A Novel Stilbene-Like Compound That Reduces Melanin through Inhibiting Melanocyte Differentiation and Proliferation without Inhibiting Tyrosinase ...
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
cosmetics
Article
A Novel Stilbene-Like Compound That Reduces
Melanin through Inhibiting Melanocyte
Differentiation and Proliferation without
Inhibiting Tyrosinase
Kristy Martinson 1 , Noah Stueven 1 , Aaron Monte 2 and Cheng-chen Huang 1, *
1 Biology Department, University of Wisconsin-River Falls, River Falls, WI 54022, USA;
kjmartinson@wisc.edu (K.M.); noah.stueven@my.uwrf.edu (N.S.)
2 Department of Chemistry and Biochemistry, University of Wisconsin-La Crosse, La Crosse, WI 54601, USA;
amonte@uwlax.edu
* Correspondence: cheng-chen.huang@uwrf.edu; Tel.: +715-425-4276; Fax: 715-425-0378
Received: 24 May 2018; Accepted: 11 July 2018; Published: 19 July 2018
Abstract: Cosmetic practices that use skin-lightening agents to obtain desired skin tones or treat
pigment abnormalities have been popular worldwide. However, the molecular and cellular
mechanisms of these agents are still largely unknown. Here we identified a family of compounds,
with the lead compound named A11, that exhibited strong pigment reduction in developing zebrafish
embryos. The pigment inhibition lasted for several days and is effective both before and after
melanogenesis. By comparison with several known skin-lightening compounds, A11 appeared to be
more potent and caused slower pigment recovery after withdrawal. A11, however, did not inhibit
tyrosinase or cause apoptosis in melanocytes. We further found that A11 suppressed proliferation
in melanocytes and reduced the number of differentiated melanocytes by activating MAPK
(mitogen-activated protein kinase) and Akt. Finally, A11 also caused melanin reduction in mammalian
melanocytes. Together, A11 might be a potent skin-lightening agent with novel mechanisms.
Keywords: skin-lightening; zebrafish; tyrosinase; melanocyte; proliferation; differentiation
1. Introduction
The desire to manipulate skin color has long been observed in human history. It is well known that
melanin, the pigment molecule that determines our skin color, plays an important role in absorbing
high energy waves, such as ultraviolet light, to protect molecules like DNA from damages induced by
the energy wave. Damages can include abnormal chemical bonding formation and DNA breaks which
can eventually lead to cancer formation. It is well documented that people with fair skin color appear
to have a significantly higher risk of skin cancer. This explains why darker (or tanned) skin is believed
to be healthier and preferred in Caucasians. In contrast, people in African and Asian countries prefer a
lighter skin color because it is commonly associated with higher social status. Therefore, while the
sunbath and tanning industry is popular in western countries, skin-lightening products and practices
are sought after in Asia.
Skin color can easily and quickly be changed due to the high turnover rate of skin cells and
the simple enzymatic pathway for melanin synthesis. Keratinocytes make up approximately 90%
of our epidermis and are responsible for even skin tone. Keratinocytes are constantly exposed to
environmental toxins and damages causing a high turnover rate. It is estimated that the turnover rate
ranges from two weeks in infants to one month for people in their thirties. Melanin pigment molecules
are synthesized in melanocytes, which only make up 1% of epidermis, and later are transported
Cosmetics 2018, 5, 45; doi:10.3390/cosmetics5030045 www.mdpi.com/journal/cosmetics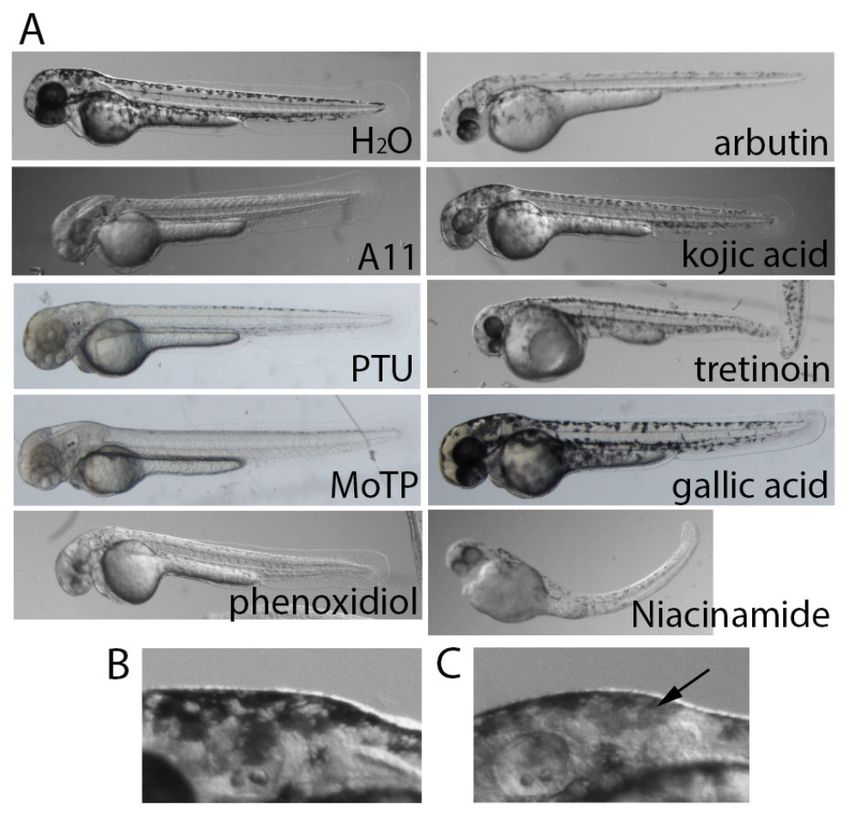
Cosmetics 2018, 5, 45 2 of 14
to keratinocytes. The biochemical synthesis pathway for melanin starts with tyrosinse amino acid
catalyzed by tyrosinase into β-3,4-dihydroxyphenylalanine (DOPA) and later into DOPAquinone.
Thus tyrosinase is the key enzyme of melanin synthesis. There are many naturally found tyrosinase
inhibitors that can effectively reduce pigment both in vitro and in vivo. For example, kojic acid
isolated from Aspergillus and Penicillum species is a strong tyrosinase inhibitor that chelates copper
which is a cofactor of tyrosinase (Battaini et al. [1]). Arbutin, derived from the fruit of the California
buckeye, is another strong tyrosinase inhibitor while aloesin, a natural derivative from aloe vera, acts
as a competitive inhibitor of tyrosinase (Jones et al. [2]). Some other pigment reduction reagents
were found to function during melanin transport or in regulating melanocyte survival (Reviewed
in Kim et al. [3]). Because of the high turnover rate of keratinocytes, topical application of these
reagents can easily generate lighter skin color within a few weeks. However, with the understanding of
melanogenesis mechanisms, it is also possible to increase skin pigmentation (Brenner and Hearing [4]).
Adverse effects of skin-lightening agents have been reported, although it is commonly considered
safe toward ectopically applied compounds. Hydroquinone, which was the very first skin-lightening
compound, is a strong tyrosinase inhibitor. Its specificity didn’t raise any concern when it was first used
on humans, but later was found to cause cytotoxicity in melanocytes, DNA damages, and likely cancers
(Eastmond et al. [5]). Derivatives of hydroquinone, the natural form called ® -arbutin and the synthetic
form α-arbutin, exhibit strong tyrosinase inhibition but with no detectable cytotoxicity. No detailed
toxicity study has been conducted for arbutin derivatives. Kojic acid, another commonly used
skin-lightening agent, has raised concerns about potentially being a carcinogen (Takizawa et al. [6]).
Tretinoin, a precursor of vitamin A which is effective in treating hyperpigmentation, was found to
cause erythema and peeling due to its function in accelerating epidermal turnover (Mezick et al. [7]).
More recently, a new product, rhododendrol from Acer nikoense and Betula platyphylla developed by
Kanebo, Inc. in Japan had to be withdrawn from market because of its unexpected result of vitiligo-like
skin disorder (Sasaki et al. [8]). To date, the list of skin-lightening agents continues growing rapidly
but only a handful of them have been carefully characterized in their toxicity and adverse effect.
