pH 8.0, 60 mM NaCl 250 10 2.61 0.2 7.32 0.2
Citrate MgCl2 Pb, pH 8.0, 100 mM NaCl 497 32 2.53 0.3 6.85 0.3
Citrate MgCl2 Pb, pH 8.0, 130 mM NaCl 299 18 1.76 0.6 6.36 0.9
Citrate CaCl2 Pb, pH 6.0 66 13 0.22 0.04 5.71 0.1
Citrate CdCl2 Pb, pH 6.0 43 1 2.34 0.03 8.05 0.3
Citrate CoCl2 Pb, pH 6.0 8.54 0.3 2.04 0.01 8.70 0.3
Citrate NiCl2 Pb, pH 6.0 3.18 0.1 1.73 0.01 8.94 0.3
isocitrate MgCl2 Pb, pH 8.0 389 60 1.85 0.7 6.30 0.7
tricarballylate MgCl2 Pb, pH 8.0 No binding
very scarce. Therefore, a series of experiments were designed to citrate and divalent cations, we studied the binding character-
cast light into this issue. Citrate recognition by McpS occurs in the istics of physiologically relevant cations, Ca2þ and Mg2þ, and of
periplasm. It was shown that the periplasmic pH varies in function Co2þ, Cd2þ and Ni2þ, which are present in natural habitats as
of the pH in the extracellular space (Wilks and Slonczewski, 2007). trace elements. This series of experiments was conducted at pH
The above titration of citrate with MgCl2 was conducted at pH 6.0 6.0, which is likely to correspond to periplasmic pH when the
and additional experiments were carried out to determine the pH bacterium is in a neutral medium.
and salt dependence of this interaction. As shown in Table 1, a In addition to the citrate–Mg2þ binding studies reported
citrate–Mg2þ interaction is seen over a pH range of 6.0–9.0 and an above, microcalorimetric titrations of citrate with Ca2þ, Co2þ,
increase in pH causes a slight increase in the binding affinity Cd2þ and Ni2þ were conducted (Figure 2, Table 1). Interestingly,
(Table 1). The buffer system so far used did not contain any all four cations bound to citrate in an entropy driven manner and
additionally added salts and subsequently the influence of NaCl an apparent stoichiometry close to 1:1 (Table 1). In all cases
on the citrate–Mg2þ interaction was determined at pH 8.0 complex formation was tighter than observed for Mg2þ. The
(Table 1). The presence of NaCl was found to only slightly reduce highest affinity was observed for complexes with Co2þ and Niþ2
the citrate–Mg2þ affinity. Data suggest that citrate–Mg2þ characterized by a KD of 8 and 3 mM, respectively. These data
complex formation occurs over a wide range of conditions suggest that complexation of citrate is a feature common to
underlining its physiological relevance. many divalent cations and that binding affinity increases with the
size of the cation.
Many divalent cations bind specifically citrate
McpS binds citrate in complex with Co2R, Cd2R and Ni2R but
The next series of experiments consisted in an evaluation of the
does not bind complexes with Mg2R and Ca2R
interaction of compounds which are similar in their properties to
Mg2þ and citrate. Initial experiments involved binding studies of To determine whether McpS-LBD recognizes all these five citrate
isocitrate and tricarballylate with Mg2þ. Isocitrate, which is also a complexes, the protein in the presence of the corresponding
weak chemoattractant of McpS (Lacal et al., 2010a), bound Mg2þ cations was titrated with the citrate–cation complexes studied
with a significantly reduced affinity as compared to citrate above. Interestingly, McpS-LBD was found to bind citrate in
(Table 1). Tricarballylate, a non-naturally occurring citrate complex with the trace elements Co2þ, Cd2þ and Ni2þ (Figure 3,
derivative, was devoid of binding. These date indicate a certain Table 1). The affinity of McpS for these complexes was reduced by
specificity for the Mg2þ binding to citrate. a factor of 1.5–3 as compared to free citrate. However, in analogy
As stated above, Ca2þ and Mg2þ were found to be the to the experiment with Mg2þ (Figure 1), citrate in complex with
prevalent divalent cations in root exudates (Lipton et al., 1987; Ca2þ is not recognized by McpS (Figure 3). In summary, studies
Qin et al., 2007). To investigate the complex formation between show that citrate in complex with trace elements Co2þ, Cd2þ and
381
J. Mol. Recognit. 2011; 24: 378–385 Copyright ß 2011 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmrJ. LACAL ET AL.
as malate (Lacal et al., 2010a). This inhibitory action is caused by
the fact that citrate per se causes a very weak response and that
citrate and malate compete for the same binding site at McpS
(Lacal et al., 2010a).
