JBC Papers in Press. Published on March 4, 2020 as Manuscript RA119.011851 The latest version is at ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
JBC Papers in Press. Published on March 4, 2020 as Manuscript RA119.011851
The latest version is at https://www.jbc.org/cgi/doi/10.1074/jbc.RA119.011851
Development of a novel β-1,6-glucan-specific detection system using functionally modified recombinant
endo-β-1,6-glucanase
Daisuke Yamanaka1, 2*, Kazushiro Takatsu1, Masahiro Kimura3, 4, Muthulekha Swamydas2, Hiroaki
Ohnishi5, Takashi Umeyama6, Fumitaka Oyama3, Michail S. Lionakis2 and Naohito Ohno1
From the 1Laboratory for Immunopharmacology of Microbial Products, School of Pharmacy, Tokyo
University of Pharmacy and Life Sciences, Hachioji, Tokyo, Japan; 2Fungal Pathogenesis Section,
Laboratory of Clinical Immunology & Microbiology (LCIM), National Institute of Allergy and Infectious
Diseases (NIAID), National Institutes of Health (NIH), Bethesda, Maryland, USA; 3Department of
Chemistry and Life Science, Kogakuin University, Hachioji, Tokyo, Japan; 4Research Fellow of Japan
Society for the Promotion of Science (DC2), Koujimachi, Chiyoda-ku, Tokyo, Japan; 5Department of
Laboratory Medicine, Kyorin University School of Medicine, Mitaka, Tokyo, Japan; 6Department of
Chemotherapy and Mycoses, National Institute of Infectious Diseases, Shinjuku-ku, Tokyo, Japan
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
Running title: Detection and quantification of β-1,6-glucan
*To whom correspondence should be addressed: Daisuke Yamanaka, Ph.D.; Laboratory for
Immunopharmacology of Microbial Products, School of Pharmacy, Tokyo University of Pharmacy and Life
Sciences, 1432-1 Horinouchi, Hachioji, Tokyo 192-0392, Japan; Tel: +81-426-76-5570; Fax: +81-426-76-
5570; E-mail: ymnkd@toyaku.ac.jp
Keywords: endo-β-1,6-glucanase, β-1,3-D-glucan, β-1,6-glucan, Candida albicans, animal model, deep
mycosis, fungi, glycobiology, glycoside hydrolase, infectious disease
ABSTRACT 1,6-glucan from both yeast and hyphal forms of the
β-1,3-D-Glucan is a ubiquitous glucose opportunistic fungal pathogen Candida albicans,
polymer produced by plants, bacteria, and most without any detectable binding to glucan lacking
fungi. It has been used as a diagnostic tool in the long β-1,6-glucan branch. We developed a
patients with invasive mycoses via a highly- sandwich ELISA-like assay with a low limit of
sensitive reagent consisting of the blood quantification for pustulan (1.5 pg/ml), and
coagulation system of horseshoe crab. However, successfully employed this assay in the
no method is currently available for measuring β- quantification of extracellular β-1,6-glucan
1,6-glucan, another primary β-glucan structure of released by >250 patient-derived strains of
fungal polysaccharides. Herein, we describe the different Candida species (including Candida
development of an economical and highly sensitive auris) in culture supernatant in vitro. We also used
and specific assay for β-1,6-glucan using a this assay to measure β-1,6-glucan in vivo in the
modified recombinant endo-β-1,6-glucanase serum and several organs in a mouse model of
having diminished glucan hydrolase activity. The systemic candidiasis. Our work describes a reliable
purified β-1,6-glucanase derivative bound to the β- method for β-1,6-glucan detection, which may
1,6-glucan pustulan with a KD of 16.4 nM. We prove useful for the diagnosis of invasive fungal
validated the specificity of this β-1,6-glucan probe infections.
by demonstrating its ability to detect cell wall β-
1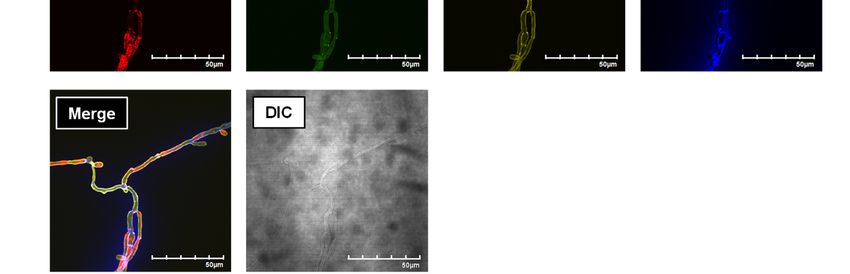
Detection and quantification of β-1,6-glucan
Introduction species early on by blood tests is important for a
β-glucan is constituted by D-glucose units favorable outcome of patients. Interestingly,
linked by β-1,3-glycosidic bonds (β-1,3-D-glucan) previous NMR analysis revealed that soluble
which is the most common β-glucan structure extracellular polysaccharides of C. albicans
produced by plants (β-1,3-/β-1,4-glucan) (1), cultured in the β-glucan-free medium were mainly
bacteria (β-1,3-glucan) (2), fungi (3) and algae (4) composed by α-mannan and β-1,6-glucan,
(β-1,6-/β-1,3-glucan); of interest, β-1,3-D-glucan suggesting that the limulus factor G reactive site
has been at the mainstream of glucan research. In (β-1,3-D-glucan) was a rather minor moiety (17).
addition, β-1,3-D-glucan-specific recognition Although commercially available diagnostic
proteins such as limulus coagulation factor G in reagents targeting mannan have already been
horseshoe crab (5), β-1,3-glucan recognition developed using a rabbit polyclonal antibody
protein (βGRP) in insects (6), and dectin-1 (7) and (CAND-TEC and UNIMEDI Candida) or a rat
immunoglobulin (8,9) in mammals are discovered monoclonal antibody (EBCA-1; PLATELIA
in a wide range of species and applied to β-1,3-D- Candida Ag and PASTREX Candida), there is no
glucan-specific detection systems (10-13). Among available tool for targeting β-1,6-glucan structure
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
these, the most commonly used in the world is a thus far. Therefore, we hypothesized that if a tool
factor G from horseshoe crab, a highly sensitive to quantify β-1,6-glucan was developed, it would
and rapid assay. be useful to compensate for the shortcomings of
A horseshoe crab (Limulus polyphemus and the LAL test and it may provide a potential avenue
Tachypleus tridentatus)-derived limulus for future diagnostic test development in clinical
amebocyte lysate (LAL) test has been evaluated practice.
since 1995 to detect β-1,3-D-glucan that is a marker In the present study, we aimed to develop a
of invasive fungal infections (14), and was new simple and convenient method for detection
approved by the US Food and Drug Administration and quantification of β-1,6-glucan. To establish a
in 2004. However, false-negative or false-positive new tool with high sensitivity at a low cost, certain
results were shown by the LAL test in some cases, conditions were required for probe candidates as
because; (i) not all pathogenic fungi release β-1,3- follows; (i) high affinity and specificity for the β-
D-glucan, and (ii) plant- or bacteria-derived β-1,3- 1,6-glucan structure, (ii) a stable monomeric
D-glucan lead to activation of limulus factor G protein, and (iii) being efficiently produced by
unintentionally (15). Therefore, additional fungal Escherichia coli. Among the different candidates,
diagnostic tests should be performed beyond β-1,3- we focused on the endo-β-1,6-glucanase (EC
D-glucan to accurately diagnose invasive fungal 3.2.1.75), which is classified into glycoside
disease in the clinic. hydrolase (GH) families 5 and 30 in the
One of the most common pathogenic fungal Carbohydrate-Active enZymes database (CAZy;
species, Candida albicans, releases a soluble www.cazy.org). Several enzymes have been
mannoprotein-β-glucan complex that can activate identified, cloned from fungi and bacteria, and
limulus factor G (16). These glucan complexes can further characterized for their structure-specific
be detected by the LAL G test and in some cases responses to β-1,6-glucan. Although natural
have helped clinical decisions to start treatment glycoside hydrolases efficiently degrade
early during fungal infection and to determine polysaccharides, we hypothesized that the
whether to administer antifungal or antibacterial elimination of the hydrolytic activity of β-1,6-
drugs when an infection is suspected. Moreover, glucanase by a point mutation in the catalytic
because first-line antifungal agents are different for domain might still retain its glucan binding activity.