A number of skin-lightening agents seem to regulate intracellular signaling pathways, which leads
to a decrease of melanin synthesis, and/or increase of melanocyte cell death, and/or possibly
melanocyte differentiation. Although the current understanding is that activation of MAPK and/or Akt
signaling pathways can lead to pigment increase (Imokawa and Ishida [9]), several new skin-lightening
agents have begun to show exceptions. For example, haginin A, a recently identified tyrosinase
inhibitor from Lespedeza cyrtobotrya, functions as a noncompetitive inhibitor of tyrosinase but also
induces the MAPK and Akt signaling pathways (Kim et al. [10]). Another study showed, however,
the extract from Rhodobacter sphaeroides inhibits melanin formation by inhibiting MAPK but activating
Akt pathways (Liu et al. [11]). Yet gallic acid was shown to inhibit melanogenesis by activating both
MAPK and Akt signaling pathways (Su et al. [12]). Therefore, it is still not yet conclusively known
how MAPK and Akt are involved in melanogenesis.
Zebrafish are a very popular tool for basic studies on pigment and color pattern formation due to
the highly conserved mechanisms of melanocyte development and melanin synthesis between fish
and human (Irion et al. [13]). The fast development of melanocytes and ease of observing pigment
change in zebrafish embryos makes it an ideal assay system for testing compounds that are potential
skin-lightening agents (Choi et al. [14]). The zebrafish embryo is also an excellent vertebrate model for
toxicity evaluation for potential drugs (Garcia et al. [15]). However, at least one fundamental difference
in skin color formation exists between fish and mammals. Unlike in mammals, where melanin
is transported from melanocytes to keratinocytes through melanosomes, zebrafish do not have a
melanosome transportation process (Irion et al. [13]), therefore compounds that show an effect on
zebrafish pigment are primarily affecting melanin synthesis and/or metabolism or melanocyte biology.
Our lab has been using an established zebrafish heart failure model (Huang et al. [16]) to
look for compounds that could attenuate heart failure progression. Among the several positive
compounds that were identified in the last few years, A11 and MEK-I were also able to cause significant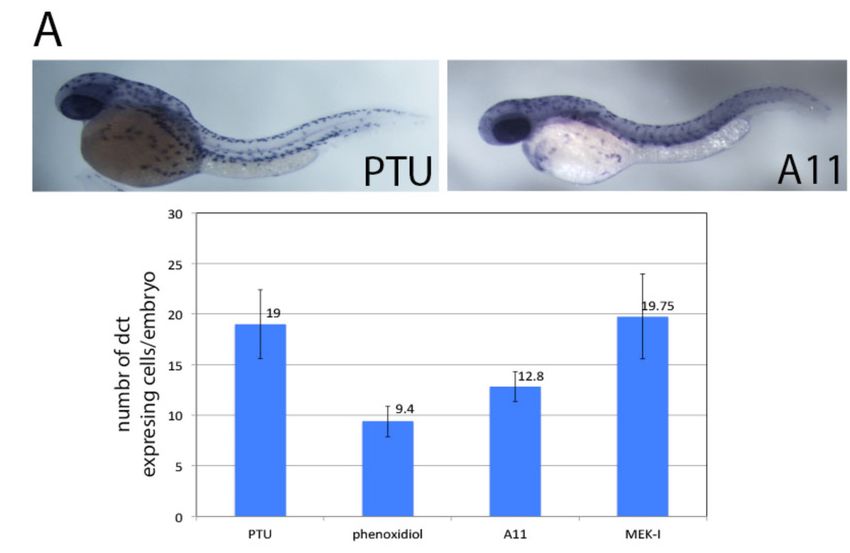
Cosmetics 2018, 5, x FOR PEER REVIEW 3 of 14
Our lab has been using an established zebrafish heart failure model (Huang et al. [16]) to look
Cosmetics 2018, 5, 45 3 of 14
for compounds that could attenuate heart failure progression. Among the several positive
compounds that were identified in the last few years, A11 and MEK-I were also able to cause
significant
reduction reduction of blackinpigment
of black pigment in theembryos.
the zebrafish zebrafish Since
embryos.
MEK-ISince
is aMEK-I
knownisinhibitor
a knownofinhibitor
the MEKof
the MEK (mitogen-activated
(mitogen-activated protein kinase protein kinase
kinase) whichkinase)
functions which
in thefunctions
MAPK pathway in the to MAPK pathway
promote melaninto
promote
synthesismelanin
(reviewed synthesis
in Imokawa(reviewed in Imokawa
and Ishida and
[9]), it is notIshida [9]), it istonot
so surprising seesoMEK-I
surprising to see
causing MEK-
pigment
I reduction. A11, however,
causing pigment reduction. has a very
A11, different
however, has chemical structure
a very different than MEK-I
chemical and than
structure belongs
MEK-Ito the
and
stilbene family which has a characteristic C6-C2-C6 structure, similar to that in resveratrol
belongs to the stilbene family which has a characteristic C6-C2-C6 structure, similar to that in resveratrol (Figure 1).
In this1).
(Figure article,
In thiswe report
article, wethe
reportskin-lightening effect effect
the skin-lightening of MEK-I and and
of MEK-I A11.A11. OurOurin in
vivo studies
vivo studiesusing
using
zebrafish embryos revealed the differential pharmacodynamics of these
zebrafish embryos revealed the differential pharmacodynamics of these two compounds. Unlike most two compounds. Unlike
most stilbenoids,
stilbenoids, A11ais
A11 is not not a tyrosinase
tyrosinase inhibitor.
inhibitor. ThroughThrough cellular
cellular and and molecular
molecular characterizations,
characterizations, we found
we A11
that found thattoA11
seems haveseems
a novel to mechanism
have a novel thanmechanism than othercompounds
other skin-lightening skin-lightening compounds
in inhibiting pigmentin
inhibiting pigment formation by potentially regulating melanocyte differentiation
formation by potentially regulating melanocyte differentiation and proliferation (more supporting data and proliferation
(more
can supporting
be found data can
in Stueven et al.be[17]).
found in Stueven et al. [17]).
Figure
Figure 1.
1. A11
A11and
and MEK-I
MEK-I cause
cause strong pigment inhibition
strong pigment inhibition inin early
earlyzebrafish
zebrafishembryos.
embryos.Zebrafish
Zebrafish
embryos were treated with A11 or
embryos were treated with A11 or MEK-I MEK-I starting from 12, 16, 20, or 24 hpf (hours post fertilization)
20, or 24 hpf (hours post fertilization)
toto48
48 hpf.
hpf. Embryos
Embryos in H22OO showed
in H showed normal
normal level of pigment.
pigment. A11 and and MEK-I
MEK-I both
bothcaused
causedstrong
strong
inhibition
inhibition in pigment
pigmentformation.
formation.However,
However, while
while A11A11 showed
showed consistent
consistent potency
potency in the different
in the different stages
stages of treatment,
of treatment, the potency
the potency of MEK-I
of MEK-I appeared
appeared weakerweaker in thehpf
in the 24–48 24–48 hpf embryo.
embryo.
2.2.Materials
Materialsand
andMethods
Methods
2.1.Zebrafish
2.1. ZebrafishHusbandry
Husbandry and
and In
In Vitro
Vitro Fertilization
The zebrafish
The zebrafish stocks
stocks used
used in this study are are maintained
maintained following
following standard
standard procedures
procedures
(Westerfield [18]) and bred by in vitro fertilization. In brief, mature male and female zebrafishwere
(Westerfield [18]) and bred by in vitro fertilization. In brief, mature male and female zebrafish were
separatedby
separated byaamesh
meshin inaa breeding
breeding tank
tank the
the night
night before
before breeding.
breeding. Soon
Soon after
afterthe
thelight
lightwas
wasturned
turnedon on
the next morning, fish were anesthetized in ~0.16% tricaine for 1–2 min. The females
the next morning, fish were anesthetized in ~0.16% tricaine for 1–2 min. The females were placed on were placed on a
paper towel to dry the body surface briefly before being transferred into a 6 cm dish.
a paper towel to dry the body surface briefly before being transferred into a 6 cm dish. The eggs were The eggs were
expelledonto
expelled ontothe
thedish
dish by
by gently
gently pressing
pressing thethe latero-ventral
latero-ventral side
side of
of the
thebelly
bellywith
withfingers.
fingers.The
Themales
males
were also dried briefly and then placed ventral side up on a sponge well. Under
were also dried briefly and then placed ventral side up on a sponge well. Under a dissection a dissection microscope,
the lateral sides
microscope, the of the belly
lateral sideswere pressed
of the bellygently
were with a pair
pressed of blunt
gently withforceps
a pairwhile holding
of blunt a capillary
forceps while
tube close to the anus to collect sperm. The sperm were immediately mixed with
holding a capillary tube close to the anus to collect sperm. The sperm were immediately mixed with the eggs and ~1 mL
of egg water was added (more details in Westerfield [18]).
