Potential Physiological Relevance of ERAD to the Biosynthesis of GPI-Anchored Proteins in Yeast - MDPI
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
International Journal of
Molecular Sciences
Review
Potential Physiological Relevance of ERAD to the Biosynthesis
of GPI-Anchored Proteins in Yeast
Kunio Nakatsukasa
Graduate School of Science, Nagoya City University, Yamanohata 1, Mizuho-cho, Mizuho-ku, Nagoya,
Aichi 467-8501, Japan; nakatsukasa@nsc.nagoya-cu.ac.jp
Abstract: Misfolded and/or unassembled secretory and membrane proteins in the endoplasmic
reticulum (ER) may be retro-translocated into the cytoplasm, where they undergo ER-associated
degradation, or ERAD. The mechanisms by which misfolded proteins are recognized and degraded
through this pathway have been studied extensively; however, our understanding of the physio-
logical role of ERAD remains limited. This review describes the biosynthesis and quality control of
glycosylphosphatidylinositol (GPI)-anchored proteins and briefly summarizes the relevance of ERAD
to these processes. While recent studies suggest that ERAD functions as a fail-safe mechanism for
the degradation of misfolded GPI-anchored proteins, several pieces of evidence suggest an intimate
interaction between ERAD and the biosynthesis of GPI-anchored proteins.
Keywords: ERAD; GPI-anchored protein; Hrd1; Doa10; Ubc6; Ubc7; Ca2+ /Mn2+ P-type ATPase;
Saccharomyces cerevisiae
1. Introduction
Secretory and membrane proteins are translocated to the endoplasmic reticulum
Citation: Nakatsukasa, K. Potential (ER), where they are folded into a three-dimensional conformation. In the ER, molecular
Physiological Relevance of ERAD to chaperones recognize unfolded polypeptides and facilitate their folding by binding to
the Biosynthesis of GPI-Anchored
amino acid patches containing exposed hydrophobic side chains [1]. Proteins that acquire
Proteins in Yeast. Int. J. Mol. Sci. 2021,
the correct conformation are transported from the ER to the Golgi apparatus. However,
22, 1061. https://doi.org/10.3390/
despite such protection, the maturation of proteins is an inherently error-prone process,
ijms22031061
and misfolded proteins are frequently generated. To remove the potentially toxic misfolded
proteins, eukaryotes have evolved two systems. The first system is the unfolded protein
Academic Editor: Jun Imai
response (UPR), which involves the upregulation of the factors that increase the protein-
Received: 21 December 2020
Accepted: 19 January 2021
folding capacity of the ER. The UPR also facilitates the transport of misfolded proteins
Published: 21 January 2021
to the lysosome/vacuole, where they are degraded. The stress conditions caused by the
accumulation of misfolded proteins (ER stress) induce membrane expansion of the ER to
Publisher’s Note: MDPI stays neutral
alleviate the stress independently of an increase in ER chaperone levels [2,3]. The second
with regard to jurisdictional claims in
system is ER-associated degradation (ERAD), by which terminally misfolded proteins
published maps and institutional affil- are specifically recognized, retained in the ER, and retro-translocated to the cytoplasm,
iations. where they are ubiquitinated and degraded by the proteasome. Components of the ERAD
machinery can also be induced by the UPR [4–6]. Thus, the UPR and ERAD constitute
two arms of the ER quality control apparatus and play critical roles in maintaining ER
homeostasis [6–12].
Copyright: © 2021 by the author.
Various “model misfolded proteins” have been developed and used for the analysis of
Licensee MDPI, Basel, Switzerland.
degradation pathways [13,14]. However, emerging evidence indicates that ERAD not only
This article is an open access article
mediates the elimination of structurally abnormal proteins in the ER, but also contributes to
distributed under the terms and the regulation of native proteins [15]. For example, ERAD targets properly folded proteins
conditions of the Creative Commons to regulate metabolic enzymes, transcription factors, and metal transporters at the plasma
Attribution (CC BY) license (https:// membrane [16–19]. To further elucidate the physiological roles of ERAD, it is imperative to
creativecommons.org/licenses/by/ identify native substrates. In addition, yeast-based genetic interaction studies may help
4.0/). discover novel associations between ERAD and other biological phenomena. This review
Int. J. Mol. Sci. 2021, 22, 1061. https://doi.org/10.3390/ijms22031061 https://www.mdpi.com/journal/ijmsInt. J. Mol. Sci. 2021, 22, 1061 2 of 14
describes the ERAD machineries in yeast and the findings of recent studies analyzing the
biogenesis and quality control of misfolded glycosylphosphatidylinositol (GPI)-anchored
proteins. The potential involvement of ERAD in manganese homeostasis, which might link
the biogenesis of GPI-anchored proteins to ERAD, is also discussed.
2. ER-Associated Degradation in Yeast
2.1. The Hrd1 Pathway
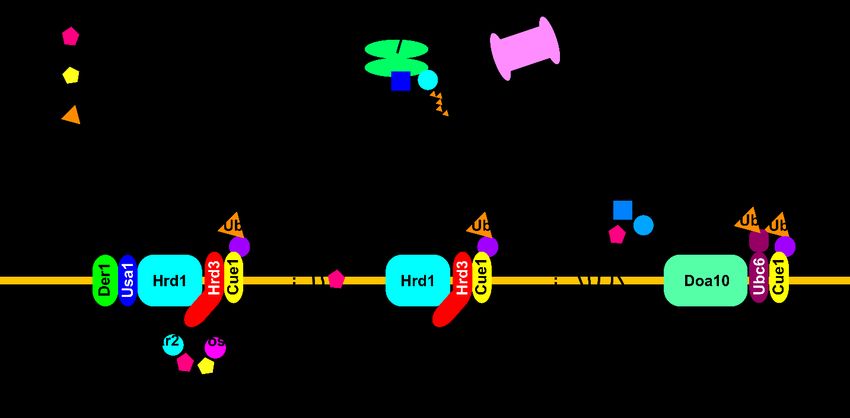
In yeast (S. cerevisiae), two dedicated ER membrane-associated E3 ligase complexes
are involved in the recognition and degradation of misfolded proteins in the ER. The
Hrd1 E3 ligase complex recognizes and targets misfolded luminal proteins, as well as
ER membrane proteins with lesions at the transmembrane domain, for ubiquitination
and degradation (Figure 1). These pathways are the so-called ERAD-L and -M pathways,
respectively [17,20–27]. The Hrd1 complex consists of Hrd3, Usa1, Der1, Dfm1, Yos9,
Kar2, Ubc7, and Cue1. Sucrose density gradient and systematic immunopurification
analyses showed that Hrd1, Hrd3, Usa1, Der1, and Yos9 comprise the core complex [25,28].
During ERAD, a misfolded substrate is first recognized by several factors, including Yos9
(luminal lectin), Der1 (transmembrane protein), Kar2 (Hsp70 chaperone), or directly by
Hrd1. The recognition mechanism for misfolded luminal glycoproteins has been studied
extensively. The N-linked glycan is trimmed by glycosidase, and the terminal α1,6-mannose
residue is recognized by the mannose 6-phosphate receptor homology (MRH) domain of
Yos9 [29–33]. Non-glycosylated misfolded substrates may be recognized by Kar2. Both
Yos9 and Kar2 bind to the luminal domain of Hrd3. In addition, unfolded and extended
polypeptide segments may be recognized by the luminal domain of Hrd3 [34]. Until
recently, differentiating the function of Hrd3 from Hrd1 stabilization was difficult because
depletion of Hrd3 results in Hrd1 instability. However, removal of the ubiquitin-like
domain of Usa1 caused Hrd1 to remain stable. This method was used to demonstrate
that Hrd3 plays a direct role in facilitating the transfer of ubiquitin to substrates [35].
Usa1 mediates the interaction of Hrd1 with Der1, which is an inactive form of rhomboid
protease, and transports substrates from the ER lumen to Hrd1 for degradation. Usa1 can
also mediate the formation of an Hrd1 oligomer, which is critical for the degradation of
ERAD-M substrates [21,27,36]. After recognition, the substrate is transferred to the Hrd1
complex for polyubiquitination. The transmembrane protein Cue1 is an ER membrane
protein that recruits the E2 ubiquitin-conjugating enzyme Ubc7 to the Hrd1 complex [37].
