Antibodies for Human Immunodeficiency Virus-1 Cure Strategies
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
The Journal of Infectious Diseases
SUPPLEMENT ARTICLE
Antibodies for Human Immunodeficiency Virus-1 Cure
Strategies
Evan Rossignol,1 Galit Alter,1, and Boris Julg1,2
1
Ragon Institute of MGH, MIT, and Harvard, Cambridge, Massachusetts, USA, and 2Massachusetts General Hospital, Infectious Disease Unit, Boston, Massachusetts, USA
Human immunodeficiency virus (HIV) infection leads to the establishment of a long-lived latent cellular reservoir. One strategy to
eliminate quiescent reservoir cells is to reactivate virus replication to induce HIV envelope glycoprotein (Env) expression on the cell
surface exposing them to subsequent antibody targeting. Via the interactions between the antibody Fc domain and Fc-γ receptors
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
(FcγRs) that are expressed on innate effector cells, such as natural killer cells, monocytes, and neutrophils, antibodies can mediate the
elimination of infected cells. Over the last decade, a multitude of human monoclonal antibodies that are broadly neutralizing across
many HIV-1 subtypes have been identified and are currently being explored for HIV eradication strategies. Antibody development
also includes novel Fc engineering approaches to increase engagement of effector cells and optimize antireservoir efficacy. In this
review, we discuss the usefulness of antibodies for HIV eradication approaches specifically focusing on antibody-mediated strategies
to target latently infected cells and options to increase antibody efficacy.
Keywords. HIV reservoir; monoclonal antibodies; Infected cell recognition; Fc function; HIV cure.
Although antiretroviral therapy (ART) has enabled pharmaco- to target various sites of vulnerability on the HIV-1 Env trimer,
logical suppression of human immunodeficiency virus (HIV) including the CD4-binding site (CD4bs) of glycoprotein (gp)
type 1 replication, it has become evident that even lifelong 120, the V2 glycan site at the apex of the Env trimer, the V3
therapy with ART will not eradicate the virus. HIV-1 infection glycan site, the membrane-proximal external region of gp41,
remains incurable because it establishes a pool of long-lived and more recently the interface region between gp120 and gp41
memory CD4+ T cells in which replication-competent virus (reviewed in [6, 7]). Data from preclinical animal models but
persists as integrated proviral DNA. This latent cellular reser- also from early-phase clinical trials suggest that bNAbs were not
voir is established within days of virus exposure, even before only capable of reducing plasma viremia, demonstrating their
virus can be detected in peripheral blood [1]. Latently infected potent antiviral activity, but were also associated with a delay in
cells are invisible to the immune system as they lack active viral viral rebound after analytical ART interruption, suggesting an
replication and therefore do not express viral proteins on the cell effect of the bNAb therapy on the cellular viral reservoir [8–10].
surface. However, once suppressive ART is stopped, these cells Antibodies against HIV might be a promising new tool for viral
are capable of rapidly reigniting new rounds of infection. The eradication. Here, we will review the recent developments in
latent reservoir therefore presents a significant hurdle for cure the use of antibodies for HIV-1 therapy, specifically focusing on
approaches. Novel concepts for viral eradication strategies com- antibody-mediated strategies to target latently infected cells and
bine pharmacological induction of the latently infected cells to options to increase antibody efficacy for HIV cure approaches.
produce virus together with immune enhancing interventions
to enable the host to clear these cells. Over the last decade there THE TARGET: RECOGNITION OF THE CELLULAR HIV
RESERVOIR
has been significant progress in the identification and develop-
ment of broadly neutralizing antibodies (bNAbs) against HIV. The process of HIV Env expression on the cell surface during
The introduction of high-throughput single B-cell receptor viral replication is poorly understood, and several mechan-
sequencing technology using HIV-1 Env probes to identify isms have been proposed (reviewed in [11]). HIV buds from
HIV-1–specific B cells [2, 3], has generated a slew of extremely the plasma membrane, a process which exposes Env on the cell
potent bNAbs [2, 4, 5]. Numerous antibodies have been reported surface. The infected cell can be recognized and labeled by anti-
bodies in this transient/vulnerable state, which can lead to re-
cruitment of innate immune cell, such as natural killer (NK)
Correspondence: Boris Julg, Ragon Institute of MGH, MIT, and Harvard, 400 Technology Sq,
cells, in an Fc-dependent fashion and elimination of the in-
Cambridge, MA 02139 (BJULG@mgh.harvard.edu). fected cell by direct effector cell functions. Figure 1 shows the
The Journal of Infectious Diseases® 2021;223(S1):S22–31 surface of an HIV-infected lymphocyte decorated with Env as
© The Author(s) 2021. Published by Oxford University Press for the Infectious Diseases Society
of America. All rights reserved. For permissions, e-mail: journals.permissions@oup.com.
detected by the fluorescently labeled antibody 2G12. The ki-
DOI: 10.1093/infdis/jiaa165 netics of viral packaging on the surface are a battle between
S22 • jid 2021:223 (Suppl 1) • Rossignol et al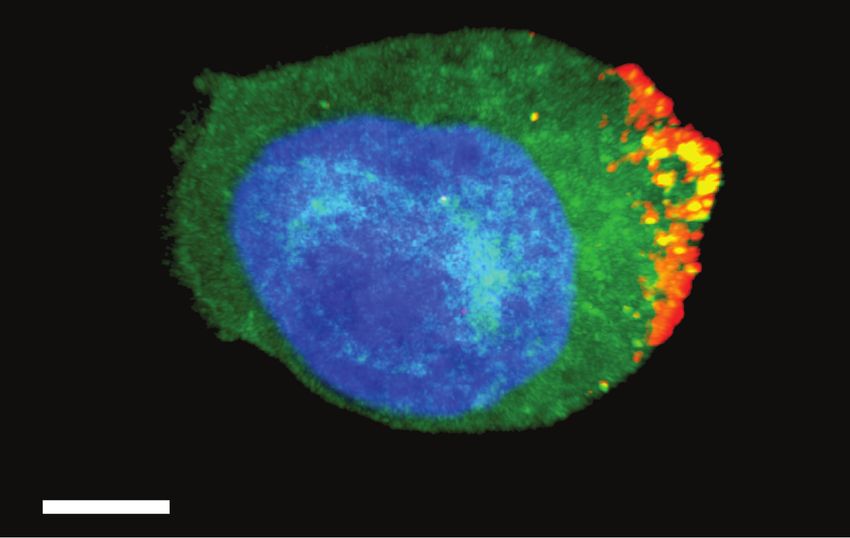
p24 spike. It is reasonable to assume that effector cells may react dif-
Env ferently to the single Fc domain presented by PGDM1400 than
DAPI to 3 presented by a fully occupied PGT121 Env spike, or 6 po-
tential Fcs presented by 2G12, because a threshold of sufficient
FcγR interactions needs to be met to activate the effector cells.