the eggs and ~1 mL of egg water was added (more details in Westerfield [18]).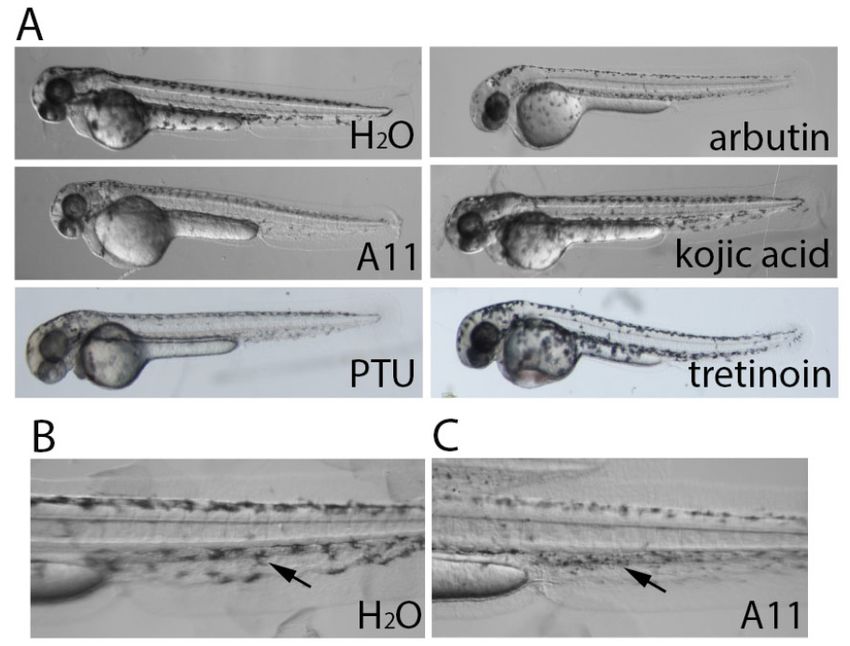
Cosmetics 2018, 5, 45 4 of 14
2.2. Chemical Treatment of Zebrafish Embryos
Zebrafish embryos were collected and arrayed 5 embryos per well of a 96-well plate in 200 µL
of egg water (distilled water containing 60 µg/mL sea salt from Coralife, CA, USA) which were
later replaced with the same volume of egg water containing the desired compound. In the recovery
experiment, embryos were set up in a 96-well plate with the compound from 24–72 hpf, after which
they were transferred to a 6 cm dish to be washed three times with egg water and allowed to continue
their development for several days. Pigment analyses were done using wild type embryos as 100%
pigment and visually comparing the pigment amount in compound-treated embryos. We typically set
up 20 embryos for each treatment group in each experiment with repetition of each experiment.
2.3. Chemicals
A11 and its analogs were synthesized by Dr. Monte at UW-La Crosse. Other chemicals used
in this study were purchased from Sigma-Aldrich (St. Louis, MI, USA): MEK-I (U0126), MoTP
(4-(4-Morpholinobutylthio)phenol, SML0047), and phenoxodiol (D7446) were all prepared at 10 mM in
DMSO, arbutin (A4256) was at 300 mM, kojic acid (K3120) at 10 mM, niacinamide (N5535) 20% (w/v),
tretinoin (PHR1187) 10% (w/v), and PTU (phenylthiourea, P7629) 0.3% (w/v) all in ddH2O.
2.4. In Vitro Tyrosinase Assay
The drug effect on tyrosinase was measured by the in vitro tyrosinase assay (Baurin et al. [19]).
Purified mushroom tyrosinase (cat. T3824, 2100 Unit/mL) and L-tyrosine (T3754, 1.5 mM) were
purchased from Sigma-Aldrich. The reactions were set up in triplicate in a 96-well plate. Each reaction
contained 30 µL L-tyrosine with 20 µL of tyrosinase and 130 µL of 0.1 M phosphate buffer pH6.5 with
or without the test compound (20 µL). The plate was then taken to a microplate reader (Bio-Rad Model
680) to measure the absorbance at 475 nm at multiple time points.
2.5. Immunohistochemistry and TUNEL Assay
Embryos were fixed in 4% paraformaldehyde at least overnight in 4 ◦ C. The fixed embryos
were washed twice with PBS, (phosphate buffered saline) then once with H2 O, 5 min/each,
and permeabilized with −20 ◦ C acetone for 7 min followed by washes in H2 O and then PBS.
The embryos were incubated with 3% BSA (bovine serum albumin) in PBST (PBS with 0.1% Tween 20)
for at least 1 h at room temperature before overnight incubation in 4 ◦ C with the phosphor-Histone
H3 (Ser10) Rabbit mAb-Alexa Fluor 555 conjugate in PBST (Cell Signaling Technology). The next day,
the embryos were washed with PBST at least 4 times for 15 min/each, then stored in ProLong Gold
Antifade Reagent with DAPI (Cell Signaling Technology, Danvers, MA, USA). Apoptotic cells were
detected by TUNEL assay using the TMR In Situ Cell Death Detection Kit from Roche.
2.6. In Situ Hybridization
In situ hybridization protocol was adopted from The Zebrafish Book (Westerfield [18]). Because
of the concern that our compounds do not cause complete albino and the residual pigment might
interfere with the in situ hybridization signal and skew our analysis, we treated embryos with desired
compounds with PTU, and those embryos treated with PTU alone were used as a positive control
in this experiment. PTU does not cause cell death or developmental defects in melanocyte. Twenty
embryos were set up for each compound and treated from 24–48 hpf then fixed for in situ hybridization.
2.7. Western Blotting
Twenty embryos were first treated with desired compound from 24–48 hpf followed by pigment
analysis using a dissection microscope. Embryos were then de-chorionized and put into 200 µL
of lysis buffer (0.2% NP-40, 100 mM Tris, 150 mM NaCl, 8 mM of EDTA, pH 7.4) with protease
inhibitor cocktail added freshly (P8340, Sigma). Embryos were grinded with plastic pestles for 5 minCosmetics 2018, 5, 45 5 of 14
and then further lysed for 2 h in 4 ◦ C on a shaker. The lysate was then centrifuged at 12,000 rpm,
4 ◦ C, for 20 min. The supernatant was transferred to a clean tube and stored in −80 ◦ C. Protein
concentration was determined using the Pierce BCA Assay Kit (23227, Thermo Scientific, Waltham,
MA, USA). 10–20 µg of proteins were loaded into 12% SDS-PAGE gel (Bio-Rad) for gel electrophoresis
and later for Western blotting using the Pierce Fast Western Blot Kit (35050, Thermo Scientific, Waltham,
MA, USA). Antibodies were purchased from Cell Signaling Technology.
2.8. Cell Culture
The mouse melanoma cell line B16F10 (ATCC CRL-6475) was purchased from ATCC (American
Tissue Culture Center, Manassas, VA, USA) and maintained in DMEM (Dulbecco/Vogt modified
Eagle’s minimal essential medium) supplemented with 10% fetal bovine serum. The human primary
epidermal melanocytes (PCS-200-013) was also purchased from ATCC and maintained in Dermal Cell
Basal Medium (PCS-200-030) supplemented with the Adult Melanocyte Growth Kit (PCS-200-042).
Trypsinization and subculture were performed following the protocols found on the ATCC website.
For chemical treatment, approximately 14,000 melanoma cells and 30,000 normal melanocytes were
seeded in each well of the 24-well plate with 0.5 mL of culture medium. Cells were set up in triplicate
and allowed to grow two days for melanoma or three days for normal melanocytes before chemical
treatment. After 24 h of chemical incubation, cells were lysed with 250 µL of 1 N KOH in the well and
180 µL of lysate was transferred to a flat-bottom 96-well plate for melanin measurement.
3. Results
3.1. A11 and MEK-I as Skin-Lightening Compounds
To understand the pharmacodynamics of A11 and MEK-I, we characterized the two compounds
in a series of experiments using zebrafish embryos. First, we tested how early the compounds are
required to produce pigment inhibition phenotypes in the developing embryos. Embryos were treated
with A11 and MEK-I at different stages, 12, 16, 20, 24 hpf (hours post fertilization) and examined for
pigment development at 48 hpf. While A11 caused similar degrees of pigment inhibition in these
embryos, MEK-I showed complete inhibition in early embryos but less inhibition in 24 hpf embryos
(Figure 1). MEK-I appeared to have no inhibition at all in the 30 hpf embryos (data not shown),
indicating MEK-I only functions in the early stage of pigment formation. Alternatively, MEK-I might
have a slower drug effect so that the inhibition was not apparent within the 16-h (from 30–48 hpf)
treatment. This was proved not to be the case as we saw no pigment inhibition by MEK-I at later time
points (data not shown).