each fungal species, diagnosing pathogenic fungal Since Neurospora-derived endo-β-1,6-glucanase
2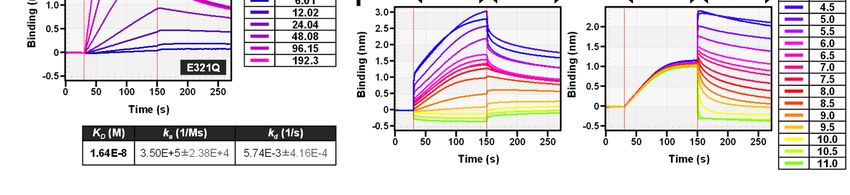
Detection and quantification of β-1,6-glucan
(Neg1), which belongs to the GH family 30 oligosaccharides including a range of glucose
subfamily 3, was well characterized (18) and first tetramers to monomers (Fig. S4A), and also
successfully expressed in E. coli (19), we first increased the amount of reducing sugar in the
attempted to evaluate modified enzymes based on reaction mixture (Fig. S4B), the glucanase
Neg1. The putative catalytic residues for the derivatives Neg1-E225Q, Neg1-E321Q and Neg1-
acid/base and the nucleophile, the common E225Q/E321Q did not increase the reducing sugar
catalytic glutamic acid residues (20-22) of GH (Fig. S4B).
family 30, which were also found in Neg1 (E225 Next, to evaluate whether the glucan
and E321), were mutated to glutamine (Q) to binding activity was preserved in the Neg1 with the
eliminate its hydrolase activity. We further different aforementioned point mutations, we
characterized the glucan binding capacity of this carried out experiments with ELISA and bio-layer
modified form of β-1,6-glucanase. Our results interferometry (BLI). As shown in Fig. 1A, all
demonstrate that the modified recombinant β-1,6- three variants showed binding to plate-coated
glucanase retained its structure-specific glucan pustulan, while they did not bind to immobilized
binding activity and thus, it can be employed as a β-1,3-glucan (laminarin). The Neg1-E321Q and
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
novel β-1,6-glucan-specific detection probe. Neg1-E225Q/E321Q variants in particular
demonstrated greater reactivity even at lower
Results concentrations (0.31–4.88 ng/ml) of solid-phased
Point mutations in the catalytic domain of endo- pustulan. The direct binding between β-1,6-glucan
β-1,6-glucanase promote its glucan-binding and Neg1 derivatives was then examined by BLI
function using a pustulan-conjugated sensor chip. Both
Pustulan is one of the most frequently used Neg1-E225Q and Neg1-E321Q showed stronger
soluble β-1,6-glucan standards. The LAL test did binding activity to pustulan compared to Neg1-
not show strong reactivity towards pustulan and E225Q/E321Q (Fig. 1B). Interestingly, wildtype
other soluble β-1,6-glucan such as mushroom- Neg1 could not retain itself on the polysaccharide
derived AgCAS due to the presence of extremely due to the strong hydrolytic activity of the enzyme.
low content of the β-1,3-glucan moiety in these We calculated the KD value of the binding of Neg1-
forms. Instead, it recognized various β-1,3-glucans E321Q to the pustulan immobilized on the
such as laminarin, single strand SPG biosensor using BLI because this variant showed
(schizophyllan) and pachyman (Fig. S1, S2), strong binding to β-1,6-glucan in both the ELISA
especially, pachyman showed approximately 500 and BLI tests, and the affinity (KD 1.64×10-8 M)
times stronger reactivity than pustulan. (Fig. S2). showed a sufficient value for further investigation
Therefore, to enable specific detection of β-1,6- as a new glucan probe (Fig. 1C). This was further
glucan structures, we aimed to develop specific supported by the results of the isothermal titration
probes by modifying the Neurospora endo-β-1,6- calorimetry (ITC) analysis carried out for reference
glucanase Neg1. We generated plasmids encoding using the free unlabeled pustulan. The affinity was
the mature form of the Neg1 protein with point not very different from the one obtained by BLI
mutations at the catalytic positions (i.e., E225, and we also confirmed that multiple proteins were
E321 or both) to glutamine (Q) (Fig. S3A) and then bound to glucose polymers (binding ratio n = 0.14,
the recombinant Neg1 and its variants were 7.1 proteins:1 ligand).
efficiently expressed in E. coli (Table S3). A single Neg1-E321Q showed thermal stability up
band with expected MW for each of the purified to 40°C, whereas its binding function was
protein was detected (Fig. S3B, C). Whereas Neg1 completely abrogated when treated at 60°C or
strongly hydrolyzed pustulan and produced higher for 5 min (Fig. 1D). Neg1-E321Q exhibited
3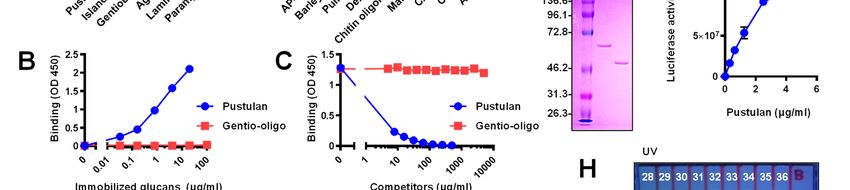
Detection and quantification of β-1,6-glucan
higher performance for ELISA at a neutral pH (pH, Neg1-E321Q to the polysaccharide was then
6 to 7) (Fig. 1E). Interestingly, the absorbance of assessed by competitive ELISA using pustulan-
ELISA was dramatically increased at a pH of 9 or coated plates. Soluble glucans mainly composed
greater (Fig. 1E), therefore, we further analyzed by β-1,6-glucan such as pustulan, islandican and
the effect of pH on the direct interaction of Neg1- AgCAS strongly inhibited the binding between
E321Q with immobilized pustulan using the BLI Neg1-E321Q and solid-phased pustulan (Fig. 2A).
method. During the association phase, strong On the other hand, linear β-1,3-glucan (i.e.,
binding was observed at pH values between 4.5 paramylon) and soluble β-1,3-glucan with β-1,6-
and 5.5, and decreased or absent binding was monoglycoside-branched side chains (i.e.,
confirmed at pH values between 9 and 11 (Fig. 1F, laminarin and SPG) did not interfere with Neg1-
left panel). Acidic conditions (pH values between E321Q binding. Binding of Neg1-E321Q to
4.5 and 5.5) also improved the stability of Neg1- pustulan was also strongly inhibited by β-1,6-/β-
E321Q binding to glucan during the dissociation 1,3-complex glucan (i.e., SCG: Sparassis β-glucan,
phase, and the dissociation rate became faster SCL: scleroglucan, BBG: baker’s yeast-derived β-
depending on the alkalinity of the test buffer (Fig. glucan), and was only slightly inhibited by AP-
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
1F, right panel). FBG (i.e., β-1,3-glucan with β-1,6-glycoside
The long-term stability of glutamine in highly branched side chains) in a concentration-
Neg1-E321Q that was substituted in the catalytic dependent manner. Moreover, extracellular
domain of this enzyme was also quantitatively polysaccharide from C. albicans (CAWS) and cell
evaluated. This was done because, if glutamine wall β-glucan from C. albicans (CSBG) and
reverts to glutamic acid over time, it would no Aspergillus (ASBG) blocked the binding of Neg1-
longer function as a probe. The parental Neg1 E321Q to pustulan. Instead, other glucans such as
exhibited the Km value of 1.1 ± 0.4 mg/ml, for the barley BG (i.e., β-1,3-/β-1,4-glucan), dextran (i.e.,
increase of reducing sugars in natural substrates α-1,4-/α-1,6-glucan), pullulan (i.e., α-1,4-/α-1,6-
similar to a previous report (18). Moreover, the glucan), chitin oligomers, α-mannan (i.e., α-1,6-/α-
glucan hydrolase activity of Neg1-E321Q, which 1,2-, α-1,3-mannan) did not show dose-dependent
was stored for more than two years after inhibition of Neg1-E321Q binding.