We have now conducted agarose plug and quantitative
capillary chemotaxis assays of P. putida KT2440 towards the
strong attractant malate in the absence or presence of either
citrate or citrate Mg2þ/complexes. Amongst the seven ligands of
McpS, malate was chosen for this series of experiments since it
caused the strongest chemotactic response in vivo and binds with
the highest affinity to McpS-LBD in vitro. The agarose plug assay
involves placing a solidified agarose plug containing chemoat-
tractant in contact with a cell suspension. The formation of rings
at a certain distance to the plug is indicative of a chemotactic
response. This type of assays provides rather a qualitative than
quantitative type of information.
When bacteria are brought in contact with malate-containing
plugs, the formation of the typical ring takes place (shown by
þþþ in Table 2). When this experiment is repeated in the
presence of citrate, which was added to both the agarose plug
and the cell suspension, only weak taxis is observed confirming
the results obtained by Lacal et al. (2010a) that showed an
inhibition of taxis towards malate in the presence of citrate
Figure 3. Microcalorimetric titration of McpS-LBD with citrate com- (Table 2). However, when this experiment is repeated in the
plexed with different divalent cations. Titration of 35 mM McpS-LBD presence of citrate/Mg2þ instead of free citrate, a certain recovery
containing 6 mM of the corresponding cationCl2 salt with 3 mM citrate of taxis towards malate is observed (Table 2). In addition, a series
in the presence of 6 mM cationCl2 salt. The injection volume was 12 ml in of control experiments were conducted which show that MgCl2
all cases. (A) CaCl2, (B) NiCl2, (C) CoCl2. Lower panel: Integrated, dilution- on its own does not interfere with taxis towards malate and that
corrected and concentration-normalized peak areas of titration raw data. citrate and MgCl2 do not appear to have any non-specific effect
Shown are the fits with the ‘One binding site model’ of the MicroCal
on bacterial motility as witnessed by the observation that the
version of ORIGIN: NiCl2 (*), CoCl2 ( ).
presence of both compounds do not interfere with taxis towards
toluene which is mediated by a different chemoreceptor (Table 2).
Ni2þ is recognized by McpS whereas citrate complexes with Mg2þ To confirm these observations, capillary assays were conducted
and Ca2þ, which are abundantly present in nature, do not bind to which provide more quantitative information. In contrast to the
this protein. plug assays the agarose containing the attractant is placed into a
capillary which is brought in contact with a bacterial suspension
and cells which migrate into this capillary are subsequently
Differential inhibition of chemotaxis towards malate by
quantified. As shown in Table 2, the capillary assays confirm the
citrate and citrate/Mg2R complexes
observation made by plug assays. In the presence of citrate, a
We have shown previously that the presence of citrate reduces significant drop in taxis towards malate is observed. This inhibition
the McpS-mediated chemotaxis towards strong attractants such is relieved by the addition of MgCl2. These data show that a
Table 2. Chemotactic responses of Pseudomonas putida KT2440 in agarose plug and modified capillary assays. Agarose plug and