Next, we tested the duration of drug effect. Embryos at 24 hpf stage were treated with MEK-I
or A11 and observed at 48 and 72 hpf. Embryos treated with A11 showed about the same pigment
inhibition at both 48 and 72 hpf stages but those in MEK-I only showed effective pigment reduction
in 48 but not in 72 hpf embryos (Figure 2A), indicating A11 has longer inhibition ability than MEK-I.
Embryos incubated in MEK-I from 12–72 hpf showed almost normal level of black pigment, but those
incubated from 20–72 hpf showed greatly reduced pigment content (Figure 2B), indicating the effective
duration of MEK-I is approximately 50 h long. By contrast, A11 caused similar level of pigment
reduction in embryos at all the stages tested, ranging from 14–72 hpf and the pigmentation inhibition
lasted at least for 4 days (data not shown). Thus, A11 has a broader time window and longer
skin-lightening effect than MEK-I. All these results suggest that A11 and MEK-I have different
inhibitory mechanisms.Cosmetics 2018, 5, 45 6 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 6 of 14
Figure
Figure 2. Differential
2. Differential pharmacodynamicsof
pharmacodynamics of A11
A11 and MEK-I.(A)
and MEK-I. (A)Embryos
Embryoswere treated
were withwith
treated A11 or
A11 or
MEK-IMEK-I
for for different
different lengthsofoftime.
lengths time. A11
A11 maintained
maintainedfairly strong
fairly pigment
strong inhibition
pigment from 24–48
inhibition from hpf to hpf
24–48
24–72 hpf while MEK-I apparently lost its potency in the 24–72 hpf embryo, compared to the MEK-I
to 24–72 hpf while MEK-I apparently lost its potency in the 24–72 hpf embryo, compared to the MEK-
24–48 hpf embryo; (B) When embryos were treated from 12 hpf for 3–4 days, A11 still maintained
I 24–48 hpf embryo; (B) When embryos were treated from 12 hpf for 3–4 days, A11 still maintained
strong pigment inhibition but MEK-I did not show any inhibition in the 12–72 or 12–96 hpf embryos.
strong pigment
Arrows pointinhibition but MEK-I
to the notochord did
defects not show
caused any inhibition in the 12–72 or 12–96 hpf embryos.
by MEK-I.
Arrows point to the notochord defects caused by MEK-I.
3.2. Stronger Potency of A11 over Most Skin-Lightening Compounds
3.2. Stronger Potency of A11 over Most Skin-Lightening Compounds
Zebrafish embryos have been used to test skin-lightening compounds due to its conserved melanin
Zebrafish embryos
development mechanisms have beenetused
(Dooley to test
al. [20]). skin-lightening
The popular compoundscompounds
human skin-lightening due to itsarbutin
conserved
melanin development
and kojic acid had been mechanisms (Dooleystrong
shown to produce et al. [20]). The popular
skin-lightening human skin-lightening
effect on developing zebrafish
embryos (Choi
compounds arbutin et al.
and [14]). Weacid
kojic next had
compared
been A11shownand to MEK-I with several
produce strong human skin-lightening
skin-lightening effect on
developing zebrafish embryos (Choi et al. [14]). We next compared A11 and MEK-I withtwo
compounds that are currently on the market: arbutin, kojic acid, niacinamide, tretinoin, and several
recently reported compounds, haginin A (Kim et al. [10], also called
human skin-lightening compounds that are currently on the market: arbutin, kojic acid, niacinamide, phenoxidiol) and MoTP
(Yang et al. [21]) as well as the well-known tyrosinase inhibitor, phenylthiourea (PTU). We found
tretinoin, and two recently reported compounds, haginin A (Kim et al. [10], also called phenoxidiol)
that these skin-lightening compounds caused different degrees of pigment reduction in zebrafish
and MoTP (Yang et al. [21]) as well as the well-known tyrosinase inhibitor, phenylthiourea (PTU).
embryos (Figure 3A). In summary, A11, arbutin, MoTP, phenoxodiol, and PTU could reliably reduce
We found that these skin-lightening compounds caused different degrees of pigment reduction in
the pigment by approximately 90–100%. MEK-I had the same effect but only when it is applied to
zebrafish
younger embryos
embryos.(Figure
Staying3A). In summary,
consistent A11, tyrosinase
with its known arbutin, MoTP,inhibitoryphenoxodiol,
activity, kojicand PTU could
acid caused
reliably reduce the pigment by approximately 90–100%. MEK-I had the same
a lighter color in all the melanocytes (Figure 3C). These comparison studies validate the potential effect but only when it
is applied to younger
skin-lightening embryos.
effect of A11 and Staying
MEK-Iconsistent
on humans. with its known tyrosinase inhibitory activity, kojic
acid caused a lighter
In the color
previous in all the we
experiment, melanocytes
also noted(Figure
that several3C). These
humancomparison
skin-lightening studies validate the
compounds
appeared to cause toxicity in developing
potential skin-lightening effect of A11 and MEK-I on humans.embryos, most of which have not been reported before.
In
Inarbutin-treated
the previous embryos,
experiment, we observed
we also cardiac
noted arrest within 2–4
that several days of
human incubation in a compounds
skin-lightening dosage
dependent manner. Unlike most cardiac toxins which commonly cause
appeared to cause toxicity in developing embryos, most of which have not been reported before. cardiac edema, there was In
no dramatic morphological defects or cardiac edema in the arbutin-treated embryos (unpublished
arbutin-treated embryos, we observed cardiac arrest within 2–4 days of incubation in a dosage
results). Niacinamide, phenoxodiol, and tretinoin appeared to disrupt other developmental processes
dependent manner. Unlike most cardiac toxins which commonly cause cardiac edema, there was no
causing severe morphological defects in multiple tissues (Figure 3A). MoTP has been shown to cause
dramatic morphological defects or cardiac edema in the arbutin-treated embryos (unpublished
melanocyte cell death in a tyrosinase-dependent manner (Yang et al. [21]). MEK-I is interesting in that
results). Niacinamide,
it caused notochord phenoxodiol,
development defects and tretinoin
(Figure 1,appeared
Hawkins to et disrupt other developmental
al. [22], Huang processes
et al. [16]), but only in
causing
earlysevere
embryos.morphological
PTU also cause defects in multiple
notochord distortiontissues
in early (Figure
embryos,3A). MoTP hasdue
presumably been shown
to its abilitytotocause
melanocyte cell death
chelate copper in a reported
ions (also tyrosinase-dependent
in Hawkins et al. manner
[22]. Older(Yang et al. treated
embryos [21]). MEK-I
with MEK-Iis interesting
or PTU in
that were
it caused
able tonotochord
develop normaldevelopment
notochords defects
and live(Figure
(Figures 1, 1Hawkins
and 3). et al. [22], Huang et al. [16]), but
only in early embryos. PTU also cause notochord distortion in early embryos, presumably due to its
ability to chelate copper ions (also reported in Hawkins et al. [22]. Older embryos treated with MEK-
I or PTU were able to develop normal notochords and live (Figures 1 and 3).Cosmetics 2018, 5, x FOR PEER REVIEW 7 of 14
Cosmetics 2018, 5, 45 7 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 7 of 14
Figure 3. Comparison between A11 and known skin-lightening compounds. (A) Zebrafish embryos
were treated with different skin-lightening compounds to show different degrees of pigment
inhibition. A11 (10 μM), PTU (0.003%), MoTP (10 μM), and phenoxidiol (10 μM) all exhibited strong
Figure 3.Figure
Comparison between
3. Comparison A11 and
between A11known skin-lightening
and known skin-lightening compounds.
compounds.(A) (A)Zebrafish
Zebrafishembryos
pigmentembryos
inhibition,
were
but ®-arbutin (100 mM), kojic acid (10 mM), tretinoin (1%), gallic acid (1 mM), and
treated with different skin-lightening compounds to show different degrees of pigment
were treated with different skin-lightening compounds to show different degrees of pigment
niacinamide
inhibition. A11 (10 µM), PTU (0.003%), MoTP (10 µM), and phenoxidiol (10 µM) all exhibited strong caused
(1%) showed weaker inhibition. Some skin-lightening compounds apparently
inhibition. A11 (10 μM), PTU® (0.003%), MoTP (10 μM), and phenoxidiol (10 μM) all exhibited strong
strong toxicity
pigment to developing
inhibition, embryos,
but -arbutin (100body
mM), curvature bymM),
kojic acid (10 niacinamide andgallic
tretinoin (1%), eye acid
and(1tail defects by
mM),
pigmentand inhibition,
niacinamide
but(1%)
®-arbutin (100 mM), kojic acid (10 mM), tretinoin (1%), gallic acid (1 mM), and
showed weaker inhibition. Some skin-lightening compounds apparently caused
tretinoin. High magnification of the head area of embryos in H2O (B) and kojic acid (C) are shown to
niacinamide
strong(1%) showed
toxicity weaker
to developing inhibition.
embryos, body Some skin-lightening
curvature by niacinamide compounds
and eye and tail apparently
defects by caused
display the lighter pigment in melanocytes (arrow) caused by kojic acid.
strong toxicity to developing embryos, body curvature by niacinamide
tretinoin. High magnification of the head area of embryos in H 2 O (B) and and eye and
kojic acid (C) are tail defects
shown to by
display the lighter pigment in melanocytes (arrow) caused by kojic acid.
tretinoin. High magnification of the head area of embryos in H2O (B) and kojic acid (C) are shown to
3.3. A11 Reduces Existing Pigment
display the lighter pigment in melanocytes (arrow) caused by kojic acid.