Hrd1 also recruits the ER membrane protein Ubx2 to the complex, which anchors the
AAA+ ATPase Cdc48/p97 to the membrane [38,39]. Then, Cdc48/p97 hydrolyzes ATP and
liberates the substrate from the ER. Inactivation of Cdc48 leads to a formation of stalled
retro-translocation complex containing Hrd1, Usa1, Hrd3, Der1, the 26S proteasome, Yos9,
ubiquitinated substrates, and Cdc48. This suggests that substrate recognition and retro-
translocation might be coupled, at least for some substrates [40]. Recognition of the integral
membrane ERAD-M substrate is mediated by the transmembrane domain of Hrd1 [41]
(Figure 1). The membrane substrates may exit the ER through a distinct pathway mediated
by the Dfm1 rhomboid protein, which can also recruit the AAA+ ATPase Cdc48/p97 to the
membrane for retro-translocation. Upon deletion of the DFM1 gene, Hrd1 levels increase
and the Hrd1 complex may be remodeled, thereby enabling a novel route of membrane
protein retro-translocation. This supports the functional flexibility of the Hrd1 complex in
response to ER stress [42,43].
2.2. The Doa10 Pathway
Another E3 ligase complex involved in yeast ERAD is Doa10, which resides in the
nuclear membrane and ER membrane. Doa10 is a 150 kDa protein with 14 transmembrane
segments [44] that targets transmembrane substrates with cytoplasmic lesions for ubiquiti-
nation and degradation, a pathway called ERAD-C (Figure 1). Substrates with abnormal
structural domains in multiple regions may be degraded depending on both the Hrd1
and Doa10 complexes [45,46]. Substrates of the Doa10 complex include single- or multi-2.2. The Doa10 Pathway
Another E3 ligase complex involved in yeast ERAD is Doa10, which resides in the
nuclear membrane and ER membrane. Doa10 is a 150 kDa protein with 14 transmembrane
segments [44] that targets transmembrane substrates with cytoplasmic lesions for ubiqui-
Int. J. Mol. Sci. 2021, 22, 1061 tination and degradation, a pathway called ERAD-C (Figure 1). Substrates with abnormal 3 of 14
structural domains in multiple regions may be degraded depending on both the Hrd1 and
Doa10 complexes [45,46]. Substrates of the Doa10 complex include single- or multi-span-
spanning
ning membrane
membrane proteins
proteins in the
in the ER ER andand
thethe inner
inner nuclearmembrane,
nuclear membrane,asaswellwellasas soluble
soluble
proteins in the cytosol and nucleoplasm [19,44,47–52]. Unlike the Hrd1 complex, which
proteins
comprises multiple
comprises multiplecomponents,
components,the theDoa10
Doa10complex
complex is relatively simple
is relatively and contains
simple three
and contains
ubiquitination
three enzymes:
ubiquitination Ubc6, Ubc7,
enzymes: Ubc6, and Cue1.
Ubc7, andHowever, in contrast
Cue1. However, to the Hrd1
in contrast pathway,
to the Hrd1
the mechanism
pathway, by which by
the mechanism thewhich
Doa10the complex
Doa10 recognizes misfolded
complex recognizes substrates
misfolded is not well
substrates is
understood. Cytosolic chaperones, such as the Hsp70 Ssa1 and
not well understood. Cytosolic chaperones, such as the Hsp70 Ssa1 and the cytosolicthe cytosolic Hsp40s Ydj1
and Hlj1,
Hsp40s facilitate
Ydj1 substrate
and Hlj1, recognition
facilitate substrate[53–55].
recognition The [53–55].
degrons The
of Doa10
degronssubstrates
of Doa10 can be
sub-
cytoplasmic [49,50] or located within the TM region [51]. During ubiquitination,
strates can be cytoplasmic [49,50] or located within the TM region [51]. During ubiquiti- Ubc6
attachesUbc6
nation, the first ubiquitin
attaches to aubiquitin
the first substrate,toand Ubc7 extends
a substrate, ubiquitin
and Ubc7 chains
extends using mostly
ubiquitin chains
lysine mostly
using 48 linkages [56].
lysine 48 Ubiquitinated
linkages [56]. substrates
Ubiquitinated are then retro-translocated
substrates to the cytosol by
are then retro-translocated
Cdc48/p97,
to the cytosolwhich is recruited
by Cdc48/p97, to the
which ER membrane
is recruited to thebyERUbx2 and/orby
membrane Dfm1
Ubx2[43,53,57].
and/or Dfm1The
mechanism underlying the retro-translocation of membrane substrates,
[43,53,57]. The mechanism underlying the retro-translocation of membrane substrates, in- including Doa10
substrates,
cluding is ill-defined.
Doa10 substrates, The
is extraction
ill-defined.ofThe Ubc6, which isofdegraded
extraction in a Doa10-dependent
Ubc6, which is degraded in a
manner [50,58], was recently reconstituted in vitro [52].
Doa10-dependent manner [50,58], was recently reconstituted in vitro [52]. The results of this
The experiment
results of
this experiment suggest that the luminal domain is unfolded by the action of cytosol
suggest that the luminal domain is unfolded by the action of Cdc48/p97 in the and
Cdc48/p97
crosses the membrane in an unfolded state.
in the cytosol and crosses the membrane in an unfolded state.
Figure
Figure 1.
1. The
The ERAD
ERAD (ER-associated
(ER-associated degradation)
degradation) pathway
pathway in
in Saccharomyces
Saccharomyces cerevisiae (see text
cerevisiae (see text for
for detail).
detail).
3. GPI-Anchored Proteins
3. GPI-Anchored Proteins
GPI anchors are structurally complex glycophospholipids that are post-translation-
GPI anchors are structurally complex glycophospholipids that are post-translationally
ally attached to the C-terminus of secretory proteins (Figure 2). The highly conserved core
attached to the C-terminus of secretory proteins (Figure 2). The highly conserved core
structure of the GPI anchor precursor (CP2: complete precursor 2), which accumulates in
structure of the GPI anchor precursor (CP2: complete precursor 2), which accumulates in
the GPI transamidase mutant, comprises four mannoses (Man1, Man2, Man3, and Man4),
the GPI transamidase mutant, comprises four mannoses (Man1, Man2, Man3, and Man4),
three ethanolamine phosphate (EtN-P) substituents on Man1, Man2, and Man3, one acyl-
three ethanolamine phosphate (EtN-P) substituents on Man1, Man2, and Man3, one acyl-
phosphatidylinositol (acyl-PI), and one glucosamine (GlcN) [59]. More than 20 genes en-
phosphatidylinositol (acyl-PI), and one glucosamine (GlcN) [59]. More than 20 genes
coding enzymes involved in the biosynthesis of GPI anchors have been identified by ge-
encoding enzymes involved in the biosynthesis of GPI anchors have been identified by
netic screening to isolate mutant cells that lack GPI proteins at the surface. GPI anchor
genetic screening to isolate mutant cells that lack GPI proteins at the surface. GPI anchor
synthesis begins on the cytosolic side of the ER membrane. The first step in GPI biosyn-
thesis is the addition of N-Acetylglucosamine (GlcNAc) to phosphatidylinositol (PI). This
transfer reaction is catalyzed by GPI-GlcNAc transferase, which consists of Gpi15, Eri1,
Gpi3, Gpi19, Gpi2, and Gpi1 [60–62]. Next, the acetyl group of GlcNAc is removed by
Gpi12, a GlcNAc-PI de-N-acetylase, to generate GlcN-PI, which is translocated by flippase
to the luminal side of the ER membrane. Inside the ER, the acyltransferase Gwt1 adds ansynthesis begins on the cytosolic side of the ER membrane. The first step in GPI biosyn-
thesis is the addition of N-Acetylglucosamine (GlcNAc) to phosphatidylinositol (PI). This
transfer reaction is catalyzed by GPI-GlcNAc transferase, which consists of Gpi15, Eri1,
Int. J. Mol. Sci. 2021, 22, 1061 Gpi3, Gpi19, Gpi2, and Gpi1 [60–62]. Next, the acetyl group of GlcNAc is removed 4 ofby
14
Gpi12, a GlcNAc-PI de-N-acetylase, to generate GlcN-PI, which is translocated by flippase
to the luminal side of the ER membrane. Inside the ER, the acyltransferase Gwt1 adds an
acyl chain,
acyl chain, which
which is
is mostly
mostlypalmitate,
palmitate,totothe
the2-position
2-positionofofthe
theinositol
inositolinin
GlcN-PI,
GlcN-PI,generating
generat-
GlcN-(acyl) PI [63]. Subsequently, Man1 and Man2 are transferred from
ing GlcN-(acyl) PI [63]. Subsequently, Man1 and Man2 are transferred from dolichol- dolicholphospho-
mannose (Dol-P-Man)
phosphomannose to GlcN-(acyl)
(Dol-P-Man) PI by GPIPImannosyl-transferase
to GlcN-(acyl) by GPI mannosyl-transferase1 (Gpi14)1and GPI
(Gpi14)
mannosyl-transferase 2 (Gpi18), respectively [64,65]. In addition, EtN-P is transferred
and GPI mannosyl-transferase 2 (Gpi18), respectively [64,65]. In addition, EtN-P is trans- from
phosphatidylethanolamine
ferred (PE) to Man1(PE)
from phosphatidylethanolamine by EtN-P transferase
to Man1 by EtN-P (Mcd4) [66]. Then,
transferase Man3
(Mcd4) is
[66].