OPEN QUESTIONS ABOUT THE EPITOPE
LANDSCAPE ON REACTIVATED INFECTED CELLS
Although neutralizing antibodies specifically target functional
5 μm Env trimers, the antibody response induced by natural HIV-1
infection consists largely of nonneutralizing antibodies (nNabs)
that bind to gp120/gp41 monomers, gp120-depleted gp41
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
Figure 1. Distribution of human immunodeficiency virus (HIV) envelope gly- stumps or nonfunctional conformational variants of the trimer
coprotein (Env) on the cell surface. Env is distributed (red) on the surface of an
HIV NLAD8-infected (p24+; green) lymphocyte and is detected by the fluorescently
on the virion surface [21]. To what degree these nonfunctional
labeled broadly neutralizing antibody 2G12 (red). Bright puncta of p24+ and Env+ Env structures are exposed on the surface of reactivated or ac-
suggest virions budding from the plasma membrane. The image is a projection of tively infected cells is not known, but antibodies specific for
confocal sections through the entire thickness of the cell. Nuclear morphological
such antigens could play a role in reservoir targeting strategies
characteristics are shown by 4′,6-diamidino-2-phenylindole (DAPI) staining (blue).
because they might increase the overall breadth of cell recogni-
virus and host; host factors such as CD4 and tetherin proteins tion. Furthermore, with latency at a cellular level generally de-
work to retain viral epitopes on the cell surface to allow suffi- fined as a state of infection without viral production, it is likely
cient exposure to antibody recognition or inhibit the release of that immune activation is necessary to activate virus produc-
viruses [12], respectively, and are antagonized by, for example, tion so that Env may be exposed for targeting by antibodies.
the HIV protein Vpu or a membrane-proximal endocytosis Currently, latency reversal agents (LRAs) such as histone
motif of gp41, which counter these defense mechanisms to pro- deacetylase (HDAC) inhibitors, protein kinase C agonists, Toll-
mote efficient viral release [13–15]. Specifically, this process is like receptor (TLR) agonists, second mitochondrial-derived
unknown in the setting of viral reactivation from latency and activator of caspase mimetics [22], and cytokine superagonists
Env synthesis and processing might differ significantly from (ie, interleukin 15 [23, 24]) are being explored (reviewed in
productively infected cells. [25–29]). Although robust induction of viral replication in la-
tently infected cells is the goal, it is critical that the LRA does
TOPOLOGY AND STOICHIOMETRY OF INFECTED not interfere with the immune response itself, that is, via off-site
CELL RECOGNITION effects. It has been demonstrated for certain HDAC inhibitors,
The topology of Env expression on the surface of infected such as romidepsin, that NK cell and cytotoxic T-lymphocyte
cells is poorly understood. On the virion surface, electron cytolytic activity is significantly decreased in the presence of the
cryomicroscopy reveals the presence of approximately 14 copies LRA, at least in vitro [30–32]. Conversely, some compounds,
of Env trimers, the main target of neutralizing antibodies, dis- such as the protein kinase C agonist prostratin or the cytokine
tributed roughly isotropically [16]. Viral assembly and bud- interleukin 15, have been demonstrated to increase NK antiviral
ding might bring these Env trimers in close proximity to each activity [33, 34]. Conceptually, the ideal LRAs for antibody-
other on one hemisphere of the virion, increasing the density based HIV cure approaches would lead to high levels of Env on
of bound antibodies and therefore potentially increasing the the surface of reservoir cells under ART, thus exposing latently
Fc accessibility by innate effector cells. Indeed, most FcγRs infected cells to antibody recognition and subsequent elimina-
have a low affinity for immunoglobulin (Ig) G, and activation tion while also stimulating innate effector cell activity.
of effector cells and subsequent cytotoxicity is dependent on
clustering of FcγRs on the cell surface. Antibody-binding stoi- PRECLINICAL AND CLINICAL EXPERIENCE WITH
ANTIBODIES FOR HIV-1 THERAPY
chiometry to Env trimers seems to be important, because more
available Fc domains will result in sufficient activating signals to Animal Models
augment, for example, antibody-dependent cellular cytotoxicity The effects of bNAbs against plasma viremia, but also cell-
(ADCC) [17]. The binding stoichiometry of neutralizing anti- associated virus, have been documented in several preclinical
bodies vary from 1 antibody per Env spike (eg, the V2 glycan studies using mice and nonhuman primate models [35–41].
antibody PGDM1400 [18]), to the more typical 3 antibodies Intravenous infusion of PGT121 in rhesus macaques infected
per spike (eg, the V3 glycan antibody PGT121 [19]), to the V3 with simian-human immunodeficiency virus (SHIV) SF-162P3
antibody 2G12 antibody, which dimerizes by domain swapping resulted not only in rapid and profound suppression of plasma
[20], creating a potential occupancy of 6 antibodies per Env viral RNA, but also in substantial reductions of proviral DNA
Antibodies for HIV-1 Cure Strategies • jid 2021:223 (Suppl 1) • S23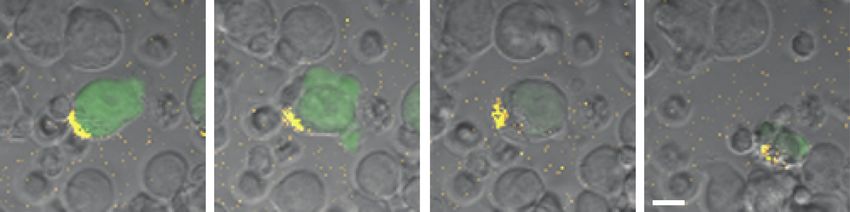
in peripheral blood, lymph nodes, and gastrointestinal mucosa (3BNC117 + 10–1074) in HIV -infected individuals receiving
[35]. In particular, the latter demonstrated that this bNAb had ART resulted in delayed viral rebound once ART was inter-
neutralizing activities but also seemed to clear infected cells, rupted [8–10]. Specifically, when the latter bNAb combination
suggesting an Fc-mediated mechanism. Indeed, in the human- was given to individuals with antibody-sensitive viral reservoirs,
ized mouse models, Bournazos and colleagues [40] demon- as 3 infusions of 30 mg/kg of each antibody every 3 weeks, viral
strated that enhanced antiviral potency of bNAbs was associated suppression was maintained for a median of 21 weeks. In con-
with preferential engagement of activating FcγRs by comparing trast, 2 individuals harboring 10–1074–resistant or 3BNC117-
the abilities of Fc-enhanced, knockout, and wild-type 3BNC117 resistant viruses rebounded early.
