Endogenous oncogenic Nras mutation initiates hematopoietic malignancies in a dose- and cell type-dependent manner
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below

LYMPHOID NEOPLASIA
Endogenous oncogenic Nras mutation initiates hematopoietic malignancies in a
dose- and cell type-dependent manner
Jinyong Wang,1 Yangang Liu,1 Zeyang Li,2 Zhongde Wang,3 Li Xuan Tan,4 Myung-Jeom Ryu,1 Benjamin Meline,5 Juan Du,1
Ken H. Young,6 Erik Ranheim,6 Qiang Chang,7 and Jing Zhang1
1McArdle Laboratory for Cancer Research, University of Wisconsin-Madison, Madison, WI; 2Department of Biochemistry, University of Wisconsin-Madison,
Madison, WI; 3Hematech Inc, Sioux Falls, SD; 4Department of Genetics, University of Wisconsin-Madison, Madison, WI; 5Department of Molecular and Cellular
Pharmacology, University of Wisconsin-Madison, Madison, WI; 6Department of Pathology & Laboratory Medicine, University of Wisconsin School of Medicine
and Public Health, University of Wisconsin Carbone Cancer Center, Madison, WI; and 7Waisman Center, University of Wisconsin-Madison, Madison, WI
Both monoallelic and biallelic oncogenic matic expression of Nras G12D/G12D but transplanted with Nras G12D/G12D bone
NRAS mutations are identified in human not Nras G12D/ⴙ leads to hyperactivation marrow cells develop TALL (TALL-homo).
leukemias, suggesting a dose-dependent of ERK, excessive proliferation of my- Although both TALL-het and -homo tu-
role of oncogenic NRAS in leukemogen- eloid progenitors, and consequently an mors acquire Notch1 mutations and are
esis. Here, we use a hypomorphic onco- acute myeloproliferative disease. Using a sensitive to a ␥-secretase inhibitor, endog-
genic Nras allele and a normal oncogenic bone marrow transplant model, we previ- enous Nras G12D/ⴙ signaling promotes
Nras allele (Nras G12Dhypo and Nras G12D, ously showed that ⬃ 95% of animals re- TALL through distinct genetic mecha-
respectively) to create a gene dose gradi- ceiving Nras G12D/ⴙ bone marrow cells nism(s) from Nras G12D/G12D. Our data
ent ranging from 25% to 200% of endoge- develop chronic myelomonocytic leuke- indicate that the tumor transformation
nous Nras G12D/ⴙ. Mice expressing Nras mia (CMML), while ⬃ 8% of recipients potential of endogenous oncogenic Nras
G12Dhypo/G12Dhypo develop normally and develop acute T-cell lymphoblastic leuke- is both dose- and cell type-dependent.
are tumor-free, whereas early embryonic mia/lymphoma [TALL] (TALL-het). Here (Blood. 2011;118(2):368-379)
expression of Nras G12D/ⴙ is lethal. So- we demonstrate that 100% of recipients
Introduction
In mammals 3 different ras gene loci encode 4 highly homologous to stronger phenotypes and significantly shorter disease latency.16,17
21-KD proteins: Hras, Nras, Kras.4A and Kras.4B.1 Ras proteins However, whether or not this conclusion would be true with
belong to the super family of small GTPases. They cycle between endogenous transcriptional regulation of the Nras locus was, until
the active GTP-bound form and the inactive GDP-bound form.2,3 now, uncertain.
Once Ras proteins are activated, they subsequently activate mul- Haigis et al generated a conditional oncogenic Nras allele,18 in
tiple downstream signaling pathways (including the Raf/MEK/ which the oncogenic mutation G12D was introduced into the
ERK and PI3K/Akt pathways) and regulate cell survival, prolifera- endogenous Nras locus and expression of oncogenic Nras is
tion, and differentiation. blocked by a floxed STOP cassette (LSL Nras G12D). In the
Oncogenic mutations in the 3 RAS genes have been identified presence of Cre recombinase, the STOP cassette is removed and
in virtually all types of human cancers, with characteristic oncogenic Nras is expressed at a level comparable with the
incidences and RAS gene associations.4 In particular, mutations wild-type (WT) Nras allele.19 Using this allele, we established a
in the KRAS and NRAS genes (but rarely in the HRAS gene) are mouse bone marrow transplantation model in which monoallelic
frequently identified in myeloid disorders (15%-60%), including oncogenic Nras is expressed in bone marrow cells alone.19 Approxi-
acute myeloid leukemia (AML),5,6 atypical chronic myeloid mately 95% of recipient mice developed a CMML-like disease
leukemia,7 chronic myelomonocytic leukemia (CMML),8-10 and after a prolonged latency. Moreover, CMML development is
juvenile myelomonocytic leukemia (JMML).8-10 In contrast, associated with aberrant GM-CSF signaling and UPD of the
although hyperactivated Ras signaling is identified in 50% of oncogenic Nras allele, both of which are reported in human CMML
patients with acute T-cell lymphoblastic leukemia/lymphoma specimens.14,20 These results highlight the relevance of our model
(TALL), oncogenic NRAS mutations only occur in ⬍ 5% of to the human disease.
these patients.4,11-13 The utility of genetically engineered alleles to study gene
Uniparental disomy (UPD) of an oncogenic RAS allele is dosages has been reported extensively in mouse.21 Here, we
reported in both primary human tumor samples and tumor cell report that using a sophisticated mouse genetic approach,
lines,14,15 suggesting a dose-dependent role of oncogenic RAS in we created a gradient of gain-of Nras signaling ranging from
tumor development. Results obtained from several mouse models 25%-200% of endogenous monoallelic expression of oncogenic
under conditions resulting in overexpression of oncogenic Nras Nras. We studied the effects of this signaling gradient on
support this hypothesis; increasing levels of oncogenic Nras leads leukemogenesis.
Submitted December 17, 2010; accepted April 28, 2011. Prepublished online payment. Therefore, and solely to indicate this fact, this article is hereby
as Blood First Edition paper, May 17, 2011; DOI 10.1182/blood-2010-12-326058. marked ‘‘advertisement’’ in accordance with 18 USC section 1734.
The publication costs of this article were defrayed in part by page charge © 2011 by The American Society of Hematology
368 BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2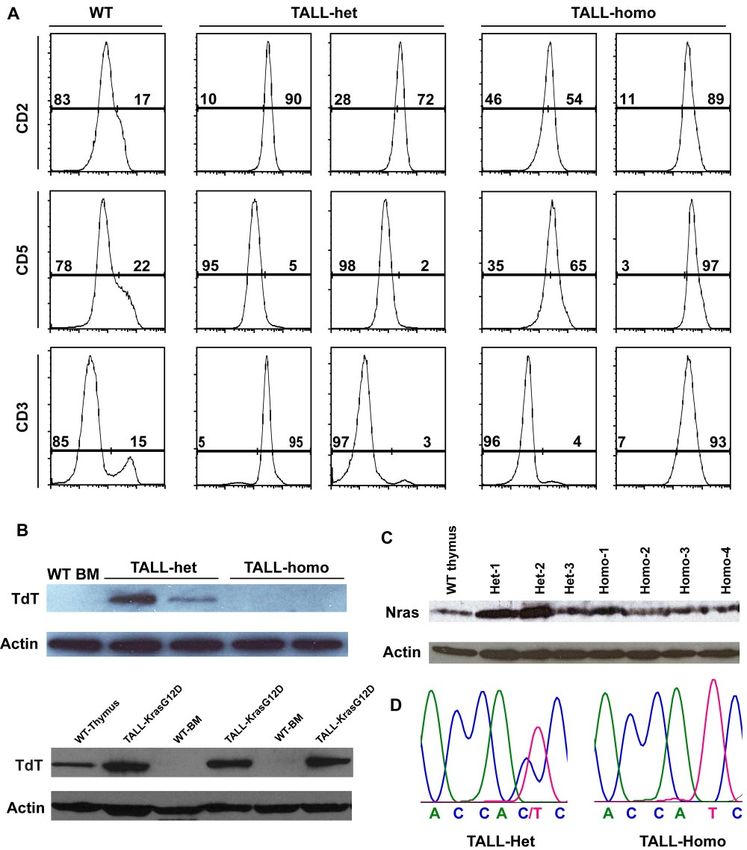
BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 ONCOGENIC NRAS-INITIATED HEMATOPOIETIC MALIGNANCIES 369
IL7R␣ (B12-1), Sca-1 (E13-161.7), TER119, CD34 (RAM34), Fc␥RII/III
(2.4G2), CD25 (3C7) and CD44 (IM7). Anti–c-Kit (2B8) antibodies were
Methods purchased from eBiosciences.