3.3. A11 Reduces Existing Pigment
To test whether the skin-lightening compounds had any effect on already formed pigment, we
3.3. A11 48
treated To test
Reduces
hpf whether
Existing
embryos the skin-lightening
Pigment
with the compounds compounds
until 72hadorany96effect
hpf.onInterestingly,
already formedwhilepigment,
most skin-
we treated 48 hpf embryos with the compounds until 72 or 96 hpf. Interestingly, while most
lightening compounds had little or no effect on existing pigment, A11 caused significant reduction of
To test whether compounds
skin-lightening the skin-lightening
had little or compounds had pigment,
no effect on existing any effectA11on already
caused formed
significant pigment, we
reduction
pigmentofwithin
pigment 24within
h of treatment
24 h of (Figure
treatment 4A).
(Figure Interestingly,
4A). Interestingly, the
the melanocytes
melanocytes in in A11-treated
A11-treated embryos embryos
treated 48 hpf embryos with the compounds until 72 or 96 hpf. Interestingly, while most skin-
appeared contracted
appeared thanthan
contracted those in control
those (Figure 4C).
4C).
lightening compounds had little or in
nocontrol
effect(Figure
on existing pigment, A11 caused significant reduction of
pigment within 24 h of treatment (Figure 4A). Interestingly, the melanocytes in A11-treated embryos
appeared contracted than those in control (Figure 4C).
Figure 4. Cont.
Figure 4. A11 reduces existing pigment. (A) Embryos at 48 hpf stage that already generated lots of
pigment were selected for this experiment to receive compound treatment until 72 hpf and then were
photographed. A11 caused significant pigment reduction compared to the control at 72 hpf (even lessCosmetics 2018, 5, 45 8 of 14
Figure 4.Figure
A11 4.reduces
A11 reducesexisting pigment.
existing pigment.(A)
(A) Embryos
Embryos atat
4848
hpfhpf stage
stage that already
that already generated
generated lots of lots of
pigment were selected for this experiment to receive compound treatment
pigment were selected for this experiment to receive compound treatment until 72 hpf and until 72 hpf and then werethen were
photographed. A11 caused significant pigment reduction compared to the control at 72 hpf (even less
photographed. A11 caused significant pigment reduction compared to the control at 72 hpf (even less
than 48 hpf normal embryo in Figure 3). High magnification of H2 O (B) and A11 (C) treated embryos
than 48 hpf normal
are shown embryo
to display thein Figure 3).
contracted High magnification
morphology of melanocytesof(arrows).
H2O (B) and A11 (C) treated embryos
Cosmetics 2018, 5, x FOR PEER REVIEW
are shown to display the contracted morphology of melanocytes (arrows). 8 of 14
3.4. Slow Pigment Recovery by A11
3.4. Slow Pigment Recovery by A11
AsAsmost skin-lightening
most compounds
skin-lightening compoundsare tyrosinase inhibitors,
are tyrosinase it has been
inhibitors, shown
it has thatshown
been pigmentation
that
could recover within 24 h after compounds were washed off. To test this notion,
pigmentation could recover within 24 h after compounds were washed off. To test this embryos were treated
notion,
with skin-lightening
embryos compounds
were treated from 20–48 or
with skin-lightening 72 hpf, washed
compounds from twice,
20–48 and
or 72transferred to egg
hpf, washed water
twice, andfor
24 transferred
h or longer.toThe
eggresults showed
water for that
24 h or pigmentation
longer. in showed
The results most embryos did recoverinquickly
that pigmentation but not in
most embryos
A11didorrecover
arbutin-treated embryos
quickly but (Figure
not in A11 5B).
or arbutin-treated embryos (Figure 5B).
Figure 5. Slow pigment recovery by A11. Embryos were treated with skin-lightening compounds
Figure 5. Slow pigment recovery by A11. Embryos were treated with skin-lightening compounds
from 24–72 hpf (A) then washed with egg water twice and allowed to develop until 96 hpf (B) in water
from 24–72 hpf (A) then washed with egg water twice and allowed to develop until 96 hpf (B) in water
to test how quickly pigment recovers after compound withdrawal. Most embryos recovered their
to test how quickly pigment recovers after compound withdrawal. Most embryos recovered their
pigment quickly except for A11 and arbutin.
pigment quickly except for A11 and arbutin.
3.5. A11 is Not a Tyrosinase Inhibitor
3.5. A11 is Not a Tyrosinase Inhibitor
It has been shown that many stilbenoids have tyrosinase inhibitory activity (Likhitwitayawuid
It Since
[23]). has A11been shown that
is a stilbene-like many we
compound, stilbenoids
tested whether haveA11tyrosinase
or MEK-I caninhibitory activity
inhibit tyrosinase
(Likhitwitayawuid [23]). Since
using an in vitro tyrosinase assay A11 is aet stilbene-like
(Baurin al. [19]). Whilecompound, we tested whether
most of the skin-lightening compoundsA11 or
MEK-I
showedcanstrong
inhibitinhibition
tyrosinaseof using an inneither
tyrosinase, vitro tyrosinase
A11 or MEK-I assay (Baurin
did et 10
at either al.μM
[19]).
andWhile
50 μM most of the
(Figure
skin-lightening
6 and data notcompounds
shown). It isshowed strong
important inhibition
to note of tyrosinase,
that phenoxodiol had aneither A11 tyrosinase
very short or MEK-I did at either
inhibition
10 efficacy
µM andbut 50 gallic acid and
µM (Figure MoTP
6 and actually
data enhanced
not shown). It istyrosinase
important activity
to note(Figure 6).
that phenoxodiol had a very
short tyrosinase inhibition efficacy but gallic acid and MoTP actually enhanced tyrosinase activity
3.6. A11
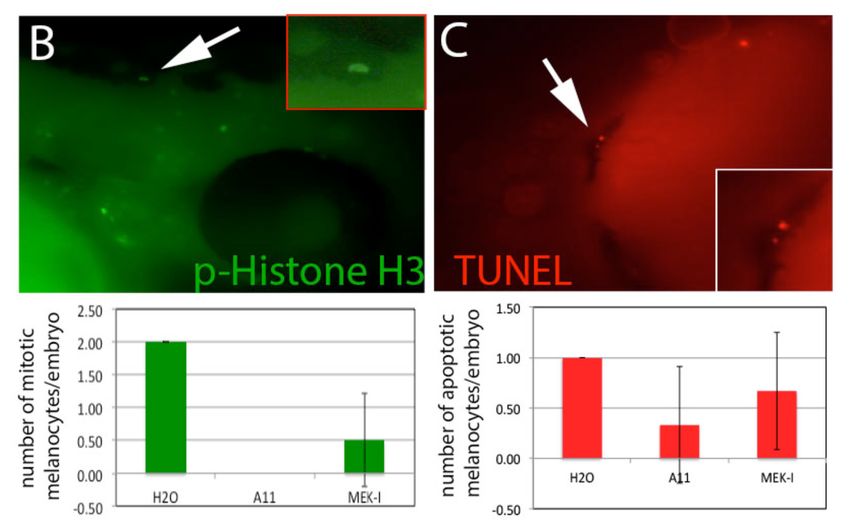
(Figure 6). Causes Defects in Melanocyte Differentiation
To further understand the mechanism of A11, we examined the development of melanocytes by
in situ hybridization for the dct (dopachrome tautomerase) gene which is expressed in differentiated
melanocytes. The result showed that the number of dct-expressing cells in A11-treated embryos is
significantly lower than in the control (Figure 7A). To test whether the fewer dct-expressing cells is
due to decreased proliferation of melanocyte, we performed immunostaining using the anti-Cosmetics 2018, 5, 45 9 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 9 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 9 of 14
Figure 6. In vitro tyrosinase assay. Reaction containing L-tyrosine and tyrosinase with or without
Figure 6. In vitro tyrosinase assay. Reaction containing L-tyrosine and tyrosinase with or without
skin-lightening compounds were set up in triplicate in a 96-well plate. The absorbance at 475 nm were
skin-lightening compounds were set up in triplicate in a 96-well plate. The absorbance at 475 nm were
taken at 20′, 60′ and 120′. Several known skin-lightening compounds (8–11) showed strong tyrosinase
taken at 200 , 600 and 1200 . Several known skin-lightening compounds (8–11) showed strong tyrosinase
inhibition, as the absorbance stayed near zero, including arbutin, kojic acid (KA), PTU, and
inhibition, as the absorbance
niacinamide stayed
(Nia). However, A11near zero,
(5) and including
MEK-I arbutin,
(4) did kojic
not show anyacid (KA), PTU,
inhibition and niacinamide
of tyrosinase at all
(Nia).asHowever, A11 functioned
the tyrosinase (5) and MEK-I (4)Hdid
like in not show any inhibition of tyrosinase at all as the tyrosinase
2O (6).