to reduce viral load. The degree of viral control after ART interruption was in
Furthermore, a rapid anti-infected cell effect by bNAbs was general variable between the studies, and antibody-resistant
demonstrated by Lu and colleagues [41], in a model meas- viruses emerged frequently. It therefore remains to be deter-
uring cell-associated RNA reduction in response to wild-type mined what effect bNAbs have on the size of the inducible viral
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
or Fc function knockout mutants that ablated effector function, reservoir. To improve antibody efficacy and reduce the risk for
as well as direct FcγR blocking. Further bNAb-directed anti- antibody therapeutic failure due to viral escape, triple-bNAb
infected cell effects were demonstrated in macaques; PGT121 cocktails, combining antibodies with complementary epitope
protected 50% of macaques from infection that were challenged target regions (eg, targeting the CD4bs, V3, and V2) are cur-
with SHIV SF162P3–infected splenocytes [42]. These animal rently being evaluated with the thought that this will elevate
data therefore confirmed the importance of Fc-mediated mech- the threshold for selecting viral variants with preexistent escape
anisms of infected cell clearance. Additional studies focused mutations or the inducing novel resistance mutations (clin-
on the direct antireservoir effects of bNAbs by performing ical trials NCT03205917 and NCT03721510). Following this
analytical treatment interruptions. For example, the combina- concept, a trispecific antibody (SAR441236), which combines
tion of the CD4bs antibody 3BNC117, the V3 glycan antibody the antigen-binding fragments (Fabs) of the CD4bs antibody
10–1074, and the V2 glycan antibody PG16 together with sev- VRC01, the membrane-proximal external region antibody
eral LRAs, abrogated viral rebound after washout of the anti- 10E8 and the V2 glycan antibody PGDM1400 is under clinical
bodies in 57% of HIV-infected humanized mice [43]. evaluation in HIV-infected viremic and ART-suppressed in-
Borducchi et al [44] treated rhesus macaques infected with dividuals (NCT03705169). Furthermore, multiple studies are
SHIV SF-162P3 with ART administered for 96 weeks, starting underway or in development to evaluate the combination of
7 days after infection. Under continued ART, macaques received bNAbs with LRAs and immunomodulators, such as TLR agon-
a TLR7 agonist (Vesatolimod), PGT121, or both, followed by ists and HDAC inhibitors (NCT03837756 and NCT03041012).
a washout period to allow PGT121 to be cleared before ART
cessation. After ART discontinuation, a subset of the macaques ANTIBODY FC FUNCTIONALITY IN HIV
(5 of 11) treated with PGT121 and TLR7 agonist did not re- Accumulating evidence demonstrates that antibody effector
bound. In addition, bNAb administration to macaques acutely functions play a critical role in conferring therapeutic efficacy
infected with SHIV AD8-EO led to controlled plasma viremia against HIV [40, 41, 46–48]. Indeed, the antibody Fc domain is
in a subset of animals, after washout of the bNAbs, in a CD8+ able to mediate a wide range of effector functions through in-
T-cell–dependent manner. Rapid reappearance of plasma vi- teraction with Fc receptors, complement proteins, and lectins.
remia occurred in these animals once CD8+ T cells were de- These functions include lysis by NK cells (ADCC), antibody-
pleted, suggesting that the bNAbs facilitated the emergence dependent complement deposition, trogocytosis (FcγR-
of potent CD8+ T-cell immunity [45]. It remains to be deter- dependent “nibbling” of the plasma membrane capable of
mined whether enhanced T-cell immunity will contribute to killing the opsonized cell [49, 50]), and phagocytosis by mono-
viral control during reservoir eradication strategies in humans,. cytes/macrophages, neutrophils, and dendritic cells (DCs) [51,
Nevertheless, the accumulating evidence for therapeutic effects 52] (Figure 2). Numerous reports demonstrate that a humoral
of bNAbs in animal models has led to a resurgence of interest in response to HIV with enhanced activity of Fc-dependent func-
evaluating bNAbs for therapeutic indications in humans. tions is critical in slowing HIV disease progression [53–57], and
enhanced Fc functionality is even correlated with the develop-
Clinical Data ment of bNAb responses [58, 59].
Several first in-human studies demonstrated that single admin-
ANTIBODY-DEPENDENT CELLULAR CYTOTOXICITY
istrations of bNAbs, including 3BNC117, the CD4bs antibody
VRC01, and 10–1074 or bNAb combinations, were able to re- ADCC is a mechanism for clearance of infected cells (Figure 3),
duce viral loads by several log10 in HIV-1–infected individuals and an enrichment of potent ADCC-mediating antibodies
not receiving ART. Furthermore, repeated dosing of single has been described in serum samples from HIV elite control-
bNAbs (eg, 3BNC117 or VRC01) or a dual-bNAb combination lers, rare individuals who are capable of spontaneous viremic
S24 • jid 2021:223 (Suppl 1) • Rossignol et alby engagement of the inhibitory FcγRIIb (reviewed in [66]).
Therefore, the balance of activating and inhibitory signals is
critical, whereas the topological arrangement of target (infected
Monocyte
cell) recognition and Fc presentation by antibodies is impor-
tant. Indeed, recognition of infected cells by bNAbs and sub-
NK cell sequent clearance by NK cells is highly heterologous [67–70].
Some groups report that infected primary cells seem to be more
susceptible to elimination by opsonization of V1-V2–targeting
bNabs PG9 and PG16, but not CD4bs antibodies [68], in con-
Cytokines
trast to target cells generated by infecting cell lines [67, 69].
Others, however show relatively high ADCC activity by CD4bs
antibody 3BNC117 against reactivated primary cells from ART-
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
treated individuals [71]. Reported differences are likely due to
confounding variables such as virus strains used and origin
(from infectious molecular clones or passaged primary virus),
target cells (primary or cell line), antibody Fc glycosylation dif-
ferences, and cellular and NK heterogeneity [72]. Furthermore,
discordant results have been reported based on the design of the
different ADCC assays [73].
