Marine natural products targeting the eukaryotic cell membrane
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The Journal of Antibiotics (2021) 74:769–785
https://doi.org/10.1038/s41429-021-00468-5
REVIEW ARTICLE
Marine natural products targeting the eukaryotic cell membrane
1
Shinichi Nishimura
Received: 30 March 2021 / Revised: 16 June 2021 / Accepted: 1 July 2021 / Published online: 7 September 2021
© The Author(s), under exclusive licence to the Japan Antibiotics Research Association 2021
Abstract
The cell membrane, with high fluidity and alternative curvatures, maintains the robust integrity to distinguish inner and outer
space of cells or organelles. Lipids are the main components of the cell membrane, but their functions are largely unknown.
Even the visualization of lipids is not straightforward since modification of lipids often hampers its correct physical
properties. Many natural products target cell membranes, some of which are used as pharmaceuticals and/or research tools.
They show specific recognition on lipids, and thus exhibit desired pharmacological effects and unique biological phenotypes.
This review is a catalog of marine natural products that target eukaryotic cell membranes. Chemical structures, biological
activities, and molecular mechanisms are summarized. I hope that this review will be helpful for readers to notice the
1234567890();,:
1234567890();,:
potential of marine natural products in the exploration of the function of lipids and the druggability of eukaryotic cell
membranes.
Introduction often show pharmacological activities, e.g., antimicrobial,
antitumor, antidiabetic, and anti-inflammatory activities, as
Chemically and biologically innovative natural products well as the modulation of the immune and the nervous
have been reported from marine-derived organisms. Marine system [4]. Many marine-derived compounds are in the
invertebrates such as sponges and tunicates are especially preclinical or the early stage of clinical development, and
important as sources from which pharmaceutical leads have some of them have been put into clinical use [3, 5]. Potent
been discovered, and the number of new compounds biological activities are exerted by binding to cellular tar-
reported from them has been constant over the last few gets with high specificity, although modes of action of most
years [1]. Recently, marine-derived fungi are showing its marine natural products remain to be unveiled.
presence in expanding the chemical diversity of marine The cell membrane is one of the major targets for anti-
natural products [1]. Retrospective analyses suggested that microbial natural products [6]. Daptomycin, produced by a
the study of a new source initially yields compounds with soil bacterium Streptomyces roseosporus, is one of the latest
little similarity to existing ones [2]. Unexplored resources, agents to be clinically approved and is used to treat severe
such as symbionts in marine invertebrates, would further infections by Gram-positive bacteria [7]. This compound
expand the chemical space [3]. Marine natural products permeabilizes bacterial membranes, although its molecular
mode of action remains incompletely understood [8].
Amphotericin B, produced by streptomycetes, is a historic
and still-used antifungal drug that targets ergosterol in the
Note: Shinichi Nishimura was awarded the Sumiki-Umezawa fungal membane [9–11]. Other polyene antifungal agents,
Memorial Award from the Japan Antibiotics Research Association in
such as nystatin and pimaricin, also target ergosterol but
2020. This review article is partly based on his award-winning
research. with slightly different sterol specificities and molecular
mechanisms [12]. However, there is an urgent need for new
Supplementary information The online version contains
agents against microbial infections because of the increased
supplementary material available at https://doi.org/10.1038/s41429-
021-00468-5. incidence of drug resistance [13].
Lipids have a high structural diversity: the theoretical
* Shinichi Nishimura number of lipids was estimated to be around 180,000 [14].
anshin@g.ecc.u-tokyo.ac.jp
In the budding yeast Saccharomyces cerevisiae, one of the
1
Department of Biotechnology, Collaborative Research Institute for eukaryotic model organisms, the size of the lipidome was
Innovative Microbiology, The University of Tokyo, Tokyo, Japan reported to be around 300 [15]. Hundreds to thousands of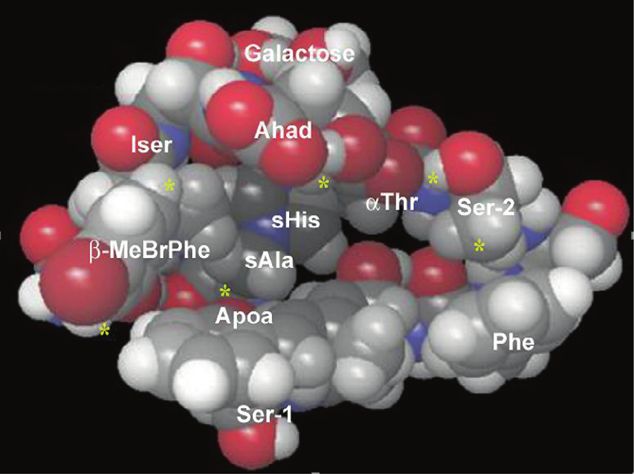
770 S. Nishimura
lipid species are distributed in the body, organs, and cells in cyanobacteria include bastimolide A and palstimolide A,
specific manners. Since lipids are not directly encoded in both with a 40-membered ring (Fig. 1) [22, 23], caylobo-
the genome, genetic approaches are not enough to under- lides with a 36-membered ring [24, 25], and palstimolide B
stand their functions and to visualize its cellular distribu- with a 24-membered ring [26]. Palstimolide A showed
tions. Chemical genetics using molecules that directly bind antiparasitic activity, bastimolides exhibited potent anti-
to lipids is an alternative to investigate the function of the malarial activity, and caylobolides showed moderate cyto-
membrane lipids [16]. New technology reconnects old toxic activity against mammalian cells. Amantelide A
compounds with membrane lipids, while new membrane- showed broad-spectrum activity against mammalian cells,
targeting molecules are being discovered in the course of bacterial pathogens and marine fungi. It is noted that the
screening for novel bioactive natural products. Furthermore, acetylation of one of the hydroxy groups gives a biologi-
our understanding of molecular mechanisms of interactions cally less potent congener amantelide B (Fig. 1) [21].
between natural products and membrane lipids have pro- Recently, amantelide A was shown to exert the anti-
gressed with the aid of newly developed analytical methods fungal action via binding to ergosterol-containing mem-
[17]. branes followed by pore formation, which is partially shared
Herein, I would like to review marine natural products with polyene antifungals such as nystatin [27]. In vitro
that target eukaryotic cell membrane. Their chemical binding assays were examined by surface plasmon reso-
structures, biological activities, and molecular mechanisms nance (SPR) experiments. In this assay, 1-palmitoyl-2-