Mice
Cell-cycle analysis
The hypomorphic LSL Nras G12D (LSL Nras G12Dhypo) mice were
constructed as following: BAC clones covering the Nras locus were Cell-cycle analysis was performed essentially as described.27 Lineage
purchased from BACPAC Resources. A 6.6 Kb fragment double-digested markers (CD3, CD4, CD8, CD19, B220, TER119, Gr1, and IgM) and
with EcoRI and HindIII was cloned into the vector pGEM-3Z (Promega). IL-7R␣ were stained with biotin conjugated antibodies followed by PECy7
G12D mutation and intron 1 mutations (changing WT sequence from conjugated streptavidin. Cells were also simultaneously stained for FITC-
gtaattgctgcttttctacag to gtaattgctgcgtttaaacag) were introduced via a site- Ki67 (BD Biosciences), PE-CD45.1, APC-cKit, PerCP-Sca1, and DAPI
directed mutagenesis kit according to the manufacturer’s instructions (Invitrogen). The stained cells were analyzed on a LSRII (BD Biosciences).
(Stratagene). A PmeI site was created in intron 1 to insert the LoxP-stop
casette-LoxP. The targeting construct was electroporated into V6.5 embry- Flow cytometric analysis of phospho-ERK1/2
onic stem (ES) cells and G418 resistant colonies were screened by genomic
Southern blot analysis. The germ line-transmitted LSL mice were generated Phosphorylated ERK1/2 in defined Lin⫺ c-Kit⫹ cells were analyzed
by tetraploid injection of correct ES cells. ES cells were screened using essentially as previously described.19 U0126 (Cell Signaling Technology)
genomic Southern analysis. The mice were backcrossed into C57BL/6 was mixed with cells for 30 minutes before GM-CSF stimulation.
genetic background for more than 10 generations. The embryos and adult
mice were genotyped using primers 5⬘ WT (5⬘-CCACGTGTATC- Murine bone marrow transplantation
GAATGGGTGCCTTAG-3⬘), 3⬘ WT (5⬘-GGGATCATATTCATCCA-
CAAAGTGG-3⬘), and 3⬘ mt (5⬘-GCGGCCAAACGCAGCATTAC-3⬘). Bone marrow transplantation was performed as described.19 For secondary
All mouse lines were maintained on a pure C57BL/6 genetic back- transplantation, recipient mice (CD45.1⫹) were sublethally irradiated
ground (N ⬎ 10). The conditional Nras G12D allele with normal expres- (650 rads). Subsets of primary bone marrow tumor cells were sorted using
sion level comparable with WT allele is described in Haigis et al.18 Mice FACSAria II (BD Biosciences). Different doses of tumor cells (CD45.2⫹)
bearing this allele were crossed to Mx1-Cre mice to generate mice carrying were mixed with 2.5 ⫻ 105 helper cells (CD45.1⫹ splenocytes) and
both alleles (LSL Nras/⫹; Mx1-Cre). LSL Nras G12D/⫹; Mx1-Cre males transplanted into recipient mice.
were crossed to LSL Nras G12D/⫹ females to generate LSL Nras
G12D/G12D; Mx1-Cre mice. Genotyping of the adult mice was performed Histopathology
as described in Haigis et al.18
Mouse organs were fixed in 10% neutral buffered formalin (Sigma-Aldrich)
Mox2 Cre mice were purchased from Jackson Laboratories. CD45.1-
and further processed at the Histology Lab of the University of Wisconsin
positive congenic C57BL/6 recipient mice were purchased from the
Carbone Cancer Center.
National Cancer Institute.
To induce Mx1-Cre expression, 5-6 week old mice were injected
intraperitoneally with 250 g of polyinosinic-polycytidylic acid (pI-pC; TCR- genomic Southern
Sigma Aldrich) every other day for 2 doses. All the experiments were Genomic DNA samples were prepared using the Puregene Genomic DNA
performed 2 days after the second injection of pI-pC unless specified. The Purification Kit (QIAGEN) per manufacturer’s instructions. Rearrangement
injected mice were monitored daily for evidence of disease. All experiments at the T-cell receptor  locus was detected as previously described.28
were conducted with the ethical approval of International Association for
Assessment and Accreditation of Laboratory Animal Care at the University
DNA sequencing of Notch1
of Wisconsin-Madison.
Genomic DNA was isolated from tumor cells using the Puregene Genomic
Sequence analysis of Nras G12 codon DNA Purification Kit (QIAGEN). Amplification of Notch1 exons 26, 27,
and 34 was performed as described previously.29 PCR products were
Total RNAs and genomic DNAs were extracted and analyzed for Nras G12 subcloned into pGEM-T Easy vector (Promega). Ten to 15 clones were
codon as described.19 randomly selected and sequenced at the Biotechnology Center (University
of Wisconsin-Madison). The sequencing data were analyzed using the
Western blot analysis Lasergene Version 8.0 software (DNASTAR).
TER119-negative fetal liver cells (enriched for erythroid progenitors) were
purified from E14.5 individual embryos using the StemSep magnetic bead
system (StemCell Technologies Inc)22 and Western blot analysis was Results
performed essentially as described.23 To detect the levels of total Nras
proteins and TdT in tumor samples, 40 and 20 g of cell lysates were loaded Mice expressing Nras G12Dhypo/G12Dhypo develop normally and
in each lane, respectively. Anti-Nras (F155) and anti-TdT (N-20) antibodies are tumor-free
were purchased from Santa Cruz Biotechnology, whereas anti–mouse
-actin antibodies were from Sigma-Aldrich. To study the dose-dependent role of endogenous oncogenic Nras in
leukemogenesis, we generated a conditional hypomorphic onco-
Flow cytometric analysis of hematopoietic tissues genic Nras allele (LSL Nras G12Dhypo; Figure 1A). Through a
computational modeling design,30 mutations in Nras intron 1 were
For lineage analysis in peripheral blood, bone marrow, spleen, and thymus created in sites adjacent to the splicing acceptor to lower the
tissues, flow cytometric analyses were performed as previously described.24
splicing efficiency of the transcript and to insert the floxed STOP
Myeloid progenitors in bone marrow and spleen were analyzed as
cassette (see “Mice” for details). Thus, the floxed STOP cassette in
previously described.25,26 The stained cells were analyzed on a FACSCali-
bur or LSRII (BD Biosciences).
our hypomorphic allele is at a different position from that in the
Directly conjugated antibodies specific for the following surface allele created by Haigis et al.18 Correct targeting in ES cells was
antigens were purchased from BD Biosciences: CD45.1 (A20), CD45.2, confirmed using Southern blot analysis with a 5⬘ end and a 3⬘ end
(104) B220 (6B2), CD19 (1D3), Thy1.2 (53-2.1), Mac-1 (M1/70), Gr-1 probe (Figure 1B). Germ line transmission of the conditional allele was
(RB6-8C5), CD4 (RM4-5), CD8 (53-6.7), CD3 (145-2C11), IgM (II/41), confirmed by genotyping PCR (data not shown). The presence of the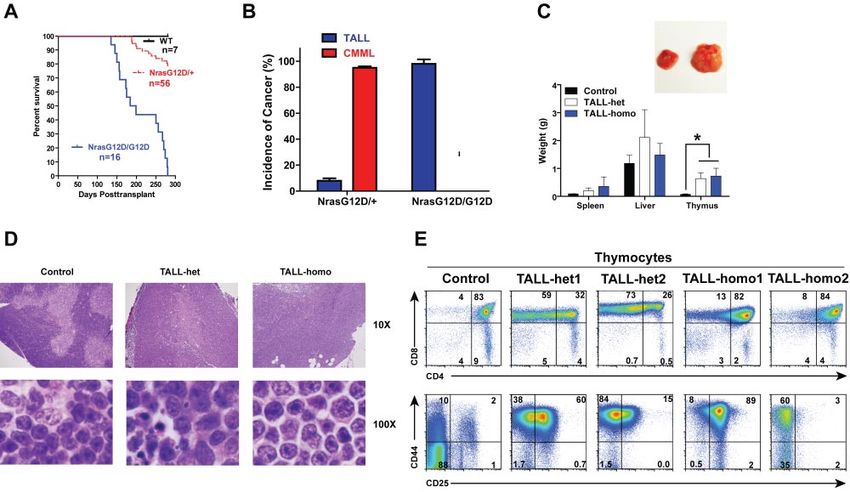