Neutrophil
In principle, neutralization is not required for ADCC-
mediating antibodies, and, hypothetically, nNabs can bind to
infected cells and mediate Fc functionality. A comparison of
NO
the ADCC activity for a panel of 9 nNabs targeting various
epitopes, showed limited ability to recognize and eliminate in-
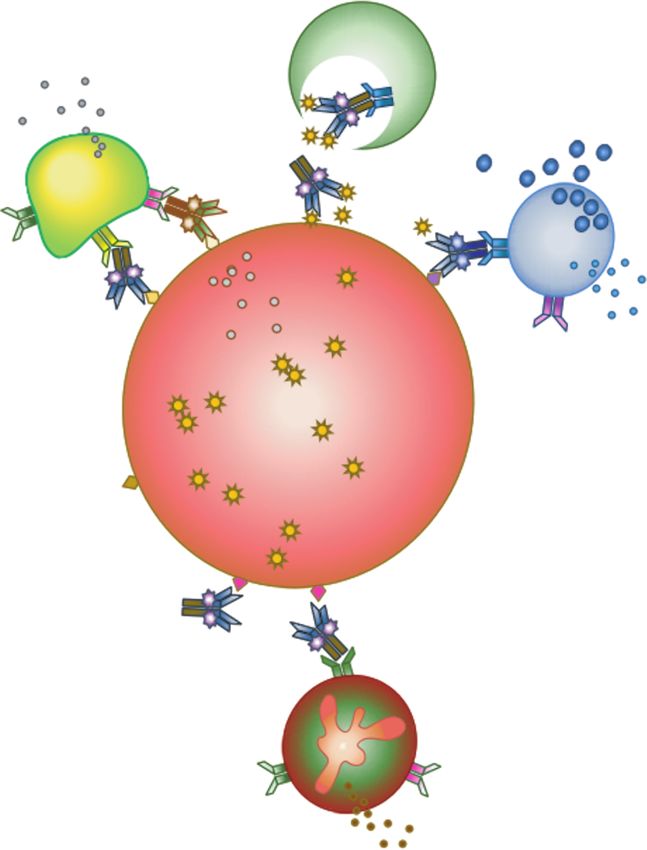
Figure 2. Fc-mediated antibody effector functions, depicting innate effector rec- fected peripheral blood mononuclear cells from ART-treated
ognition of immune complexes of antibodies and infected cells. Antibody-opsonized patients, compared with bNAbs [71]. One explanation might be
cells display the Fc in a conformation recognized by innate effectors, such as mono- that current bNAbs have been selected for high-affinity binding
cytes, natural killer (NK) cells, and neutrophils, which are capable of responses that
vary by effector type. Abbreviation: NO, Nitric Oxide.
to the Env trimer, but the same efforts have not been applied
to the identification of nNabs directed against nontrimer tar-
gets. Investing more in the identification of novel antibodies
control [54, 56, 60–64], but they also have been associated with that bind nontrimer Env structures with highest affinity might
antibody-mediated protection against infection in the RV144 result in a new class of therapeutics that can be used in cellular
HIV vaccine study [65]. ADCC is dependent on the ability of reservoir targeting strategies. Specifically, sequential antibody
the antibody to recognize infected cells and present the Fc do- binding to different epitopes may act in concert to enhance
main in a conformation accessible to FcγRIIIa (CD16a) on NK ADCC [74–76]. For example, binding of certain CD4bs and
cells. Activation of NK degranulation depends on cross-linking glycan-targeting bNAbs to the Env trimer results in exposure
of FcγRIIIa, leading to intracellular signaling events that led of CD4-induced epitopes [77], creating a hypothetical scenario
to NK cell activation and proliferation, and it can be inhibited in which a bNAb could enhance ADCC by exposing epitopes to
1.3 4.3 20.3 58.7
Figure 3. Time-lapse confocal microscopy of a live human immunodeficiency virus (HIV)–infected (green) cell and antibody-dependent cellular cytotoxicity (ADCC) by natural
killer (NK) cells. Purified primary NK cells (smaller cells) were coincubated with HIV-1 JR-CSF–infected CEM cells that express green fluorescent protein (GFP) when infected.
Envelope glycoprotein recognition was mediated by a mix of AF647-conjugated 2G12 and unlabeled 2G12 (1:5 ratio). NK cells appear to be associated with the yellow en-
velope patch (first panel), the cell blebs (second panel), show diminished GFP expression (third panel), and are ultimately destroyed by NK cells (fourth panel). Numbers
indicate time in minutes after coculture of effectors and targets; scale bar represents 10 μm.
Antibodies for HIV-1 Cure Strategies • jid 2021:223 (Suppl 1) • S25the patient’s own ADCC-capable antibody response or to pas- Amino acid sequence modifications of the IgG1 Fc domain
sively infused antibody therapeutics. focus on increasing or decreasing affinity for FcγRs. The most
commonly used Fc mutation, M428L/N434S (commonly short-
PHAGOCYTOSIS ened to LS), increases antibody half-life in vivo by increasing
Antibody-dependent phagocytosis (ADCP) (reviewed in [78]) affinity for the neonatal Fc receptor [91] while retaining binding
is another Fc function capable of clearance of HIV virions and and functional capacity to FcγRIIIA and C1q [92]. Currently,
HIV-infected cells. This function is principally carried out multiple antibodies with LS mutations are being explored clin-
via the FcγRIIa on monocytes, macrophages, neutrophils, eo- ically in HIV [93, 94]. Other Fc mutations can increase (or
sinophils, mast cells, and DCs. Phagocytic activity occurs in reduce) Fc functionality [86]. Examples include Fc mutations
response to FcγRIIa stimulation and is inhibited by FcγRIIb. S298A/E333A/K334A (AAA) and G236A/S239D/A330L/I332E
Similar to findings with ADCC, ADCP activity is higher in HIV (GASDALIE), which increase ADCC activity by increasing
elite controllers [57]. ADCP can directly phagocytose antibody- affinity for FcγRIIIA [40, 95–97], (reviewed in [85]). In ad-
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
opsonized virions [79], and it can also result in antigen pres- dition to increasing affinity for FcγRIIIA, the GASDALIE Fc
entation, and interferon α signaling in DCs. Macrophages are mutations exhibit >10-fold increases in affinity for FcγRIIA as
capable of engulfing whole infected cells, but this can lead to well and slightly decreases affinity for the inhibitory receptor
productive infection in the macrophage [80]. ADCP activity FcγRIIB [40]. The use of function-enhanced antibodies in the
has certain advantages over ADCC, for example, in lymph fight against HIV is still in nascent stages, as demonstrated by
nodes, which are considered a sanctuary site of the viral res- protection studies [40, 96]; collectively, however, these data
ervoir [81], NK cells are infrequently found, whereas macro- highlight the potential for modulation of antireservoir func-
phages and neutrophils are more abundant [82]. In monoclonal tional activity.
antibody-based HIV cure approaches, Fc domains that stimu- Similar to amino acid mutations, the glycosylation of the Fc
late ADCP add another dimension to the fight against the res- domain modulates FcγR affinity and result in altered Fc func-
ervoir, through the recruitment of effectors with inflammatory tionality (reviewed in [98]). Deglycosylation has been demon-
and signaling potential. strated to abolish FcγR binding [99], and ADCC is significantly
increased in antibodies lacking core fucose [85, 100–105].