added to Man2 by GPI mannosyl-transferase 3 (Gpi10), and Man4 is added to Man3 by
Then, Man3 is added to Man2 by GPI mannosyl-transferase 3 (Gpi10), and Man4 is added
Smp3 before the addition of EtN-P to Man3 by EtN-P transferase 3 [67–69]. This complex
to Man3 by Smp3 before the addition of EtN-P to Man3 by EtN-P transferase 3 [67–69].
comprises Gpi13, which is a catalytic subunit, and the stabilizing subunit Gpi11. Finally,
This complex comprises Gpi13, which is a catalytic subunit, and the stabilizing subunit
EtN-P is added to Man2 by EtN-P transferase enzyme 2, which consists of the catalytic
Gpi11. Finally, EtN-P is added to Man2 by EtN-P transferase enzyme 2, which consists of
subunit Gpi7 and the stabilizing subunit Gpi11 [70,71].
the catalytic subunit Gpi7 and the stabilizing subunit Gpi11 [70,71].
Figure2.2.The
Figure The pathway
pathway of of
GPIGPI (glycosylphosphatidylinositol)
(glycosylphosphatidylinositol) biosynthesis
biosynthesis in in Saccharomyces
Saccharomyces cerevisiae
cerevisiae (see
(see texttext
forfor de-
detail).
tail).
Precursors of GPI-anchored proteins have a signal for GPI anchoring at the C-terminus
and Precursors of GPI-anchored
a conventional signal sequenceproteins
for have a signal for GPI
ER translocation anchoring
at the at the The
N-terminus. C-termi-
GPI
nus and a conventional signal sequence for ER translocation at the N-terminus.
anchoring sequence contains a C-terminal hydrophobic domain that is separated from The GPI
anchoring sequence
the upstream contains a C-terminal
GPI-attachment site (the “ωhydrophobic domain
site”) by a short thatof
stretch is hydrophilic
separated from the
amino
upstream
acids [72].GPI-attachment site (the
Soon after translation “ω precursor
of the site”) by aprotein
short stretch of hydrophilic
by ribosomes amino acids
and its translocation
into the ER membrane are completed, the C-terminus of the protein is conjugated to the
amine group of EtN-P by the GPI transamidase complex, which consists of five essential
proteins: Gpi8, Gpi17, Gpi16, Gab1, and Gaa1 [73–77]. Of these, Gpi8, a catalytic subunit
that is homologous to caspase-like cysteine proteases, cleaves the C-terminus of substrate
proteins. This reaction is a prerequisite for the transamidation reaction [78–80]. After
attachment, the GPI anchor is subject to a sequence of remodeling reactions on both theInt. J. Mol. Sci. 2021, 22, 1061 5 of 14
lipid and sugar moieties. These reactions occur exclusively inside the ER and are catalyzed
by Bst1, Per1, Gup1, and Cwh43 in yeast [81–84]. Bst1 is a phosphatidylinositol deacylase
that mediates inositol deacylation. This step is required for downstream lipid remodeling.
Per1 removes the unsaturated fatty acid at the sn2 position, and Gup1 adds C26 fatty
acids. Cwh43 is responsible for replacing diacylglycerol (DAG) with ceramide, which is a
major lipid (saturated and very long inositolphosphoceramide) component of mature GPI
anchors in yeast. Subsequently, Cdc1 and Ted1 remove the EtN-P of the first and second
mannose, respectively [85,86]. These two enzymes are homologs of mammalian PGAP5,
a membrane-spanning enzyme that possesses a metal-containing phosphoesterase motif
in the luminal domain and removes EtN-P from the second mannose in the ER [87]. The
removal of EtN-P from the second mannose is a prerequisite for the recognition of GPI-
anchored proteins by the p24 complex and their exit from the ER. The localization of Cdc1
to the cis/medial Golgi apparatus, and not to the ER, was demonstrated recently. Removal
of EtN-P from the second mannose by Ted1 in the ER and from the first mannose by Cdc1
in the Golgi apparatus may serve as a quality assurance signal for GPI-anchored proteins.
4. Quality Control of GPI-Anchored Proteins
In yeast, a mutant version of Gas1, β-1,3-glucanosyltransferase, which is referred to as
Gas1*, is a well-studied model substrate for the quality control of misfolded GPI-anchored
proteins. Gas1* contains a point mutation (G291R) that renders the protein misfolded
and leads to its degradation [88,89]. Accumulating evidence suggests that Gas1* is not an
efficient ERAD substrate. Only a small fraction is delivered to the ERAD pathway, while the
vast majority of proteins escape Hrd1-dependent degradation; however, they are rapidly
recognized by the p24 complex, including Emp24, exported from the ER, and delivered to
the vacuole for degradation. p24 family proteins are conserved transmembrane proteins of
~24 kDa that function as cargo receptors for GPI-anchored proteins. The budding yeast p24
family is composed of three p24α (Erp1, Erp5, Erp6), one p24β (Emp24), three p24γ (Erp2,
Erp3, Erp4), and one p24δ (Erv25) [90]. These are single-transmembrane proteins with a
short (10–20 amino acids) C-terminal cytoplasmic tail. The cytoplasmic tail can interact
with both COPI (Coat Protein I) and COPII (Coat Protein II) subunits. The luminal portion
of these proteins may contribute to the formation of p24 oligomer and also participate in the
cargo recognition. In mammalian cells, the misfolded version of the prion protein (PrP*),
which is a GPI-anchored protein, is not degraded by ERAD but rather exported from the
ER despite the misfolding [91]. PrP* is recognized by the p24 complex and delivered to the
plasma membrane, from where it is transported to the lysosome for degradation [92]. When
the ER is loaded with high amounts of misfolded proteins and its capacity is saturated
during ER stress, misfolded PrP* dissociates from resident ER chaperones and is rapidly
released into the secretory pathway in a process called rapid ER stress-induced export
(RESET) [92]. This response is faster than the activation of the UPR and reduces the load of
aberrant proteins in the ER, thereby maintaining protein homeostasis in the ER.
Then, what is the difference between misfolded GPI-anchored proteins and general
ERAD substrates? When GPI anchor attachment is impaired, the misfolded protein moiety,
which is free from the ER membrane, is rarely targeted to the vacuole. Instead, a consider-
able portion of these proteins is delivered to the ERAD pathway. The protein moiety of
Gas1* is delivered to the Hrd1-dependent degradation pathway. In mammals, when GPI
anchor attachment is prevented, PrP* is targeted for ERAD. Therefore, the protein moiety of
the misfolded GPI-anchored proteins can be disposed by ERAD. It is possible that the GPI
anchor sterically obstructs step(s) during ERAD, including retro-translocation and/or other
steps. However, when GPI anchor remodeling proceeds incorrectly in cells lacking Bst1,
Cwh43, or Ted1, GPI-anchored proteins are not efficiently exported from the ER; instead,
they are retained in the ER and delivered to the Hrd1-dependent ERAD pathway [93].
This observation supports the idea that a GPI anchor does not pose a steric obstruction to
the ERAD of misfolded GPI-anchored proteins. The ER retention time of misfolded GPI-
anchored proteins may be determined by the remodeling status of the GPI anchor, which isInt. J. Mol. Sci. 2021, 22, 1061 6 of 14
directly coupled to ER export. To ensure the correct sequential synthesis of GPI-anchored
proteins, it would be beneficial to remove those possessing an immature GPI anchor from
the ER. Thus, the quality control of GPI-anchored proteins may be more severe than that of
normal secretory proteins. This idea is further supported by the recent observation that
Cdc1, a remodeling enzyme for GPI-anchored proteins, resides in the cis/medial Golgi
apparatus, where additional quality control systems might be required [94]. In addition, a
recent study showed that PrP* is trafficked from the ER to lysosomes in a complex with
ER-derived chaperones, including calnexin and cargo receptors [95]. These interaction
partners are critical for rapid endocytosis. Resident ER factors not only protect misfolded
GPI-anchored proteins from aggregation during trafficking, but also ensure that they are
subject to quality control at the plasma membrane and endocytosis to lysosomes.