are described. This review is partly the update of the pre- oleoyl-sn-glycero-3-phosphatidylcholine (POPC)-based
vious review, focusing on marine natural products [16]. liposomes were captured on the sensor chip and then
Readers interested in helix forming peptides, ribosomal compounds were eluted as analyte. Amantelide A showed
peptides, compounds targeting bacterial cell membrane, significant binding to POPC liposomes, which was mark-
which are not included in this review, should refer to pre- edly increased by the inclusion of 20 mol% ergosterol in the
vious reviews [6, 18–20]. liposome (Fig. 2). The increased affinity was also observed
when membranes contained cholesterol, suggesting that the
Amantelide A from marine cyanobacteria cytotoxicity to mammalian cells might be due to its affinity
to cholesterol-containing membranes. Furthermore, addition
Amantelides are polyhydroxylated macrolides isolated from of epicholesterol (3α-cholesterol, Fig. 2) also enhanced the
marine cyanobacteria [21]. Amantelide A is a 40-membered binding of amantelide A to the membrane. These results
macrolide consisting of a 1,3-diol and contiguous 1,5-diol suggest that amantelide A may have a unique mode of
units (Fig. 1). Other polyol metabolites derived from membrane binding, since sterol-targeting compounds
Fig. 1 Chemical structures of a
membrane-binding polyhydroxy
metabolites. a 40-Membered
polyhydroxy macrolactams from
marine cyanobacteria. b
Amphidinol 3 from marine
dinoflagellates
amantelide A (R = H)
amantelide B (R = Ac)
bastimolide A palstimolide A
b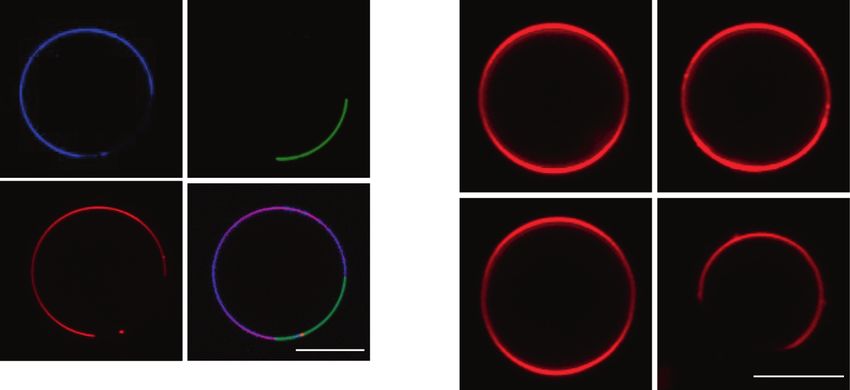
Marine natural products targeting the eukaryotic cell membrane 771
Fig. 2 Chemical structures of
sterols
usually require the β-configuration of the 3-OH group in stereochemistry of the 3-OH group of sterols in the mem-
sterols (see below) [28, 29]. After binding to the membrane, brane. Channel-current recording experiments were con-
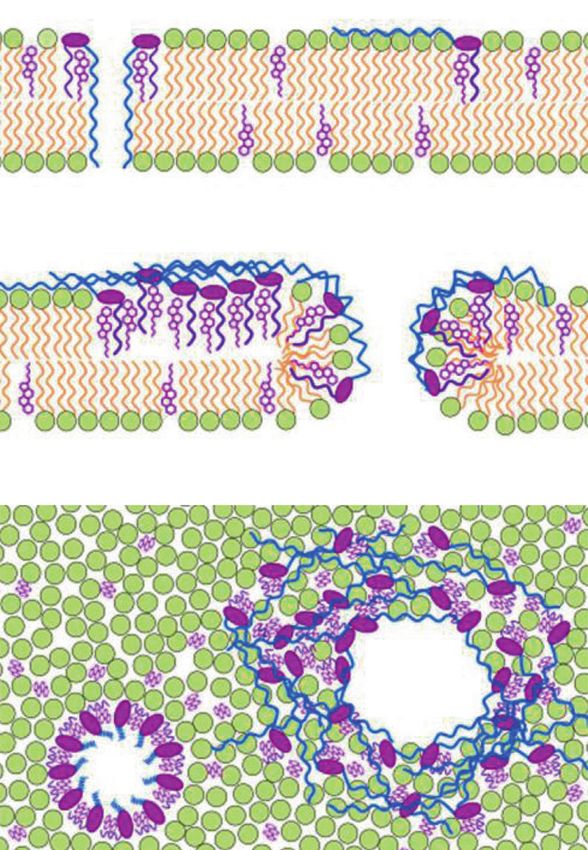
amantelide A seems to form a pore or a lesion with a dia- ducted to show the significantly higher activity of AM3 in
meter of 2.0‒3.8 nm. The size of the pore/lesion was similar an ergosterol-containing membrane [54]. The polyene por-
to that of amphidinol 3 (see next section), whose pore/lesion tion of AM3 was suggested to recognize the sterol hydro-
size was estimated to be 2.0‒2.9 nm [30]. The hydroxy phobic region to some extent (Fig. 3a) [29].
group at C-33 should be involved in the unique membrane- The pore made by AM3 was expected to be a barrel-
binding mode, since the C-33 acetoxy derivative amantelide stave or toroidal type (Fig. 3b). A toroidal pore model, in
B lost sterol-dependent membrane affinity and biological which the lipid monolayer bends continuously from the
activities. Drastic difference between activities of amante- outer to the inner leaflets of the membrane, was assumed
lide A and B predicts a highly specific, unique binding since the membrane thickness had little effect on the
mode of amantelide A with lipid membranes. membrane permeabilizing activity of AM3 [53, 55, 56].
However, solid-state 31P NMR experiments did not support
Amphidinol 3 from marine dinoflagellates this model: distinctive isotropic signals did not appear in the
presence of AM3, suggesting that AM3 binds to the
Amphidinols (AMs) are polyhydroxy polyene metabolites membrane without substantially changing the orientation of
isolated from dinoflagellates Amphidinium spp. They have the surrounding lipids [29]. This result supported the
two tetrahydropyran rings that are linked by a carbon chain, transmembrane barrel-stave pore formation by AM3
to which a polyene chain is attached. So far, more than 20 (Fig. 3b). In the barrel-stave model, AM3 takes a hairpin-
amphidinols and many congeners have been reported from like conformation with a relatively rigid turn structure at the
dinoflagellates [31–49]. Among them, amphidinol 3 (AM3) middle region between the two tetrahydropyran rings
has the most potent hemolytic and antifungal activities. The (Fig. 3a) [55, 56]. The turned region may recognize the 3β-
stereochemistry of AM3 was first reported in 1999 [50], but OH group of sterols through hydrogen bonds [29]. The
was revised several times. Final revision reported in polyol chain penetrates the membrane to form the hydro-
2017 settled on the definite structure shown in Fig. 1, and philic inner pore, while the hydrophobic polyene structure
the first total synthesis was accomplished in 2020 [51, 52]. likely interacts with the steroidal skeleton. Recent channel-
AM3 forms pores in lipid membranes in a sterol- recording experiments showed that AM3 forms both barrel-
dependent manner [53], which was shown by observing stave type transmembrane channels and toroidal-like pores
leakage of calcein dye from liposomes [29]. Strong dye (jumbo pores) in a concentration dependent manner; a
leakage activity was observed in ergosterol- or cholesterol- barrel-stave pore is formed at a lower concentration of
containing liposomes, but not in ones containing epicho- AM3, while a jumbo pore forms at a higher concentration
lesterol (Fig. 2). Physical studies employing the SPR (Fig. 3b) [54]. In the jumbo pore model, the AM3 polyol
experiments and solid-state 2H NMR measurements indi- chain may exist on the membrane surface to increase the
cated that AM3 directly interacts with cholesterol and local membrane curvature and facilitate the pore formation
ergosterol in membranes, but has much weaker interaction [54]. Recently, AM3 was shown to preferentially bind to
with epicholesterol [29]. AM3 strictly recognizes the cholesterol in liquid-disordered phase, than that in liquid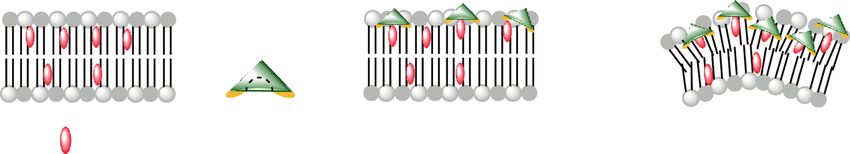
772 S. Nishimura
Fig. 3 Membrane interaction a b
model of amphidinol 3 (AM3). a
Binding model of AM3 and
cholesterol. The turned region of
AM3 likely recognizes the 3β-
OH group of cholesterol. b
Binding model of AM3 in the
membrane. The hydrophobic
polyene chain (purple) interacts
with sterols, and the hydrophilic
polyol chain (blue) penetrates
the membrane or resides on the
membrane surface (top and
middle). When the concentration
of amphidinol 3 is low, barrel-
stave pores are formed (top). At
high concentrations of the
compound, jumbo pores are
generated (middle). Top view of
the barrel-stave (left) and jumbo
pore (right) are shown (bottom).
Figures in (b) are reproduced
from ref. [54]
Fig. 4 Chemical structures of
heronamides.