FC-ENGINEERED ANTIBODIES Hypergalactosylation (G2 glycans) improves ADCP [106] and
Using antibodies as monoclonal therapeutics presents an op- ADCC to a lesser relative extent than afucosylation, but it can
portunity to optimize the Fc domain, resulting in enhanced Fc be synergistically combined with afucosylation for increased
functionality over IgG1 [83]. Although in vivo data are limited FcγRIIIa affinity [105]. Approaches under development include
in the HIV field, there are multiple Fc-engineered monoclonal glycoengineering antibodies (eg, by using specifically engin-
antibodies approved for cancer [84, 85]. Fc enhancement can be eered production cell lines, tobacco plants, etc) and applying
accomplished in 3 principal ways: by altering the isotype, the chemoenzymatic approaches to generate antibodies with specific
amino acid sequence of the Fc domain, or the Fc glycans, all glycan signatures (reviewed in [107]), and the first monoclonal
of which modulate affinity to FcγRs and complement protein antibodies (eg, with nonfucosylated glycan mixtures) have been
C1q [85, 86]. Most antibodies in clinical use for treatment of approved for clinical use to eliminate blast cells during lym-
autoimmune disorders or cancers are IgG1, but there is growing phoma [100, 108]. As the principles behind antibody-mediated
appreciation of the importance of other isotypes. Passive immu- elimination of cancer cells are similar to what is attempted with
nization of monoclonal IgM and IgA at mucosal surfaces has antibody-mediated clearance of HIV-infected cells, the poten-
been demonstrated to offer superior protection against mucosal tial for functional enhancement in vivo by glycoengineering of
transmission of SHIV in macaques, compared with IgG1, poten- antireservoir antibodies should be explored.
tially owing to the superior viral capture and avidity of IgM [87,
VIRAL DIVERSITY IN THE LATENT RESERVOIR
88]. IgG3 variants of the V2-directed bNAb CAP256-VRC26.25
exhibit increased neutralization potency and breadth, as well as Env sequence diversity in the HIV reservoir is a major obstacle
increased ADCP and antibody-dependent complement deposi- for antibody-based cure approaches. ART cessation in HIV-
tion compared with IgG1 [89]. IgG1 and IgG3 are topologically infected patients leads to a rebound of diverse viruses from
similar in structure, with the exception of the extended hinge multiple tissue sources and compartments [109], with the con-
region of IgG3 separating the Fab and Fc domains. The length sequence that reactivated viral populations are being dispersed
of this flexible domain modulates phagocytic activity [90] but via the blood and repopulate tissues throughout the body [110].
may contribute to decreased antibody half-life of IgG3 rela- Although there is evidence for viral compartmentalization—for
tive to IgG1. However, the increased Fab and Fc functions of example, in the central nervous system (CNS) compared with
IgG3 could potentially complement IgG1 in the development of blood [111]—a recent study performing rapid autopsies in de-
antireservoir monoclonal antibody combinations. ceased HIV-infected individuals combined with tissue reservoir
S26 • jid 2021:223 (Suppl 1) • Rossignol et alanalysis found evidence of identical intact full-length Env pro- antibody resistance mutations, inaccessibility of antibodies to
viruses within and across tissues, but the differences across (for example) the CNS, and limitations in innate effector cell
compartments varied between individuals [110]. Beside the fact availability within certain tissue compartments might represent
that intravenously administered antibodies do not access the obstacles that need to be overcome. Future studies in this field
CNS owing to the blood-brain barrier, the CNS or potentially may benefit from advances in LRA development, resulting in
other tissue compartments could harbor a reservoir of bNAb- products that are capable of robustly inducing viral activation
resistant viruses that could migrate to blood, therefore refueling and increasing infected cell visibility for the immune system,
systemic viremia. as well as from novel antibody Fc modifications that robustly
To test the susceptibility of peripheral blood reservoir viruses boost effector functions.
to different bNAbs, Ren et al [70] characterized the ability of 14
anti-Env antibodies to neutralize virus outgrown from infected Notes
peripheral blood mononuclear cells from 36 ART-treated HIV- Acknowledgments. We thank Hacheming Compere for
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
infected individuals but also assessed the ability of these anti- proofreading the manuscript.
bodies to bind to cells infected with the same viruses ex vivo. Financial support. This work was supported by the National
They demonstrated that neutralization susceptibility and in- Institute of Health (grant AI138790 to B. J.).
fected cell-binding efficacy was quite heterogenous, with single- Supplement sponsorship. This supplement is sponsored by
antibody neutralization breadths ranging from 0% to 64% of the Harvard University Center for AIDS Research (CFAR), an
viruses neutralized (80% inhibitory concentration, ≤10 μg/mL) NIH funded program (P30 AI060354), and the Ragon Institute
and from 0% to 89% for binding of infected cells [70]. of MGH, MIT and Harvard. The content is solely the responsi-
Identifying viral resistance in patients before antibody treat- bility of the authors and does not necessarily represent the offi-
ment is therefore critical to rational design of antibody com- cial views of the National Institutes of Health.
bination regimens for HIV reservoir eradication strategies. Potential conflicts of interest. All authors: No reported
Defining reservoir susceptibility using quantitative viral out- conflicts. All authors have submitted the ICMJE Form for
growth assays and testing of neutralization sensitivity of these Disclosure of Potential Conflicts of Interest. Conflicts that the
viruses in standard TZM-bl assays is time consuming and ex- editors consider relevant to the content of the manuscript have
pensive (reviewed in [112]). Furthermore, this approach does been disclosed.
not guarantee that all viral variants that could emerge are con-
sidered, because not every replication-competent provirus can References
and will be induced for outgrowth under in vitro conditions 1. Whitney JB, Hill AL, Sanisetty S, et al. Rapid seeding of the
[113]. Several computational concepts have therefore been viral reservoir prior to SIV viraemia in rhesus monkeys.
developed to use machine learning algorithms for predicting Nature 2014; 512:74–7.
bNAb susceptibility based on patient-derived proviral Env 2. Walker LM, Phogat SK, Chan-Hui PY, et al; Protocol G
sequences [114–117], and the models have achieved high pre- Principal Investigators. Broad and potent neutralizing anti-
diction accuracy that might facilitate the selection of personal- bodies from an African donor reveal a new HIV-1 vaccine
ized antibody regimens for cure approaches. Nevertheless, we target. Science 2009; 326:285–9.
will need a better understanding of the landscape of neutraliza- 3. Wu X, Yang ZY, Li Y, et al. Rational design of envelope iden-
tion resistance of the HIV reservoir across tissue compartments tifies broadly neutralizing human monoclonal antibodies to
and methods to robustly assess these before antibody therapy. HIV-1. Science 2010; 329:856–61.