5. Potential Physiological Relevance of ERAD to the Biosynthesis of
GPI-Anchored Proteins
As mentioned above, emerging evidence suggests that ERAD functions as a fail-
safe mechanism for the degradation of misfolded GPI-anchored proteins when the vac-
uole/lysosomal route is impaired. However, several observations support the physiological
relevance of ERAD to the biosynthesis of GPI-anchored proteins.
5.1. Genetic Interactions between GPI and ERAD Genes
Systematic studies in yeast have suggested several positive and negative genetic
interactions between genes encoding ERAD components and GPI biosynthetic factors [96].
Negative genetic interactions refer to double mutants that exhibit a more severe fitness
defect than expected [97]. Conversely, positive genetic interactions refer to double mutants
with a less severe growth defect than anticipated. These genes may contain genes encoding
components of the same nonessential protein complex [97]. Genes required for relatively
the later steps of GPI biosynthesis tend to interact with ERAD genes. For example, HRD1
shows positive genetic interactions with GPI8, GPI10, GPI11, and GPI17. Similarly, HRD3
shows positive genetic interactions with GPI8, GPI10, and GPI17, and negative genetic
interactions with GPI19. UBC7 shows positive genetic interactions with GPI13, GPI16, and
GPI17 [96]. Other interactions between major ERAD components and factors involved
in the biosynthesis of GPI-anchored proteins are listed in Table 1. Although the reasons
for these genetic interactions are currently unknown, one possible explanation is that
endogenous substrates that accumulate upon ERAD deficiency positively or negatively
affect the GPI deletion phenotype.
Table 1. Positive and negative genetic interactions between genes encoding ERAD components and
GPI biosynthetic factors. N: negative genetic interactions; P: positive genetic interactions.
GPI2 GPI8 GPI10 GPI11 GPI13 GPI16 GPI17 GPI19
HRD1 P P P P
HRD3 P P P N
UBC7 P P P
USA1 P P P P
DER1 N P P
YOS9 P N N
DOA10 P
In mammals, genome-wide CRISPR-Cas9 (Clustered Regularly Interspaced Short
Palindromic Repeats/CRISPR-Associated Proteins 9) genetic screening suggests that dis-
ruption of HRD1 or several other ERAD components enhances GPI synthesis in GPI-
transamidase-deficient cells [98]. The proposed scenario is that cells use ERAD to suppress
GPI synthesis by degrading unknown protein factor(s) or endogenous substrate(s) that
normally enhance the biosynthesis of GPI. Disruption of ERAD may cause the accumu-Int. J. Mol. Sci. 2021, 22, 1061 7 of 14
lation of such factor(s), which can lead to increased free GPIs including its biosynthetic
intermediates as well as mature forms in GPI-transamidase-deficient cells [98].
5.2. Quality Control of Proteins that Harbor the GPI Anchoring Signal in the Cytosol
Molecular recognition events, including protein targeting to the organelle, are inher-
ently imperfect because of intrinsic limits on specific binding. Indeed, during protein
targeting to the ER, many secretory proteins fail to associate with the signal recognition
particle (SRP) and can be detected in the cytosol before their translocation [99,100]. Under
normal conditions in mammalian cells, the efficiency of ER translocation is not high: it
may range from 60% to 95% [101]. This implies that the cell must be equipped with a
quality control system to monitor a significant number of un-translocated proteins in the
cytosol. The existence of a surveillance system that targets SRP-independent substrates
for degradation is also likely. These would include a precursor form of GPI-anchored
proteins whose C-terminal signal is hydrophobic. In yeast, GPI-anchored proteins that are
not translocated to the ER are degraded on the cytosolic face of the ER. This degradation
pathway is termed prERAD (Pre-insertional ERAD) and relies on opposing forces of the
ubiquitin ligase Doa10 and the deubiquitinating enzyme Ubp1 [102].
5.3. Exit of GPI-Anchored Proteins from the ER Is Affected by the Perturbation of
Manganese Homeostasis
Studies of mammalian cells show that PGAP5, an ER membrane protein, has a metal-
containing phosphate esterase motif in the lumen and requires manganese ions (Mn2+ )
for its enzymatic activity [87]. PGAP5 catalyzes the removal of the second EtN-P after
GPI is transferred to the protein. The removal of EtN-P from GPI-glycan by PGAP5 is
required for the efficient transport of GPI-anchored proteins from the ER to the Golgi
apparatus. As mentioned above, S. cerevisiae has two putative homologs of PGAP5, Cdc1
and Ted1. Cdc1 is a Mn2+ -dependent enzyme that interacts with genes involved in GPI
fatty acid remodeling [85,103]. Cdc1 was recently shown to have mannose-ethanolamine
phosphate phosphodiesterase activity, and it is responsible for the removal of EtN-P from
Man1 [85]. Transport of Gas1 from the ER is delayed in ted1∆ cells [104,105], although it
is not currently clear whether Ted1 is a manganese-dependent enzyme. Perturbation of
manganese homeostasis by depletion of Spf1, a P5-type ATPase that regulates manganese
transport into the ER, causes the defective export of GPI-anchored proteins from the ER,
and their accumulation in the ER [106]. These observations in mammals and yeast suggest
that manganese homeostasis is critical for the biosynthesis of GPI-anchored proteins.
5.4. Possible Involvement of ERAD in the Maintenance of Manganese Homeostasis
I found a genetic interaction between ERAD and Pmr1, a P-type Ca2+ - and Mn2+ -
transporting ATPase that is localized in the Golgi membrane [107], in yeast. Pmr1 is a
prototypic member of the Ca2+ -ATPase family of transporting ATPases, which are found in
a variety of organisms, including fungi, Caenorhabditis elegans, Drosophila melanogaster, and
mammals [108–110]. Both the calcium and manganese ions that are transported by Pmr1
into the Golgi lumen enable proper processing and trafficking of proteins through the secre-
tory pathway [107,111–113]. While calcium is mainly required for protein trafficking [114],
manganese is an essential enzymatic co-factor for glycosyltransferases that catalyze protein
glycosylation in the secretory pathway [112,115]. Pmr1 transports excess cytosolic man-
ganese into the Golgi lumen and mediates its export from the cell via secretory pathway
vesicles. Thus, Pmr1 contributes to the cellular detoxification of manganese [115]. Yeast
cells with a deleted PMR1 gene (pmr1∆ cells) show a pleiotropic phenotype. For example,
cells lacking Pmr1 are sensitive to ethylene glycol-bis(β-aminoethyl ether)-N,N,N0 ,N0 -
tetraacetic acid (EGTA), an effective chelator of calcium, manganese, and other divalent
metal ions [107]. Depletion of Pmr1 results in the accumulation of manganese and calcium
in the cytoplasm as well as depletion of these ions from the Golgi apparatus. Moreover,
cells lacking Pmr1 display a defect in carboxy peptidase Y (CPY) trafficking [112] and a
defect in the degradation of CPY*, a typical ERAD-L substrate [116]. Defects in the humanYeast cells with a deleted PMR1 gene (pmr1∆ cells) show a pleiotropic phenotype. For
example, cells lacking Pmr1 are sensitive to ethylene glycol-bis(β-aminoethyl ether)-
N,N,N′,N′-tetraacetic acid (EGTA), an effective chelator of calcium, manganese, and other
divalent metal ions [107]. Depletion of Pmr1 results in the accumulation of manganese
Int. J. Mol. Sci. 2021, 22, 1061 and calcium in the cytoplasm as well as depletion of these ions from the Golgi apparatus. 8 of 14
Moreover, cells lacking Pmr1 display a defect in carboxy peptidase Y (CPY) trafficking
[112] and a defect in the degradation of CPY*, a typical ERAD-L substrate [116]. Defects
in the human
ortholog of PMR1,ortholog
ATP2C1,of PMR1, ATP2C1, are
are associated associated
with with Hailey–Hailey
Hailey–Hailey disease, an
disease, an autosomal
autosomalblistering
dominant dominant blistering
skin disorder skin disorder [108,117].
[108,117].
AAprevious
previouslarge-scale
large-scaleanalysis
analysissuggested
suggestedaagenetic
geneticinteraction betweenPMR1
interactionbetween PMR1and and
UBC7
UBC7[118].
[118]. To confirm this this observation,
observation,I Iconstructed ubc7∆,
constructedubc7∆, pmr1∆,
pmr1∆, andand ubc7∆pmr1∆
ubc7∆pmr1∆ mu-
mutant strains
tant strains andand analyzed
analyzed their
their growth
growth (Supplementary
(Supplementary Materials).