BE-14106 / GT32-A: R = OH heronamide C: R = OH
GT32-B: R = H 8-deoxyheronamide C: R = H
heronamide F
8R,9S-heronamide C
heronamide A heronamide B
ordered phase. This might be due to the weaker lipid- Heronamides from marine actinomycetes
cholesterol interaction in disordered membrane than that in
ordered membrane. As a consequence, AM3 enhances the Heronamides are 20-membered polyene macrolactams,
order of the disordered phase and modulates phase separa- including BE-14106/GT32-A [58, 59] and the 8-deoxy
tion [57]. Both of the pore formation and the modulated congener GT32-B [60], ML409 [61], heronamide C [62] and
membrane order could affect the function of the cell F [63], and 8-deoxyheronamide C [64], all of which were
membrane. reported from Streptomyces species (Fig. 4). HeronamidesMarine natural products targeting the eukaryotic cell membrane 773
are relatively unstable and intramolecular cyclization compounds, thus heronamides were expected to target the
undergoes to furnish tri- or tetracyclic compounds, such as cell membrane [72, 73].
heronamides A and B (Fig. 4) [62, 63, 65–70]. 1H NMR Membrane affinity of heronamides was analyzed by SPR
spectra of heronamide C and 8-deoxyheronamide C gave experiments as described above [64]. Heronamides bound
heavily overlapping signals for olefinic protons, which irreversibly to liposomes consisting of 1,2-dimyristoyl-sn-
hampered correct assignment of the stereochemistry [62]. In glycero-3-phosphocholine (DMPC) or sphingomyelin
contrast, tri- or tetracyclic compounds derived from the (SM), both of which have saturated hydrocarbon chains
monocyclic compounds showed relatively dispersed olefinic (Fig. 5a). In contrast, heronamides showed weak binding to
protons, for example, heronamide A from heronamide C. liposomes consisting of 1,2-dioleoyl-sn-glycero-3-phos-
Taking advantage of this conversion, the stereochemistries phocholine (DOPC) or 1-palmitoyl-2-oleoyl-sn-glycero-3-
of heronamide C and BE-14106 were deduced [59, 64]. phosphocholine (POPC). DOPC and POPC have unsatu-
Total synthesis of heronamide C was reported, which rated acyl chains. Presence of 20 mol% of ergosterol in the
unambiguously determined the chemical structure [64, 66]. liposome did not affect the membrane affinity of her-
BE-14106 showed antibacterial and antifungal activities onamide C and 8-deoxyheronamide C. In addition, her-
[58, 60, 71]. BE-14106 showed cytotoxicity to murine onamides may not distinguish the membrane fluidity. SPR
tumor cell lines and human lymphoma cells, while it did not experiments were conducted at 30 °C. At this temperature,
show acute toxicity in mice after 100 mg kg−1 was intra- DMPC membranes display a liquid-disordered phase
peritoneally administered. Heronamide C showed a rever- whereas DMPC membranes containing cholesterol have a
sible non-cytotoxic effect on mammalian cell morphology mixture of liquid-disordered and ordered phases. SM
[62]. 8-Deoxyheronamide C was discovered in a screen for membranes are in a gel phase. Thus, heronamides likely
membrane-targeting antifungal compounds. To obtain new favor lipid molecules possessing saturated hydrocarbon
membrane-targeting molecules, microbial culture extracts chains.
were screened to identify a marine-derived actinomycete The molecular size and the rigid structure of her-
Streptomyces sp., whose culture extract showed less onamides suggest that they can act as pseudo-sterols
toxicity to the fission yeast cells lacking erg2 gene than to (Fig. 5b). Cholesterol are preferentially localized in mem-
wild-type cells. Bioassay-guided fractionation afforded branes consisting of lipids that have saturated hydrocarbon
8-deoxyheronamide C. Cells lacking ergosterol biosynthetic chains when compared with membranes that contain lipids
genes such as erg2 are tolerant to membrane-binding having unsaturated chains [74–77]. In the case of
a b
SM DMPC POPC DOPC
c d
0.1 1 10 100
anti-yeast activity (MIC, μM)
Fig. 5 Binding of heronamides to lipid membranes. a Chemical yeast assay using wild-type cells are shown [59, 64, 66]. Asterisk (*)
structures of phospholipids used in the binding test using SPR [64]. indicates these two derivatives were not active at 40 μM. d
SM and DMPC membranes showed high affinity to heronamides, Structure–activity relationships of the heronamides. Details are
while POPC and DOPC membranes did not. b A schematic for the described in the main text. b Reproduced from ref. [16] by permission
molecular interactions between cholesterol, heronamides, and phos- of The Royal Society of Chemistry
pholipids. c Anti-yeast activities of heronamides. MIC values in anti-774 S. Nishimura
heronamides, the hydrophobic ring and tail are inserted into this phenotype. Bgs1 is a 1,3-β-glucan synthase [79], while
the lipid membranes, whereas hydroxy groups are located at Rho1 is an small GTPase that activates 1,3-β-glucan syn-
the hydrophilic, shallow area of the membranes. Hydroxy thase [80]. Notably, a similar phenotype was reported in
groups can form hydrogen bonding with lipid head groups, sphingolipid metabolism mutant cells. The temperature-
which stabilizes the lateral interaction between heronamides sensitive mutant of css1, encoding sphingolipid-
and phospholipids. Structure–activity relationships of her- phospholipase C, displayed accumulation of cell wall
onamide support this pseudo-cholesterol model (Fig. 5c, d). material at the restrictive temperature [81]. Curiously, this
Heronamide C showed anti-yeast activity, which is as phenomenon was also induced by theonellamides (TNMs),
potent as amphotericin B, and 20–40 times more potent than whose cellular target is ergosterol (see below) [82]. These
8-deoxyheronamide C (Fig. 5c). When hydroxy groups in data suggest that the cell wall biosynthesis machinery is
heronamide C were acetylated, both membrane affinity and regulated by the so-called lipid raft, a membrane micro-
anti-yeast activity was lost. The intact macrolactam ring was domain consisting of sphingolipids with saturated hydro-
requisite since heronamide A and heronamide B lost carbon chains, sterols, and proteins [83]. In fact, Bgs1 was
activities [64]. 8R,9S-heronamide C, a synthetic congener, shown to be insoluble in non-ionic detergents [84], which is
showed 80 times less potent activity than heronamide C one characteristic of lipid-raft proteins. Molecular basis for
[66, 78]. This synthetic congener showed opposite Cotton this unique morphological change by heronamides and
effects in the CD spectrum when compared with natural TNMs is under investigation in our group.
heronamide C, indicating that the conformation of the
macrolactam ring is important to embed the compound in Papuamides from a marine sponge
the membrane. Otherwise, the hydroxy groups in 8R,9S-
heronamide C may not have been placed in a proper Papuamides are marine sponge-derived cyclic depsipeptides
direction for exhibiting high membrane affinity. The struc- (Fig. 6). Papuamides A-D were isolated from Papua New
ture of the tail also affects the antifungal activity. BE- Guinea collections of the sponges Theonella mirabilis and
14107, with a shorter hydrocarbon tail by one acetate unit, Theonella swinhoei [85], and papuamides C-F were from
showed four times less potent activity than heronamide C. Melophlus sp. collected in the Solomon Islands [86].