functioned like in H2 O (6).
3.6. A11 Causes Defects in Melanocyte Differentiation
To further understand the mechanism of A11, we examined the development of melanocytes by
in situ hybridization for the dct (dopachrome tautomerase) gene which is expressed in differentiated
melanocytes. The result showed that the number of dct-expressing cells in A11-treated embryos is
significantly lower
Figure 6. Inthan
vitroin the control
tyrosinase assay.(Figure
Reaction7A). To test
containing whether
L-tyrosine andthe fewer dct-expressing
tyrosinase with or without cells is due
to decreased proliferation of melanocyte, we performed immunostaining usingatthe
skin-lightening compounds were set up in triplicate in a 96-well plate. The absorbance 475anti-phosphorylated
nm were
taken at 20′, 60′ and 120′. Several known skin-lightening compounds (8–11) showed strong tyrosinase
histone H3inhibition,
antibody.as The result showed that embryos treated with A11 had lower level of proliferative
the absorbance stayed near zero, including arbutin, kojic acid (KA), PTU, and
melanocytes (Figure 7B). To test ifA11
niacinamide (Nia). However, A11 (5)induced
and MEK-Iapoptosis in melanocytes,
(4) did not show any inhibition ofTUNEL
tyrosinaseexperiments
at all were
performedasand showedfunctioned
the tyrosinase no significant
like in Hapoptosis
2O (6). in A11-treated embryos (Figure 7C).
Figure 7. Cont.Cosmetics 2018, 5, 45 10 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 10 of 14
Figure
Figure 7. 7. Inhibition
Inhibition of melanocyte
of melanocyte differentiation
differentiation and proliferation
and proliferation by (A)
by A11. A11.In(A)
situIn situ
hybridization
hybridization
using dct using revealed
gene riboprobe dct gene fewer
riboprobe revealed fewer
differentiated differentiated
melanocytes melanocytes in A11-PTU-
in A11-PTU-treated embryos than
treated embryos than those in PTU alone. The quantitative data is shown in the chart below. MEK-I
those in PTU alone. The quantitative data is shown in the chart below. MEK-I has no effect on
has no effect on melanocyte differentiation; (B) Immunostaining using rabbit mAb for
melanocyte differentiation; (B) Immunostaining using rabbit mAb for phosphorylated histone H3
phosphorylated histone H3 (p-Histone H3) on embryos treated with A11 or MEK-I showed fewer
(p-Histone H3) on Arrow
mitotic cells. embryos treated
points with A11
to a melanocyte orwas
that MEK-I showed
positive fewer mitotic
with p-Histone cells.view
H3 (enlarged Arrow points to
in the
a melanocyte thatTUNEL
insert). (C) was positive with
assay reveals p-Histone
little H3 (enlarged
or no difference view
in the number of in the insert).
melanocytes (C) TUNEL
between control assay
and A11-treated
reveals little embryos.inArrow
or no difference points toofanmelanocytes
the number apoptotic melanocyte
between (enlarged
controlview
andinA11-treated
the insert). embryos.
Arrow points to an apoptotic melanocyte (enlarged view in the insert).
3.7. A11 Activates MAPK and Akt Pathways in Fish Embryo
MAPK MAPK
3.7. A11 Activates signaling pathway
and has been in
Akt Pathways shown
Fish to positively regulate pigment formation (Imokawa
Embryo
and Ishida [9]). We performed Western blotting to examine how A11 and MEK-I influence MAPK
MAPK signaling
activity pathway
to regulate pigment has been shown
formation. to positively
MEK-I showed expected regulate pigment
inhibition of MAPKformation (Imokawa
activity which
is consistent with the current literature. To our surprise, A11 caused
and Ishida [9]). We performed Western blotting to examine how A11 and MEK-I influence a distinguishably higher level of MAPK
phosphorylated MAPK (activated MAPK). Another signaling pathway
activity to regulate pigment formation. MEK-I showed expected inhibition of MAPK activity whichthat is known to regulate
pigment formation is Akt. Studies established that activation of Akt causes suppression of melanin
is consistent with the current literature. To our surprise, A11 caused a distinguishably higher
synthesis in G361 melanoma cells (Oka et al. [24]). We also found that A11 slightly increases
level ofphosphorylated
phosphorylated Akt. MAPK (activated
MEK-I, however, MAPK).
decreases Another signaling
the phosphorylated pathway
Akt (Figure that
8). Note thatisdue
known to
regulatetopigment formation is Akt. Studies established that activation of Akt
the high content of lipids and proteins in the embryonic yolk, the amount of pMAPK and pAkt causes suppression of
melaninproteins
synthesis in G361
appeared muchmelanoma
less and thecells
bands(Oka
wereet fainter
al. [24]). Wegel
in the also
thanfound that A11
we expected. slightlywe
However, increases
were able to
phosphorylated Akt.observe
MEK-I,a similar increase
however, of pMAPK
decreases theand pAkt by A11 using
phosphorylated AktB16F10 mouse
(Figure 8). melanoma
Note that due to
the highcells (Stueven et al. [17]).
content of lipids and proteins in the embryonic yolk, the amount of pMAPK and pAkt proteins
appeared much less and the bands were fainter in the gel than we expected. However, we were able to
observe a similar increase of pMAPK and pAkt by A11 using B16F10 mouse melanoma cells (Stueven
et al. [17]).Cosmetics 2018, 5, 45 11 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 11 of 14
Cosmetics 2018, 5, x FOR PEER REVIEW 11 of 14
Figure 8. Activation of MAPK and Akt by A11. Western blotting using anti-total MAPK, anti-P-
Figure 8. Activation of MAPK and Akt by A11. Western blotting using anti-total MAPK, anti-P-MPAK,
MPAK,8.orActivation
Figure anti-Akt antibodies
of MAPK showed that
by A11 caused higher levelusing
of phosphorylated MAPK (p-
or anti-Akt antibodies showed thatand
A11Akt
caused A11. Western
higher level ofblotting
phosphorylated anti-total
MAPKMAPK,
(p-MAPK)anti-P-
and
MAPK) and Akt (pAkt). MEK-I inhibited MAPK activation strongly and caused inhibition of Akt.
AktMPAK,
(pAkt).orMEK-I
anti-Akt antibodies
inhibited MAPKshowed that A11
activation caused
strongly higher
and caused level of phosphorylated
inhibition of Akt. PTUMAPK
showed (p-no
PTU showed
MAPK) no effect
and Akt on MAPK and Akt. Total MAPK was used as an internal control which showed
effect on MAPK and(pAkt). MEK-I
Akt. Total inhibited
MAPK wasMAPK
used asactivation
an internalstrongly
controland caused
which inhibition
showed of Akt.
nearly equal
nearly
PTU equal amount
showed no ofonproteins
effect MAPK in
andeach
Akt.lane.
Total MAPK was used as an internal control which showed
amount of proteins in each lane.
nearly equal amount of proteins in each lane.