quantitative capillary assays were carried out as described in Materials and Methods. Results given are the means of at least three
individual experiments
Chemotactic response
Compounds present in Agarose Capillary assay
Compounds immobilized in agarose bacterial suspension plug (cells in capillary)
Buffer Buffer — 200 40
Malate (0.7 mM) Buffer þþþ 2100 400
Malate (0.7 mM), citrate (1 mM) Citrate (1 mM) þ 400 50
Malate (0.7 mM), citrate (1 mM), MgCl2 (2 mM) Citrate (1 mM), MgCl2 (2 mM) þþ 2000 350
Malate (0.7 mM), MgCl2 (2 mM) MgCl2 (2 mM) þþþ 2100 350
Toluene (1 mM) Buffer þþ 900 150
Toluene (1 mM), citrate (1 mM) Citrate (1 mM) þþ 1000 150
Toluene (1 mM), MgCl2 (2 mM) MgCl2 (2 mM) þþ 950 300
þþþ, strong taxis; þþ, intermediate taxis; þweak taxis
382
wileyonlinelibrary.com/journal/jmr Copyright ß 2011 John Wiley & Sons, Ltd. J. Mol. Recognit. 2011; 24: 378–385CITRATE RECOGNITION BY McpS
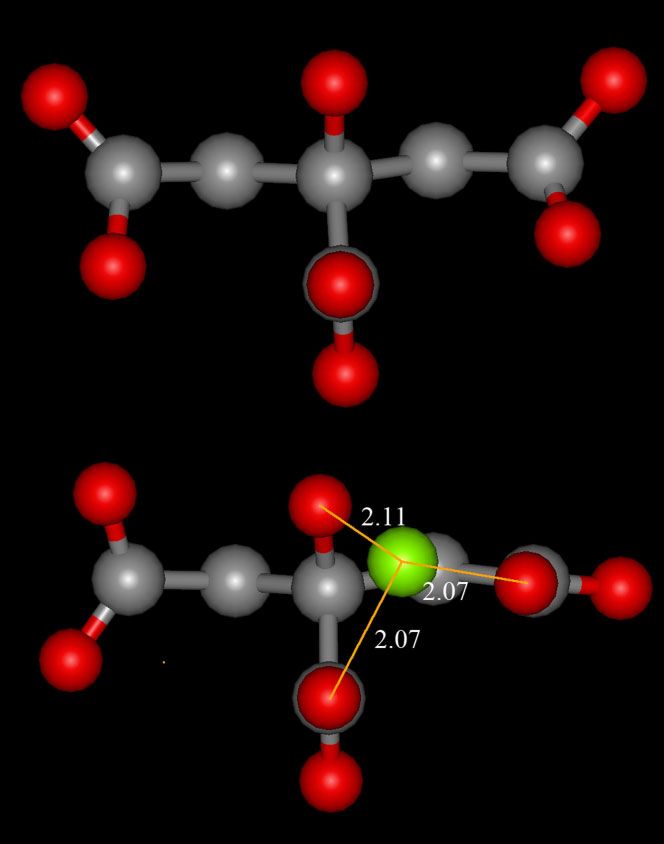
modulation of taxis towards the strong attractant is modulated by symmetrical structure. The planes formed by the carboxyl groups
Mg2þ. This modulation is mediated by the differential recognition at C1 and C5 are almost perpendicular to the C–OH bond. The
of citrate and citrate/Mg2þ complexes by the chemoreceptor. carboxyl groups are only weakly restricted in their rotational
freedom. Mg2þ establishes three interactions with citrate: two
interactions with the carboxyl groups at C5 and C6 and a third
DISCUSSION interaction with the hydroxyl group. The bond distances between
Mg2þ and these three sites are almost identical (Figure 4). The
Pseudomonas putida KT2440, the organism under investigation, is most important change in the Mg2þ bound citrate structure is a
a saprophyte and is also able to efficiently colonize roots and rotation of the C5 carboxyl group which is almost perpendicular
seeds (Molina et al., 2000). This strain uses organic acids present to the carboxyl group at C1. As a consequence of this interaction,
in root exudates as the primary carbon source during rhizosphere the carboxyl group at C5 is largely restricted in its rotational
colonization (Lugtenberg et al., 2001) and shows chemotaxis flexibility. This pronounced rotation of the C5 carboxyl group
toward six different TCA cycle intermediates. Amongst the 26 might be the reason for the lacking binding of citrate/Mg2þ and
chemoreceptors of this strain, McpS was found to be the only citrate/Ca2þ complexes to McpS-LBD. It might be hypothesized
receptor for TCA cycle intermediates (Lacal et al., 2010a). Citrate is that the binding of larger cations causes a less important rotation
one of the most abundant organic acids in the natural habitat of P. of the C5 carboxyl group, which might account for the fact that
putida, i.e. plant surfaces and root exudates, where it is present at protein binding occurs, although with weaker affinity as
lower millimolar concentrations reaching in some cases 10 mM compared to free citrate. A verification of this hypothesis will
(Lipton et al., 1987; Johnson et al., 1994; Lucas Garcı́a et al., 2001; need to await the resolution of crystal structures of McpS with a
Kamilova et al., 2006). McpS-LBD was found to bind citrate with an variety of ligands which is an ongoing research activity in our
affinity of 109 mM. With respect to the high citrate concentration laboratory.