370 WANG et al BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 Figure 1. Construction of a conditional hypomorphic Nras G12D allele. (A) Schematic diagram of WT Nras allele, targeting vector, and constructed LSL (LoxP-STOP cassette-LoxP) Nras G12Dhypo allele. The asterisk shows the substitution of amino acid aspartic acid for glycine by mutation of GGT to GAT at the codon 12. Please see “Mice” for the details of intron 1 mutations. The positions of the probes for Southern blotting are shown. (B) Southern blot analysis of BglII digested genomic DNA isolated from different ES cell clones to confirm correct targeting at the endogenous Nras locus. The WT allele is denoted by the 10.7 KB fragment. The correctly targeted LSL allele is indicated by the 7.4 KB fragment using the 5⬘ internal probe and by the 5.6 Kb fragment using the 3⬘ external probe. (C) Direct sequencing with a reverse primer of genomic DNA isolated from WT and germ line transmitted LSL Nras G12Dhypo mice demonstrates the presence of G12D mutation at the Nras locus. Arrows indicate the WT and mutated nucleotides at the codon 12. (D) Evaluation of recombination efficiency of Mox2-Cre and expression level of Nras G12D hypo allele in E14.5 fetal liver erythoid progenitors. Because Mox2-Cre recombines the conditional LSL cassette and leads to Nras G12Dhypo expression, we refer the compound mice harboring both LSL and Mox2-Cre alleles as Nras G12Dhypo/⫹; Mox2-Cre/⫹. We further crossed Nras G12Dhypo/⫹; Mox2-Cre/⫹ mice to LSL Nras G12Dhypo/⫹ (LSL/⫹) mice and generate Nras G12Dhypo/LSL progenies inherit a recombined Nras G12Dhypo allele and a nonrecombined LSL allele from parents but does not carry Mox2-Cre allele. To simplify the genotyping results, we omit the Mox2-Cre status. Southern blot analysis of SpeI digested genomic DNA using the 3⬘ external probe confirmed the recombination efficiency at the endogenous Nras locus (top panel). Expression levels of total Nras were measured by Western blotting and normalized against actin using the Molecular Analyst Version 1.4 software (Bio-Rad). G12D point mutation in germ line transmitted mice was confirmed by Blood Web site; see the Supplemental Materials link at the top of genomic sequencing (Figure 1C). the online article). None of these mice display any morphologic We examined the expression level of oncogenic Nras at the abnormalities. We killed 5 LSL Nras G12Dhypo/G12Dhypo;Mox2- protein level. We crossed LSL Nras G12Dhypo/⫹ mice to Mox2- Cre/⫹ mice at 12 months and 10 at 18-24 months, and examined Cre/⫹ mice to generate compound mice carrying both alleles. the thymus, spleen, peripheral blood, liver, lung, kidney, and Mox2-Cre efficiently activates oncogenic Nras expression in intestine. We did not observe any hyperplasia phenotypes (data not epiblasts beginning at E5, as judged by Southern blot analysis shown). Our results indicate that expression of Nras G12Dhypo/ (Figure 1D top panel).31 Analysis of E14.5 fetal liver erythroid G12Dhypo is insufficient to cause developmental abnormalities and progenitors showed that Nras G12Dhypo/⫹ expresses oncogenic cancers. Nras at a level equivalent to 25%-40% of single copy of the Nras We obtained another conditional oncogenic Nras allele (LSL WT allele (Figure 1D bottom panel). Thus, Nras G12Dhypo/ Nras G12D) independently generated by Haigis et al.18 This allele G12Dhypo expresses 50%-80% of single copy of the Nras WT allele. expresses oncogenic Nras at a comparable level to the WT Nras To our surprise, despite efficient recombination at the Nras locus, allele.19 We crossed LSL Nras G12D/⫹ mice to Mox2-Cre. None of both LSL Nras G12Dhypo/⫹; Mox2-Cre/⫹ mice and LSL Nras LSL Nras G12D/⫹; Mox2-Cre/⫹ mice were born alive (supplemen- G12Dhypo/G12Dhypo; Mox2-Cre/⫹ mice were viable and generated tal Table 2), suggesting that early embryonic expression of Nras with the expected percentages (supplemental Table 1, available on G12D/⫹ leads to embryonic lethality. Our data are consistent with
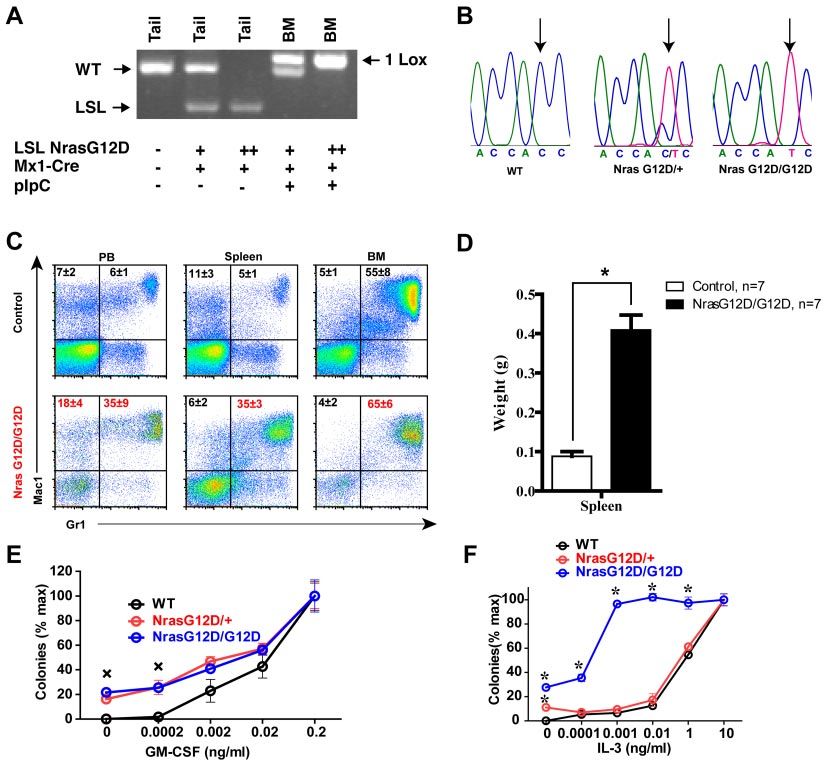
BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 ONCOGENIC NRAS-INITIATED HEMATOPOIETIC MALIGNANCIES 371
Figure 2. Somatic activation of Nras G12D/G12D but not Nras G12D/ⴙ leads to acute myeloproliferative disease. Five- to 6-week-old mice were injected with pI-pC as
described in “Mice.” Two days after the second pI-pC injection, different tissues were isolated and analyzed. Nras G12D/⫹ and Nras G12D/G12D refer to pI-pC treated
compound mice expressing monoallelic and biallelic oncogenic Nras, respectively, as described in “Somatic activation of Nras G12D/G12D but not Nras G12D/⫹ leads to an
acute myeloproliferative disease.” (A) Genotyping analysis of genomic DNA to detect WT allele, LSL allele, and recombined LSL allele (1 LoxP allele). (B) Total RNA was
extracted from bone marrow cells. Direct sequencing of RT-PCR amplified Nras gene using a reverse primer to confirm the sequences at the codon 12. Arrows indicate the WT
and mutated nucleotides at the codon 12. (C) Flow cytometric analysis of peripheral blood (PB), spleen and bone marrow (BM) cells isolated from control (n ⫽ 5) and Nras
G12D/G12D (n ⫽ 5) mice using myeloid lineage specific markers. Debris and unlysed red blood cells (low forward scatter) and dead cells (propidium iodide positive) were
excluded from analysis. Data are presented as averages ⫹ SDs. (D) Splenomegaly in Nras G12/G12D mice. Results are presented as the average of spleen weights ⫹ SD.
*P ⬍ .01. (E, F) 5 ⫻ 104 bone marrow cells isolated from control, Nras G12D/⫹, and Nras G12D/G12D mice were plated in duplicate in semisolid medium with or without
GM-CSF (E) or IL-3 (F). The data are presented as average percentages (from multiple mice of each group) of maximum number of colonies formed in culture with 0.2 ng/mL of
GM-CSF or 10 ng/mL of IL-3. Student t test was performed. Error bars show SD. (E) Crosses indicate P ⬍ .01. (F) *P ⬍ .05.
a recent report from Dr Martin Zenker’s group.32 In this report, endogenous locus.33 We refer to pI-pC–treated compound mice
they failed to identify a germ line oncogenic NRAS mutation in as Nras G12D/⫹ and Nras G12D/G12D mice, respectively, and
917 Noonan Syndrome patients who are negative for previously pI-pC–treated Mx1-Cre mice as WT control mice throughout
known mutations in the Ras pathway, suggesting that germ line this manuscript.
oncogenic NRAS leads to embryonic/fetal lethality. pI-pC treatment of LSL Nras G12D/⫹; Mx1-Cre and LSL Nras
G12D/G12D; Mx1-Cre mice efficiently induced oncogenic Nras
Somatic activation of Nras G12D/G12D but not Nras G12D/ⴙ expression in bone marrow (Figure 2A-B). Two days after the
leads to an acute myeloproliferative disease
second injection of pI-pC, genotypic analysis of genomic DNA
We further studied oncogenic Nras-initiated hematopoietic malig- showed that the recombined allele was readily detectable in bone
nancies using the normally expressed LSL Nras G12D allele.18 We marrow and gain of the recombined allele was associated with the
crossed LSL Nras G12D/⫹ mice to Mx1-Cre transgenic mice to loss of the LSL allele (Figure 2A). Sequencing of RT-PCR
generate compound mice (LSL Nras G12D/⫹; Mx1-Cre) and amplification products confirmed that oncogenic Nras was effi-
further crossed LSL Nras G12D/⫹; Mx1-Cre mice to LSL Nras ciently transcribed at the mRNA level (Figure 2B).