The hydrophobic interactions expected between her- Chlorinated congeners, mirabamides A-H, some of which
onamides and the phospholipids might be easily changed by are glycosylated, were also reported from sponges (Fig. 6a)
the structure/length of the tail. [87, 88]. The depsipeptide ring of papuamides and mir-
In fission yeast, heronamides induced drastic morpholo- abamides consists of seven amino acids, to which a linear
gical changes: cell wall material abnormally accumulated at tetrapeptide is attached. The N-terminus of the tetrapeptide
both cell tips and septa. This material might be 1,3-β-glucan chain in papuamides A-D and mirabamides A-E and G was
because genetic perturbation of Bgs1 or Rho1 suppressed capped by a polyketide chain, 2,3-dihydroxy-2,6,8-
a
papuamide B mirabamide A
b
Cho1
serine + DAG-CDP
Fig. 6 Chemical structures of papuamides and biosynthesis of phos- 2S, 3S configuration is drawn. b Phosphatidylserine is synthesized by
phatidylserine. a Total synthesis established the structure of papua- Cho1 from serine and DAG-CDP. Cells lacking the cho1 gene con-
mide B [91]. Mirabamide A is a chlorinated, glycosylated congener of ferred altered sensitivity to papuamide B. DAG-CDP: 1,2-diacyl-sn-
papuamides. Since the stereochemistry of 2,3-diaminobutanoic acid in glycero-3-cytidine-5’-diphosphate
papuamides A, B and mirabamide A was reported to be same [85, 87],Marine natural products targeting the eukaryotic cell membrane 775 trimethyldeca-(4Z,6E)-dienoic acid. Several amino acids are involved in the regulation of phospholipid asymmetry nonproteinogenic. Stereochemistries of the amino acids [102, 103]. Papuamide A was used in a screening for drugs were analyzed by chemical degradation and derivatization that perturb fungal PS synthase [92]. Since PS synthase is of the resulting products, followed by chromatographic essential for virulence in Candida albicans [104], com- comparisons of the derivatives with synthetic or commer- pounds that antagonize the growth inbitory activity of cially available standards [85, 89, 90]. Total synthesis papuamide A by inhibiting PS synthesis were expected to established the stereochemistry of these cyclodepsipeptides be drug candidates. [91]. Papuamides and mirabamides inhibit HIV-1 infection Papuamides inhibit growth of the yeasts Saccharomyces [85, 87, 88]. The anti-HIV activity was shown to be exerted cerevisiae and Candida albicans [92, 93]. The cellular through inhibiting virus entry by directly interacting with target of papuamides in yeast is phosphatidylserine (PS), the virus [105]. Papuamides also showed potent cytotoxicity which was revealed by a budding yeast chemical genomic against human cancer cells. In spite of the potency, the analysis coupled with a traditional genetic approach target molecule in the anti-HIV activity, and cytotoxicity (Fig. 6b) [93]. Parsons et al. collected chemical–genetic remains to be elucidated [105]. interactions by testing the activity of 82 compounds and crude natural product extracts against ~4800 haploid dele- Saponins from sea cucumber tion mutants [93]. Papuamide B showed more than 300 chemical–genetic interactions with genes whose gene Marine invertebrates and plants contain saponins, which ontology annotations were enriched by vesicle-mediated often show high sterol-binding and membrane lytic activ- transport, cell wall organization and biogenesis, and protein ities [106–109]. Holotoxins, saponins from the sea modification [94]. This result suggested that papuamide B cucumber Stichopus japonicus, are a part of the formulation may modulate intracellular membrane trafficking or perturb of a drug used to treat athlete’s foot (Fig. 7) [110, 111]. some events on the cell surface. Hierarchical analysis of the Holotoxins exhibit potent growth inhibition against various chemical–genetic interaction profile of papuamide B and a fungi, including Trichophyton rubrum and Candida albi- set of 132 genome-wide genetic interaction profiles [95] cans [112]. Saponins in sea cucumbers have deterrent effect unveiled that papuamide B treatment and cell-surface against predators, likely as an aposematic signal [113]. In mutants have similar cellular defects. The binding target addition, sea cucumber saponins were shown to work as was identified through classical genetic approach. Papua- kairomones that attract a symbiotic crab [114]. Several mide B-resistant mutants were obtained by inoculating saponins were shown to bind to sterols, and the stoichio- wild-type cells with high concentrations of papuamide B. metry was assessed to be 1:1 for some plant saponins The drug-resistant strain was revealed to be a cho1 null [109, 115–117]. Importantly, their sterol specificity and mutant by genetic analyses. The CHO1 gene encodes biological activities are affected by minor structural differ- phosphatidylserine synthase, and the null mutant lacks the ences of the aglycon and the sugar portion [117]. The ability to synthesize PS (Fig. 6b) [96–98]. Based on the number of natural saponins is huge; e.g., more than 700 chemical genomic analysis in which papuamide B was triterpene glycosides have been reported from sea cucum- suggested to target cell-surface molecules, the compound bers and more than 1000 from marine organisms [106–109]. was expected to recognize PS directly. In a dye leakage Below, only a couple of representatives are introduced. experiment using liposomes, papuamide B induced dye Stichlorosides are antifungal saponins from the sea leakage of PC liposomes containing 10 mol% PS, which cucumber Stichopus chloronotus (Fig. 7) [118]. Stichloro- was about 100-fold more potent than those against PC side was rediscovered in the analysis of the compendium of liposomes with 10% PE or pure PC liposomes. Taken chemical–genetic interaction profiles generated by testing together, papuamide B was concluded to target PS to the collection of viable yeast haploid deletion mutants compromise the yeast cell membrane integrity. So far, the [119]. Parsons et al. [93] found that a crude extract of a sea recognition mechanism of PS by pauamides is not known. cucumber and another extract of a marine sponge showed Papuamides are one of the rare molecules that recognize similar chemical–genetic interaction profiles. They purified glycerophospholipids with high specificity, thus this natural stichloroside and theopalauamide from the extracts and product is a useful research tool. For example, the amount showed that these two chemically distinct compounds may of PS in the outer leaflet of the plasma membrane can be share their cellular target. Later, the same group developed a assessed by testing cellular sensitivity to papuamides, i.e., molecular barcoded yeast open reading frame (MoBY- mutant cells with tolerance to papuamides are expected to ORF) library, by which they identified the gene associated have decreased amount of PS. This strategy allowed sci- with the drug-resistant phenotype of theopalauamide/sti- entists to investigate the cellular function of aminopho- chloroside-resistant (theoR) mutant [119]. The identified spholipid translocases [99–101] and to explore new factors gene was MVD1, encoding mevalonate pyrophosphate
776 S. Nishimura
holotoxin A stochloroside C1 neothyonidioside
psolusoside A psolusoside B
Fig. 7 Chemical structures of marine-derived saponins
decarboxylase, an essential enzyme involved in an early a
step of the ergosterol biosynthesis pathway. In the dye
leakage assay using ergosterol-containing liposome, sti-
chloroside showed ergosterol-enhanced leakage, suggesting
that this compound targets membrane ergosterol to exert
squalamine
antifungal activity.
Neothyonidioside is a fungicidal triterpene glycoside
b
from the sea cucumber Australostichopus mollis (Fig. 7).
This compound is potently cytotoxic to S. cerevisiae but
does not permeabilize cellular membranes [120]. Mutant
cells with reduced ergosterol content conferred resistance to DMPS
neothyonidioside while exogenous ergosterol suppressed its Fig. 8 Chemical structures of squalamine (a) and 1,2-dimyristoyl-sn-
toxicity, suggesting that ergosterol is its direct target. Pso- glycero-3-phospho-L-serine (DMPS) (b). Squalamine and α-synuclein
lusoside A, isolated from the holothurian Psolus fabricii, competed on the lipid vesicles of DMPS
has a similar structure with that of neothyonidioside but the
number of sulfates differs (Fig. 7). Psolusoside A formed a
complex with cholesterol and showed potent hemolytic Squalamine has a steroidal skeleton with a sulfated side
activity and moderate inhibitory activity to the rat brain Na chain, and a spermidine group is substituted at the C-3
+
, K+-ATPase [121]. In contrast, psolusoside B, isolated position. The structure was readily confirmed by total
from the same organism, failed to form a complex with synthesis, 1 year after its discovery [126]. Squalamine
cholesterol, and its biological activities were less potent shows multiple biological activities, e.g., antimicrobial
than psolusoside A [121, 122]. Binding to cholesterol seems activities against fungi, protozoa, and both Gram-negative
to be the cause of the biological activities of psolusoside A, and Gram-positive bacteria [123], broad-spectrum antiviral
although the molecular mechanism remains to be activity [127], anti-angiogenesis [128], and reduction of α-
elucidated. synuclein aggregation [129].
The molecular mechanism of the bioactivities of squa-
Squalamine from shark and lamprey lamine is not fully understood, but the membrane affinity of
squalamine, at least partly, seems to contribute to them. The
Squalamine was originally isolated from tissues of the net charge of this molecule is positive, and shows high
dogfish shark Squalus acanthias (Fig. 8) [123, 124]. Later, affinity to the anionic phospholipid, such as phosphati-
this compound was found in the plasma membrane of white dylglycerol (PG) and phosphatidylserine (PS) [130, 131]. α-
blood cells in the sea lamprey Petromyzon marinus [125]. Synuclein is a 140-residue intrinsically disordered protein,Marine natural products targeting the eukaryotic cell membrane 777
whose aggregation to form amyloid fibrils is the hallmark of [141]. The characteristic bicyclic structure is bridged by a
Parkinson’s disease. Vesicles prepared from 1,2-dimyr- histidinoalanine residue. Several congeners possess a sugar
istoyl-sn-glycero-3-phospho-L-serine (DMPS) (Fig. 8) group on the imidazole ring, e.g., theonellamide A (Fig. 9).
enhance the rate of the primary nucleation step that initiates The sugar group was not required for the biological activ-
amyloid formation by three orders of magnitude or more ities of TNMs, such as their potent antifungal activity and
[132]. Squalamine dramatically affects α-synuclein aggre- moderate cytotoxicity [133–138, 142–144], suggesting that
gation in vitro and in vivo [129]. The positively charged the bicyclic peptide framework is responsible for its unique
squalamine and α-synuclein likely competed on the lipid biological activities.