Materials). However,
However, double
double mu-
mutant cells
tant cells grewnormally
grew normallycompared
comparedwith with isogenic
isogenic wild-type
wild-type strains or or single
singlemutant
mutant
strains,
strains,atatleast
leastononyeast
yeastextract-peptone-dextrose
extract-peptone-dextrose(YPD) (YPD)medium
medium(Figure(Figure3).
3).Next,
Next,I Itested
tested
the EGTA sensitivity of these strains because pmr1∆
the EGTA sensitivity of these strains because pmr1∆ cells were previously showntotobe
cells were previously shown be
sensitive
sensitivetotoEGTA.
EGTA.Consistently,
Consistently, pmr1∆
pmr1∆ cells were
cells sensitive
were to EGTA,
sensitive to EGTA,whereas
whereascellscells
withwith
the
deleted UBC7UBC7
the deleted genegene
grewgrewsimilarly to wild-type
similarly cells (Figure
to wild-type 3). Interestingly,
cells (Figure I found
3). Interestingly, that
I found
the
thatdeletion of UBC7
the deletion suppressed
of UBC7 the EGTA
suppressed sensitivity
the EGTA of pmr1∆
sensitivity cells,cells,
of pmr1∆ indicating that the
indicating that
deletion of UBC7 suppresses the EGTA sensitivity of pmr1∆ cells. To
the deletion of UBC7 suppresses the EGTA sensitivity of pmr1∆ cells. To further confirm further confirm this
result, I analyzed
this result, the genetic
I analyzed interactions
the genetic between
interactions PMR1PMR1
between and other ERADERAD
and other components.
compo-
As in Figure 3, the EGTA sensitivity of pmr1∆
nents. As shown in Figure 3, the EGTA sensitivity of pmr1∆ cells was rescued depletion
shown cells was rescued by the by the de-
of Hrd1, of
pletion a retro-translocation channel for
Hrd1, a retro-translocation luminalfor
channel ERAD substrates.
luminal A similar trend
ERAD substrates. was
A similar
observed for Hrd3, a component of the Hrd1 complex, and for
trend was observed for Hrd3, a component of the Hrd1 complex, and for Doa10, a mem- Doa10, a membrane E3
ligase
branethat targets
E3 ligase misfolded
that membrane
targets misfolded proteins with
membrane cytoplasmic
proteins lesions. These
with cytoplasmic results
lesions. These
suggest a potential physiological role of ERAD in maintaining calcium
results suggest a potential physiological role of ERAD in maintaining calcium and man- and manganese
homeostasis in the Golgi
ganese homeostasis in theapparatus.
Golgi apparatus.
Figure3.
Figure 3. Deletion
Deletionofofmajor
majorERAD
ERADcomponents
componentssuppresses the EGTA
suppresses the EGTA(Ethylene glycol-bis(β-ami-
(Ethylene glycol-bis(β-
noethyl ether)-N,N,N′,N′-tetra-acetic acid) sensitivity of pmr1∆ cells. Serial 10-fold dilutions
aminoethyl ether)-N,N,N0 ,N0 -tetra-acetic acid) sensitivity of pmr1∆ cells. Serial 10-fold of of
dilutions
yeast cultures were spotted onto yeast extract-peptone-dextrose medium (YPD) or YPD containing
yeast cultures were spotted onto yeast extract-peptone-dextrose medium (YPD) or YPD containing
2 mM EGTA. Plates were incubated for 3–7 days. The strains used here were BY4741 (wild-type)
2and
mMitsEGTA.
mutantPlates were incubated for 3–7 days. The strains used here were BY4741 (wild-type) and
derivatives.
its mutant derivatives.
The mechanism by which depletion of ERAD might rescue the EGTA-sensitive phe-
The mechanism by which depletion of ERAD might rescue the EGTA-sensitive pheno-
notype of pmr1∆ is currently unclear. One possible scenario is that a native luminal and/or
type of pmr1∆ is currently unclear. One possible scenario is that a native luminal and/or
membrane protein(s) accumulates upon ERAD deficiency and affects the pmr1∆ pheno-
membrane protein(s) accumulates upon ERAD deficiency and affects the pmr1∆ phenotype.
type. Furthermore, there may be many kinds of substrates that accumulate in ERAD-de-
Furthermore, there may be many kinds of substrates that accumulate in ERAD-defective
fective cells and affect the phenotype of pmr1∆ cells. One candidate is Cdc1, which sup-
cells and affect the phenotype of pmr1∆ cells. One candidate is Cdc1, which suppresses
presses
the EGTAthe EGTA sensitivity
sensitivity of pmr1∆of pmr1∆
cells whencells when overexpressed
overexpressed [85,119–121].
[85,119–121]. Consist-
Consistently, N-
ently, N-terminal hemagglutinin-tagged Cdc1 is stabilized in the Hrd3 mutant
terminal hemagglutinin-tagged Cdc1 is stabilized in the Hrd3 mutant [85]. However, [85]. How-
a
ever, a recent study showed that Cdc1 localizes to cis and medial Golgi when
recent study showed that Cdc1 localizes to cis and medial Golgi when tagged with the tagged with
the fluorescent
fluorescent mNeonmNeon protein
protein at or
at its N- itsC-terminus
N- or C-terminus [94], although
[94], although a previous
a previous study
study showed
that Cdc1 localizes to the ER when tagged with the myc-epitope at the N-terminus [120].
The reason for these discrepancies is not clear; however, the removal of EtN-P from Man 1
would be a critical step for the quality control of GPI-anchored proteins before they are
delivered to the plasma membrane. Nonetheless, it will be essential to test the localization
and stability of a purely endogenous untagged version of Cdc1 to fully understand the
observed phenomena.Int. J. Mol. Sci. 2021, 22, 1061 9 of 14
6. Conclusions
Unidentified endogenous ERAD substrates are key to understanding the potential
relationship between ERAD and the biosynthesis of GPI-anchored proteins. The simple
genetic interaction reported by systematic studies implies the existence of an endogenous
ERAD substrate that may affect the phenotype of cells defective in GPI biosynthesis.
Mammals may express a putative positive regulator of GPI biosynthesis whose stability is
regulated by ERAD. The rescue of the EGTA sensitivity of pmr1∆ cells by deletion of ERAD
components could also be explained by the accumulation of endogenous substrates. This
suggests that ERAD could control manganese homeostasis, which is critical for the ER exit
of GPI-anchored proteins. Current data indicate that the role of ERAD is not limited to the
degradation of misfolded proteins and may play a critical role in the regulation of cellular
phenomena, even under normal growth conditions. The identification of an endogenous
substrate of Hrd1, Doa10, as well as the dedicated ubiquitin ligases in mammals [15],
would be important to fully understand the physiological roles of ERAD.
Supplementary Materials: Supplementary materials can be found at https://www.mdpi.com/1422
-0067/22/3/1061/s1.
Funding: This work was supported by the Toray Science Foundation, the Toyoaki Scholarship
Foundation, and JSPS KAKENHI, Grant Numbers 15K18503, 18K19306, and 19H02923.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: Data is contained within the article or supplementary material.
Acknowledgments: I would like to thank Takumi Kamura and Maiko Shimizu for the initial study
of this project.
Conflicts of Interest: The author declares no conflict of interest.
Abbreviations
ERAD Endoplasmic reticulum-associated degradation
EGTA Ethylene glycol-bis(β-aminoethyl ether)-N,N,N 0 ,N 0 -tetra-acetic acid
GPI Glycosylphosphatidylinositol
EtN-P Ethanolamine phosphate
Man Mannose
GlcN Glucosamine
GlcNAc N-Acetylglucosamine
UPR Unfolded protein response
prERAD Pre-insertional ERAD
References
1. Rüdiger, S.; Buchberger, A.; Bukau, B. Interaction of Hsp70 chaperones with substrates. Nat. Struct. Mol. Biol. 1997, 4, 342–349.
[CrossRef] [PubMed]
2. Rutkowski, D.; Kaufman, R.J. A trip to the ER: Coping with stress. Trends Cell Biol. 2004, 14, 20–28. [CrossRef] [PubMed]
3. Ron, D.; Walter, P. Signal integration in the endoplasmic reticulum unfolded protein response. Nat. Rev. Mol. Cell Biol. 2007, 8,
519–529. [CrossRef] [PubMed]
4. Vembar, S.S.; Brodsky, J.L. One step at a time: Endoplasmic reticulum-associated degradation. Nat. Rev. Mol. Cell Biol. 2008, 9,
944–957. [CrossRef]
5. Smith, M.H.; Ploegh, H.L.; Weissman, J.S. Road to Ruin: Targeting Proteins for Degradation in the Endoplasmic Reticulum.