in the natural environment of this bacterium, this affinity is, Citrate present in the bacterial habitats is of significant
therefore, of physiological relevance. physiological relevance, which is underlined by the fact that several
A key finding of this work consists in the demonstration that protein families have evolved which are able to recognize citrate.
citrate complexation with Mg2þ abolished recognition by McpS. Bivalent cations as well as citrate were found to be present in root
The three-dimensional structures of free citrate and citrate in exudates in millimolar concentrations (Lipton et al., 1987; Lucas
complex with Mg2þ have been solved (Johnson, 1965). As shown Garcı́a et al., 2001 Kamilova et al., 2006; Qin et al., 2007). Isothermal
in Figure 4, the unliganded form of citrate is an almost Titration Calorimetry (ITC) data show that citrate binds bivalent
cations with affinities ranging between 3 and 500 mM, which implies
that under physiological conditions the majority of citrate is
complexed whereas a fraction will remain ligand-free. Amongst the
different citrate recognizing proteins, there is no clear consensus as
to the specificity of the recognition of free and metal bound citrate
and the identification of physiological reasons for these differences
in binding specificity is not a straightforward issue.
In general, there are three major families of proteins which
recognize extracytosolic citrate, which are citrate transporters,
sensor kinases of two component systems and chemoreceptors.
Information on the specificity of citrate binding for members of all
of these families is available. There are, for example, transporters
which transport exclusively uncomplexed citrate of which the
CitS transporter of Klebsiella pneumoniae is an example. This
protein was shown to exclusively recognize Hcitrate2 (Kästner et
al., 2003). Other transporters recognize citrate metal complexes
with a defined metal specificity. Within the latter category,
significant differences exist in the specificity for the type of
metal–citrate complex. For example, the CitH transporter of B.
subtilis transports citrate in complex with Ca2þ, Sr2þ, Mu2þ, Cd2þ
and Pb2þ, whereas citrate in complex with Mg2þ, Zn2þ, Ni2þ and
Co2þ are not transported (Krom et al., 2000). Interestingly, the
CitM transporter of the same organism acts specifically on the
metal–citrate complexes which are not transported by CitH
(Boorsma et al., 1996; Krom et al., 2000). A similar situation is also
found in other species such as Enterococcus faecalis (Blancato et
al., 2006). The ability of a microorganism to transport different
citrate complexes would allow the colonization of different
niches where the availability of an abundant carbon and energy
Figure 4. Three dimensional structure of free citrate and in complex depends on the presence of ions. Therefore, the capacity to use
with Mg2þ. Structures were retrieved from the Cambridge Structural transport system with overlapping substrate specificity can
database. Upper part: structure of free citrate (refcolde: CitraC); lower confer colonization advantages to microbes.
part: structure of citrate in complex with Mg2þ, shown in green (refcode The second class of citrate sensing proteins are sensor kinases
MgcitD) (Johnson, 1965). of two component regulator systems (TCS) in facultative
383
J. Mol. Recognit. 2011; 24: 378–385 Copyright ß 2011 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmrJ. LACAL ET AL.