purification, did not exhibit any Km value (Fig. We next aimed to understand how many β-
S4C). We also proved that the modified Neg1- 1,6-glucose units are necessary for the binding to
E321Q retained its sugar-binding activity (data not Neg1-E321Q. A previous study showed that
shown) and completely lost its glucan hydrolase another endo-β-1,6-glucanase (i.e., BT3312) from
activity upon long-term storage experiment for Bacteroides thetaiotaomicron was active on
both natural and synthetic substrates (Fig. S4D to gentiotriose (DP 3) but not on gentiobiose (DP 2)
F). Taken together, our data show that endo-β-1,6- (21). Neg1 also produced glucose monomers and
glucanase exerts β-1,6-glucan binding activity dimers by hydrolyzing pustulan (Fig. S4A),
upon loss of its cleavage function via suggesting that it could be active on gentiotriose.
modifications of its catalytic site. This modified As expected, Neg1-E321Q did not bind to
endo-β-1,6-glucanase exhibits stable activity even gentiobiose (Fig. 2A), however, it also did not bind
after long-term storage and thus showed promise to immobilized gentio-oligo mix that contains
for use as a novel β-1,6-glucan probe. glucose dimers to hexamers (Fig. S4A) in the
direct ELISA-like assay (Fig. 2B). The result of the
Structure- and size-dependent binding of Neg1- competitive ELISA also supported the above result
E321Q to β-glucan because even high concentrations of gentio-oligo
The structural specificity of the binding of mix (5 mg/ml) could not inhibit the interaction
4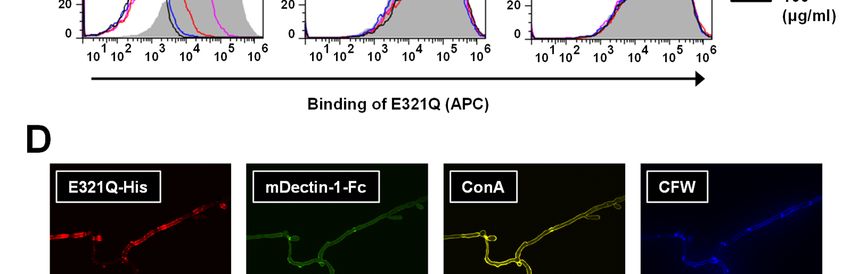
Detection and quantification of β-1,6-glucan
between Neg1-E321Q and solid-phased pustulan inhibited by the addition of soluble β-1,6-glucan,
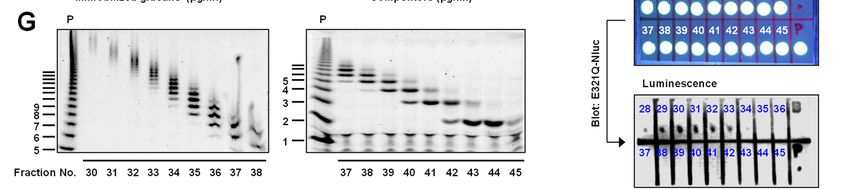
(Fig. 2C). Therefore, we prepared oligosaccharides but not of β-1,3-glucan or mannan (Fig. 3C); this
with larger-molecular weights from pustulan by finding indicates that the binding between Neg1-
hydrolysis with acid and separation by HPLC. E321Q and the yeast cell-surface is mediated in a
Glucose polymers in each fraction were analyzed β-1,6-glucan-specific manner. In addition, Neg1-
by fluorophore-assisted carbohydrate E321Q bound to the hyphal form of C. albicans.
electrophoresis (FACE) after fluorophore-labeling Of interest, the hyphal areas recognized by Neg1-
(Fig. 2G) and used in the dot blot assay. To avoid E321Q were somewhat different from those
an excessive washing process in this assay, we recognized by dectin-1-Fc which stains β-1,3-
further designed Oplophorus luciferase glucan, concanavalin A which stains mannan, and
(NanoLuc)-fused Neg1-E321Q (Neg1-E321Q- calcofluor white which stains chitin (Fig. 3D).Next,
Nluc) (Fig. 2D) and confirmed the ability of to quantify the extracellular polysaccharides
purified Neg1-E321Q-Nluc (Fig. 2E and Table S3) released from C. albicans, we prepared biotin-
to bind β-1,6-glucan by ELISA (Fig. 2F). The labeled Neg1-E321Q and assembled a sandwich
fluorophore-conjugated negative-charge glucose ELISA. By comparison of the horseradish
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
polymer in each fraction was spotted on the peroxidase (HRP) substrate, we applied both
positively-charged nylon membrane (Fig. 2H, colorimetric and chemiluminescent methods using
upper panel), incubated with Neg1-E321Q-Nluc unlabeled Neg1-E321Q-coated microplates and
and the luciferase activity was observed in the biotin-labeled Neg1-E321Q with streptavidin-
acid-degraded pustulan (P) and fraction no. 28–34 HRP and found reactivity to pustulan
(Fig. 2H, lower panel). According to this result, the concentrations ranging from 1.4 to 1,000 pg/ml
minimum unit of β-1,6-glucose polymer that can (Fig. S5A, B). The limit of quantification of the
be recognized by Neg1-E321Q is DP 11–15 (i.e., colorimetric and chemiluminescent methods was
the major bands in fraction no. 34). Collectively, 32.1 and 1.5 pg/ml, respectively (Fig. S5C, D).
these results suggest that Neg1-E321Q has a strong Accordingly, we decided to use the
structural specificity and molecular size chemiluminescent method for our subsequent
dependence for binding to polysaccharides. experiments. The standard curve from a broad
range of pustulan concentrations (i.e., 30.5 pg/ml
Applying the β-1,6-glucanase Neg1-E321Q to the to 22.2 ng/ml) is shown in Fig. 4A. Then, yeast
quantification of Candida β-1,6-glucan colonies of C. albicans strain NBRC1385 were
Because Neg1-E321Q reacted with inoculated in RPMI 1640 medium containing 10%
pathogenic fungal-related polysaccharides, FBS and cultured at 37°C for 24 h for the hyphal
particularly with Candida cell wall (CSBG) and form to develop in order to measure the naturally
extracellular (CAWS) glucan (Fig. 2A), we next produced extracellular polysaccharides by
aimed to employ the β-1,6-glucanase assay for a Candida hyphae (Fig. 4B). The supernatants of the
potential diagnostic application. First, to culture medium with or without Candida were then
demonstrate whether there is direct interaction tested by sandwich ELISA, which showed
between Neg1-E321Q and the cell-surface of C. reactivity to the Candida supernatant, but not to the
albicans, we carried out flow cytometric (for the Candida-free medium, in a dilution-dependent
yeast form) and microscopic (for the hyphal form) manner (Fig. 4C). Furthermore, the diluted
analyses using the C. albicans strain NBRC1385. supernatant was measured by both the β-1,6-
Notably, Neg1-E321Q bound to the cell wall of the glucan ELISA and the β-1,3-D-glucan LAL test.