G12D/⫹ mice to generate compound mice (LSL Nras G12D/ Two days after the second injection of pI-pC, all of the Nras
G12D; Mx1-Cre). Administration of pI-pC in compound mice G12D/⫹ mice were grossly unremarkable; both white blood cell
stimulates endogenous IFN production and thus induces Cre counts and differentials were normal.19 In contrast, myeloprolifera-
expression from the IFN-␣/–inducible Mx1 promoter, which in tive phenotypes were prominent in all of the Nras G12D/G12D
turn leads to the expression of oncogenic Nras from its mice (n ⫽ 20). Diseased animals showed marked splenomegaly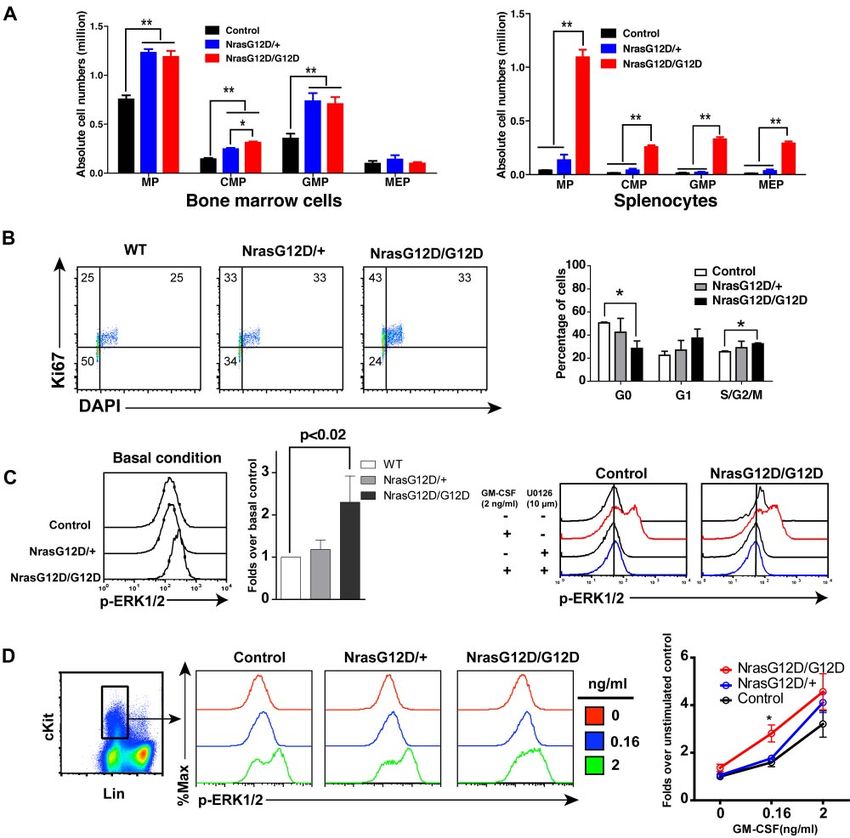
372 WANG et al BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2
(Figure 2D). Flow cytometric analysis using myeloid cell–specific (p-ERK1/2) levels in vivo (Figure 3C). P-ERK1/2 in Lin–/low c-Kit⫹
markers demonstrated a predominantly granulocytic/monocytic cells (enriched for MPs) were analyzed using multiparameter flow
myeloid hyperplasia in the peripheral blood, spleen, and bone cytometry.19 As expected, the ERK pathway was hyperactivated in
marrow of diseased mice (Figure 2C). These phenotypes closely Nras G12D/G12D cells over control and Nras G12D/⫹ cells. To
resembled the acute myeloproliferative disease (MPD) developed determine whether the higher intensity of p-ERK1/2 staining in
in Kras G12D mice in a similar experimental setting.24,34,35 As in Nras G12D/G12D cells truthfully represents elevated p-ERK1/2
Kras G12D mice, MPD develops in Nras G12D/G12D mice level, we added U0126, a MEK inhibitor, before fixation or
without pI-pC injections (n ⫽ 5). GM-CSF stimulation (Figure 3C). Our data show that in the
To determine whether Nras G12D/G12D myeloid progenitors presence of U0126, the p-ERK1/2 level in Nras G12D/G12D cells
display any abnormal growth patterns, bone marrow cells isolated was shifted back to a level comparable with that in control cells,
from control, Nras G12D/⫹, or Nras G12D/G12D mice were indicating an elevated ERK1/2 activation in Nras G12D/G12D
plated in semisolid cultures in the presence of various concentra- cells in vivo. Second, total bone marrow cells were deprived of
tions of mGM-CSF (Figure 2E) or IL-3 (Figure 2F). Bone marrow serum and cytokines and stimulated with various concentrations of
cells from Nras G12D/⫹ and Nras G12D/G12D mice, but not GM-CSF (Figure 3D). The cells were fixed and permeabilized after
controls, formed significant numbers of CFU-GM colonies in the cytokine stimulation. P-ERK1/2 was analyzed in Lin–/low c-Kit⫹
absence of exogenous cytokines. In the presence of GM-CSF or cells. We found that Nras G12D/G12D does not constitutively
IL-3, Nras G12D/⫹ and Nras G12D/G12D colonies were signifi- activate ERK. Rather, it significantly hyperactivates ERK at a low
cantly enlarged compared with that of controls in a dose-dependent concentration of GM-CSF. The magnitude of ERK hyperactivation
manner. Nras G12D/⫹ cells did not show characteristic hypersensi- is similar to that of basal in vivo levels shown in Figure 3C.
tivity to both cytokines, whereas Nras G12D/G12D cells displayed We also examined the activation of Stat5 in Nras G12D/G12D
hypersensitivity to IL-3 but not to GM-CSF. These results are in cells (supplemental Figure 2). Our results demonstrate that activa-
sharp contrast to those of mice expressing oncogenic Kras in a tion of Stat5 in Nras G12D/G12D cells is indistinguishable from
similar experimental system34,35; their bone marrow cells show that in control or Nras G12D/⫹ cells. This result is similar to that
hypersensitivity to both IL-3 and GM-CSF. obtained from Kras G12D cells.36
Oncogenic Nras hyperactivates ERK and promotes cell Nras G12D/G12D mice die with a myeloproliferative disease
proliferation in myeloid progenitors in a dose-dependent manner
After pI-pC injections, control, Nras G12D/⫹, and Nras G12D/
We analyzed the myeloid progenitors (MPs, Lin⫺ IL7R␣⫺ Sca1⫺ G12D mice were kept for an extended period of time. Approxi-
c-Kit⫹) in the bone marrow and spleen of control, Nras G12D/⫹ mately 50% of Nras G12D/⫹ mice died with either histiocytic
and Nras G12D/G12D mice. In bone marrow, the percentages of sarcoma (predominant) or a chronic MPD (occasional) closely
MPs in control and Nras G12D/G12D mice were comparable with resembling CMML at 12 months,19 whereas all of the Nras
each other (P ⫽ .11) but significantly lower than that of Nras G12D/G12D mice died with an acute MPD (Figure 4A). At a
G12D/⫹ mice (P ⬍ .005) (supplemental Figure 1). However, moribund stage, Nras G12D/G12D mice displayed a ⬃ 7-fold
because of the hypercellularity in Nras G12D/G12D bone marrow, increase in their average spleen weights (Figure 4B). Extramedul-
the absolute numbers of MPs in both Nras G12D/⫹ and Nras lary hematopoiesis was evident with various proportions of granu-
G12D/G12D mice were comparable with each other but ⬃ 2-fold locytic, monocytic, erythroid, and megakaryocytic lineage cells
over that of control mice (Figure 3A). The increased numbers of (Figure 4C). Flow cytometric analysis using myeloid cell specific
MPs in Nras G12D/⫹ and Nras G12D/G12D mice were mainly markers revealed a predominantly granulocytic/monocytic myeloid
because of the expansion of common myeloid progenitor (CMP) hyperplasia in the peripheral blood, spleen, and bone marrow of
and granulocyte-monocyte progenitor (GMP) compartments. In diseased mice (supplemental Figure 3). Complete blood counts
spleen, the percentages of MPs in control and Nras G12D/⫹ mice were performed on peripheral blood samples obtained from control
were comparable with each other (P ⫽ .43) but lower than that of and Nras G12D/G12D mice (Figure 4D). The median white blood
Nras G12D/G12D mice (supplemental Figure 1). Because of the cell count in Nras G12D/G12D mice was significantly elevated
marked splenomegaly in Nras G12D/G12D mice, the absolute (18.4 ⫻ 103/L at 4 weeks after pI-pC injections and 53 ⫻ 103/L
numbers of MPs, CMPs, GMPs, and MEPs (megakaryocyte- at a moribund stage versus 7.8 ⫻ 103/L in controls). The median
erythroid progenitors) in these animals were all much higher than platelet count in Nras G12D/G12D mice was significantly lower
those of control and Nras G12D/⫹ mice (Figure 3A). than that in controls (936 ⫻ 103/L at 4 weeks after pI-pC
To determine whether the expansion of the MP compartment is injections and 640 ⫻ 103/L at a moribund stage versus
associated with the hyperproliferation of these cells, we performed 1451 ⫻ 103/L in controls). Similarly as Nras G12D/G12D mice at
flow cytometry-based cell-cycle analysis of MPs in control, Nras 2 days after pI-pC treatment, moribund mice showed a significantly
G12D/⫹ and Nras G12D/G12D bone marrow (Figure 3B).27 expanded myeloid compartment in both bone marrow and spleen
Through intracellular staining for Ki67 and DNA content (using (Figure 4E).