anaerobic bacteria such as E. coli and Klebsiella. These TCS intermediates. However, the magnitude of chemotaxis indicates
regulate the expression of genes involved in citrate transport and that taxis to only malate, fumarate, succinate and oxaloacetate
metabolism. Examples are the highly citrate specific TCS CitA/CitB are of physiological relevance. Citrate was found to be present in
of Klebsiella pneumoniae (Kaspar et al., 1999) and the DcuS/DcuR root and seed exudates at very high concentrations and the
system of E. coli (Krämer et al., 2007) which recognizes several physiological relevance of citrate binding to McpS, resulting in a
C4-dicarboxylates and citrate. CitA/CitB regulates amongst others very significant reduction in the chemotaxis towards the four
the expression of the above mentioned CitS transporter. Citrate strong attractants, was unclear (Lacal et al., 2010a). Analysis of
binding by CitA is accomplished by a PAS domain, which is root exudates suggests that citrate is primarily present in
located in the periplasm (Reinelt et al., 2003). CitA was found to complex with Mg2þ and Ca2þ (Lipton et al., 1987; Lucas Garcı́a
bind free citrate and with some reduced affinity also the et al., 2001; Kamilova et al., 2006; Qin et al., 2007). This work shows
citrate–Mg2þ complex. that McpS does not recognize both complexes and that citrate
The third class of extracytoplasmic citrate binding proteins are complexed to Mg2þ does not inhibit taxis towards the strong
chemoreceptors of which Tcp is the most studied example attractant malate. This work thus permits a reassessment of the
(Yamamoto and Imae, 1993; Iwama et al., 2000; Iwama et al., inhibitory action of citrate on the taxis towards strong attractants.
2006). This receptor employs a TarH type of domain for citrate On the other hand, it remains to be established why the organism
sensing (Interpro signature IPR003122) which forms a 4-helix under study has not evolved a chemotactic mechanism which
bundle structure (Yeh et al., 1996). Iwama et al. (2006) showed permits an efficient taxis towards citrate which is abundantly
that this receptor binds citrate both in its free form and in present in natural habitats of this bacterium.
complex with magnesium ions. The authors demonstrate that
free and Mg2þ bound citrate bind to the same site at Tcp but in
two distinct manners involving different sets of amino acids.
Acknowledgements
Our data reveal yet another mode of citrate recognition:
Binding occurs for free citrate as well as in complex with larger The authors acknowledge financial support from the BBVA
cations, whereas complexes with the small cations, Mg2þ and foundation and the Andalusian Regional Government Junta de
Ca2þ, are not recognized. This is in strong contrast to the other Andalucı́a (EDFR grant P09-RNM-4509 to TK and EDFR Bio-191 to
citrate chemoreceptor Tcp. McpS is different to most studied the group ‘Degradation of toxic organic compounds’). They wish
chemoreceptors since it has a relatively broad ligand spectrum to thank Dr Jose A. Gavira for his help in retrieving the structures
and recognizes with high specificity six different TCA cycle of citrate.
REFERENCES
Alvarez-Ortega C, Harwood CS. 2007. Identification of a malate chemor- Johnson JF, Allan DL, Vance CP. 1994. Phosphorus stress-induced proteoid
eceptor in Pseudomonas aeruginosa by screening for chemotaxis roots show altered metabolism in Lupinus albus. Plant Physiol. 85:
defects in an energy taxis-deficient mutant. Appl. Environ. Microbiol. 315–317.
73: 7793–7795. Kamilova F, Kravchenko LV, Shaposhnikov AI, Azarova T, Makarova N,
Blancato VS, Magni C, Lolkema JS. 2006. Functional characterization and Lugtenberg B. 2006. Organic acids, sugars, and L-tryptophane in
Me ion specificity of a Ca-citrate transporter from Enterococcus exudates of vegetables growing on stonewool and their effects on
faecalis. FEBS J. 273: 5121–5130. activities of rhizosphere bacteria. Mol. Plant Microbe Interact. 19:
Boorsma A, van der Rest ME, Lolkema JS, Konings WN. 1996. Secondary 250–256.
transporters for citrate and the Mg2þ-citrate complex in Bacillus Kaspar S, Perozzo R, Reinelt S, Meyer M, Pfister K, Scapozza L, Bott M.
subtilis are homologous proteins. J. Bacteriol. 178: 6216–6222. 1999. The periplasmic domain of the histidine autokinase CitA
Bott M. 1997. Anaerobic citrate metabolism and its regulation in entero- functions as a highly specific citrate receptor. Mol. Microbiol. 33:
bacteria. Arch. Microbiol. 167: 78–88. 858–872.