vesicles. Notably, squalamine almost completely sup- The cellular target of TNMs responsible for its antifungal
pressed the toxicity of α-synuclein oligomers in human activity is ergosterol, which was determined by yeast che-
neuroblastoma cells by inhibiting their interactions with mical genomic analyses employing a budding yeast system
lipid membranes, as well as muscle paralysis in a nematode [119] and a fission yeast system [82]. In the budding yeast
worm Caenorhabditis elegans strain overexpressing α- omics study, a MoBY-ORF library was utilized as described
synuclein. These findings suggested that membrane- in the section of saponin. It was found that theopalauamide
targeting reagents such as squalamine could have the may have a common molecular target with stichloroside
potential to act as therapeutic agents for Parkinson’s disease (Fig. 7): two metabolites showed similar chemical-genetic
and other conditions associated with the pathogenic aggre- profiles, and mutant cells resistant to stichloroside were also
gation of α-synuclein. resistant to theopalauamide [93]. The MoBY-ORF com-
plementation assay unveiled that the resistant mutant have a
Theonellamides (TNMs) from marine sponges mutation in MVD1, a gene encoding mevalonate pyropho-
sphate decarboxylase, which is involved in an early step of
Target ID of TNMs the ergosterol biosynthesis pathway. In the fission yeast
system, the ORFeome overexpression strain collection was
TNMs are bicyclic peptides consisting of 12 amino acids used, by which the effect of gene overexpression against
(Fig. 9) [133–136]. Theonegramide, theopalauamide and drug efficacy was measured [145, 146]. The sensitivity of
isotheopalauamide are congeners with minor modifications, ORFeome strains to TNM was tested to generate a chemical
all of which were isolated from marine sponges of Theo- genomic profile. Comparison of the chemical genomic
nella swinhoei [137, 138]. Cell fractionation analysis sug- profiles of TNM and reference compounds with known
gested the bacterial origin of theopalauamide [139, 140]. mechanisms of action suggested that TNM may have a
Recently, metagenomics analysis identified the uncultivated common target molecule with polyene macrolides,
Entotheonella sponge symbionts as producers of TNMs amphotericin B, and nystatin. In both omics studies, lipid
theonellamide A: R = OH
theopalauamide: R = H
TNM-AMCA:
TNM-BF:
theonellamide F
Fig. 9 Chemical structures of natural TNMs and fluorescently-labeled TNMs (fTNMs). Representatives of natural products and fTNMs derived
from TNM-A (TNM-AMCA and TNM-BF) are shown778 S. Nishimura
binding experiments using fluorescently-labeled TNMs interactions between TNM-A and cholesterol. 1H NMR
(fTNMs, Fig. 9) unambiguously showed that TNMs target chemical shift changes of TNM-A upon addition of 25-
ergosterol in yeast. hydroxycholesterol revealed that TNM-A likely has a
hydrophobic cavity that can accommodate the sterol ali-
Interaction mode of TNMs to sterol-containing membranes cyclic part, including the 3β-OH group (Fig. 10b) [147].
Authors suggested that the TNM-Chol complex becomes
TNM-BF, one of fTNMs, showed binding to ergosterol that less polar by Chol interaction, resulting in a greater accu-
was attached on hydrophobic plates [82]. In this assay, mulation of the peptide in the membrane.
structure-affinity relationship of sterols were examined Membrane binding of TNM modulates the membrane
using various sterol derivatives to show the importance of curvature and/or membrane fluidity. 31P NMR experiments
the 3β-hydroxy group (Figs. 2, 10a). Ergosterol, choles- and dynamic light scattering measurements showed the
terol, 5α-cholest-7-en-3β-ol and cholestanol showed inclusion of TNM-A in liposomes promotes smaller vesicle
equivalent binding to TNM-BF, indicating that double formation. Furthermore, confocal microscopy experiments
bonds in the ring B are not requisite, and the minor struc- using giant unilamellar vesicles (GUVs) showed that the
tural differences in the hydrocarbon tail had no significant addition of TNM-A deformed the membrane [148]. It is
effects. When the hydroxy group was acetylated, oxidized plausible that the compound binds to the membrane surface,
to ketone, or removed, TNM-BF showed no significant accumulates there through direct interaction with the 3β-OH
binding. In addition, epicholesterol possessing 3α-hydroxy moiety of sterols, and consequently changes the local
group showed no binding to TNM-BF. membrane curvature (Fig. 11a) [147–149]. Another micro-
The mode of interaction of TNM with sterols in the scopic analyses employing GUVs and fTNM demonstrated
membrane was analyzed by SPR experiments, solution and that TNMs are localized at cholesterol-containing liquid-
solid-state NMR measurements, and microscopic analyses. disordered domains rather than cholesterol-rich liquid
In the SPR experiments [28], POPC liposomes were ordered domains (Fig. 11b). This can be due to the weaker
immobilized on the SPR sensor chip. TNM-A showed lipid-cholesterol interaction in disordered domains than that
strong affinity to liposomes containing cholesterol or in ordered domains, which might be a general rule for
ergosterol, whereas epicholesterol did not enhance the sterol-targeting bioactive molecules as AM3 also showed a
membrane binding. The kinetic analyses revealed that the similar tendency [57]. Notably, in cholesterol abundant
initial binding of TNM-A to the membrane surfaces was GUVs, TNM-A induced phase separation of the membrane
markedly promoted by the presence of 3β-sterols. Direct (Fig. 11c) [150].
interactions between TNM-A and 3β-sterols were observed
in solid-state 2H NMR experiments [28]. Only a much Visualization of membrane sterols by TNM derivatives
weaker interaction of TNM-A with epicholesterol was
detected. These results revealed that TNM-A mostly fTNMs, which were derived from natural TNM-A through
recognizes the 3β-OH moieties of sterols, but do not strictly modifying D-galactose, have been used to detect membrane
distinguish the steroid skeleton or the side chain. Recently, ergosterols in eukaryotic model organisms, budding yeast,
solution NMR experiments was employed to analyze the and fission yeast (Fig. 9) [82, 119, 151]. Cholesterol in
a b
Fig. 10 Binding modes of TNMs to 3β-sterols. a Structure-affinity binding pocket or a cavity. Yellow asterisks denote hydrogen atoms
relationships of sterols. 3β-Hydroxy group is essential (orange), double that show significant changes in the NMR experiments upon the
bonds in the ring B are not required (green), the minor structural addition of 25-hydroxycholesterol. Reprinted from ref. [147], Copy-
variation in the tail (blue) has little impact on the binding of TNM-BF. right 2019, with permission from Elsevier
b A hypothetical 3D structure of TNM-A that possesses a hydrophobicMarine natural products targeting the eukaryotic cell membrane 779
cultured mammalian cells and tissues can also be detected cytoskeleton-, and energy-dependent phenomenon [150].
under fluorescence microscopy [152–155]. Furthermore, Modulation of the membrane order was observed after
TNM-BF possessing BODIPY is used to detect cholesterol TNM-A treatment, which was consistent with the micro-
under electron microscopy (Fig. 9) [156, 157]. Proteinous scopic observation using GUVs. Analysis using TNMs
probes, such as perfringolysin O derivatives, can also detect revealed that cholesterol maintains proper membrane order
sterols under electron microscopy [158, 159]. Comparative for regulating cell shape.
analysis using fTNMs and proteinous probes would unveil
novel sterol functions since they have different membrane-
binding properties [158]. Summary and perspectives
Cellular effects of TNMs Amphiphilic compounds tend to reside in lipid membranes.
Rigid and planar conformation of molecules also support
TNMs induce unique morphological changes in yeast and membrane residence [160]. Many chemicals including both
mammalian cells. In the fission yeast Schizosaccharomyces natural and synthetic ones potentially target cell mem-
pombe, TNM-F induced overproduction of 1,3-β-D-glucan in branes. To recognize a lipid molecule as a guest in a
a Rho1- and Bgs1-dependent manner [82]. This phenotype 1:1 stoichiometry, the host molecule is required to be large
was not observed when cells were treated with polyene enough to construct a cavity, e.g., cinnamycin [16]. In this
antifungals, amphotericin B and nystatin. As described regard, natural products have an advantage since its mole-
above, similar effects were observed by heronamides, which cular size is variable, unrestricted by Lipinski’s rule of five
target phospholipids with saturated hydrocarbon chains [64]. [161]. From marine organisms, many membrane-targeting
In mammalian cells, TNM-A induces shrinking of cells compounds have been reported, although only a part of
transiently without cytotoxicity, which is a cholesterol-, them, whose molecular mechanisms were recently reported,
a
b c
TNM-AMCA EGFP-Lysenin 0 min 20 min
control
TNM-A
rhodamine-DOPE merge
Fig. 11 Membrane order recognized and modulated by TNM. a A are shown. GUVs were incubated with 1 μM TNM-AMCA (blue) and
possible mechanism of TNM-A-induced membrane morphological EGFP-lysenin (green). Scale bar represents 10 μm. c Phase separation
alteration. TNM-A efficiently binds to the surface of a lipid bilayer of lipid membranes by TNM-A. GUVs (DOPC/SM/Chol/Rh-DOPE,
containing cholesterol. The complex may show deeper association in 24:25:50:1) were incubated with 1 μM TNM-A for 20 min, and con-
the membrane and alter the membrane curvature. b Localization of focal sections in the equatorial plane of GUVs are shown. Scale bar,
TNM-AMCA in liquid-disordered domains. Confocal sections in the 10 μm. Reprinted from refs. [147, 150]. Copyright 2015 and 2019,
equatorial plane of a GUV (DOPC/SM/Chol/Rh-DOPE = 33:33:33:1) with permission from Elsevier780 S. Nishimura
are included in this review. In spite of their specific and Compliance with ethical standards
potent biological activities, the mechanism of their mole-
cular recognition remains to be elusive. Precise under- Conflict of interest The author declares no competing interests.
standing of the molecular mechanism will be helpful for the
Publisher’s note Springer Nature remains neutral with regard to
rational design of molecules which selectively recognize jurisdictional claims in published maps and institutional affiliations.
lipids that are specifically detected in sites of disease or
found in pathogens.