Science 2011, 334, 1086–1090. [CrossRef]
6. Ruggiano, A.; Foresti, O.; Carvalho, P. Quality control: ER-associated degradation: Protein quality control and beyond. J. Cell Biol.
2014, 204, 869–879. [CrossRef]
7. Oikonomou, C.; Hendershot, L.M. Disposing of misfolded ER proteins: A troubled substrate’s way out of the ER. Mol. Cell
Endocrinol. 2020, 500, 110630. [CrossRef]
8. Sun, Z.; Brodsky, J.L. Protein quality control in the secretory pathway. J. Cell Biol. 2019, 218, 3171–3187. [CrossRef]Int. J. Mol. Sci. 2021, 22, 1061 10 of 14
9. Mehrtash, A.B.; Hochstrasser, M. Ubiquitin-dependent protein degradation at the endoplasmic reticulum and nuclear envelope.
Semin. Cell Dev. Biol. 2019, 93, 111–124. [CrossRef]
10. Berner, N.; Reutter, K.-R.; Wolf, D.H. Protein Quality Control of the Endoplasmic Reticulum and Ubiquitin–Proteasome-Triggered
Degradation of Aberrant Proteins: Yeast Pioneers the Path. Annu. Rev. Biochem. 2018, 87, 751–782. [CrossRef]
11. Christianson, J.C.; Ye, Y. Cleaning up in the endoplasmic reticulum: Ubiquitin in charge. Nat. Struct. Mol. Biol. 2014, 21, 325–335.
[CrossRef] [PubMed]
12. Wu, X.; Rapoport, T.A. Mechanistic insights into ER-associated protein degradation. Curr. Opin. Cell Biol. 2018, 53, 22–28.
[CrossRef] [PubMed]
13. Nakatsukasa, K.; Brodsky, J.L. The Recognition and Retrotranslocation of Misfolded Proteins from the Endoplasmic Reticulum.
Traffic 2008, 9, 861–870. [CrossRef] [PubMed]
14. Nakatsukasa, K.; Okumura, F.; Kamura, T. Proteolytic regulation of metabolic enzymes by E3 ubiquitin ligase complexes: Lessons
from yeast. Crit. Rev. Biochem. Mol. Biol. 2015, 50, 489–502. [CrossRef] [PubMed]
15. Qi, L.; Tsai, B.; Arvan, P. New Insights into the Physiological Role of Endoplasmic Reticulum-Associated Degradation. Trends Cell
Biol. 2017, 27, 430–440. [CrossRef]
16. Adle, D.J.; Wei, W.; Smith, N.; Bies, J.J.; Lee, J. Cadmium-mediated rescue from ER-associated degradation induces expression of
its exporter. Proc. Natl. Acad. Sci. USA 2009, 106, 10189–10194. [CrossRef]
17. Hampton, R.Y.; Gardner, R.G.; Rine, J. Role of 26S proteasome and HRD genes in the degradation of 3-hydroxy-3-methylglutaryl-
CoA reductase, an integral endoplasmic reticulum membrane protein. Mol. Biol. Cell 1996, 7, 2029–2044. [CrossRef]
18. Rape, M.; Hoppe, T.; Gorr, I.; Kalocay, M.; Richly, H.; Jentsch, S. Mobilization of Processed, Membrane-Tethered SPT23
Transcription Factor by CDC48UFD1/NPL4, a Ubiquitin-Selective Chaperone. Cell 2001, 107, 667–677. [CrossRef]
19. Foresti, O.; Ruggiano, A.; Hannibal-Bach, H.K.; Ejsing, C.S.; Carvalho, P. Sterol homeostasis requires regulated degradation of
squalene monooxygenase by the ubiquitin ligase Doa10/Teb4. eLife 2013, 2, e00953. [CrossRef]
20. Wu, X.; Siggel, M.; Ovchinnikov, S.; Mi, W.; Svetlov, V.; Nudler, E.; Liao, M.; Hummer, G.; Rapoport, T.A. Structural basis of
ER-associated protein degradation mediated by the Hrd1 ubiquitin ligase complex. Science 2020, 368, eaaz2449. [CrossRef]
21. Knop, M.; Finger, A.; Braun, T.; Hellmuth, K.; Wolf, D.H. Der1, a novel protein specifically required for endoplasmic reticulum
degradation in yeast. EMBO J. 1996, 15, 753–763. [CrossRef] [PubMed]
22. Bordallo, J.; Plemper, R.K.; Finger, A.; Wolf, D.H. Der3p/Hrd1p Is Required for Endoplasmic Reticulum-associated Degradation
of Misfolded Lumenal and Integral Membrane Proteins. Mol. Biol. Cell 1998, 9, 209–222. [CrossRef] [PubMed]
23. Bays, N.W.; Gardner, R.G.; Seelig, L.P.; Joazeiro, C.A.; Hampton, R.Y. Hrd1p/Der3p is a membrane-anchored ubiquitin ligase
required for ER-associated degradation. Nat. Cell Biol. 2001, 3, 24–29. [CrossRef] [PubMed]
24. Gauss, R.; Jarosch, E.; Sommer, T.; Hirsch, C. A complex of Yos9p and the HRD ligase integrates endoplasmic reticulum quality
control into the degradation machinery. Nat. Cell Biol. 2006, 8, 849–854. [CrossRef]
25. Carvalho, P.; Goder, V.; Rapoport, T.A. Distinct Ubiquitin-Ligase Complexes Define Convergent Pathways for the Degradation of
ER Proteins. Cell 2006, 126, 361–373. [CrossRef]
26. Kanehara, K.; Xie, W.; Ng, D.T.W. Modularity of the Hrd1 ERAD complex underlies its diverse client range. J. Cell Biol. 2010, 188,
707–716. [CrossRef]
27. Mehnert, M.; Sommer, T.; Jarosch, E. Der1 promotes movement of misfolded proteins through the endoplasmic reticulum
membrane. Nat. Cell Biol. 2014, 16, 77–86. [CrossRef]
28. Gauss, R.; Sommer, T.; Jarosch, E. The Hrd1p ligase complex forms a linchpin between ER-lumenal substrate selection and Cdc48p
recruitment. EMBO J. 2006, 25, 1827–1835. [CrossRef]
29. Bhamidipati, A.; Denic, V.; Quan, E.M.; Weissman, J.S. Exploration of the Topological Requirements of ERAD Identifies Yos9p as
a Lectin Sensor of Misfolded Glycoproteins in the ER Lumen. Mol. Cell 2005, 19, 741–751. [CrossRef]
30. Szathmary, R.; Bielmann, R.; Nita-Lazar, M.; Burda, P.; Jakob, C.A. Yos9 Protein Is Essential for Degradation of Misfolded
Glycoproteins and May Function as Lectin in ERAD. Mol. Cell 2005, 19, 765–775. [CrossRef]
31. Quan, E.M.; Kamiya, Y.; Kamiya, D.; Denic, V.; Weibezahn, J.; Kato, K.; Weissman, J.S. Defining the Glycan Destruction Signal for
Endoplasmic Reticulum-Associated Degradation. Mol. Cell 2008, 32, 870–877. [CrossRef] [PubMed]
32. Jakob, C.A.; Bodmer, D.; Spirig, U.; Bättig, P.; Marcil, A.; Dignard, D.; Bergeron, J.J.; Thomas, D.Y.; Aebi, M. Htm1p, a mannosidase-
like protein, is involved in glycoprotein degradation in yeast. EMBO Rep. 2001, 2, 423–430. [CrossRef] [PubMed]
33. Nakatsukasa, K.; Nishikawa, S.-I.; Hosokawa, N.; Nagata, K.; Endo, T. Mnl1p, an α-Mannosidase-like Protein in YeastSaccha-
romyces cerevisiae, Is Required for Endoplasmic Reticulum-associated Degradation of Glycoproteins. J. Biol. Chem. 2001, 276,
8635–8638. [CrossRef] [PubMed]
34. Xie, W.; Kanehara, K.; Sayeed, A.; Ng, D.T.W. Intrinsic Conformational Determinants Signal Protein Misfolding to the Hrd1/Htm1
Endoplasmic Reticulum–associated Degradation System. Mol. Biol. Cell 2009, 20, 3317–3329. [CrossRef] [PubMed]
35. Vashistha, N.; Neal, S.E.; Singh, A.; Carroll, S.M.; Hampton, R.Y. Direct and essential function for Hrd3 in ER-associated
degradation. Proc. Natl. Acad. Sci. USA 2016, 113, 5934–5939. [CrossRef]
36. Horn, S.C.; Hanna, J.; Hirsch, C.; Volkwein, C.; Schütz, A.; Heinemann, U.; Sommer, T.; Jarosch, E. Usa1 Functions as a Scaffold of
the HRD-Ubiquitin Ligase. Mol. Cell 2009, 36, 782–793. [CrossRef]
37. Biederer, T.; Volkwein, C.; Sommer, T. Role of Cue1p in Ubiquitination and Degradation at the ER Surface. Science 1997, 278,
1806–1809. [CrossRef]Int. J. Mol. Sci. 2021, 22, 1061 11 of 14
38. Schuberth, C.; Buchberger, A. Membrane-bound Ubx2 recruits Cdc48 to ubiquitin ligases and their substrates to ensure efficient
ER-associated protein degradation. Nat. Cell Biol. 2005, 7, 999–1006. [CrossRef]
39. Neuber, O.; Jarosch, E.; Volkwein, C.; Walter, J.; Sommer, T. Ubx2 links the Cdc48 complex to ER-associated protein degradation.