DAPI), we could define G0 (Ki67⫺, 2n DNA content), G1 (Ki67⫹,
2n), and S/G2/M (Ki67⫹, ⬎ 2n) subpopulations within the MP Recipient mice transplanted with Nras G12D/G12D bone
population. Our results demonstrated an oncogenic Nras dose- marrow cells develop TALL with a 100% penetrance
dependent decrease in the proportion of MPs in G0 with a
corresponding increase in the proportions in G1 and S/G2/M. To study the hematopoietic cell-autonomous role of oncogenic
We studied whether the hyperproliferation of MPs corresponds Nras signaling in leukemogenesis, we transplanted 2.5 ⫻ 105 bone
to the hyperactivation of ERK, the major signaling component marrow cells (CD45.2ⴙ) isolated from control, Nras G12D/⫹, or
downstream of oncogenic Nras (Figure 3C-D). First, total bone Nras G12D/G12D mice along with same number of WT competitor
marrow cells were freshly isolated and immediately fixed and cells (CD45.1ⴙ) into lethally irradiated recipient mice (Figure 5A).
permeabilized (basal condition) to measure phospho-ERK1/2 Approximately 95% of recipient mice transplanted with NrasBLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 ONCOGENIC NRAS-INITIATED HEMATOPOIETIC MALIGNANCIES 373 Figure 3. Oncogenic Nras signaling engages myeloid progenitors into cell cycle and leads to their expansion in a dose-dependent manner. Different tissues were isolated and analyzed 2 days after pI-pC injections. (A) Quantitative analysis of myeloid progenitor (MP) compartment in bone marrow and spleen of control, Nras G12D/⫹, and Nras G12D/G12D mice. CMP indicates common myeloid progenitor; GMP, granulocyte-monocyte progenitor; and MEP, megakaryocyte-erythroid progenitor. Results are presented as averages ⫹ SDs. Student t test was performed: *P ⬍ .05, and **P ⬍ .01. (B) Cell cycle analysis of MPs in bone marrow of control, Nras G12D/⫹, and Nras G12D/G12D mice. Cell-cycle phases are defined as G0 (Ki67⫺, DAPIlo), G1 (Ki67⫹, DAPIlo), and S/G2/M (Ki67⫹, DAPIhi). The percentages of MPs in individual cell-cycle phases are indicated on the density plots. Average values ⫹ SDs are shown in the right graph. Student t test was performed: *P ⬍ .05. (C,D) Phospho-flow analysis of p-ERK1/2 in Lin⫺/low c-Kit⫹ bone marrow cells of control, Nras G12D/⫹, and Nras G12D/G12D mice 2 days after the second pI-pC injection. (C) Total bone marrow cells were freshly isolated and stimulated with or without 2ng/mL of GM-CSF at 37°C for 10 minutes. Basal condition is defined as without GM-CSF stimulation. U0126 was mixed with cells for 30 minutes before fixation or GM-CSF stimulation. Levels of p-ERK1/2 were measured using phospho-specific flow cytometry. Nonneutrophil Lin⫺/low c-Kit⫹ cells were gated for data analysis. Results obtained from one representative experiment are shown (left panel). Quantification of 6 independent experiments is shown as average values ⫹ SDs (middle panel). Solid lines indicate the median intensity of p-ERK1/2 in control cells without GM-CSF stimulation (right panels). (D) Total bone marrow cells were serum- and cytokine-starved for 1 hour and stimulated with various concentrations of GM-CSF (0, 0.16 and 2 ng/mL) at 37°C for 10 minutes. Gating strategy and plots of p-ERK1/2 are representative of 4 independent experiments. To quantify the activation of ERK1/2, median intensities of p-ERK1/2 at different GM-CSF concentrations in different animals are compared with control cells at 0 ng/mL, which is arbitrarily set at 1. Average values ⫾ SDs are shown in the right graph. Student t test was performed: *P ⬍ .05. G12D/⫹ cells developed CMML 6-24 months after transplantation none of them developed sustained CMML-like phenotypes nor did and ⬃ 8% of recipient mice developed acute TALL (TALL-het) they develop AML. 6-7 months after transplantation (some of the animals developed We characterized both TALL-het and TALL-homo mice (Figure both of the diseases; Figure 5A-B).19 In contrast, all of the recipient 5C-E). Both groups of animals showed variable hepatosplenomegly mice transplanted with Nras G12D/G12D cells died of TALL with T-cell infiltration at the moribund stage, whereas their thymi (TALL-homo) 5-9 months after transplantation (Figure 5A-B) and were consistently enlarged 8- to 10-fold over controls (Figure 5C).
374 WANG et al BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2
Figure 4. All the Nras G12D/G12D mice die with a severe myeloproliferative disease. After pI-pC injections, control and Nras G12D/G12D mice were kept for an extended
period of time until Nras G12D/G12D mice reached a moribund stage. (A) Kaplan-Meier comparative survival analysis of control and Nras G12D/G12D mice. Cumulative
survival was plotted against days after the first pI-pC injection. (B) Splenomegaly in Nras G12D/G12D mice. Results are presented as averages of spleen weights ⫹ SDs.
(C) Representative histologic H&E sections from spleen show an extensive infiltration of myelomonocytic cells and extramedullary hematopoiesis in Nras G12D/G12D mice.
(D) Complete blood count was performed on peripheral blood samples drawn from control and Nras G12D/G12D mice. *P ⬍ .05. (E-F) Quantitative analysis of myeloid
progenitor (MP) compartment in bone marrow (E) or spleen (F) of control and moribund Nras G12D/G12D mice as described in Figure 3. Results are presented as averages ⫹
SDs. *P ⬍ .05.
Compared with control thymus, which showed a distinct pattern of individual diseased mice that had been transplanted with Nras
cortical and medullar architecture and was filled with maturing G12D/⫹ or Nras G12D/G12D cells (supplemental Figure 5). Bone
T cells with dense nuclear staining, the TALL-het and -homo marrow cells (1 ⫻ 104) were transplanted into sublethally irradi-
thymus demonstrated a completely effaced thymic architecture ated secondary recipient mice. All secondary recipient mice
filled with actively proliferating T-cell blasts and scattered tangible- developed TALL and died within 2 months after transplantation
body macrophages (Figure 5D). We further analyzed both TALL- (supplemental Figure 5A). Further fractionation of tumor cells
het and -homo thymocytes with various T-cell markers (Figure 5E). demonstrated that the activity of tumor initiating cells is restricted
Both groups of tumor samples were double positive for CD4 and in CD8⫹ T cells (data not shown), and particularly enriched in
CD8 and positive for CD44, similar to the T-cell disease initiated CD8⫹ Sca1⫺ cKit⫺ cells (supplemental Figure 5B).
by endogenous oncogenic Kras mutation.24,37
To examine the development of T-cell malignancies, we ana-
lyzed recipient mice transplanted with Nras G12D/G12D cells Both TALL-het and TALL-homo tumors are predominantly
4 months after transplantation (supplemental Figure 4). At this monoclonal and acquire clonal secondary Notch1 mutations
pre-TALL stage, the thymus size was normal compared with To determine whether the oncogenic Nras-initiated TALL is clonal
controls. However, there was significant expansion of CD4⫺CD8⫺ in origin, we performed genomic Southern analysis of the TCR
cells. These double-negative cells started to show up-regulation locus in multiple TALL-het and TALL-homo tumor samples
of CD44. (Figure 6A).24 The probe hybridizes to the V region of the T-cell
receptor  locus.28 Our results indicate that the tumors are
Oncogenic Nras-initiated T-cell diseases are transplantable to
secondary recipients
predominantly monoclonal.