3.8. A11 Also Causes Melanin Reduction in Mammalian Cells
3.8. A11 Also Causes Melanin Reduction in Mammalian Cells
3.8. A11 Also we
Finally, Causes Melanin
tested Reduction
whether A11 hasin Mammalian
a similar drug Cellseffect on mammalian cells. We chose to use
Finally,
B16F10 wewe
mouse
Finally,
tested whether
melanoma A11
cells which
tested whether
has
areahighly
A11 has
similar
a similar
drug
pigmented effect
drug effect
on
andon mammalian
metastatic
mammalian
cells.
cancer cells
cells. We
We
that chose to use
havetobeen
chose use
B16F10
B16F10mouse
widely used
mouse melanoma
formelanoma cells
skin-lighteningwhich are
compound
cells which highly pigmented
test pigmented
are highly and humanand and metastatic
primary cancer
melanocytes
metastatic cells
which
cancer cells that
thatare have been
derived
have been
widely
fromused
widely the forfor
used skin-lightening
epidermis of humancompound
skin-lightening foreskin.
compound Intest
both
test and
and human
cases,
human primary
we primary melanocytes
observedmelanocytes which
reduction ofwhich
melanin arederived
are byderived
A11
from the epidermis
comparable to of
other human foreskin.
skin-lightening In
compoundsboth cases,
(Figure we
9). observed reduction
from the epidermis of human foreskin. In both cases, we observed reduction of melanin by A11 of melanin by A11
comparable to other skin-lightening compounds
comparable to other skin-lightening compounds (Figure 9). (Figure 9).
Figure 9. A11 also causes pigment reduction in mammalian melanocytes. (A) Cultured mouse
melanoma cellsalso
B16F10 were treated reduction
with different skin-lightening compounds for 24 h. The cells
Figure 9. 9.A11
Figure A11 causes
also causes pigment
pigment reduction in in mammalian
mammalian melanocytes.
melanocytes. (A)
(A)Cultured
Culturedmouse
mouse
were lysed with
melanoma cells KOH
B16F10 and the melanin
were was measured
treated with with spectrophotometer
different skin-lightening at 475
compounds fornm.
24 A11 showed
h. The cells
melanoma cells B16F10 were treated with different skin-lightening compounds for 24 h. The cells were
stronglysed
were reduction
with KOHof melanin
and the at 40 μMwas
melanin (A11-40)
measuredbut with
no reduction at 10 μM (A11-10).
spectrophotometer at 475 nm.Several skin-
A11 showed
lysed with KOH and the melanin was measured with spectrophotometer at 475 nm. A11 showed strong
lightening compounds also showed strong reduction of melanin. MEK-I, however,
strong reduction of melanin at 40 μM (A11-40) but no reduction at 10 μM (A11-10). Several skin- showed no
reduction
melanin ofreduction
melanininatB16F10
40 µMcells;
(A11-40)
(B) but no human
Normal reduction at 10 µMwere
melanocytes (A11-10). Several
treated with skin-lightening
skin-lightening
lightening compounds also showed strong reduction of melanin. MEK-I, however, showed no
compounds
compounds. alsoA11,
showed
PTU, strong reduction
kojiccells;
acid, and of melanin.
arbutin MEK-I,
showed however,
melanin showed
inhibition. no melanin
Again, MEK-I reduction
did not
melanin reduction in B16F10 (B) Normal human melanocytes were treated with skin-lightening
in B16F10
show cells; (B)
inhibition Normal
at all. human
FSK0392 is melanocytes
an A11 analog were
which treated
also with
showed skin-lightening
strong compounds.
inhibition. A11,
compounds. A11, PTU, kojic acid, and arbutin showed melanin inhibition. Again, MEK-I did not
PTU, kojic acid, and arbutin showed melanin inhibition. Again, MEK-I did not show inhibition at all.
show inhibition at all. FSK0392 is an A11 analog which also showed strong inhibition.
FSK0392 is an A11 analog which also showed strong inhibition.Cosmetics 2018, 5, 45 12 of 14
4. Discussion
4.1. A11 as a Potent and Safer Skin-Lightening Compound
Through the comparison studies of A11 and other skin-lightening compounds, we found that A11
seems to be a potent yet safe compound for cosmetic product. A11 at 10 µM inhibits almost 90% of
pigment in zebrafish embryos up to 5 days post fertilization. A11 at 40 µM also shows strong melanin
inhibition in mammalian cells comparable to other skin-lightening compounds, such as kojic acid
and arbutin. Unlike niacinamide or tretinoin, which caused strong toxicity in developing zebrafish
embryos, A11 shows no detectable toxicity up to 40 µM. Through the late treatment experiment
(Figure 4), we found that only a few skin-lightening compounds could reduce existing pigment,
which included A11. In the pigment recovery experiment, A11-treated embryos recovered slower
than other skin-lightening treated embryos. During our studies, we found that phenoxidiol was
the other compound that could reduce existing pigment, but the phenoxidiol-treated embryos were
very sick and died in a few days, with phenoxidiol toxicity being irreversible. A11 can effectively
reduce melanin in mammalian cells, both in highly pigmented melanoma cells and normal human
melanocytes. Together, we conclude that A11 seems superior than other skin-lightening compounds.
4.2. Possible Cellular Mechanism of A11
Studies show that most skin-lightening compounds reduce melanin synthesis by simply inhibiting
tyrosinase enzyme activity with little toxicity on melanocytes. Those compounds, including resveratrol,
could be competitive or noncompetitive inhibitor of tyrosinase. Some skin-lightening compounds,
however, bind to tyrosinase which metabolizes into toxic product and kills melanocytes, such as
hydroquinone, MoTP (Yang et al. [21]), and rhododendrol (Sasaki et al. [8]). A11 did not inhibit
tyrosinase or cause melanocyte apoptosis, which makes A11 a very interesting skin-lightening
compound. Our studies clearly show that A11 inhibits melanocyte proliferation (Figure 7). However,
that cannot be the only mechanism because it does not explain how A11 can reduce existing pigment.
By examining the number of differentiated melanocytes using in situ hybridization, we further
discovered that A11 actually caused fewer differentiated melanocytes (Figure 7). These results
together lead to an interesting model where A11 possibly causes reduction of melanocyte number by
inhibiting melanocyte differentiation. If this model is correct, we anticipate a reduction of expression of
melanocyte differentiation genes. Indeed, we recently published another paper using mouse melanoma
cells to clearly see reduction of the Mitf and tyrosinase genes by A11 (Stueven et al. [17]).
4.3. MAPK Signaling Pathway in Melanogenesis
Only several skin-lightening compounds have demonstrated the ability to inhibit tyrosinase
in addition to activating or inhibiting intracellular signal cascades leading to the transcriptional
inhibition of melanin synthesis genes, such as haginin A (Kim et al. [10]), gallic acid (Su et al. [12]),
and hydroquinone (Inoue et al. [25]). These studies focused on the MAPK and Akt signaling pathways.
Unfortunately, those and our results do not quite agree with the current molecular mechanism for
melanogenesis. In the review article by Imokawa and Ishida [9], UV light and α-MSH (melanocyte
stimulating hormone) induce melanin synthesis by activating MAPK and PKC pathways but haginin
A and gallic acid both activate MAPK and Akt to reduce melanin (Kim et al. [10]). In our study,
we show that MEK-I reduces pigment and inhibits MAPK at the same time in zebrafish embryo. A11,
however, reduces pigment and activates MAPK and Akt simultaneously. One potential explanation
for this is that the activation of MAPK and Akt by A11 does not directly lead to melanin synthesis,
but instead there is another as-yet unidentified signaling pathway(s) involved in melanin synthesis
that is regulated by A11.Cosmetics 2018, 5, 45 13 of 14
Author Contributions: Conceptualization, A.M. and C.-c.H.; Methodology, K.M., N.S., C.-c.H.; Validation, K.M,
N.S., C.-c.H.; Formal Analysis, C.-c.H.; Investigation, C.-c.H.; Resources, A.M. and C.-c.H.; Data Curation, A.M.
and C.-c.H.; Writing-Original Draft Preparation, C.-c.H.; Writing-Review & Editing, C.-c.H.; Supervision, C.-c.H.;
Project Administration, C.-c.H.; Funding Acquisition, C.-c.H.
Funding: This research was funded by the University of Wisconsin System grant number 106-SYS-06-8000-4 and
UW-River Falls grant number [PRJ82VD].
Acknowledgments: This project was supported by a Faculty Research Grant (PRJ82VD) to C.-c.H., and Summer
Research Scholarships to K.M. and N.S. from University of Wisconsin-River Falls and a Prototype Development
grant (106-SYS-06-8000-4) to C.-c.H. from the University of Wisconsin System. The authors thank Michael J. Martin
for comments and suggestions on the manuscript.