yeast form of C. albicans in a dose-dependent Notably, the Neg1-E321Q sandwich ELISA test
manner (Fig. 3A, B) and this binding was clearly could measure β-1,6-glucan in both 250-fold and
5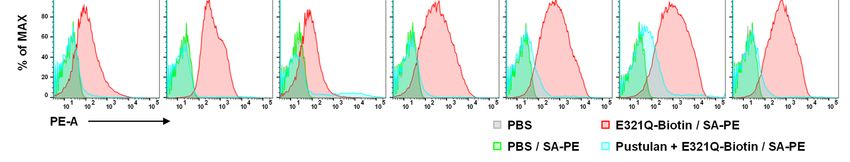
Detection and quantification of β-1,6-glucan
2,000-fold diluted Candida supernatants (Fig. 4D). ELISA could detect naturally produced β-1,6-
While the 250-fold diluted Candida supernatant glucan in vivo in mice infected systemically with
also contained detectable β-1,3-D-glucan, the the C. albicans strain SC5314. First, we confirmed
2,000-fold diluted Candida supernatant only abundant production of β-1,6-glucan following in
contained measurable β-1,6-glucan but not β-1,3- vitro culture of C. albicans strain SC5314 by
D-glucan (Fig. 4E). ELISA (Fig. 5A). Next, we measured the
Since we found that that extracellular concentration of β-1,6-glucan in the serum and
polysaccharides could be detected in the culture homogenized kidney, spleen, liver and brain of WT
supernatant even at a 2,000-fold dilution, that mice at days 3, 6 and 9 post-infection with C.
culture supernatant was then injected albicans SC5314 and found significant increases
intravenously into mice to determine whether these compared to uninfected control mice; the β-1,6-
circulating polysaccharides could be detected in glucan concentration peaked at day 6 after
vivo in the mouse blood. After 1, 10 and 30 min of infection at the peak of fungal proliferation in the
injection, we detected β-1,6-glucan in the serum by model (Fig. 5B to F) (23). We also examined β-1,6-
the sandwich ELISA using Neg1-E321Q. Instead, glucan levels in Candida-infected Cx3cr1-
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
β-1,6-glucan was not detected in blood by 24 h deficient mice that exhibit greater tissue fungal
after administration (Fig. 4F). Importantly, we did burden and mortality relative to WT mice (24); the
not detect a non-specific signal in the serum of β-1,6-glucan content in serum and tissues of
mice injected with the Candida-free medium. Cx3cr1-deficient mice tended to be higher than
In order to exclude the possibility that our that of WT mice (Fig. S6). Together, these data
ELISA might measure metabolic products other show that β-1,6-glucan is produced in vivo in blood
than β-1,6-glucan that could be released by C. and various organs of Candida-infected mice and
albicans, we examined the C. albicans Cabig1Δ its temporal kinetics can be measured by our
strain BIG104 which is known to have impaired β- ELISA using Neg1-E321Q.
1,6-glucan biosynthesis, together with the
reconstituted C. albicans strain BIG105 that has β-1,6-glucan is produced by a large number of
intact β-1,6-glucan biosynthesis. The two strains clinical Candida strains irrespective of species
grew similarly (Fig. 4G) and both produced β-1,3- We have thus far have shown that β-1,6-
glucan as measured by the LAL test (Fig. 4H). glucan can be detected in the culture supernatants
Instead, when we measured β-1,6-glucan in the of three strains of C. albicans (i.e., NBRC1385,
supernatant of both strains by the sandwich ELISA BIG105, SC5314) by a sandwich ELISA using
using Neg1-E321Q, we detected β-1,6-glucan only Neg1-E321Q. Because no information exists with
in the reconstituted C. albicans strain BIG105, but regard to the ability of all C. albicans strains to
not in the β-1,6-glucan-deficient C. albicans produce β-1,6-glucan, we next examined levels of
Cabig1Δ strain BIG104 (Fig. 4I). Together, this β-1,6-glucan (and of β-1,3-D-glucan as control) in
finding indicates that our sandwich ELISA 32 strains of C. albicans obtained from NBRC (n
specifically detects β-1,6-glucan, even within a = 9) and the Kyorin University Hospital (n = 23).
crude biological specimen derived from C. The strains were cultured for 24 h in vitro and
albicans. analyzed using both our ELISA method and the
LAL test. As shown in Fig. 6, both β-1,6-glucan
β-1,6-glucan is detected in the serum and tissue and β-1,3-D-glucan were detected in all tested C.
homogenates of Candida-infected mice by an albicans isolates, and we found a positive
ELISA-like assay correlation between β-1,6-glucan and β-1,3-D-
We next wondered whether our sandwich glucan levels in the tested strains.
6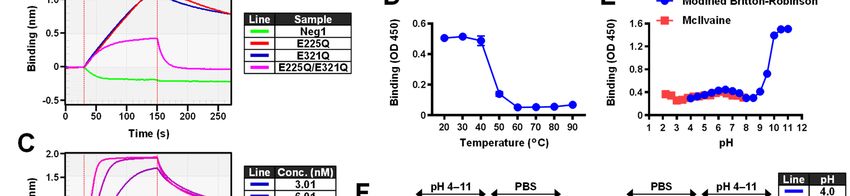
Detection and quantification of β-1,6-glucan
We next expanded our investigation in 224 detect β-1,6-glucan in vivo in the mouse model of
other Candida clinical isolates across all Candida systemic candidiasis provide the foundation for the
species to determine the extent and strain future development and testing of β-1,6-glucan as
specificity of β-1,6-glucan production. For that, a potentially useful diagnostic test in humans with
132 C. albicans and 92 non-albicans Candida invasive candidiasis.
species [C. glabrata (n = 35), C. dubliniensis (n =
15), C. parapsilosis (n = 11), C. krusei (n = 11), C. Discussion
auris (n = 11), C. tropicalis (n = 9)] were cultured In recent years, invasive fungal infections
for 24 h and the level of β-1,6-glucan production such as candidiasis and aspergillosis have emerged
was measured for each strain by our ELISA. as important causes of morbidity and mortality in
Notably, β-1,6-glucan was detected at high levels acutely ill patients in the intensive care unit and in
in the culture supernatants of all tested strains of C. immunosuppressed patients with cancer, and
albicans, C. dubliniensis, C. parapsilosis, C. hematopoietic stem cell or solid organ
tropicalis, and C. auris (Fig. 7A). C. krusei strains transplantation. Mortality in patients affected by
released β-1,6-glucan to a lower extent relative to invasive fungal infections remains unacceptably
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
the aforementioned Candida species. C. glabrata high (>40%) despite administration of potent
produced the least amount of β-1,6-glucan relative antifungal therapy (25,26). A major cause for the
to all other Candida species; yet, the β-1,6-glucan high mortality in these patients is the delayed
measured in C. glabrata supernatants was initiation of antifungal treatment, which is caused
significantly greater compared to that in the by the suboptimal performance of current
Candida-free medium. Since the growth rate of C. diagnostic tests for invasive fungal infections (27).
glabrata can be slower than that of other Candida Specifically, fungal isolation and identification by
species, we extended its incubation period to 72 h culture and/or histopathological examination is
and observed a slight, yet significant, increase in β- hampered by low sensitivity and even when
1,6-glucan production in the culture medium in positive, it typically takes several days to identify
most tested C. glabrata strains (Fig. 7B). the infecting pathogen. PCR testing appears
Our data revealed that all Candida species sensitive but it is not standardized for clinical use
produce β-1,6-glucan but the extent of the (27). The recent advent of serological tests that
production varies in different Candida species. We measure fungal polysaccharides such as β-1,3-D-
wondered whether the amount of β-1,6-glucan glucan and galactomannan has improved
exposed on the cell wall of various Candida might diagnostic accuracy in certain settings but still has
mirror the species-specific β-1,6-glucan limitations (27). Therefore, new diagnostic tests
production. For that, we employed FACS and used are needed to facilitate timely diagnosis of invasive
Neg1-E321Q as the probe to bind to yeast forms of fungal infections and improve patient outcomes.