Bourret RB, Stock AM. 2002. Molecular information processing: lessons Kästner CN, Prummer M, Sick B, Renn A, Wild UP, Dimroth P. 2003. The
from bacterial chemotaxis. J. Biol. Chem. 277: 9625–9628. citrate carrier CitS probed by single-molecule fluorescence spec-
de Weert S, Vermeiren H, Mulders IH, Kuiper I, Hendrickx N, Bloemberg GV, troscopy. Biophys. J. 84: 1651–1659.
Vanderleyden J, De Mot R, Lugtenberg BJ. 2002. Flagella-driven Krämer J, Fischer JD, Zientz E, Vijayan V, Griesinger C, Lupas A, Unden G.
chemotaxis towards exudate components is an important trait for 2007. Citrate sensing by the C4-dicarboxylate/citrate sensor kinase
tomato root colonization by Pseudomonas fluorescens. Mol. Plant DcuS of Escherichia coli: binding site and conversion of DcuS to
Microbe Interact. 15: 1173–1180. a C4-dicarboxylate- or citrate-specific sensor. J. Bacteriol. 189:
Hazelbauer GL, Falke JJ, Parkinson JS. 2008. Bacterial chemoreceptors: 4290–4298.
high-performance signaling in networked arrays. Trends Biochem. Sci. Krell T. 2008. Microcalorimetry: a response to challenges in modern
33: 9–19. biotechnology. Microb. Biotechnol. 1: 126–136.
Hugdahl MB, Beery JT, Doyle MP., 1988. Chemotactic behavior of Campy- Krom BP, Warner JB, Konings WN, Lolkema JS. 2000. Complementary metal
lobacter jejuni. Infect. Immun. 56: 1560–1566. ion specificity of the metal-citrate transporters CitM and CitH of
Iwama T, Ito Y, Aoki H, Sakamoto H, Yamagata S, Kawai K, Kawagishi I. 2006. Bacillus subtilis. J. Bacteriol. 182: 6374–6381.
Differential recognition of citrate and a metal-citrate complex by the Lacal J, Alfonso C, Liu X, Parales RE, Morel B, Conejero-Lara F, Rivas G,
bacterial chemoreceptor Tcp. J. Biol. Chem. 281: 17727–17735. Duque E, Ramos JL, Krell T. 2010a. Identification of a chemoreceptor
Iwama T, Nakao KI, Nakazato H, Yamagata S, Homma M, Kawagishi I. 2000. for TCA cycle intermediates: ligand induced dimerization of chemo-
Mutational analysis of ligand recognition by tcp, the citrate chemo- receptor sensor domain determines magnitude of chemotaxis. J. Biol.
receptor of Salmonella enterica serovar typhimurium. J. Bacteriol. 182: Chem. 285: 23126–23136.
1437–1441. Lacal J, Garcı́a-Fontana C, Muñoz-Martı́nez F, Ramos JL, Krell T. 2010b.
Johnson CK. 1965. X-ay crystal analysis of the substrates of aconitase. v. Sensing of environmental signals: classification of chemoreceptors
magnesium citrate decahydrate (Mg(H2O)6)(MgC6H5O7(H2O))2. 2H2O. according to the size of their ligand binding regions. Environ. Micro-
Acta Crystallogr. 18: 1004–1018. biol. 182: 2873–2884. DOI: 10.1111/j.1462-2920.2010.02325
384
wileyonlinelibrary.com/journal/jmr Copyright ß 2011 John Wiley & Sons, Ltd. J. Mol. Recognit. 2011; 24: 378–385CITRATE RECOGNITION BY McpS
Liao H, Wan H, Shaff J, Wang X, Yan X, Kochian LV. 2006. Phosphorus and Reinhold B, Hurek T, Fendrik I. 1985. Strain-specific chemotaxis of Azos-
aluminum interactions in soybean in relation to aluminum tolerance. pirillum spp. J. Bacteriol. 162: 190–195.