Nat. Cell Biol. 2005, 7, 993–998. [CrossRef]
40. Nakatsukasa, K.; Brodsky, J.L.; Kamura, T. A stalled retrotranslocation complex reveals physical linkage between substrate
recognition and proteasomal degradation during ER-associated degradation. Mol. Biol. Cell 2013, 24, 1765–1775. [CrossRef]
41. Sato, B.K.; Schulz, D.; Do, P.H.; Hampton, R.Y. Misfolded Membrane Proteins Are Specifically Recognized by the Transmembrane
Domain of the Hrd1p Ubiquitin Ligase. Mol. Cell 2009, 34, 212–222. [CrossRef] [PubMed]
42. Neal, S.; Jaeger, P.A.; Duttke, S.H.; Benner, C.; Glass, C.K.; Ideker, T.; Hampton, R.Y. The Dfm1 Derlin Is Required for ERAD
Retrotranslocation of Integral Membrane Proteins. Mol. Cell 2018, 69, 306–320.e4. [CrossRef] [PubMed]
43. Neal, S.E.; Syau, D.; Nejatfard, A.; Nadeau, S.; Hampton, R.Y. HRD Complex Self-Remodeling Enables a Novel Route of
Membrane Protein Retrotranslocation. iScience 2020, 23, 101493. [CrossRef] [PubMed]
44. Kreft, S.G.; Wang, L.; Hochstrasser, M. Membrane Topology of the Yeast Endoplasmic Reticulum-localized Ubiquitin Ligase
Doa10 and Comparison with Its Human Ortholog TEB4 (MARCH-VI). J. Biol. Chem. 2006, 281, 4646–4653. [CrossRef]
45. Huyer, G.; Piluek, W.F.; Fansler, Z.; Kreft, S.G.; Hochstrasser, M.; Brodsky, J.L.; Michaelis, S. Distinct machinery is required in
Saccharomyces cerevisiae for the endoplasmic reticulum-associated degradation of a multispanning membrane protein and a
soluble luminal protein. J. Biol. Chem. 2004, 279, 38369–38378. [CrossRef]
46. Gnann, A.; Riordan, J.R.; Wolf, D.H. Cystic Fibrosis Transmembrane Conductance Regulator Degradation Depends on the Lectins
Htm1p/EDEM and the Cdc48 Protein Complex in Yeast. Mol. Biol. Cell 2004, 15, 4125–4135. [CrossRef]
47. Ravid, T.; Kreft, S.G.; Hochstrasser, M. Membrane and soluble substrates of the Doa10 ubiquitin ligase are degraded by distinct
pathways. EMBO J. 2006, 25, 533–543. [CrossRef]
48. Vashist, S.; Ng, D.T. Misfolded proteins are sorted by a sequential checkpoint mechanism of ER quality control. J. Cell Biol. 2004,
165, 41–52. [CrossRef]
49. Furth, N.; Gertman, O.; Shiber, A.; Alfassy, O.S.; Cohen, I.; Rosenberg, M.M.; Doron, N.K.; Friedler, A.; Ravid, T. Exposure of
bipartite hydrophobic signal triggers nuclear quality control of Ndc10 at the endoplasmic reticulum/nuclear envelope. Mol. Biol.
Cell 2011, 22, 4726–4739. [CrossRef]
50. Swanson, R.; Locher, M.; Hochstrasser, M. A conserved ubiquitin ligase of the nuclear envelope/endoplasmic reticulum that
functions in both ER-associated and Matalpha2 repressor degradation. Genes Dev. 2001, 15, 2660–2674. [CrossRef]
51. Habeck, G.; Ebner, F.A.; Shimada-Kreft, H.; Kreft, S.G. The yeast ERAD-C ubiquitin ligase Doa10 recognizes an intramembrane
degron. J. Cell Biol. 2015, 209, 261–273. [CrossRef] [PubMed]
52. Schmidt, C.C.; Vasic, V.; Stein, A. Doa10 is a membrane protein retrotranslocase in ER-associated protein degradation. eLife 2020,
9, e56945. [CrossRef] [PubMed]
53. Nakatsukasa, K.; Huyer, G.; Michaelis, S.; Brodsky, J.L. Dissecting the ER-Associated Degradation of a Misfolded Polytopic
Membrane Protein. Cell 2008, 132, 101–112. [CrossRef] [PubMed]
54. Han, S.; Liu, Y.; Chang, A. Cytoplasmic Hsp70 Promotes Ubiquitination for Endoplasmic Reticulum-associated Degradation of a
Misfolded Mutant of the Yeast Plasma Membrane ATPase, PMA1. J. Biol. Chem. 2007, 282, 26140–26149. [CrossRef]
55. Metzger, M.B.; Maurer, M.J.; Dancy, B.M.; Michaelis, S. Degradation of a Cytosolic Protein Requires Endoplasmic Reticulum-
associated Degradation Machinery. J. Biol. Chem. 2008, 283, 32302–32316. [CrossRef]
56. Weber, A.; Cohen, I.; Popp, O.; Dittmar, G.; Reiss, Y.; Sommer, T.; Ravid, T.; Jarosch, E. Sequential Poly-ubiquitylation by
Specialized Conjugating Enzymes Expands the Versatility of a Quality Control Ubiquitin Ligase. Mol. Cell 2016, 63, 827–839.
[CrossRef]
57. Nakatsukasa, K.; Kamura, T. Subcellular Fractionation Analysis of the Extraction of Ubiquitinated Polytopic Membrane Substrate
during ER-Associated Degradation. PLoS ONE 2016, 11, e0148327. [CrossRef]
58. Walter, J.; Urban, J.; Volkwein, C.; Sommer, T. Sec61p-independent degradation of the tail-anchored ER membrane protein Ubc6p.
EMBO J. 2001, 20, 3124–3131. [CrossRef]
59. Pittet, M.; Conzelmann, A. Biosynthesis and function of GPI proteins in the yeast Saccharomyces cerevisiae. Biochim. Et Biophys.
Acta (BBA)-Mol. Cell Biol. Lipids 2007, 1771, 405–420. [CrossRef]
60. Leidich, S.D.; Kostova, Z.; Latek, R.R.; Costello, L.C.; Drapp, D.A.; Gray, W.; Fassler, J.S.; Orlean, P. Temperature-sensitive Yeast
GPI Anchoring Mutants gpi2 and gpi3 Are Defective in the Synthesis of N-Acetylglucosaminyl Phosphatidylinositol. J. Biol.
Chem. 1995, 270, 13029–13035. [CrossRef]
61. Yan, B.C.; Westfall, B.A.; Orlean, P. Ynl038wp (Gpi15p) is theSaccharomyces cerevisiae homologue of human Pig-Hp and
participates in the first step in glycosylphosphatidylinositol assembly. Yeast 2001, 18, 1383–1389. [CrossRef] [PubMed]
62. Newman, H.A.; Romeo, M.J.; Lewis, S.E.; Yan, B.C.; Orlean, P.; Levin, D.E. Gpi19, the Saccharomyces cerevisiae Homologue of
Mammalian PIG-P, Is a Subunit of the Initial Enzyme for Glycosylphosphatidylinositol Anchor Biosynthesis. Eukaryot. Cell 2005,
4, 1801–1807. [CrossRef] [PubMed]
63. Orlean, P.; Menon, A.K. Thematic review series: Lipid Posttranslational Modifications.GPI anchoring of protein in yeast and
mammalian cells, or: How we learned to stop worrying and love glycophospholipids. J. Lipid Res. 2007, 48, 993–1011. [CrossRef]
[PubMed]Int. J. Mol. Sci. 2021, 22, 1061 12 of 14
64. Maeda, Y.; Watanabe, R.; Harris, C.L.; Hong, Y.; Ohishi, K.; Kinoshita, K.; Kinoshita, T. PIG-M transfers the first mannose to
glycosylphosphatidylinositol on the lumenal side of the ER. EMBO J. 2001, 20, 250–261. [CrossRef]
65. Kang, J.Y.; Hong, Y.; Ashida, H.; Shishioh, N.; Murakami, Y.; Morita, Y.S.; Maeda, Y.; Kinoshita, T. PIG-V Involved in Transferring
the Second Mannose in Glycosylphosphatidylinositol. J. Biol. Chem. 2005, 280, 9489–9497. [CrossRef]
66. Gaynor, E.C.; Mondésert, G.; Grimme, S.J.; Reed, S.I.; Orlean, P.; Emr, S.D. MCD4Encodes a Conserved Endoplasmic Reticulum
Membrane Protein Essential for Glycosylphosphatidylinositol Anchor Synthesis in Yeast. Mol. Biol. Cell 1999, 10, 627–648.