representative strains from the 7 different Candida Besides its decreased sensitivity and
species (Fig. 7C). This binding was specific as it specificity, another important limitation of the
was inhibited by the addition of pustulan. All tested LAL test that measures β-1,3-D-glucan is a large
strains from the 7 Candida species isolated from decline in horseshoe crab population due to
patients had detectable β-1,6-glucan on their cell commercial harvesting. As such, although the LAL
wall using this approach. Taken together, our data C test kit for measuring endotoxin has been
show that β-1,6-glucan can be produced and reconstructed by animal-free recombinant proteins
detected by our Neg1-E321Q-based ELISA assay (28) (for example, PyroGene rFC/Lonza and
in >250 clinical isolates of various Candida PyroSmart/Seikagaku Corporation), the LAL G
species. This finding together with the ability to test that measures β-1,3-D-glucan is still made
7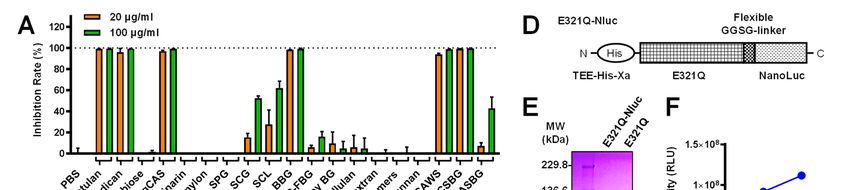
Detection and quantification of β-1,6-glucan
from blue blood collected from living horseshoe ELISA method. The absence of non-specific
crabs. reactivity of our modified β-1,6-glucanase to
In the present study, we developed a β-1,6- Candida-produced exopolysaccharide was proven
glucan detection system with an animal-free by our analysis of C. albicans strain BIG104 which
recombinant protein. Before focusing on β-1,6- lacks β-1,6-glucan biosynthesis. Interestingly, the
glucanase, we had considered other candidates for amount of released β-1,6-glucan varied depending
a β-1,6-glucan probe, such as the Musa acuminata- on the Candida species, but was detected in all
derived lectin (29), yeast-derived K1/K2 killer medically important Candida species, with higher
toxins (30) and a monoclonal antibody (31). levels in C. albicans, C. dubliniensis, C.
However, we excluded these candidates for the parapsilosis and C. tropicalis, intermediate levels
following reasons; the lectin has insufficient in C. krusei and C. auris, and lower levels in C.
structure specificity, killer toxins are structurally glabrata. The presence of β-1,6-glucan in the cell
unstable, and a monoclonal antibody requires high wall of C. glabrata was confirmed by our FACS
cost for sufficient production in high-quality grade. results and is consistent with a previous report (33),
As expected, Neurospora β-1,6-glucanase-derived however, the production level of extracellular β-
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
genetically engineered enzymes were efficiently 1,6-glucan was lower in C. glabrata. Although the
expressed in E. coli (Table S3) without any growth rate may have contributed to this lower
refolding process, and also exhibited expected production of β-1,6-glucan, other factors such as
binding capacity for pustulan and various β-1,6- the greater evolutionary distance on the
glucans. Moreover, they could easily be fused with phylogenetic tree may also be operative (34). To
other small proteins like Oplophorus luciferase clarify how much β-1,6-glucan is contained in the
(NanoLuc; Fig. 2D) and these modified enzymes naturally released exopolysaccharides from major
were also found to exhibit stable glucan binding pathogenic fungi of humans including Aspergillus
activity without regaining glycolytic function even spp., Mucorales spp., Cryptococcus spp., and
after long-term storage. For these reasons, we Pneumocystis spp., further investigation will be
propose that the modified β-1,6-glucanase has a required in the future.
potential application and can be utilized in new In developing a serum diagnostic method,
fields. A future direction of research could focus on it is necessary to consider the biological
further enhancing the design of improved probes metabolism rate of the target molecule. The
based on modifying the glucanase with a higher pharmacokinetic information pertaining to C.
affinity to glucan. albicans cell wall β-glucan has previously been
A potential advantage of detecting β-1,6- reported (35) and the clearance of vascular
glucan during infection is that it does not respond Candida-derived β-glucan was rapid in the rabbit
to β-glucans from plant (β-1,3-/β-1,4-glucan) or (half-life of 1.4 to 1.8 min). Interestingly, anti-β-
bacteria (β-1,3-glucan). In addition, our β-1,6- glucan antibodies have been detected in humans
glucanase derivative did not respond to gentio- (36) and other animals (37), and their presence
oligosaccharides (DP 2–10), a finding that suggests may affect the clearance rate of β-glucan from the
that low molecular weight oligos derived from bloodstream. Our data on the clearance of injected
food additives or botanical glycosides like a crocin β-1,6-glucan from the serum of mice indicates that
in saffron (Crocus sativus) (32) should not produce the polysaccharide could be detected after 30 min
non-specific reactions in the β-1,6-glucan test. of intravenous administration. To further
Furthermore, the use of an immunoassay is facile. understand the pharmacokinetics of naturally
Indeed, β-1,6-glucan released from cultured released β-1,6-glucan and other β-glucans with a
Candida strains could be easily detected by our variety of composition ratios of β-1,3-glucan and
8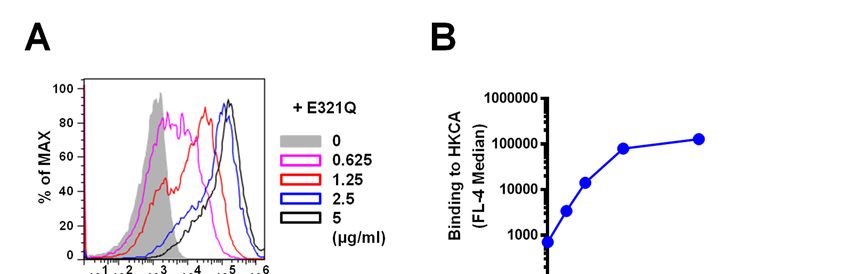
Detection and quantification of β-1,6-glucan
β-1,6-glucan, future research will be required. Laboratory Animal Care-accredited animal facility
Importantly, β-1,6-glucan could be detected with at the NIAID under SPF conditions and housed in
our probe in vivo from serum and several organs in accordance with the procedures outlined in the
the mouse model of systemic candidiasis. These Guide for the Care and Use of Laboratory Animals
preclinical data show promise for the potential under the auspices of a protocol approved by the
development of a β-1,6-glucan-based detection Animal Care and Use Committee of the NIAID
system as a diagnostic modality for future clinical (LCIM14E). Eight to 12-week-old female mice
use. were infected with C. albicans strain SC5314.
In conclusion, we have found that a point Study protocols for Candida yeasts isolated from
mutation at amino acid position 321 (glutamic patients at the Kyorin University Hospital (895,
acid to glutamine) in the endo-β-1,6-glucanase 16-22) and the NIH Clinical Center (11-I-0187)
Neg1 from N. crassa promotes its function as a β- were approved by the Institutional Review Board
1,6-glucan-specific binding protein and provides committees at each study center. The study was
a probe that has the potential for future diagnostic performed in accordance with the Declaration of
development. We are currently developing an Helsinki.