Exudation of specific organic acids from different regions of the intact Revelles O, Wittich RM, Ramos JL. 2007. Identification of the initial steps in
root system. Plant Physiol. 141: 674–684. D-lysine catabolism in Pseudomonas putida. J Bacteriol. 189:
Lipton DS, Blanchar RW, Blevins DG. 1987. Citrate, malate, and succinate 2787–2792.
concentration in exudates from P-Sufficient and P-Stressed Medicago Robinson JB, Bauer WD. 1993. Relationships between C4 dicarboxylic acid
sativa L. seedlings. Plant Physiol. 85: 315–317. transport and chemotaxis in Rhizobium meliloti. J. Bacteriol. 175:
Lucas Garcı́a JA, Barbas C, Probanza A, Barrientos ML, Gutierrez Mañero FJ. 2284–2291.
2001. Low molecular weight organic acids and fatty acids in root Vı́lchez S, Molina L, Ramos C, Ramos JL. 2000. Proline catabolism by
exudates of two Lupinus cultivars at flowering and fruiting stages. Pseudomonas putida: cloning, characterization, and expression of the
Phytochem. Anal. 12: 305–311. put genes in the presence of root exudates. J. Bacteriol. 182: 91–99.
Lugtenberg BJ, Dekkers L, Bloemberg GV. 2001. Molecular determinants of Wadhams GH, Armitage JP. 2004. Making sense of it all: bacterial che-
rhizosphere colonization by Pseudomonas. Annu. Rev. Phytopathol. motaxis. Nat. Rev. Mol. Cell. Biol. 5: 1024–1037.
39: 461–490. Warner JB, Lolkema JS. 2002. Growth of Bacillus subtilis on citrate and
Molina L, Ramos C, Duque E, Ronchel MC, Garcı́a JM, Wyke L, Ramos JL. isocitrate is supported by the Mg2þ-citrate transporter CitM. Micro-
2000. Survival of Pseudomonas putida KT2440 in soil and in the biology 148: 3405–3412.
rhizosphere of plants under greenhouse and environmental con- Wilks JC, Slonczewski JL. 2007. pH of the cytoplasm and periplasm of
ditions. Soil Biol. Biochem. 32: 315–321. Escherichia coli: rapid measurement by green fluorescent protein
Parales RE, Ditty JL, Harwood CS. 2000. Toluene-degrading bacteria are fluorimetry. J. Bacteriol. 189: 5601–5607.
chemotactic towards the environmental pollutants benzene, toluene, Yamamoto K, Imae Y. 1993. Cloning and characterization of the Salmonella
and trichloroethylene. Appl. Environ. Microbiol. 66: 4098–4104. typhimurium-specific chemoreceptor Tcp for taxis to citrate and from
Qin R, Hirano Y, Brunner I. 2007. Exudation of organic acid anions from phenol. Proc. Natl. Acad. Sci. USA 90: 217–221.
poplar roots after exposure to Al, Cu and Zn. Tree Physiol. 27: 313–320. Yeh JI, Biemann HP, Privé GG, Pandit J, Koshland DE Jr. 1996. High-
Reinelt S, Hofmann E, Gerharz T, Bott M, Madden DR. 2003. The structure of resolution structures of the ligand binding domain of the wild-type
the periplasmic ligand-binding domain of the sensor kinase CitA bacterial aspartate receptor. J. Mol. Biol. 262: 186–201.
reveals the first extracellular PAS domain. J. Biol. Chem. 278: Yurgel SN, Kahn ML. 2004. Dicarboxylate transport by rhizobia. FEMS
39189–39196. Microbiol Rev. 28: 489–501.
385
J. Mol. Recognit. 2011; 24: 378–385 Copyright ß 2011 John Wiley & Sons, Ltd. wileyonlinelibrary.com/journal/jmrYou can also read

















