References
We have not fully mined the biological potential of
marine natural products. This is partly due to the limited 1. Carroll AR, Copp BR, Davis RA, Keyzers RA, Prinsep MR.
amount of reported compounds, which hampers annotating Marine natural products. Nat Prod Rep. 2021;38:362–413.
unique biological activities and molecular targets. Sys- 2. Pye CR, Bertin MJ, Lokey RS, Gerwick WH, Linington RG. Ret-
rospective analysis of natural products provides insights for future
tematic screening system employing omics materials and in
discovery trends. Proc Natl Acad Sci USA. 2017;114:5601–6.
silico computational analyses are promising ways for 3. McCauley EP, Piña IC, Thompson AD, Bashir K, Weinberg M,
comprehensive analyses using minute amounts of bioactive Kurz SL, et al. Highlights of marine natural products having
compounds [162, 163]. Finding new compounds is another parallel scaffolds found from marine-derived bacteria, sponges,
and tunicates. J Antibiot. 2020;73:504–25.
issue, which is being tackled by developing new technol-
4. Mayer AMS, Guerrero AJ, Rodríguez AD, Taglialatela-Scafati
ogy, e.g., genome mining, awaking silent/cryptic metabolic O, Nakamura F, Fusetani N. Marine Pharmacology in 2016-
pathways [164, 165], and quick structure identification 2017: marine compounds with antibacterial, antidiabetic, anti-
based on curated compound databases [166]. The rate of fungal, anti-inflammatory, antiprotozoal, antituberculosis and
antiviral activities; affecting the immune and nervous systems,
discovery of unusual structures is not changing, and there
and other miscellaneous mechanisms of action. Mar Drugs.
seems to be much left to be discovered [2]. In fact, new 2021;19:49.
membrane-targeting molecules were discovered from a 5. Lu WY, Li HJ, Li QY, Wu YC. Application of marine natural
combined culture system and an unexplored source organ- products in drug research. Bioorg Med Chem. 2021;35:116058.
6. Epand RM, Walker C, Epand RF, Magarvey NA. Molecular
ism [167–169]. Unique compound libraries coupled with
mechanisms of membrane targeting antibiotics. Biochim Bio-
innovative screening approaches will identify many more phys Acta. 2016;1858:980–7.
membrane-targeting compounds. 7. Heidary M, Khosravi AD, Khoshnood S, Nasiri MJ, Soleimani S,
Saponins in sea cucumbers are harmful for most organ- Goudarzi M. Daptomycin. J Antimicrob Chemother. 2018;73:1–11.
8. Taylor SD, Palmer M. The action mechanism of daptomycin.
isms but attractants for the symbiotic crab [114]. Squala-
Bioorg Med Chem. 2016;24:6253–68.
mine, which shows broad antimicrobial activities and 9. Ostrosky-Zeichner L, Casadevall A, Galgiani JN, Odds FC, Rex
inhibits α-synuclein aggregation, is thought to be a host JH. An insight into the antifungal pipeline: selected new mole-
defense metabolite in the dogfish shark and sea lamprey cules and beyond. Nat Rev Drug Discov. 2010;9:719–27.
10. Odds FC, Brown AJ, Gow NA. Antifungal agents: mechanisms
[123, 125]. Tachyplesin-I, whose derivatives are investi-
of action. Trends Microbiol. 2003;11:272–9.
gated as anticancer leads, is a host defense ribosomal pep- 11. Ostrosky-Zeichner L, Marr KA, Rex JH, Cohen SH. Ampho-
tide in the horseshoe crab [170, 171]. So far, however, roles tericin B: time for a new “gold standard”. Clin Infect Dis.
of most marine natural products in the marine environments 2003;37:415–25.
12. te Welscher YM, ten Napel HH, Balagué MM, Souza CM,
are hidden. Even in the case of TNMs and papuamides,
Riezman H, de Kruijff B, et al. Natamycin blocks fungal growth
whose pharmacological targets are known, their ecological by binding specifically to ergosterol without permeabilizing the
roles in sponge holobionts are not known. Investigating membrane. J Biol Chem. 2008;283:6393–401.
roles in the natural environment would give us serendipi- 13. Perlin DS, Rautemaa-Richardson R, Alastruey-Izquierdo A. The
global problem of antifungal resistance: prevalence, mechanisms,
tous clues to utilize these molecules in a more rational way
and management. Lancet Infect Dis. 2017;17:e383–92.
to keep sustainable development of humanities. 14. Yetukuri L, Ekroos K, Vidal-Puig A, Oresic M. Informatics and
computational strategies for the study of lipids. Mol Biosyst.
2008;4:121–7.
Acknowledgements I would like to thank Profs Minoru Yoshida 15. Ejsing CS, Sampaio JL, Surendranath V, Duchoslav E, Ekroos
(RIKEN; Univ. Tokyo) and Hideaki Kakeya (Kyoto Univ.). Research K, Klemm RW, et al. Global analysis of the yeast lipidome by
presented here was conducted in their laboratories with talented quantitative shotgun mass spectrometry. Proc Natl Acad Sci
members. I especially thank Drs Yuko Arita, Ryosuke Sugiyama, Mr. USA. 2009;106:2136–41.
Kohei Fujita, and Takahiro Nakatani. I am grateful to Profs Shigeki 16. Nishimura S, Matsumori N. Chemical diversity and mode of
Matsunaga (Univ. Tokyo), Toshihide Kobayashi (CNRS), Nobuaki action of natural products targeting lipids in the eukaryotic cell
Matsumori (Kyushu Univ.), and Hendrik Luesch (Univ. Florida) for membrane. Nat Prod Rep. 2020;37:677–702.
exciting collaborations, and Prof. William Fenical (UCSD) for his 17. Murata M, Sugiyama S, Matsuoka S, Matsumori N. Bioactive
warm encouragement. I thank Mayuki Sasaki for reading this paper. structure of membrane lipids and natural products elucidated by a
Research presented here was supported in part by JSPS KAKENHI chemistry-based approach. Chem Rec. 2015;15:675–90.
(18K06717, 17H06401, 16K13086, 25702048), Nagase Science and 18. Ciumac D, Gong H, Hu X, Lu JR. Membrane targeting cationic
Technology Foundation, the Institute for Fermentation, Osaka (IFO), antimicrobial peptides. J Colloid Interface Sci. 2019;537:163–85.
and Suntory Institute for Bioorganic Research.Marine natural products targeting the eukaryotic cell membrane 781
19. Naito A, Matsumori N, Ramamoorthy A. Dynamic membrane polyhydroxyl chain and their membrane-permeabilizing activ-
interactions of antibacterial and antifungal biomolecules, and ities. Bioorg Med Chem. 2006;14:6548–54.
amyloid peptides, revealed by solid-state NMR spectroscopy. 37. Meng Y, Van Wagoner RM, Misner I, Tomas C, Wright JL.