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
immunoassay-based rapid glucan detection
system with glucanase and magnetic beads, Materials
because the LAL test usually gets the results Gentiobiose, dimethylamine borane
within 90 min, while the β-1,6-glucan ELISA (DMAB) and 1,3-diaminopropane dihydrochloride
requires 4 h. In addition, we are in the process of (DAP-2HCl) were purchased from Tokyo
characterizing the structure of the natural form of Chemical Industry Co., Ltd. (Tokyo, Japan). Clear
the exopolysaccharide from various fungi using and white plate for the β-1,6-glucan ELISA were
both the conventional β-1,3-D-glucan test and our purchased from Greiner Bio-one (Frickenhausen,
β-1,6-glucan detection system. Germany). The peroxidase substrate, 3,3',5,5'-
tetramethylbenzidine (TMB) was purchased from
Experimental procedures KPL Inc. (MD, USA). The soluble β-1,6-glucan
Study approval polymer, pustulan from Lasallia pustulata was
For the kinetic analysis of blood obtained from Calbiochem (CA, USA) and
concentration of intravenously injected β-1,6- InvivoGen (CA, USA). Laminarin (4,38), mannan
glucan, female ICR mice were purchased from (39), barley BG (40), DMSO, and calcofluor white
Japan SLC (Shizuoka, Japan), housed in a specific (CFW) were purchased from Sigma-Aldrich (MO,
pathogen-free (SPF) environment, and were used USA). Bovine serum albumin (BSA) was
at 7 to 10 weeks of age. The animal experimental purchased from Sigma-Aldrich and Fisher
protocol was approved by the Committee for Scientific (NJ, USA). Sonifilan (schizophyllan,
Laboratory Animal Experiments at Tokyo SPG) (41) that have been used clinically as
University of Pharmacy and Life Sciences (P18- anticancer β-glucan in Japan were obtained from
34) and experiment was performed in accordance Kaken Pharmaceutical Co., Ltd. (Tokyo, Japan).
with the experiment guidelines provided by the We purchased scleroglucan (SCL) (42) from
Tokyo University of Pharmacy and Life Sciences. CarboMer, Inc. (CA, USA), pullulan (43) from
The mouse model of systemic candidiasis has been Pfanstiehl Laboratories Inc. (IL, USA) and dextran
previously described (23). C57BL/6 WT and T500 (44) from Pharmacia (Uppsala, Sweden).
Cx3cr1-deficient mice were purchased from Aureobasidium pullulans-derived β-glucan, AP-
Taconic Farms (NY, USA) and were maintained at FBG (45,46) was gifted from ADEKA Corporation
an American Association for the Accreditation of (Tokyo, Japan). Baker’s yeast cell wall glucan
9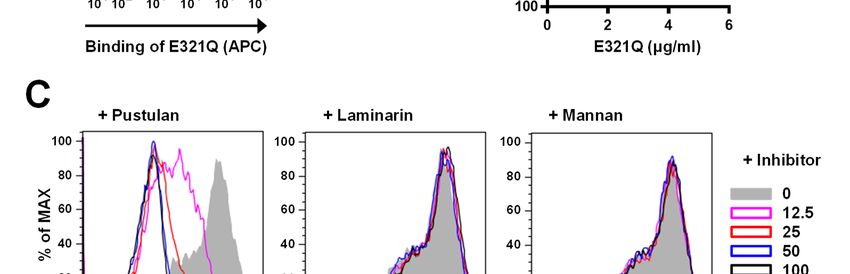
Detection and quantification of β-1,6-glucan
(BBG) (47) was a gift from Oriental Yeast Co., Ltd. released from C. albicans NBRC 1385 (17) were
(Tokyo, Japan). Paramylon (48) and gentio- prepared according to previous reports. Chitin
oligosaccharides (DP 2–6, mix) were purchased oligomers were prepared through acetone
from Wako Pure Chemical Industries, Ltd. (Osaka, precipitation after hydrolysis in concentrated
Japan). Structural characteristics of β-glucan and hydrochloric acid as described previously (56).
non-β-glucan used in this study is listed in Table
S1. Plasmid preparation
The mature form of recombinant endo-β-
Fungal strains 1,6-glucanase (Neg1, GH30_3, EC 3.2.1.75) was
C. albicans NBRC 1385 as a standard prepared as previously reported (19) with slight
strain for in vitro culture, and C. albicans NBRC modifications. The β-1,6-glucanase coding gene
0692, 0759, 1060, 1061, 1393, 1397, 1594, 1974, (neg1) was amplified by PCR using PrimeSTAR
Neurospora crassa NBRC 6068 and Aspergillus Max DNA Polymerase (Takara Bio Inc., Siga,
niger NBRC 6342 were obtained from NITE Japan), primers pCold-IF-NEG1M-F and pCold-
biological resource center (NBRC, Chiba, Japan). IF-NEG1-R with template cDNA prepared from N.
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
The Cabig1Δ strain C. albicans BIG104 that crassa NBRC 6068. The PCR amplicon was
lacking CaBig1p, resulting in repression of β-1,6- purified and cloned into linearized pCold I DNA
glucan biosynthesis and its reconstituted strain C. vector (Takara Bio Inc.) (1–300, 361–4407,
albicans BIG105 were created in the previous amplified with primer sets pColdI-n361-F and
study (49). Two hundred-fourteen Candida yeasts pColdI-n300-R) using In-Fusion HD Cloning Kit
(132 C. albicans; 35 C. glabrata; 15 C. (Clontech Laboratories, Inc., CA, USA), and then
dubliniensis; 11 C. parapsilosis; 11 C. krusei; 9 C. transformed into E. coli DH5α competent cells,
tropicalis; 1 C. auris) and 23 strains of C. albicans cultured in LB broth containing ampicillin (100
isolated from patients at the NIH Clinical Center μg/ml) and purified as Neg1-His6-tag fusion
and Kyorin University Hospital, respectively, were protein-expressing plasmid vector (pCold-Neg1).
tested for quantification of β-1,6-glucan in the The point mutation at the catalytic domain (21) of
culture supernatant in vitro. Ten additional isolates Neg1, Glu225 (acid/base) and/or Glu321
of C. auris (AR Bank#0381 to #0390) were (nucleophile) was induced using basic directional
obtained from the FDA-CDC Antibiotic cloning methods. Linear vector and DNA inserts
Resistance Isolate Bank (GA, USA) and were also for glucanase variants were amplified by PCR
used for in vitro culture test as mentioned above. using primer sets (vector for all variants: NEG1-
For all mouse challenge experiments, C. albicans Mu-F and NEG1-Mu-R, insert for E225Q: NEG1-
SC5314 was used. 225Q-F and NEG1-321E-R, insert for E321Q:
NEG1-225E-F and NEG1-321Q-R, insert for
Preparation of polysaccharide fractions E225Q/E321Q: NEG1-225Q-F and NEG1-321Q-
The β-1,6-glucan islandican from R) with pCold-Neg1 as a template plasmid.
Penicillium islandicum (50), Agaricus Oplophorus gracilirostris-derived low molecular
brasiliensis-derived AgCAS (51) that is rich in β- weight luciferase, NanoLuc (57) (Nluc, 19 kDa)-
1,6-glucan, and the branched β-1,3-glucan SCG fused Neg1-E321Q was designed for the dot blot
from Sparassis crispa (52,53) were prepared as assay. The DNA sequence encoding Nluc and stop
previously described. Solubilized β-glucan codon removed Neg1-E321Q encoding linear
purified from cell wall, CSBG from C. albicans vector was amplified with primer sets (pCold-NL-
NBRC 1385 (54) and ASBG from A. niger NBRC IF-F and pCold-NL-IF-R, pColdI-n361-F and
6342 (55), and the exopolysaccharide CAWS NEG1-FS-R, respectively) and joined by linker
10Detection and quantification of β-1,6-glucan
peptide (GGSGGGSGG) sequence. The protein- by adding the primary amine moiety as previously
expressing plasmid vectors were prepared as described with modifications (58,59). Briefly, 20
described above and DNA sequence was mg of DMAB dissolved in 100 μl acetic acid at
confirmed using BigDye Terminator v3.1 Cycle 80°C was mixed with 5 mg (0.25 μmol) of pustulan
Sequencing Kit (Thermo Fisher Scientific Inc., and heated with 8 ml of DAP-2HCl solution (147
MA, USA) and an ABI3130xl DNA analyzer mg, dissolved in DMSO) at 80°C for 1 h. Water
(Applied Biosystems, CA, USA). All sequences of was added to the reaction mixture, dialyzed
primer sets used in this study are listed in Table S2. (MWCO: 3,500 Da) against water thrice, changed
the buffer to PBS and mixed with biotin-(AC5)2-
Preparation of endo-β-1,6-glucanase and its Osu (0.5 μmol) at RT for 3 h. The reaction mixture
derivatives was further dialyzed against water and then,
SHuffle express competent E. coli cells lyophilized biotin-labeled pustulan was
(New England Biolabs Inc., MA, USA) harboring reconstituted in PBS at 1 mg/ml by boiling.