Until then, selection of bNabs/bNAb combinations with the 4. Walker LM, Huber M, Doores KJ, et al; Protocol G Principal
broadest viral coverage possible may still be advantageous. Investigators. Broad neutralization coverage of HIV by mul-
tiple highly potent antibodies. Nature 2011; 477:466–70.
CONCLUSIONS
5. Scheid JF, Mouquet H, Feldhahn N, et al. Broad diversity
Antibodies have the potential to harness the innate immune of neutralizing antibodies isolated from memory B cells in
system against the reactivated HIV reservoir and result in HIV-infected individuals. Nature 2009; 458:636–40.
viral remission. The ideal antibody combination for reservoir 6. Burton DR, Mascola JR. Antibody responses to envelope
targeting recognizes all reactivated cells and efficiently medi- glycoproteins in HIV-1 infection. Nat Immunol 2015;
ates destruction through the recruitment of innate effector 16:571–6.
cells. This will require exquisite breadth in viral coverage 7. Sok D, Burton DR. Recent progress in broadly neutralizing
combined with the ability to bind to Env structures on reacti- antibodies to HIV. Nat Immunol 2018; 19:1179–88.
vated cells while allowing easy accessibility of the Fc domain 8. Bar KJ, Sneller MC, Harrison LJ, et al. Effect of HIV anti-
to FcγRs. The vast Env sequence diversity across the globe body VRC01 on viral rebound after treatment interruption.
but also within each host’s reservoir, with potential archived N Engl J Med 2016; 375:2037–50.
Antibodies for HIV-1 Cure Strategies • jid 2021:223 (Suppl 1) • S279. Mendoza P, Gruell H, Nogueira L, et al. Combination 24. Mcbrien JB, Mavigner M, Franchitti L, et al. Robust and
therapy with anti-HIV-1 antibodies maintains viral sup- persistent reactivation of SIV and HIV by N-803 and deple-
pression. Nature 2018; 561:479–84. tion of CD8+ cells. Nature 2020; 578:154–9.
10. Scheid JF, Horwitz JA, Bar-On Y, et al. HIV-1 antibody 25. Sengupta S, Siliciano RF. Targeting the latent reservoir for
3BNC117 suppresses viral rebound in humans during treat- HIV-1. Immunity 2018; 48:872–95.
ment interruption. Nature 2016; 535:556–60. 26. Sadowski I, Hashemi FB. Strategies to eradicate HIV from
11. Freed EO. HIV-1 assembly, release and maturation. Nat Rev infected patients: elimination of latent provirus reservoirs.
Microbiol 2015; 13:484–96. Cell Mol Life Sci 2019; 76:3583–600.
12. Neil SJ, Zang T, Bieniasz PD. Tetherin inhibits retrovirus 27. Campbell GR, Spector SA. DIABLO/SMAC mimetics se-
release and is antagonized by HIV-1 Vpu. Nature 2008; lectively kill HIV-1-infected resting memory CD4+ T
451:425–30. cells: a potential role in a cure strategy for HIV-1 infection.
13. Arias JF, Heyer LN, von Bredow B, et al. Tetherin antag- Autophagy 2019; 15:744–6.
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
onism by Vpu protects HIV-infected cells from antibody- 28. Macedo AB, Novis CL, Bosque A. Targeting cellular and
dependent cell-mediated cytotoxicity. Proc Natl Acad Sci U tissue HIV reservoirs with Toll-like receptor agonists. Front
S A 2014; 111:6425–30. Immunol 2019; 10:2450.
14. Bredow B von, Arias JF, Heyer LN, et al. Envelope glyco- 29. Bashiri K, Rezaei N, Nasi M, Cossarizza A. The role of la-
protein internalization protects human and simian immu- tency reversal agents in the cure of HIV: a review of current
nodeficiency virus-infected cells from antibody-dependent data. Immunol Lett 2018; 196:135–9.
cell-mediated cytotoxicity. J Virol 2015; 89:10648–55. 30. Kelly-Sell MJ, Kim YH, Straus S, et al. The histone
15. Pham TNQ, Lukhele S, Dallaire F, Perron G, Cohen ÉA. deacetylase inhibitor, romidepsin, suppresses cellular im-
Enhancing virion tethering by BST2 sensitizes productively mune functions of cutaneous T-cell lymphoma patients.
and latently HIV-infected T cells to ADCC mediated by Am J Hematol 2012; 87:354–60.
broadly neutralizing antibodies. Sci Rep 2016; 6:1–15. 31. Pace M, Williams J, Kurioka A, et al. Histone deacetylase in-
16. Zhu P, Liu J, Bess J Jr, et al. Distribution and three-dimen- hibitors enhance CD4 T cell susceptibility to NK cell killing
sional structure of AIDS virus envelope spikes. Nature but reduce NK cell function. PLoS Pathog 2016; 12:1–22.
2006; 441:847–52. 32. Jones RB, O’Connor R, Mueller S, et al. Histone deacetylase
17. Klein JS, Webster A, Gnanapragasam PN, Galimidi RP, inhibitors impair the elimination of HIV-infected cells by
Bjorkman PJ. A dimeric form of the HIV-1 antibody 2G12 cytotoxic T-lymphocytes. PLoS Pathog 2014; 10:e1004287.
elicits potent antibody-dependent cellular cytotoxicity. 33. Garrido C, Spivak AM, Soriano-Sarabia N, et al. HIV
AIDS 2010; 24:1633–40. latency-reversing agents have diverse effects on natural
18. Lee JH, Andrabi R, Su CY, et al. A broadly neutralizing an- killer cell function. Front Immunol 2016; 7:1–11.
tibody targets the dynamic HIV envelope trimer apex via a 34. Garrido C, Abad-Fernandez M, Tuyishime M, et al.
long, rigidified, and anionic β-hairpin structure. Immunity Interleukin-15-stimulated natural killer cells clear HIV-
2017; 46:690–702. 1-infected cells following latency reversal ex vivo. J Virol
19. Garces F, Lee JH, Val N de, et al. Affinity maturation of a 2018; 92:e00235–18.
potent family of HIV antibodies is primarily focused on 35. Barouch DH, Whitney JB, Moldt B, et al. Therapeutic efficacy
accommodating or avoiding glycans. Immunity 2015; of potent neutralizing HIV-1-specific monoclonal antibodies
43:1053–63. in SHIV-infected rhesus monkeys. Nature 2013; 503:224–8.