Biochim Biophys Acta Gen Subj. 2018;1862:307–23. Structure and biosynthesis of amphidinol 17, a hemolytic com-
20. Marquette A, Bechinger B. Biophysical investigations elucidat- pound from Amphidinium carterae. J Nat Prod. 2010;73:409–15.
ing the mechanisms of action of antimicrobial peptides and their 38. Nuzzo G, Cutignano A, Sardo A, Fontana A. Antifungal
synergism. Biomolecules. 2018;8:18. amphidinol 18 and its 7-sulfate derivative from the marine
21. Salvador-Reyes LA, Sneed J, Paul VJ, Luesch H. Amantelides A dinoflagellate Amphidinium carterae. J Nat Prod.
and B, polyhydroxylated macrolides with differential broad- 2014;77:1524–7.
spectrum cytotoxicity from a guamanian marine cyanobacterium. 39. Cutignano A, Nuzzo G, Sardo A, Fontana A. The missing piece
J Nat Prod. 2015;78:1957–62. in biosynthesis of amphidinols: first evidence of glycolate as a
22. Shao CL, Linington RG, Balunas MJ, Centeno A, Boudreau P, starter unit in new polyketides from Amphidinium carterae. Mar
Zhang C, et al. Bastimolide A, a potent antimalarial polyhydroxy Drugs. 2017;15:157.
macrolide from the marine cyanobacterium Okeania hirsuta. J 40. Satake M, Cornelio K, Hanashima S, Malabed R, Murata M,
Org Chem. 2015;80:7849–55. Matsumori N, et al. Structures of the largest amphidinol homo-
23. Keller L, Siqueira-Neto JL, Souza JM, Eribez K, LaMonte GM, logues from the dinoflagellate Amphidinium carterae and
Smith JE, et al. Palstimolide A: a complex polyhydroxy mac- structure-activity relationships. J Nat Prod. 2017;80:2883–8.
rolide with antiparasitic activity. Molecules. 2020;25:1604. 41. Martínez KA, Lauritano C, Druka D, Romano G, Grohmann T,
24. MacMillan JB, Molinski TF. Caylobolide A, a unique 36- Jaspars M, et al. Amphidinol 22, a new cytotoxic and antifungal
membered macrolactone from a Bahamian Lyngbya majuscula. amphidinol from the dinoflagellate Amphidinium carterae. Mar
Org Lett. 2002;4:1535–8. Drugs. 2019;17:E385.
25. Salvador LA, Paul VJ, Luesch H. Caylobolide B, a macrolactone 42. Paul GK, Matsumori N, Konoki K, Sasaki M, Murata M,
from symplostatin 1-producing marine cyanobacteria Phormi- Tachibana K. Harmful and toxic algal blooms. Yasumoto T,
dium spp. from Florida. J Nat Prod. 2010;73:1606–9. Oshima Y, Fukuyo Y, editors. I.O.C. of UNESCO: Paris; 1996
26. Shao CL, Mou XF, Cao F, Spadafora C, Glukhov E, Gerwick L, p. 503–506.
et al. Bastimolide B, an antimalarial 24-membered marine 43. Huang SJ, Kuo CM, Lin YC, Chen YM, Lu CK. Carteraol E, a
macrolide possessing a tert-butyl group. J Nat Prod. potent polyhydroxyl ichthyotoxin from the dinoflagellate
2018;81:211–5. Amphidinium carterae. Tetrahedron Lett. 2009;50:2512–5.
27. Elsadek LA, Matthews JH, Nishimura S, Nakatani T, Ito A, Gu 44. Huang XC, Zhao D, Guo YW, Wu HM, Lin LP, Wang ZH, et al.
T, et al. Genomic and targeted approaches unveil the cell Lingshuiol, a novel polyhydroxyl compound with strongly
membrane as a major target of the antifungal cytotoxin aman- cytotoxic activity from the marine dinoflagellate Amphidinium
telide A. Chembiochem. 2021;22:1790–9. sp. Bioorg Med Chem Lett. 2004;14:3117–20.
28. Espiritu RA, Matsumori N, Murata M, Nishimura S, Kakeya H, 45. Huang XC, Zhao D, Guo YW, Wu HM, Trivellone E, Cimino G.
Matsunaga S, et al. Interaction between the marine sponge cyclic Lingshuiols A and B, two new polyhydroxy compounds from the
peptide theonellamide A and sterols in lipid bilayers as viewed Chinese marine dinoflagellate Amphidinium sp. Tetrahedron
by surface plasmon resonance and solid-state (2)H nuclear Lett. 2004;45:5501–4.
magnetic resonance. Biochemistry. 2013;52:2410–8. 46. Washida K, Koyama T, Yamada K, Kita M, Uemura D. Kar-
29. Espiritu RA, Matsumori N, Tsuda M, Murata M. Direct and atungiols A and B, two novel antimicrobial polyol compounds,
stereospecific interaction of amphidinol 3 with sterol in lipid from the symbiotic marine dinoflagellate Amphidinium sp. Tet-
bilayers. Biochemistry. 2014;53:3287–93. rahedron Lett. 2006;47:2521–5.
30. Houdai T, Matsuoka S, Matsumori N, Murata M. Membrane- 47. Hanif N, Ohno O, Kitamura M, Yamada K, Uemura D. Sym-
permeabilizing activities of amphidinol 3, polyene-polyhydroxy biopolyol, a VCAM-1 inhibitor from a symbiotic dinoflagellate
antifungal from a marine dinoflagellate. Biochim Biophys Acta. of the jellyfish Mastigias papua. J Nat Prod. 2010;73:1318–22.
2004;1667:91–100. 48. Kubota T, Takahashi A, Tsuda M, Kobayashi J. Luteophanol D,
31. Satake M, Murata M, Yasumoto T, Fujita T, Naoki H. Amphi- new polyhydroxyl metabolite from marine dinoflagellate
dinol, a polyhydroxypolyene antifungal agent with an unprece- Amphidinium sp. Mar Drugs. 2005;3:113–8.
dented structure, from a marine dinoflagellate, Amphidinium- 49. Inuzuka T, Yamada K, Uemura D. Amdigenols E and G, long
klebsii. J Am Chem Soc. 1991;113:9859–61. carbon-chain polyol compounds, isolated from the marine
32. Paul GK, Matsumori N, Murata M, Tachibana K. Isolation and dinoflagellate Amphidinium sp. Tetrahedron Lett.
chemical-structure of amphidinol-2, a potent hemolytic com- 2014;55:6319–23.
pound from marine dinoflagellate Amphidinium-klebsii. Tetra- 50. Murata M, Matsuoka S, Matsumori N, Paul GK, Tachibana K.
hedron Lett. 1995;36:6279–82. Absolute configuration of amphidinol 3, the first complete
33. Paul GK, Matsumori N, Konoki K, Murata M, Tachibana K. structure determination from amphidinol homologues: Applica-
Chemical structures of amphidinols 5 and 6 isolated from marine tion of a new configuration analysis based on carbon-hydrogen
dinoflagellate Amphidinium klebsii and their cholesterol- spin-coupling constants. J Am Chem Soc. 1999;121:870–1.
dependent membrane disruption. Mar Biotechnol. 1997;5:124–8. 51. Wakamiya Y, Ebine M, Murayama M, Omizu H, Matsumori N,
34. Morsy N, Matsuoka S, Houdai T, Matsumori N, Adachi S, Murata M, et al. Synthesis and stereochemical revision of the
Murata M, et al. Isolation and structure elucidation of a new C31-C67 fragment of Amphidinol 3. Angew Chem Int Ed.
amphidinol with a truncated polyhydroxyl chain from Amphi- 2018;57:6060–4.
dinium klebsii. Tetrahedron 2005;61:8606–10. 52. Wakamiya Y, Ebine M, Matsumori N, Oishi T. Total synthesis
35. Echigoya R, Rhodes L, Oshima Y, Satake M. The structures of of Amphidinol 3: a general strategy for synthesizing Amphidinol
five new antifungal and hemolytic amphidinol analogs from analogues and structure-activity relationship study. J Am Chem
Amphidinium carterae collected in New Zealand. Harmful Soc. 2020;142:3472–8.
Algae. 2005;4:383–9. 53. Morsy N, Houdai T, Konoki K, Matsumori N, Oishi T, Murata
36. Morsy N, Houdai T, Matsuoka S, Matsumori N, Adachi S, Oishi M. Effects of lipid constituents on membrane-permeabilizing
T, et al. Structures of new amphidinols with truncated activity of amphidinols. Bioorg Med Chem. 2008;16:3084–90.782 S. Nishimura
54. Iwamoto M, Sumino A, Shimada E, Kinoshita M, Matsumori N, two distinct PKS systems and amino acid processing enzymes
Oiki S. Channel formation and membrane deformation via sterol- for generation of the aminoacyl starter unit. Chem Biol.
aided polymorphism of Amphidinol 3. Sci Rep. 2017;7:10782. 2009;16:1109–21.