each plasmid were cultured at 37°C in LB broth
with ampicillin (100 μg/ml) until the OD600 Measurement of (1→3)-β-D-glucan (LAL assay)
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
reached 0.4 and then, isopropyl β-D-1- The concentration of β-1,3-D-glucan was
thiogalactopyranoside was added at final analyzed by the chromogenic kinetic method,
concentration of 0.01 mM, and further incubated Fungitec G Test MKII "Nissui" (Nissui
(180 rpm) at 15°C for 24 h. The cells were Pharmaceutical Co., Ltd., Tokyo, Japan) with
collected and resuspended in PBS containing pachyman (60) as a standard glucan according to
phenylmethylsulphonyl fluoride 0.2 mM and the manufacturer’s instructions. The reaction of
dithiothreitol 1 mM. After sonication repeated NaOH-diluted samples and the LAL reagent in the
thrice for 30 s at 50 watt on ice, the insoluble β-glucan-free 96-well microplates (Toxipet plate
fraction was removed by centrifugation (10,000 96 F, Seikagaku Corporation, Tokyo, Japan) was
rpm, 20 min, 4°C), and the supernatant was applied monitored by Wellreader MP-96 (Seikagaku
to TALON metal affinity resin (Clontech Corporation) at 37°C for 30 min.
Laboratories, Inc.). After washing with sodium
phosphate buffer (pH 7.0), His6-taged Neg1 (52 Verification of modified β-1,6-glucanase as the
kDa) and its derivatives were eluted by structure-specific probe
150 mM imidazole-containing buffer, dialyzed To verify the binding ability and its
against PBS (MWCO: 3,500 Da) and protein structure specificity of β-1,6-glucanase variants to
concentration was measured by Pierce BCA glucans, direct and competitive ELISA-like assays
protein assay kit (Thermo Fisher Scientific Inc.). were carried out. In brief, for direct ELISA,
The yields of recombinant glucanase and its pustulan or laminarin (0–5,000 ng/ml) in 0.1 M
derivatives are summarized in Table S3. sodium carbonate buffer (pH 9.5) were added to a
96-well clear plate and incubated at 4°C. The next
Biotinylation of modified glucanase and pustulan day, the plate was washed by PBS containing
Neg1-E321Q-His (400 μg/ml) was 0.05% Tween 20 (PBST) and blocked with 1%
biotinylated by mixing with a 5-fold molar excess BSA-PBST (BPBST) by incubating for 1 h. Solid-
of biotin-(AC5)2-Osu (Dojindo, Kumamoto, Japan) phased glucans were reacted with recombinant
in PBS at room temperature (RT) for 1 h and then modified Neg1 in BPBST (2 μg/ml) for 1 h,
stored at 4°C until use. For a quantitative washed and the HRP-conjugated anti-His tag
measurement of glucanase affinity, biotinylation of antibody (BioLegend, CA, USA) was added to the
pustulan at the reducing terminus was carried out plate. After 1 h, the plate was washed and the
11Detection and quantification of β-1,6-glucan
binding of modified enzymes to solid-phase baseline was determined by incubating with assay
glucans was monitored using the peroxidase buffer (PBS containing 0.1% BSA and 0.005%
substrate TMB, and color development was Tween 20) in the tube for 30 s. Biotin-pustulan (10
stopped with 1 M phosphoric acid; the optical μg/ml) in 4 μl of assay buffer was loaded to sensor
density was measured at 450 nm using a microplate chips for 120 s and washed by assay buffer for 30
reader (MTP450; Corona Electric, Ibaraki, Japan). s (baseline). For the association assay, sensor chip
For competitive ELISA, various glucans (20, 100 and glucanase variants at 1 μg/ml (19.23 nM) in 4
μg/ml, final concentrations) were mixed with μl of assay buffer were incubated for 120 s. Then,
Neg1-E321Q-His (0.5 μg/ml, final concentration) the assay buffer was applied to the sensor chip for
and pre-incubated for 1 h. The pustulan (0.5 μg/ml) 120 s to collect dissociation data. To calculate the
coated 96-well clear plate was blocked, washed association rate constant (ka), dissociation rate
and incubated with the above mixture of E321Q- constant (kd) and equilibrium dissociation
His and glucan for 1 h. After washing, the binding constants (KD) of E321Q-His, the data from 2-fold
of E321Q-His to the immobilized pustulan was serially diluted five concentrations (3.01, 6.01,
assessed as described above. 12.02, 24.04, 48.08, 96.15, 192.3 nM) of test
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
samples were collected. The data was analyzed
pH and thermal stability of modified β-1,6- using BLItz Pro software (Pall ForteBio Inc.). To
glucanase assess the binding affinity of E321Q-His towards
Stability of Neg1-E321Q was evaluated the unlabeled-soluble pustulan, isothermal titration
by ELISA assay under different conditions. Briefly, calorimetry (ITC) analysis was performed using an
Neg1-E321Q-His (1 μg/ml) in PBS was pre-treated Affinity ITC instrument (TA Instruments, DE,
in varied temperature (20–90°C) for 5 min, cooled USA). E321Q-His and pustulan were co-dialyzed
on ice and an equal volume of BPBST was added. twice against PBS. The average molecular weight
A 96-well clear plate was coated with pustulan (0.5 (Mn: 34,000 Da) of pustulan after dialysis was
μg/ml), blocked by BPBST, incubated with heat- determined by HPLC with similar settings as
treated Neg1-E321Q-His (0.5 μg/ml). To evaluate described in a previous report (47). ITC was
pH-stability, untreated Neg1-E321Q-His (0.5 performed at 25°C with 20 subsequent injections
μg/ml) in 1% BSA-containing various pH of 2.5 μl each. The data were analyzed using
environments with McIlvaine (pH 2.2–7.8) or NanoAnalyze software (TA Instruments, DE,
modified Britton-Robinson (pH 4.0–11) buffer USA) as an independent model.
solution (61,62) were added to each well. Plate
remaining Neg1-E321Q-His was monitored by β-1,6-glucan ELISA for gentio-oligosaccharides
anti-His-tag-mAb-HRP (0.5 mg/ml, BioLegend) A 96-well clear plate was coated with
and TMB. gentio-oligosaccharides (0–100 μg/ml) or pustulan
(0–20 μg/ml), blocked by BPBST, incubated with
Measurement of the affinities of β-1,6-glucanase biotinylated-E321Q (2 μg/ml) and detected using
derivatives streptavidin-HRP (BioLegend) and TMB. The
The binding affinity and the kinetics of reverse experiment was carried out to exclude the
modified Neg1 to the immobilized pustulan were possibility that differential binding was due to the
monitored using the bio-layer interferometry (BLI) size of glucans in the plate. The pustulan (1 μg/ml)
biosensor (BLItz system; Pall ForteBio Inc., CA, coated 96-well clear plate was blocked, treated
USA). The affinities of Neg1 variants were with mixture of biotinylated-E321Q (0.5 μg/ml,
measured as follows: The streptavidin-coated final concentration) and various concentration of
biosensor chips were pre-hydrated and the initial gentio-oligosaccharides (0–5,000 μg/ml, final
12Detection and quantification of β-1,6-glucan
concentrations) or pustulan (0–500 μg/ml, final with Neg1-E321Q-Nluc (2 μg/ml) in blocking
concentrations). The biotinylated-E321Q binding buffer. After washing, membrane remained E321Q
to solid phased-pustulan was detected as described was detected using Nano-Glo luciferase substrate
above. (Promega, WI, USA) in the chemiluminescent
substrate (ImmunoStar; Wako Pure Chemical
Fluorophore-assisted carbohydrate Industries, Ltd.). The images were scanned using a
electrophoresis (FACE) C-DiGit Blot Scanner (LI-COR Biotechnology).