20. Wu Y, West AP Jr, Kim HJ, Thornton ME, Ward AB, 36. Horwitz JA, Halper-Stromberg A, Mouquet H, et al. HIV-1
Bjorkman PJ. Structural basis for enhanced HIV-1 neutrali- suppression and durable control by combining single broadly
zation by a dimeric immunoglobulin G form of the glycan- neutralizing antibodies and antiretroviral drugs in human-
recognizing antibody 2G12. Cell Rep 2013; 5:1443–55. ized mice. Proc Natl Acad Sci U S A 2013; 110:16538–43.
21. Moore PL, Crooks ET, Porter L, et al. Nature of nonfunc- 37. Julg B, Pegu A, Abbink P, et al. Virological control by the
tional envelope proteins on the surface of human immuno- CD4-binding site antibody N6 in simian-human immu-
deficiency virus type 1. J Virol 2006; 80:2515–28. nodeficiency virus-infected rhesus monkeys. j Virol 2017;
22. Nixon CC, Mavigner M, Sampey GC, et al. Systemic HIV 91:1–15.
and SIV latency reversal via non-canonical NF-κB signalling 38. Shingai M, Nishimura Y, Klein F, et al. Antibody-mediated
in vivo. Nature 2019; 578:160–5. immunotherapy of macaques chronically infected with
23. Jones RB, Mueller S, O’Connor R, et al. A subset of latency- SHIV suppresses viraemia. Nature 2013; 503:277–80.
reversing agents expose HIV-infected resting CD4+ T-cells 39. Klein F, Halper-Stromberg A, Horwitz JA, et al. HIV therapy
to recognition by cytotoxic T-lymphocytes. PLoS Pathog by a combination of broadly neutralizing antibodies in hu-
2016; 12:e1005545. manized mice. Nature 2012; 492:118–22.
S28 • jid 2021:223 (Suppl 1) • Rossignol et al40. Bournazos S, Klein F, Pietzsch J, Seaman MS, 56. Ackerman ME, Mikhailova A, Brown EP, et al. Polyfunctional
Nussenzweig MC, Ravetch JV. Broadly neutralizing anti- HIV-specific antibody responses are associated with spon-
HIV-1 antibodies require Fc effector functions for in vivo taneous HIV control. PLoS Pathog 2016; 12:1–14.
activity. Cell 2014; 158:1243–53. 57. Ackerman ME, Dugast AS, McAndrew EG, et al. Enhanced
41. Lu CL, Murakowski DK, Bournazos S, et al. Enhanced phagocytic activity of HIV-specific antibodies correlates
clearance of HIV-1-infected cells by broadly neutralizing with natural production of immunoglobulins with skewed
antibodies against HIV-1 in vivo. Science 2016; 352:1001–4. affinity for FcγR2a and FcγR2b. J Virol 2013; 87:5468–76.
42. Parsons MS, Lloyd SB, Lee WS, et al. Partial efficacy of a 58. Lofano G, Gorman MJ, Yousif AS, et al. Antigen-specific
broadly neutralizing antibody against cell-associated SHIV antibody Fc glycosylation enhances humoral immunity
infection. Sci Transl Med 2017; 9: eaaf1483. via the recruitment of complement. Sci Immunol 2018;
43. Halper-Stromberg A, Lu CL, Klein F, et al. Broadly neutral- 3:eaat7796.
izing antibodies and viral inducers decrease rebound from 59. Richardson SI, Chung AW, Natarajan H, et al. HIV-specific
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
HIV-1 latent reservoirs in humanized mice. Cell 2014; Fc effector function early in infection predicts the develop-
158:989–99. ment of broadly neutralizing antibodies. PLoS Pathog 2018;
44. Borducchi EN, Liu J, Nkolola JP, et al. Antibody and TLR7 14:1–25.
agonist delay viral rebound in SHIV-infected monkeys. 60. Madhavi V, Wren LH, Center RJ, et al. Breadth of HIV-1
Nature 2018; 563:360–4. Env-specific antibody-dependent cellular cytotoxicity:
45. Nishimura Y, Gautam R, Chun TW, et al. Early antibody relevance to global HIV vaccine design. AIDS 2014;
therapy can induce long-lasting immunity to SHIV. Nature 28:1859–70.
2017; 543:559–63. 61. Madhavi V, Wines BD, Amin J, et al. HIV-1 Env- and Vpu-
46. Hessell AJ, Hangartner L, Hunter M, et al. Fc receptor but specific antibody-dependent cellular cytotoxicity responses
not complement binding is important in antibody protec- associated with elite control of HIV. J Virol 2017; 91:1–16.
tion against HIV. Nature 2007; 449:101–4. 62. Lambotte O, Pollara J, Boufassa F, et al. High antibody-
47. Hessell AJ, Poignard P, Hunter M, et al. Effective, low-titer dependent cellular cytotoxicity responses are correlated
antibody protection against low-dose repeated mucosal with strong CD8 T cell viral suppressive activity but not
SHIV challenge in macaques. Nat Med 2009; 15:951–4. with B57 status in HIV-1 elite controllers. PLoS ONE 2013;
48. Flerin NC, Bardhi A, Zheng JH, et al. Establishment of a 8:1–5.
novel humanized mouse model to investigate in vivo activa- 63. Alsahafi N, Ding S, Richard J, et al. Nef proteins from HIV-1
tion and depletion of patient-derived HIV latent reservoirs. elite controllers are inefficient at preventing antibody-
J Virol 2019; 93:1–18. dependent cellular cytotoxicity. J Virol 2016; 90:2993–3002.
49. Taylor RP, Lindorfer MA. Fcγ-receptor-mediated 64. Ackerman ME, Crispin M, Yu X, et al. Natural variation in
trogocytosis impacts mAb-based therapies: historical prec- Fc glycosylation of HIV-specific antibodies impacts anti-
edence and recent developments. Blood 2015; 125:762–6. viral activity. J Clin Invest 2013; 123:2183–92.
50. Richardson SI, Crowther C, Mkhize NN, Morris L. 65. Haynes BF, Gilbert PB, McElrath MJ, et al. Immune-
Measuring the ability of HIV-specific antibodies to mediate correlates analysis of an HIV-1 vaccine efficacy trial. N Engl
trogocytosis. J Immunol Methods 2018; 463:71–83. J Med 2012; 366:1275–86.