We next sequenced the Notch1 gene for mutations in the
To determine whether oncogenic Nras-initiated T-cell malignan- heterodimerization (exons 26 and 27) or PEST (exon 34) domains
cies are transplantable, we isolated bone marrow cells from that have been previously observed in both human and murineBLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 ONCOGENIC NRAS-INITIATED HEMATOPOIETIC MALIGNANCIES 375 Figure 5. Oncogenic Nras initiates TALL in a dose-dependent manner. Lethally irradiated mice (CD45.1⫹) were transplanted with 2.5 ⫻ 105 total bone marrow cells of control, Nras G12D/⫹, or Nras G12D/G12D mice along with same number of competitor cells. (A) Kaplan-Meier survival curves of reconstituted mice. Cumulative survival was plotted against days after transplantation. (B) Disease distribution patterns in recipient mice transplanted with Nras G12D/⫹ or Nras G12D/G12D cells. (C) Hepatosplenomegaly and enlarged thymi in TALL-het and -homo mice. (Top) Enlarged thymus in a representative recipient mouse that developed a TALL disease. (Bottom) Results are presented as averages of spleen, liver, or thymus weights ⫹ SDs. Student t test was performed: *P ⬍ .001. (D) Representative histologic H&E sections of thymus from control, TALL-het, and TALL-homo mice. (E) Flow cytometric analysis of total thymocytes of representative control, TALL-het, and TALL-homo mice. models of TALL. No mutations were detected at the het- Figure 6B). Consistent with a recent report,39 4 of 5 Notch1 erodimerization cleavage site, but 2 of 4 TALL-het tumors and mutation-positive tumors contain heterogeneous Notch1 muta- 3 of 4 TALL-homo tumors harbored clonal PEST domain tions (supplemental Figure 6C). mutations (Figure 6B). These mutations are all insertional muta- tions, which lead to frame shifts and premature stops in the PEST Oncogenic Nras-initiated TALL cells are sensitive to domain. Compared with TALL-het and TALL-homo, oncogenic ␥-secretase inhibitor and Ras downstream effector inhibitors Kras-initiated TALL has a significantly shorter latency; the dis- eased animals die 1.5-4 months after transplantation.24,37,38 To determine whether the growth of TALL-het and -homo cells Similarly as shown before,37 these tumors are predominantly depends on abnormal Notch1 signaling, we treated various TALL oligoclonal (supplemental Figure 6A) and 5 of 10 tumors cells with GSI XXI, a small molecule inhibitor of ␥-secretase. The contain Notch1 mutations in the PEST domain (supplemental tested cells included TALL-het primary tumor cells, and cell lines Figure 6. Oncogenic Nras-initiated TALL tumors contain clonal Notch1 mutations. (A) Southern blot analysis of genomic DNA obtained from TALL-het and TALL-homo tumors. The blot was hybridized with a probe to the V region of T-cell receptor . (B) Sequence analysis of the exon 34 of Notch1 in control, TALL-het, and TALL-homo thymocytes.
376 WANG et al BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2
derived from TALL-het and TALL-homo tumors, respectively. All ity. Second, at its normal endogenous level, the tumor transforming
of them carried a Notch1 mutation in the PEST domain. All of the capability of oncogenic Nras is not only dose-dependent but also
tested cells were sensitive to inhibition of Notch signaling at cell type–dependent. Third, although both Nras G12D/⫹ and Nras
concentrations as low as 0.05M (supplemental Figure 7A). The G12D/G12D initiate TALL, tumor cells appear to be transformed at
decrease in cell growth resulted from increased apoptotic cell death different developmental stages and through distinct genetic mecha-
as demonstrated by a dose-dependent increase in annexin V- nisms of tumor progression.
positive cells (data not shown).
To investigate the effects of inhibition of Ras downstream Somatic expression of Nras G12D/ⴙ versus Nras G12D/G12D
pathways, we also treated the TALL-homo cell line with the MEK
inhibitor, U0126 (supplemental Figure 7B). Treatment for 48 hours Although the Ras genes have long been established as proto-
caused a dose-dependent inhibition of cell growth. Moreover, oncogenes, substantial evidence indicates that WT Ras genes act as
because both Notch1 and Ras signaling pathways affect PI3K tumor suppressor genes in different tumor models.45,46 Compared
signaling,40,41 we treated TALL-homo cells with rapamycin (supple- with Nras G12D/⫹, Nras G12D/G12D not only doubles the dose of
mental Figure 7C) and LY294002 (supplemental Figure 7D). oncogenic Nras but also loses the potential restraint from the WT
Again, both inhibition treatments resulted in a dose-dependent Nras gene. The latter might also contribute to the acute MPD
suppression of cell growth. The specificity of XXI to inhibit Notch phenotypes in Nras G12D/G12D mice.
signaling and U0126 to block ERK1/2 activation is shown in
supplemental Figure 7E. Somatic versus bone marrow-specific expression of Nras
G12D/G12D
Endogenous Nras G12D/ⴙ signaling promotes TALL in the
In our new model, we found that widespread expression of Nras
presence of an intact WT Nras allele
G12D/G12D in mice leads to acute MPD with a complete
We characterized both TALL-het and -homo tumors with T-cell penetrance (Figures 2-4), whereas bone marrow-specific expres-
markers expressed at different developmental stages (Figure 7A). sion of Nras G12D/G12D in recipient mice results in 100%
TALL-het tumors are surface CD2⫹ CD5⫺ CD3⫹/⫺, whereas penetrant TALL (Figure 5). A similar phenotypic switch is also seen
TALL-homo tumors are CD2⫹ CD5⫹ CD3⫹/⫺, suggesting that in the case of endogenous oncogenic Kras.24,34,35,37,38 Several
TALL-het and -homo cells may be transformed at different possibilities might account for this observation. First, the MPD
developmental stages. We further analyzed the expression of phenotypes in primary Nras G12D/G12D mice are likely compli-
terminal deoxynucleotidyl transferase (TdT) in both types of cated by systemic interferon-mediated responses, induced Nras
tumor cells (Figure 7B). TdT is usually expressed in primitive G12D/G12D expression in nonhematopoietic cells, and the simul-
T-lymphocytes in the normal thymus as well as in most of cases of taneous expression of Nras G12D/G12D in 80%-90% of myeloid
TALL in both human and mouse.42,43 We found that despite being cells. Thus, the initiation of MPD phenotypes with 100% pen-
morphologically similar and consistent with human TALL, both etrance in the primary mice might be a transient phenomenon
oncogenic Kras-initiated TALL and oncogenic Nras-initiated TALL- attributable to microenvironmental factors and does not necessarily
het are TdT-positive, whereas oncogenic Nras-initiated TALL- imply the long-term maintenance in a hematopoietic-cell autono-
homo tumors are TdT-negative. This suggests a potential role for a mous manner. Thus, we believe that it is critical to adopt the bone
dose-dependent survival/differentiation signal of oncogenic Nras in marrow transplantation system to study the cell-autonomous role of
neoplastic T-cell precursors. genes in leukemogenesis. Second, it is possible that either the MPD
Because up-regulation of oncogenic Nras protein through state or the pI-pC treatment hinders the engraftment of MPD-
genetic (eg UPD of the oncogenic Nras allele) and/or epigenetic initiating cells and thus contributes to our results. Third, it is also
mechanisms is often observed during tumor development in both likely that MPD phenotypes are maintained by genetically altered
human and mouse,14,19,44 we tested whether this mechanism is also hematopoietic stem cells (HSCs). Nras G12D/G12D signaling
involved in the formation of TALL-het and/or TALL-homo tumors. might substantially alter HSC behaviors so that these HSCs no
Western blot analysis showed that TALL-homo tumors express longer sustain MPD phenotypes in recipient mice until they reach a
comparable levels of Nras as control thymocytes, whereas 2 of lethal stage.
3 TALL-het tumors express significantly higher levels of Nras than It is nevertheless surprising that thus far none of the recipient
controls (Figure 7C). Further sequencing analysis of mRNA mice with Nras G12D/G12D cells has succumbed to a myeloid
transcripts and genomic DNA demonstrated that both WT and disease. At the dose of 2.5 ⫻ 105 cells and under the same
oncogenic Nras alleles are intact and transcribed in TALL-het experimental procedure, 10 of 72 recipient mice transplanted with
tumor cells (Figure 7D, n ⫽ 5). Therefore, the up-regulation of Kras G12D cells died with JMML-like phenotypes (B.M. and J.D.,
oncogenic Nras protein in TALL-het tumors is associated with unpublished data, April 2011), whereas 0 of 64 recipient mice of
concomitant up-regulation of WT Nras protein. Nras G12D/G12D cells developed a sustained MPD (P ⫽ .002). It
is likely that Nras G12D/G12D signaling is insufficient to initiate a
strong myeloid disease in a timely manner after transplantation.