The reducing end of oligosaccharide
sample was labeled by fluorophore 8- Binding of modified β-1,6-glucanase to C.
aminonaphthalene-1,3,6-trisulphonic acid (ANTS; albicans cell wall
Invitrogen, CA, USA) and separated by PAGE Heat-killed C. albicans (HKCA) strain
(63,64). NBRC1385 grown in yeast extract, peptone, and
dextrose (YPD) medium (48 h, yeast form) was
Separation of hydrolyzed-pustulan using HPLC repeatedly washed by PBS. Insoluble fungal body
Pustulan (1 mg/ml) was hydrolyzed by (300 μg/ml, final concentration) was mixed with
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
boiling for 20 min with hydrochloric acid (0.1 N). Neg1-E321Q-His (0–5 μg/ml, final
Gel permeation chromatography was performed to concentrations) in 50 μl of flow cytometry staining
separate hydrolyzed pustulan on an HPLC system buffer for 30 min. A competitive assay was used to
which consisted of a Waters 510 pump (Waters, prove the glucan-specific reaction, in which
MA, USA), a CTO-6A column oven (Shimadzu, E321Q-His (5 μg/ml, final concentration) was pre-
Kyoto, Japan) and a Shodex GS-220 HQ column mixed with different concentration of pustulan,
(7.5 mm × 300 mm, Showa Denko, Tokyo, Japan). laminarin or mannan (0–100 μg/ml, final
The separation was performed at 60°C using H2O concentrations). Subsequently, cell wall binding
as the eluent at a flow rate of 0.5 ml/min (56). E321Q-His was detected using anti-His-tag mAb-
Samples were fractionated every minute and Biotin (MBL Co., Ltd., Aichi, Japan) and
lyophilized by Centrifugal concentrator CC-105 streptavidin-APC (BioLegend). FACS was
(Tomy seiko Co., Ltd., Tokyo, Japan) and performed using a BD Accuri C6 flow cytometer
dissolved in H2O to adjust the concentration. with BD CSampler Software (BD Biosciences, CA,
Oligosaccharide was labeled by ANTS and USA), and data were analyzed using FlowJo
analyzed by FACE as described above. software (Tree Star Inc., OR, USA). Formalin-
killed C. albicans NBRC1385 grown in RPMI
Dot blot assay 1640 medium (Life Technologies Inc., CA, USA)
To disclose the interaction between containing 10% heat-inactivated fetal bovine
modified β-1,6-glucanase and trace amounts of serum (FBS) (Equitech-Bio, TX, USA) (24 h,
oligosaccharide, ANTS-labeled acid-degraded hyphae form) was repeatedly washed by PBS,
pustulan in each fraction separated by HPLC was resuspended in 2% BSA-PBS and incubated with
spotted (1.5 μl) onto a nylon membrane (0.45 μm, Neg1-E321Q-His (5 μg/ml) and dectin-1-Fc
positively charged, Wako Pure Chemical (purified fusion protein of mouse dectin-1
Industries, Ltd.) with a border drawn by carbohydrate-recognition domain and human
WesternSure Pen (LI-COR Biotechnology, NE, IgG1-Fc domain expressed in silkworm, 2.5
USA) in advance. The membrane was gently μg/ml) as the β-1,3-glucan probe. After washing, C.
washed by ultra-pure water containing 0.05% albicans was further treated with anti-His-tag
Tween 20, blocked with 1% casein sodium in mAb-Biotin. To detect cell wall mannan, 2 mg of
washing buffer for 60 min, washed and incubated concanavalin A (Con A) (Wako Pure Chemical
13Detection and quantification of β-1,6-glucan
Industries, Ltd.) was labeled by 100 μg of NHS- μg/ml) or Neg1-E321Q-His plus pustulan (100
Rhodamine (Thermo Fisher Scientific Inc.) in PBS μg/ml). After washing, cells were further incubated
at 4°C (overnight) and Tris buffer was added to in buffer only or PE-conjugated streptavidin (0.4
quench the reaction. Then, cells were washed and μg/ml, Miltenyi biotec, Bergisch Gladbach,
stained with streptavidin-APC (BioLegend), anti- Germany), and then analyzed by cytometer (BD
human IgG1-Fc-FITC (BioLegend), Con A- Fortessa and Diva software; BD Biosciences) and
Rhodamine (10 μg/ml) and 5 μg/ml of CFW for FlowJo software. Yeasts suspended in PBS were
total chitin at 4°C. The unbound reagents were counted and further cultured in 10% FBS
removed by three PBS washes and stained cells containing RPMI 1640 medium (105 yeasts/ml) for
were mounted onto microscope slides with 24–72 h at 37°C. After centrifugation at 1,800 (×g)
ProLong Diamond Antifade Mountant (Thermo for 10 min at 4°C, fungal free supernatants were
Fisher Scientific Inc.). The image data was collected and kept frozen at -20°C until used for β-
collected using a confocal laser scanning glucan test. The remaining fungal body was fixed
microscope (Olympus FV1000; Olympus, Tokyo, in formalin, photographed with EVOS FL Cell
Japan). Imaging System (Thermo Fisher Scientific Inc.) to
Downloaded from http://www.jbc.org/ by guest on December 31, 2020
prove the proliferation. The culture supernatant
Quantification of β-1,6-glucan by a Sandwich was boiled for 5 min before use for β-glucan test.
ELISA-like assay
A 96-well white plate was coated with Murine model of systemic candidiasis
Neg1-E321Q-His (2 μg/ml) by overnight Mouse experiments were performed as
incubation at 4°C. The plate was washed with previously described (24). Briefly, the C. albicans
PBST and incubated for 1 h with BPBST. After strain SC5314 was grown in YPD medium
washing, diluted specimen and standard β-1,6- containing penicillin and streptomycin (Mediatech
glucan (pustulan, InvivoGen) were added to the Inc., VA, USA) in a shaking incubator at 30°C.
plate and incubated for 1 h at RT. Biotinized- Cells were centrifuged, washed in PBS, counted,
Neg1-E321Q-His (2 μg/ml) in BPBST was added and injected (105 yeast cells) into C57BL/6 mice
to the washed plate and incubated for 1 h. The plate via the lateral tail vein. The serum, kidney, spleen,
was then washed and treated with streptavidin- liver, and brain were harvested before infection and
HRP (BioLegend or R&D Systems, MN, USA) in at days 3, 6, and 9 after infection and the organs
BPBST for 20 min. After removing the unbound were homogenized using a tissue homogenizer
enzyme, the peroxidase substrate (SuperSignal (Omni International, Inc., GA, USA) into 1.5 ml of
ELISA femto substrate; Thermo Fisher Scientific PBS with 0.5% Tween20 and a protease inhibitor
Inc.) was added and luminescence signals were cocktail (Roche Applied Science, Upper Bavaria,
measured using a microplate reader (GloMax; Germany) and centrifuged at 15,682 (×g) for 10
Promega or Spark; TECAN, Männedorf, min at 4°C. The clarified supernatants and serum
Switzerland). were frozen at -80°C until use.
In vitro culture of clinical Candida strains Pretreatment of specimen for quantification of β-
Yeasts were inoculated onto YPD agar 1,6-glucan
plates, cultured at RT for few days, and then yeast The appropriate volume of PBS (equal
colonies were suspended in formalin or sterile PBS. volume for serum, 4 times volume for organs) was
Representative clinical isolated yeasts (105 cells) in added to the serum and supernatants of organs
formalin were washed twice with FACS buffer, from Candida-infected mice, and were boiled for 5
incubated with buffer only, Neg1-E321Q-His (8 min, mixed and centrifuged at 14,000 rpm for 10
14You can also read

















