[CrossRef]
67. Sütterlin, C.; Escribano, M.V.; Gerold, P.; Maeda, Y.; Mazón, M.J.; Kinoshita, T.; Schwarz, R.T.; Riezman, H. Saccharomyces
cerevisiae GPI10, the functional homologue of human PIG-B, is required for glycosylphosphatidylinositol-anchor synthesis.
Biochem. J. 1998, 332, 153–159. [CrossRef]
68. Flury, I.; Benachour, A.; Conzelmann, A. YLL031c Belongs to a Novel Family of Membrane Proteins Involved in the Transfer
of Ethanolaminephosphate onto the Core Structure of Glycosylphosphatidylinositol Anchors in Yeast. J. Biol. Chem. 2000, 275,
24458–24465. [CrossRef]
69. Grimme, S.J.; Westfall, B.A.; Wiedman, J.M.; Taron, C.H.; Orlean, P. The Essential Smp3 Protein Is Required for Addition of the
Side-branching Fourth Mannose during Assembly of Yeast Glycosylphosphatidylinositols. J. Biol. Chem. 2001, 276, 27731–27739.
[CrossRef]
70. Benachour, A.; Sipos, G.; Flury, I.; Reggiori, F.; Canivenc-Gansel, E.; Vionnet, C.; Conzelmann, A.; Benghezal, M. Deletion of GPI7,
a Yeast Gene Required for Addition of a Side Chain to the Glycosylphosphatidylinositol (GPI) Core Structure, Affects GPI Protein
Transport, Remodeling, and Cell Wall Integrity. J. Biol. Chem. 1999, 274, 15251–15261. [CrossRef]
71. Imhof, I.; Flury, I.; Vionnet, C.; Roubaty, C.; Egger, D.; Conzelmann, A. Glycosylphosphatidylinositol (GPI) Proteins ofSaccha-
romyces cerevisiaeContain Ethanolamine Phosphate Groups on the α1,4-linked Mannose of the GPI Anchor. J. Biol. Chem. 2004,
279, 19614–19627. [CrossRef] [PubMed]
72. Chen, R.; Knez, J.J.; Merrick, W.C.; Medof, M.E. Comparative efficiencies of C-terminal signals of native glycophosphatidylinositol
(GPI)-anchored proproteins in conferring GPI-anchoring. J. Cell. Biochem. 2002, 84, 68–83. [CrossRef] [PubMed]
73. Ohishi, K.; Inoue, N.; Kinoshita, T. PIG-S and PIG-T, essential for GPI anchor attachment to proteins, form a complex with GAA1
and GPI8. EMBO J. 2001, 20, 4088–4098. [CrossRef]
74. Benghezal, M.; Benachour, A.; Rusconi, S.; Aebi, M.; Conzelmann, A. Yeast Gpi8p is essential for GPI anchor attachment onto
proteins. EMBO J. 1996, 15, 6575–6583. [CrossRef] [PubMed]
75. Fraering, P.; Imhof, I.; Meyer, U.; Strub, J.-M.; Van Dorsselaer, A.; Vionnet, C.; Conzelmann, A. The GPI Transamidase Complex of
Saccharomyces cerevisiae Contains Gaa1p, Gpi8p, and Gpi16p. Mol. Biol. Cell 2001, 12, 3295–3306. [CrossRef]
76. Hamburger, D.; Egerton, M.; Riezman, H. Yeast Gaa1p is required for attachment of a completed GPI anchor onto proteins. J. Cell
Biol. 1995, 129, 629–639. [CrossRef]
77. Hong, Y.; Ohishi, K.; Kang, J.Y.; Tanaka, S.; Inoue, N.; Nishimura, J.-I.; Maeda, Y.; Kinoshita, T. Human PIG-U and Yeast Cdc91p
Are the Fifth Subunit of GPI Transamidase That Attaches GPI-Anchors to Proteins. Mol. Biol. Cell 2003, 14, 1780–1789. [CrossRef]
78. Meyer, U.; Benghezal, M.; Imhof, I.; Conzelmann, A. Active Site Determination of Gpi8p, a Caspase-Related Enzyme Required for
Glycosylphosphatidylinositol Anchor Addition to Proteins†. Biochemistry 2000, 39, 3461–3471. [CrossRef]
79. Ohishi, K.; Inoue, N.; Maeda, Y.; Takeda, J.; Riezman, H.; Kinoshita, T. Gaa1p and Gpi8p Are Components of a Glycosylphos-
phatidylinositol (GPI) Transamidase That Mediates Attachment of GPI to Proteins. Mol. Biol. Cell 2000, 11, 1523–1533. [CrossRef]
80. Kang, X.; Szallies, A.; Rawer, M.; Echner, H.; Duszenko, M. GPI anchor transamidase of Trypanosoma brucei: In vitro assay of the
recombinant protein and VSG anchor exchange. J. Cell Sci. 2002, 115, 2529–2539.
81. Tanaka, S.; Maeda, Y.; Tashima, Y.; Kinoshita, T. Inositol Deacylation of Glycosylphosphatidylinositol-anchored Proteins Is
Mediated by Mammalian PGAP1 and Yeast Bst1p. J. Biol. Chem. 2004, 279, 14256–14263. [CrossRef] [PubMed]
82. Fujita, M.; Umemura, M.; Yoko-o, T.; Jigami, Y. PER1 Is Required for GPI-Phospholipase A2 Activity and Involved in Lipid
Remodeling of GPI-anchored Proteins. Mol. Biol. Cell 2006, 17, 5253–5264. [CrossRef] [PubMed]
83. Bosson, R.; Jaquenoud, M.; Conzelmann, A. GUP1 of Saccharomyces cerevisiae Encodes an O-Acyltransferase Involved in
Remodeling of the GPI Anchor. Mol. Biol. Cell 2006, 17, 2636–2645. [CrossRef] [PubMed]
84. Umemura, M.; Fujita, M.; Yoko-o, T.; Fukamizu, A.; Jigami, Y. Saccharomyces cerevisiae CWH43Is Involved in the Remodeling of
the Lipid Moiety of GPI Anchors to Ceramides. Mol. Biol. Cell 2007, 18, 4304–4316. [CrossRef]
85. Vazquez, H.M.; Vionnet, C.; Roubaty, C.; Conzelmann, A. Cdc1 removes the ethanolamine phosphate of the first mannose of
GPI anchors and thereby facilitates the integration of GPI proteins into the yeast cell wall. Mol. Biol. Cell 2014, 25, 3375–3388.
[CrossRef] [PubMed]
86. Manzano-Lopez, J.; Perez-Linero, A.M.; Aguilera-Romero, A.; Martin, M.E.; Okano, T.; Silva, D.V.; Seeberger, P.H.; Riezman, H.;
Funato, K.; Goder, V.; et al. COPII Coat Composition Is Actively Regulated by Luminal Cargo Maturation. Curr. Biol. 2015, 25,
152–162. [CrossRef]
87. Fujita, M.; Maeda, Y.; Ra, M.; Yamaguchi, Y.; Taguchi, R.; Kinoshita, T. GPI Glycan Remodeling by PGAP5 Regulates Transport of
GPI-Anchored Proteins from the ER to the Golgi. Cell 2009, 139, 352–365. [CrossRef]
88. Fujita, M.; Yoko-o, T.; Jigami, Y. Inositol Deacylation by Bst1p Is Required for the Quality Control of Glycosylphosphatidylinositol-
anchored Proteins. Mol. Biol. Cell 2006, 17, 834–850. [CrossRef]You can also read

















