Conflicts of Interest: The authors declare no conflict of interest.
References
1. Battaini, G.; Monzani, E.; Casella, L.; Santagostini, L.; Pagliarin, R. Inhibition of the catecholase activity of
biomimetic dinulcear copper complexes by kojic acid. J. Biol. Inorg. Chem. 2000, 5, 262–268. [CrossRef]
[PubMed]
2. Jones, K.; Hughes, J.; Hong, M.; Jia, Q.; Orndorff, S. Modulation of melanogenesis by aloesin: A competitive
inhibitor of tyrosinase. Pigment Cell Res. 2002, 15, 335–340. [CrossRef] [PubMed]
3. Kim, H.J.; Choi, H.-R.; Kim, D.-S.; Park, K.-C. Topical hypopigmenting agents for pigmentary disorders and
their mechanisms of action. Ann. Dermatol. 2012, 24, 1–6. [CrossRef] [PubMed]
4. Brenner, M.; Hearing, V.J. Modifying skin pigmentation-approaches through intrinsic biochemistry and
exogenous agents. Drug Discov. Today Dis. Mech. 2008, 5, e189–e199. [CrossRef] [PubMed]
5. Eastmond, D.A.; Rupa, D.S.; Hasegawa, L.S. Detection of hyperdiploidy and chromosome breakage
in interphase human lymphocytes following exposure to the benzene metabolite hydroquinone using
multicolor fluorescence in situ hybridization with DNA probes. Mutat. Res. 1994, 322, 9–20. [CrossRef]
6. Takizawa, T.; Mitsumori, K.; Tamura, T.; Nasu, M.; Ueda, M.; Imai, T.; Hirose, M. Hepatocellular tumor
induction in heterozygous p53-deficient CBA mice by a 26-week dietary administration of kojic acid.
Toxicol. Sci. 2003, 73, 287–293. [CrossRef] [PubMed]
7. Mezick, J.A.; Bhatia, M.C.; Capetola, R.M. Topical and systemic effects of retinoids on hor-filled utriculus
size in the rhino mouse, A model to quantify “anti-keratinizing” effect of retinoids. J. Investig. Dermatol.
1984, 83, 110–113. [CrossRef] [PubMed]
8. Sasaki, M.; Kondo, M.; Sato, K.; Umeda, M.; Kawabata, K.; Takahashi, Y.; Suzuki, T.; Matsunaga, K.;
Inoue, S. Rhododendrol, a depigmentation-inducing phenolic compound, exerts melanocyte cytotoxicity via
a tyrosinase-dependent mechanism. Pigment Cell Melanoma Res. 2004, 27, 754–763. [CrossRef] [PubMed]
9. Imokawa, G.; Ishida, K. Inhibitors of intracellular signaling pathways that lead to stimulated epidermal
pigmentation: Perspective of anti-pigmenting agents. Int. J. Mol. Sci. 2014, 15, 8293–8315. [CrossRef]
[PubMed]
10. Kim, J.H.; Baek, S.H.; Kim, D.H.; Choi, T.Y.; Yoon, T.J.; Hwang, J.S.; Kim, M.R.; Kwon, H.J.; Lee, C.H.
Down regulation of melanin synthesis by haginin A and its application to in vivo lightening model.
J. Investig. Dermatol. 2008, 128, 1227–1235. [CrossRef] [PubMed]
11. Lu, W.-S.; Kuan, Y.-D.; Chiu, K.-H.; Wang, W.-K.; Chang, F.-H.; Liu, C.-H.; Lee, C.-H. The extract of Rhodobacter
sphaeroides inhibits melanogenesis through the MEK/ERK signaling pathway. Mar. Drugs 2013, 11, 1899–1908.
[CrossRef] [PubMed]
12. Su, T.-R.; Lin, J.-J.; Tsai, C.-C.; Huang, T.-K.; Yang, Z.-Y.; Wu, M.-O.; Zheng, Y.-Q.; Su, C.-C.; Wu, Y.-J. Inhibition
of melanogenesis by gallic acid: Possible involvement of the PI3K/Akt, MEK/ERK and Wnt/b-catenin
signaling pathways in B16F10 cells. Int. J. Mol. Sci. 2013, 14, 20443–20458. [CrossRef] [PubMed]
13. Irion, U.; Singh, A.P.; Nusslein-Volhard, C. The developmental genetics of vertebrate color pattern formation:
Lessons from zebrafish. Curr. Top. Dev. Biol. 2016, 117, 141–169. [PubMed]
14. Choi, T.-Y.; Kim, J.-H.; Ko, D.H.; Kim, C.H.; Hwang, J.-S.; Ahn, S.; Kim, S.Y.; Kim, C.-D.; Lee, J.-H.;
Yoon, T.-J. Zebrafish as a new model for phenotype-based screening of melanogenic regulatory compounds.
Pigment Cell Res. 2007, 20, 120–127. [CrossRef] [PubMed]
15. Garcia, G.R.; Noyes, P.D.; Tanguay, R.L. Advancements in zebrafish applications for 21st century toxicology.
Pharmacol. Ther. 2016, 161, 11–21. [CrossRef] [PubMed]Cosmetics 2018, 5, 45 14 of 14
16. Huang, C.-C.; Monte, A.; Cook, J.M.; Kabir, M.S.; Peterson, K.P. Zebrafish heart failure models for the
evaluation of chemical probes and drugs. Assay Drug Dev. 2013, 11, 561–572. [CrossRef] [PubMed]
17. Stueven, N.A.; Schlaeger, N.M.; Monte, A.P.; Hwang, S.-P.L.; Huang, C.-C. A novel stilbene-like
compound that inhibits melanoma growth by regulating melanocyte differentiation and proliferation.
Toxicol. Appl. Pharm. 2017, 337, 30–38. [CrossRef] [PubMed]
18. Westerfield, M. The Zebrafish Book; A Guide for the Laboratory Use of Zebrafish (Danio rerio); University of
Oregon Press: Eugene, OR, USA, 2007.
19. Baurin, N.; Arnoult, E.; Scior, T.; Do, Q.T.; Bernard, P. Preliminary screening of some tropical plants for
anti-tyrosinase activity. J. Ethnopharmacol. 2002, 82, 155–158. [CrossRef]
20. Dooley, C.M.; Schwarz, H.; Mueller, K.P.; Mongera, A.; Konantz, M.; Neuhauss, S.C.F.; Nusslein-Volhard, C.;
Geisler, R. Slc45a2 and V-ATPase are regulators of melanosomal pH homeostasis in zebrafish, providing
a mechanism for human pigment evolution and disease. Pigment Cell Melanoma Res. 2013, 26, 205–217.
[CrossRef] [PubMed]
21. Yang, C.-T.; Johnson, S.L. Small molecule-induced ablation and subsequent regeneration of larval zebrafish
melanocytes. Development 2006, 133, 3563–3573. [CrossRef] [PubMed]
22. Hawkins, T.A.; Cavodeassi, F.; Erdelyi, F.; Szabo, G.; Lele, A. The small molecule Mek1/2 inhibitor U0126
disrupts the chrodamesoderm to notochord transition in zebrafish. BMC Dev. Biol. 2008, 8, 42. [CrossRef]
[PubMed]
23. Likhitwitayawuid, K. Stilbenes with tyrosinase inhibitory activity. Curr. Sci. 2008, 94, 44–52.
24. Oka, M.; Nagai, H.; Ando, H.; Fukunaga, M.; Matsumura, M.; Araki, K.; Ogawa, W.; Miki, T.;
Sakaue, M.; Tsukamoto, K.; Konishi, H.; Kikkawa, U.; Ichihashi, M. Regulation of melanogenesis through
phosphatidylinositol 3-kinase-Akt pathway in human G361 melanoma cells. J. Investig. Dermatol. 2000,
115, 699–703. [CrossRef] [PubMed]
25. Inoue, Y.; Hasegawa, S.; Yamada, T.; Date, Y.; Mizutani, H.; Nakata, S.; Matsunaga, K.; Akamatsu, H. Analysis
of the effects of hydroquinone and arbutin on the differentiation of melanocytes. Biol. Pharm. Bull. 2013,
36, 1722–1730. [CrossRef] [PubMed]
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access
article distributed under the terms and conditions of the Creative Commons Attribution
(CC BY) license (http://creativecommons.org/licenses/by/4.0/).You can also read

















