55. Houdai T, Matsuoka S, Morsy N, Matsumori N, Satake M, 72. Hampsey M. A review of phenotypes in Saccharomyces cere-
Murata M. Hairpin conformation of amphidinols possibly visiae. Yeast. 1997;13:1099–133.
accounting for potent membrane permeabilizing activities. Tet- 73. Iwaki T, Iefuji H, Hiraga Y, Hosomi A, Morita T, Giga-Hama Y,
rahedron. 2005;61:2795–802. et al. Multiple functions of ergosterol in the fission yeast Schi-
56. Houdai T, Matsumori N, Murata M. Structure of membrane- zosaccharomyces pombe. Microbiology. 2008;154:830–41.
bound Amphidinol 3 in isotropic small bicelles. Org Lett. 74. Simons K, Vaz WL. Model systems, lipid rafts, and cell mem-
2008;10:4191–4. branes. Annu Rev Biophys Biomol Struct. 2004;33:269–95.
57. Hieda M, Sorada A, Kinoshita M, Matsumori N. Amphidinol 3 75. Huang J, Feigenson GW. A microscopic interaction model of
preferentially binds to cholesterol in disordered domains and maximum solubility of cholesterol in lipid bilayers. Biophys J.
disrupts membrane phase separation. Biochem Biophys Rep. 1999;76:2142–57.
2021;26:100941. 76. Mesmin B, Maxfield FR. Intracellular sterol dynamics. Biochim
58. Kojiri K, Nakajima S, Suzuki H, Kondo H, Suda H. A new Biophys Acta. 2009;1791:636–45.
macrocyclic lactam antibiotic, BE-14106. I. Taxonomy, isola- 77. McConnell HM, Radhakrishnan A. Condensed complexes of
tion, biological activity and structural elucidation. J Antibiot. cholesterol and phospholipids. Biochim Biophys Acta.
1992;45:868–74. 2003;1610:159–73.
59. Fujita K, Sugiyama R, Nishimura S, Ishikawa N, Arai MA, 78. Sakanishi K, Itoh S, Sugiyama R, Nishimura S, Kakeya H,
Ishibashi M, et al. Stereochemical assignment and biological Iwabuchi Y, et al. Total synthesis of the proposed structure of
evaluation of BE-14106 unveils the importance of one acetate heronamide C. Eur J Org Chem. 2014;2014:1376–80.
unit for the antifungal activity of polyene macrolactams. J Nat 79. Cortes JC, Ishiguro J, Duran A, Ribas JC. Localization of the
Prod. 2016;79:1877–80. (1,3)β-D-glucan synthase catalytic subunit homologue Bgs1p/
60. Takahashi I, Oda Y, Nishiie Y, Ochiai K, Mizukami T. GT32-B Cps1p from fission yeast suggests that it is involved in septation,
new 20-membered macrocyclic lactam antibiotic. J Antibiot. polarized growth, mating, spore wall formation and spore ger-
1997;50:186–8. mination. J Cell Sci. 2002;115:4081–96.
61. Jørgensen H, Degnes KF, Dikiy A, Fjaervik E, Klinkenberg G, 80. Arellano M, Duran A, Perez P. Rho 1 GTPase activates the (1-3)
Zotchev SB. Insights into the evolution of macrolactam bio- beta-D-glucan synthase and is involved in Schizosaccharomyces
synthesis through cloning and comparative analysis of the bio- pombe morphogenesis. EMBO J. 1996;15:4584–91.
synthetic gene cluster for a novel macrocyclic lactam, ML-449. 81. Feoktistova A, Magnelli P, Abeijon C, Perez P, Lester RL,
Appl Environ Microbiol. 2010;76:283–93. Dickson RC, et al. Coordination between fission yeast glucan
62. Raju R, Piggott AM, Conte MM, Capon RJ. Heronamides A-C, formation and growth requires a sphingolipase activity. Genetics.
new polyketide macrolactams from an Australian marine-derived 2001;158:1397–411.
Streptomyces sp. A biosynthetic case for synchronized tandem 82. Nishimura S, Arita Y, Honda M, Iwamoto K, Matsuyama A,
electrocyclization. Org Biomol Chem. 2010;8:4682–9. Shirai A, et al. Marine antifungal theonellamides target 3β-
63. Zhang W, Li S, Zhu Y, Chen Y, Chen Y, Zhang H, et al. Her- hydroxysterol to activate Rho1 signaling. Nat Chem Biol.
onamides D-F, polyketide macrolactams from the deep-sea-derived 2010;6:519–26.
Streptomyces sp. SCSIO 03032. J Nat Prod. 2014;77:388–91. 83. Lingwood D, Simons K. Lipid rafts as a membrane-organizing
64. Sugiyama R, Nishimura S, Matsumori N, Tsunematsu Y, Hattori principle. Science. 2010;327:46–50.
A, Kakeya H. Structure and biological activity of 84. Liu J, Tang X, Wang H, Oliferenko S, Balasubramanian MK.
8-deoxyheronamide C from a marine-derived Streptomyces sp.: The localization of the integral membrane protein Cps1p to the
heronamides target saturated hydrocarbon chains in lipid mem- cell division site is dependent on the actomyosin ring and the
branes. J Am Chem Soc. 2014;136:5209–12. septation-inducing network in Schizosaccharomyces pombe.
65. Sugiyama R, Nishimura S, Kakeya H. Stereochemical reassign- Mol Biol Cell. 2002;13:989–1000.
ment of heronamide A, a polyketide macrolactam from Strep- 85. Ford PW, Gustafson KR, McKee TC, Shigematsu N, Maurizi
tomyces sp. Tetrahedron Lett. 2013;54:1531–3. LK, Pannell LK, et al. Papuamides A−D, HIV-inhibitory and
66. Kanoh N, Itoh S, Fujita K, Sakanishi K, Sugiyama R, Terajima cytotoxic depsipeptides from the sponges Theonella mirabilis
Y, et al. Asymmetric total synthesis of heronamides A-C: ste- and Theonella swinhoei collected in Papua New Guinea. J Am
reochemical confirmation and impact of long-range stereo- Chem Soc. 1999;121:5899–909.
chemical communication on the biological activity. Chemistry. 86. Prasad P, Aalbersberg W, Feussner KD, Van Wagoner RM.
2016;22:8586–95. Papuamides E and F, cytotoxic depsipeptides from the marine
67. Yu P, Patel A, Houk KN. Transannular [6 + 4] and ambimodal sponge Melophlus sp. Tetrahedron. 2011;67:8529–31.
cycloaddition in the biosynthesis of heronamide A. J Am Chem 87. Plaza A, Gustchina E, Baker HL, Kelly M, Bewley CA. Mir-
Soc. 2015;137:13518–23. abamides A-D, depsipeptides from the sponge Siliquariaspongia
68. Booth TJ, Alt S, Capon RJ, Wilkinson B. Synchronous intra- mirabilis that inhibit HIV-1 fusion. J Nat Prod.
molecular cycloadditions of the polyene macrolactam polyketide 2007;70:1753–60.
heronamide C. Chem Commun. 2016;52:6383–6. 88. Lu Z, Van Wagoner RM, Harper MK, Baker HL, Hooper JN,
69. Ding N, Han L, Jiang Y, Li G, Zheng Z, Cao B, et al. Her- Bewley CA, et al. Mirabamides E-H, HIV-inhibitory depsipep-
onamides G-L, polyene macrolactams from Streptomyces tides from the sponge Stelletta clavosa. J Nat Prod.
niveus. RSC Adv. 2018;8:17121–31. 2011;74:185–93.
70. Zhang C, Wang X, Chen Y, He Z, Yu P, Liang Y. Dynamical 89. Oku N, Krishnamoorthy R, Benson AG, Ferguson RL, Lipton
trajectory study of the transannular [6+4] and Ambimodal MA, Phillips LR, et al. Complete stereochemistry of neampha-
cycloaddition in the biosynthesis of heronamides. J Org Chem. mide A and absolute configuration of the beta-methoxytyrosine
2020;85:9440–5. residue in papuamide B. J Org Chem. 2005;70:6842–7.
71. Jørgensen H, Degnes KF, Sletta H, Fjaervik E, Dikiy A, Her- 90. Makino K, Nagata E, Hamada Y. Synthesis of tripeptide
findal L, et al. Biosynthesis of macrolactam BE-14106 involves hydrolysate from papuamide A: determination of absoluteYou can also read

















