Discussion Tumor transforming activity of oncogenic Nras is both
dose-dependent and cell type–dependent
In this study, we created a gradient of oncogenic Nras signaling
ranging from 25%-200% of endogenous monoallelic expression of Overexpression of oncogenic Nras (at least 6-12 fold over endoge-
Nras G12D (supplemental Figure 8). First, we show that expressing nous Nras G12D/⫹) in mouse bone marrow cells efficiently
up to 80% of Nras G12D/⫹ neither affects normal mouse induces CMML/AML within 3-6 months after transplantation.17
embryonic development nor results in tumors. In contrast, early Moreover, increasing overexpression levels of oncogenic Nras
embryonic expression of Nras G12D/⫹ leads to embryonic lethal- leads to stronger phenotypes and significantly shorter diseaseBLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 ONCOGENIC NRAS-INITIATED HEMATOPOIETIC MALIGNANCIES 377 Figure 7. TALL-het tumors are generated through a distinct genetic mechanism from TALL-homo tumors. (A) Flow cytomytric analysis of TALL-het and -homo tumor cells with T-cell markers expressed at different developmental stages. Two representative tumors of 5 for each tumor type are shown. (B) Western blot analysis of TdT expression levels in control cells, oncogenic Nras mutation-initiated TALL-het and -homo tumors (top panels), and oncogenic Kras mutation-initiated TALL-KrasG12D tumors (bottom panels). (C) Western blot analysis of total Nras expression levels in control, TALL-het, and -homo thymocytes. (D) Direct sequencing of RT-PCR amplified products in TALL-het and -homo tumors at the Nras G12 codon. Results are representative of 5 animals for each tumor type. latency, suggesting that stronger oncogenic Nras signaling results CMML transformation to AML. The failure to accelerate CMML in greater tumor transforming potential. Contrary to results from formation by Nras G12D/G12D might be attributed to the confound- oncogenic Nras overexpression, our bone marrow transplant mod- ing emergence of fatal TALL. In contrast, in the T-cell lineage, Nras els show that the tumor transforming potential of oncogenic Nras G12D/G12D signaling dramatically increases the TALL penetrance signaling is not only dose-dependent but also cell type-dependent. but does not significantly shorten the disease latency (Figure 5). In myeloid cells, compared with endogenous Nras G12D/⫹, Nras Apparently, T cells require stronger signaling than myeloid cells G12D/G12D does not accelerate CMML formation nor lead to during leukemia development. Our observation is consistent with
378 WANG et al BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2
the case of different BCR-ABL forms; p190 BCR-ABL shows Notch1 mutations, such mutations in TALL-het tumors are clonal
stronger tyrosine kinase activity and is often associated with ALL, but heterogeneous in oncogenic Kras-initiated T-cell diseases. The
whereas p210 BCR/ABL displays weaker tyrosine kinase activity clonality of Notch1 mutations correlates to the clonality of the
and are predominantly identified in CML patients.47 Our data TALL tumors. The differences observed in these 2 TALL models
emphasize the importance of constructing and studying physiologic regarding tumor latency, penetrance, and clonality likely result
mouse models for human cancers. from the dramatic signaling strengths elicited by endogenous Nras
G12D/⫹ signaling and Kras G12D/⫹ signaling.19,36
T-cell leukemia/lymphoma initiating cells Taken together, our study shows a dose- and cell type–
dependent role of endogenous oncogenic Nras signaling in hemato-
It remains controversial whether TALL is maintained by a rare
poietic malignancies (supplemental Figure 8).
population of tumor cells called tumor-initiating cells (TICs) or by
the majority of tumor cells. Pten deficiency-initiated TALL is
driven by TICs, whose activity is enriched in a CD3⫹ cKitmid
Acknowledgments
subpopulation of tumor cells.48 In contrast, TALL isolated from
E-Nras transgenic mice is sustained by virtually all the tumor We are grateful to Dr Harvey F. Lodish and Dr Rudolf Jaenisch for
cells.49 Interestingly, our TALL models show intermediate frequen- their generous help and support of this project at the early stages.
cies of TICs (supplemental Figure 3). Collectively, these results We thank Dr Zefeng Wang for computational modeling and Dr
suggest that the frequency of TICs in TALL is genetic alteration- Laurie Jackson-Grusby for providing us with the STOP cassette
dependent. Considerably more work is required to address this construct. We are grateful to Drs Kevin Haigis and Tyler Jacks for
issue and the results might vary from case to case. providing us the conditional oncogenic Nras mice. We thank Drs
Norman Drinkwater, Lily Huang, and Shannon Kenney for helpful
Differential mechanisms underlying the generation of TALL-het
discussion and critical comments on the manuscript. We are
and -homo tumors
grateful to Dr Norman Drinkwater for his help in statistical
Although both Nras G12D/⫹ and Nras G12D/G12D initiate analysis.
TALL, the tumor cells appear to be transformed through distinct This work was supported by a Howard Temin Award from the
mechanisms. Two of 3 TALL-het tumors acquired enhanced National Cancer Institute, a Shaw Scientist Award from the Greater
oncogenic Nras signaling through up-regulation of both WT and Milwaukee Foundation, a research grant from the Elsa Pardee
oncogenic Nras proteins (Figure 7C). However, such mechanism is Foundation and from the Wendy Will Case Cancer Fund, an ASH
not detected in TALL-homo tumors. This mechanism is also Scholar Award from the American Society of Hematology, and a
distinct from the findings in Nras G12D/⫹ initiated CMML, in pilot project grant from the American Cancer Society Institutional
which 2 of 5 mice up-regulated the expression of oncogenic Nras Research Grant to J.Z. This project was also supported in part by
protein through UPD of the oncogenic Nras allele.19 It is not clear the UW Institute for Clinical and Translational Research, funded
what leads to the differential mechanism(s) of up-regulating through a National Institutes of Health/NCRR Clinical and Transla-
oncogenic Nras signaling in myeloid cells versus T cells. Nonethe- tional Science Award, 1UL1RR025011.
less, we anticipate that oncogenic NRAS mutations are rare in
TALL because of the low incidence of TALL-het and rare biallelic
NRAS mutations. Indeed, we and others found that oncogenic Authorship
NRAS mutations occur in ⬍ 5% of TALL patients (Bos,4 Roden-
Contribution: J.W. was responsible for experimental design, execu-
huis,11 Neri,12 Lubbert13; Z.L., unpublished results, April 2011).
tion, and writing the manuscript; Z.W. and Q.C. were responsible
Nras G12D/ⴙ initiated versus Kras G12D/ⴙ initiated TALL for ES cell culture and tetraploid injection to generate LSL Nras
G12Dhypo allele; Z.L., Y.L., L.X.T., J.D., M.-J.R., and B.M. were
Morphologically and phenotypically, Nras G12D/⫹ initiated T-cell responsible for experimental execution; K.H.Y. and E.A.R. were
diseases (TALL-het) closely resemble those initiated by Kras responsible for histopathologic analysis and editing of the manu-
G12D/⫹.24,37 Both of them are CD4, CD8 double positive but script; and J.Z. was responsible for experimental design and
prone to be CD8 single positive during lymphoma development, writing of the manuscript.
CD44 positive, TdT positive, and transplantable to secondary Conflict-of-interest disclosure: The authors declare no compet-
recipient mice. Notch1 mutations in the PEST domain are identified ing financial interests.
in ⬃ 50% of each tumor type and tumor cells containing Notch1 The current affiliation for K.H.Y. is Department of Hemato-
mutations are sensitive to ␥-secretase inhibitor XXI. On the other pathology, University of Texas M. D. Anderson Cancer Center,
hand, TALL-het occurs in only ⬃ 8% of recipient mice 6-7 months Houston, TX.
after transplantation, whereas oncogenic Kras initiates T-cell Correspondence: Jing Zhang, McArdle Laboratory for Cancer
diseases 1.5-3.5 months after transplantation with an almost Research, 1400 University Ave, Rm 417A, University of Wisconsin-
complete penetrance. Although both types of tumors contain Madison, Madison, WI; e-mail: zhang@oncology.wisc.edu.
References
1. Barbacid M. ras genes. Annu Rev Biochem. GTPase superfamily: conserved structure and tinct cytogenetic subgroups but does not influ-
1987;56:779-827. molecular mechanism. Nature. 1991;349(6305): ence outcome in patients younger than 60 years.
117-127. Blood. 2005;106(6):2113-2119.
2. Bourne HR, Sanders DA, McCormick F. The
GTPase superfamily: a conserved switch for di- 4. Bos JL. Ras oncogenes in human cancer: a re- 6. Auewarakul CU, Lauhakirti D, Tocharoentanaphol
verse cell functions. Nature. 1990;348(6297):125- C. Frequency of RAS gene mutation and its coop-
view. Cancer Res. 1989;49(17):4682-4689.
erative genetic events in Southeast Asian adult
132.
5. Bowen DT, Frew ME, Hills R, et al. RAS mutation acute myeloid leukemia. Eur J Haematol. 2006;
3. Bourne HR, Sanders DA, McCormick F. The in acute myeloid leukemia is associated with dis- 77(1):51-56.BLOOD, 14 JULY 2011 䡠 VOLUME 118, NUMBER 2 ONCOGENIC NRAS-INITIATED HEMATOPOIETIC MALIGNANCIES 379
7. Vardiman JW. Myelodysplastic/myeloproliferative 21. Smithies O, Kim HS, Takahashi N, Edgell MH. nous promoter induces a myeloproliferative dis-
diseases. Cancer Treatment Research. 2004; Importance of quantitative genetic variations in ease. J Clin Invest. 2004;113(4):528-538.
121:13-43. the etiology of hypertension. Kidney Int. 2000; 36. Van Meter ME, Diaz-Flores E, Archard JA, et al.
8. Elliott MA. Chronic neutrophilic leukemia and 58(6):2265-2280. K-RasG12D expression induces hyperprolifera-
chronic myelomonocytic leukemia: WHO defined. 22. Zhang J, Socolovsky M, Gross AW, Lodish HF. tion and aberrant signaling in primary hematopoi-
Best Pract Res Clin Haematol. 2006;19(3):571- Role of Ras signaling in erythroid differentiation of etic stem/progenitor cells. Blood. 2007;109(9):
593. mouse fetal liver cells: functional analysis by a 3945-3952.
9. Reuter CW, Morgan MA, Bergmann L. Targeting flow cytometry-based novel culture system. 37. Kindler T, Cornejo MG, Scholl C, et al.
the Ras signaling pathway: a rational, mecha- Blood. 2003;102(12):3938-3946. K-RasG12D-induced T-cell lymphoblastic lym-
nism-based treatment for hematologic malignan- 23. Zhang J, Lodish HF. Identification of K-ras as the phoma/leukemias harbor Notch1 mutations and
cies? Blood. 2000;96(5):1655-1669. major regulator for cytokine-dependent Akt acti- are sensitive to gamma-secretase inhibitors.
vation in erythroid progenitors in vivo. Proc Natl Blood. 2008;112(8):3373-3382.
10. Emanuel PD. Juvenile myelomonocytic leukemia
and chronic myelomonocytic leukemia. Leuke- Acad Sci U S A. 2005;102(41):14605-14610. 38. Sabnis AJ, Cheung LS, Dail M, et al. Oncogenic
mia. 2008;22(7):1335-1342. 24. Zhang J, Wang J, Liu Y, et al. Oncogenic Kras- Kras initiates leukemia in hematopoietic stem
induced leukemogeneis: hematopoietic stem cells. PLoS Biol. 2009;7(3):e59.
11. Rodenhuis S, Bos JL, Slater RM, Behrendt H,
van ’t Veer M, Smets LA. Absence of oncogene cells as the initial target and lineage-specific pro- 39. Ashworth TD, Pear WS, Chiang MY, et al.
amplifications and occasional activation of N-ras genitors as the potential targets for final leukemic Deletion-based mechanisms of Notch1 activation
in lymphoblastic leukemia of childhood. Blood. transformation. Blood. 2009;113(6):1304-1314. in T-ALL: key roles for RAG recombinase and a
1986;67(6):1698-1704. 25. Akashi K, Traver D, Miyamoto T, Weissman IL. A conserved internal translational start site in
clonogenic common myeloid progenitor that gives Notch1. Blood. 2010;116(25):5455-5464.
12. Neri A, Knowles DM, Greco A, McCormick F,
rise to all myeloid lineages. Nature. 2000; 40. Rodriguez-Viciana P, Warne PH, Dhand R, et al.
Dalla-Favera R. Analysis of RAS oncogene muta-
404(6774):193-197. Phosphatidylinositol-3-OH kinase as a direct tar-
tions in human lymphoid malignancies. Proc Natl
26. Miyamoto T, Iwasaki H, Reizis B, et al. Myeloid or get of Ras. Nature. 1994;370(6490):527-532.
Acad Sci U S A. 1988;85(23):9268-9272.
lymphoid promiscuity as a critical step in hemato- 41. Gutierrez A, Look AT. NOTCH and PI3K-AKT
13. Lubbert M, Mirro J Jr, Miller CW, et al. N-ras gene
poietic lineage commitment. Dev Cell. 2002;3(1): pathways intertwined. Cancer Cell. 2007;12(5):
point mutations in childhood acute lymphocytic
137-147. 411-413.
leukemia correlate with a poor prognosis. Blood.
1990;75(5):1163-1169. 27. Bereshchenko O, Mancini E, Moore S, et al. He- 42. Suzumiya J, Ohshima K, Kikuchi M, Takeshita M,
matopoietic stem cell expansion precedes the Akamatsu M, Tashiro K. Terminal deoxynucleoti-
14. Dunbar AJ, Gondek LP, O’Keefe CL, et al. 250K dyl transferase staining of malignant lymphomas
single nucleotide polymorphism array karyotyping generation of committed myeloid leukemia-initiat-
ing cells in C/EBPalpha mutant AML. Cancer in paraffin sections: a useful method for the diag-
identifies acquired uniparental disomy and ho- nosis of lymphoblastic lymphoma. J Pathol. 1997;
mozygous mutations, including novel missense Cell. 2009;16(5):390-400.
182(1):86-91.
substitutions of c-Cbl, in myeloid malignancies. 28. Roberts CW, Leroux MM, Fleming MD, Orkin SH.
Highly penetrant, rapid tumorigenesis through 43. Morse HC, 3rd Anver MR, Fredrickson TN, et al.
Cancer Res. 2008;68(24):10349-10357.
conditional inversion of the tumor suppressor Bethesda proposals for classification of lymphoid
15. Edkins S, O’Meara S, Parker A, et al. Recurrent neoplasms in mice. Blood. 2002;100(1):246-258.
gene Snf5. Cancer Cell. 2002;2(5):415-425.
KRAS codon 146 mutations in human colorectal
29. O’Neil J, Calvo J, McKenna K, et al. Activating 44. Sarkisian CJ, Keister BA, Stairs DB, Boxer RB,
cancer. Cancer Biol Ther. 2006;5(8):928-932.
Notch1 mutations in mouse models of T-ALL. Moody SE, Chodosh LA. Dose-dependent onco-
16. MacKenzie KL, Dolnikov A, Millington M, gene-induced senescence in vivo and its evasion
Blood. 2006;107(2):781-785.
Shounan Y, Symonds G. Mutant N-ras induces during mammary tumorigenesis. Nat Cell Biol.
myeloproliferative disorders and apoptosis in 30. Wang Z, Rolish ME, Yeo G, Tung V, Mawson M, 2007;9(5):493-505.
bone marrow repopulated mice. Blood. 1999; Burge CB. Systematic identification and analysis
45. Zhang Z, Wang Y, Vikis HG, et al. Wildtype Kras2
93(6):2043-2056. of exonic splicing silencers. Cell. 2004;119(6):
can inhibit lung carcinogenesis in mice. Nat Genet.
831-845.
17. Parikh C, Subrahmanyam R, Ren R. Oncogenic 2001;29(1):25-33.
NRAS rapidly and efficiently induces CMML- and 31. Tallquist MD, Soriano P. Epiblast-restricted Cre
46. Diaz R, Ahn D, Lopez-Barcons L, et al. The N-ras
AML-like diseases in mice. Blood. 2006;108(7): expression in MORE mice: a tool to distinguish
proto-oncogene can suppress the malignant phe-
2349-2357. embryonic vs. extra-embryonic gene function.
notype in the presence or absence of its onco-
Genesis. 2000;26(2):113-115.
18. Haigis KM, Kendall KR, Wang Y, et al. Differential gene. Cancer Res. 2002;62(15):4514-4518.
effects of oncogenic K-Ras and N-Ras on prolif- 32. Cirstea IC, Kutsche K, Dvorsky R, et al. A re-
47. Li S, Ilaria RL Jr, Million RP, Daley GQ,
eration, differentiation and tumor progression in stricted spectrum of NRAS mutations causes
Van Etten RA. The P190, P210, and P230 forms
the colon. Nat Genet. 2008;40(5):600-608. Noonan syndrome. Nat Genet. 2010;42(1):27-29.
of the BCR/ABL oncogene induce a similar
19. Wang JY, Liu YG, Li ZY, et al. Endogenous onco- 33. Kuhn R, Schwenk F, Aguet M, Rajewsky K. Induc- chronic myeloid leukemia-like syndrome in mice
genic Nras mutation leads to aberrant GM-CSF ible gene targeting in mice. Science. 1995; but have different lymphoid leukemogenic activity.
signaling in granulocytic/monocytic precursors in 269(5229):1427-1429. J Exp Med. 1999;189(9):1399-1412.
a murine model of chronic myelomonocytic leuke- 34. Braun BS, Tuveson DA, Kong N, et al. Somatic 48. Guo W, Lasky JL, Chang CJ, et al. Multi-genetic
mia. Blood. 2010;116(26):5991-6002. activation of oncogenic Kras in hematopoietic events collaboratively contribute to Pten-null leu-
20. Kotecha N, Flores NJ, Irish JM, et al. Single-cell cells initiates a rapidly fatal myeloproliferative dis- kaemia stem-cell formation. Nature. 2008;
profiling identifies aberrant STAT5 activation in order. Proc Natl Acad Sci U S A. 2004;101(2): 453(7194):529-533.
myeloid malignancies with specific clinical and 597-602. 49. Kelly PN, Dakic A, Adams JM, Nutt SL, Strasser A.
biologic correlates. Cancer Cell. 2008;14(4):335- 35. Chan IT, Kutok JL, Williams IR, et al. Conditional Tumor growth need not be driven by rare cancer
343. expression of oncogenic K-ras from its endoge- stem cells. Science. 2007;317(5836):337.You can also read

















