51. Bournazos S, Ravetch JV. Diversification of IgG effector 66. Nimmerjahn F, Ravetch JV. Anti-inflammatory actions of
functions. Int Immunol 2017; 29:303–10. intravenous immunoglobulin. Annu Rev Immunol 2008;
52. Parsons MS, Chung AW, Kent SJ. Importance of Fc-mediated 26:513–33.
functions of anti-HIV-1 broadly neutralizing antibodies. 67. Bruel T, Guivel-Benhassine F, Amraoui S, et al. Elimination
Retrovirology 2018; 15:1–12. of HIV-1-infected cells by broadly neutralizing antibodies.
53. Baum LL, Cassutt KJ, Knigge K, et al. HIV-1 gp120-specific Nat Commun 2016; 7:10844.
antibody-dependent cell-mediated cytotoxicity correlates with 68. Mujib S, Liu J, Rahman AKMN, et al. Comprehensive cross-
rate of disease progression. J Immunol 1996; 157:2168–73. clade characterization of antibody-mediated recognition,
54. Lambotte O, Ferrari G, Moog C, et al. Heterogeneous complement-mediated lysis, and cell-mediated cytotoxicity
neutralizing antibody and antibody-dependent cell cyto- of HIV-1 envelope-specific antibodies toward eradication
toxicity responses in HIV-1 elite controllers. AIDS 2009; of the HIV-1 reservoir. J Virol 2017; 91:1–23.
23:897–906. 69. Bredow B von, Arias JF, Heyer LN, et al. Comparison of
55. Huang J, Kang BH, Ishida E, et al. Identification of a CD4- antibody-dependent cell-mediated cytotoxicity and virus
binding-site antibody to HIV that evolved near-pan neu- neutralization by HIV-1 Env-specific monoclonal anti-
tralization breadth. Immunity 2016; 45:1108–21. bodies. J Virol 2016; 90:6127–39.
Antibodies for HIV-1 Cure Strategies • jid 2021:223 (Suppl 1) • S2970. Ren Y, Korom M, Truong R, et al. Susceptibility to neutral- 85. Saunders KO. Conceptual approaches to modulating an-
ization by broadly neutralizing antibodies generally correl- tibody effector functions and circulation half-life. Front
ates with infected cell binding for a panel of clade B HIV Immunol 2019; 10:1–20.
reactivated from latent reservoirs. J Virol 2018; 92:1–20. 86. Wang X, Mathieu M, Brezski RJ. IgG Fc engineering to
71. Bruel T, Guivel-Benhassine F, Lorin V, et al. Lack of ADCC modulate antibody effector functions. Protein Cell 2018;
breadth of human nonneutralizing anti-HIV-1 antibodies. J 9:63–73.
Virol 2017; 91:1–19. 87. Gong S, Tomusange K, Kulkarni V, et al. Anti-HIV IgM
72. Bernard NF, Kiani Z, Tremblay-McLean A, Kant SA, protects against mucosal SHIV transmission. AIDS 2018;
Leeks CE, Dupuy FP. Natural killer (NK) cell education 32:F5–F13.
differentially influences HIV antibody-dependent NK cell 88. Watkins JD, Sholukh AM, Mukhtar MM, et al; CAVD Project
activation and antibody-dependent cellular cytotoxicity. Group. Anti-HIV IgA isotypes: differential virion capture
Front Immunol 2017; 8:1–11. and inhibition of transcytosis are linked to prevention of mu-
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
73. Lewis GK, Ackerman ME, Scarlatti G, et al. Knowns and cosal R5 SHIV transmission. AIDS 2013; 27:F13–20.
unknowns of assaying antibody-dependent cell-mediated 89. Richardson SI, Lambson BE, Crowley AR, et al. IgG3 en-
cytotoxicity against HIV-1. Front Immunol 2019; 10:1–12. hances neutralization potency and Fc effector function of
74. Anand SP, Prévost J, Baril S, et al. Two families of Env anti- an HIV V2-specific broadly neutralizing antibody. PLoS
bodies efficiently engage Fc-gamma receptors and elimi- Pathog 2019; 15:e1008064.
nate HIV-1-infected cells. J Virol 2019; 93:1–17. 90. Chu TH, Crowley AR, Backes I, et al. Hinge length con-
75. Pollara J, Bonsignori M, Moody MA, et al. HIV-1 vaccine- tributes to the phagocytic activity of HIV-specific IgG1 and
induced C1 and V2 Env-specific antibodies synergize for IgG3 antibodies. PLoS Pathog 2020; 16:e1008083.
increased antiviral activities. J Virol 2014; 88:7715–26. 91. Zalevsky J, Chamberlain AK, Horton HM, et al. Enhanced
76. Davis-Gardner ME, Gardner MR, Alfant B, Farzan M. antibody half-life improves in vivo activity. Nat Biotechnol
eCD4-Ig promotes ADCC activity of sera from HIV-1- 2010; 28:157–9.
infected patients. PLoS Pathog 2017; 13:1–16. 92. Booth BJ, Ramakrishnan B, Narayan K, et al. Extending
77. Derking R, Ozorowski G, Sliepen K, et al. Comprehensive human IgG half-life using structure-guided design. MAbs
antigenic map of a cleaved soluble HIV-1 envelope trimer. 2018; 10:1098–110.
PLoS Pathog 2015; 11:1–22. 93. Caskey M. Broadly neutralizing antibodies for the treat-
78. Tay MZ, Wiehe K, Pollara J. Antibody dependent cel- ment and prevention of HIV infection. Curr Opin HIV
lular phagocytosis in antiviral immune responses. Front AIDS 2020; 15:49–55.
Immunol 2019; 10:1–18. 94. Gaudinski MR, Coates EE, Houser KV, et al. Safety and
79. Tay MZ, Liu P, Williams LTD, et al. Antibody-mediated pharmacokinetics of the Fc-modified HIV-1 human mono-
internalization of infectious HIV-1 virions differs among clonal antibody VRC01LS: a phase 1 open-label clinical trial
antibody isotypes and subclasses. PLoS Pathog 2016; in healthy adults. PLoS Med 2018; 15:1–20.
12:1–25. 95. Shields RL, Namenuk AK, Hong K, et al. High resolution
80. Baxter AE, Russell RA, Duncan CJ, et al. Macrophage infec- mapping of the binding site on human IgG1 for Fc gamma
tion via selective capture of HIV-1-infected CD4+ T cells. RI, Fc gamma RII, Fc gamma RIII, and FcRn and design of
Cell Host Microbe 2014; 16:711–21. IgG1 variants with improved binding to the Fc gamma R. J
81. Fukazawa Y, Lum R, Okoye AA, et al. B cell follicle sanc- Biol Chem 2001; 276:6591–604.
tuary permits persistent productive simian immunodefi- 96. Santra S, Tomaras GD, Warrier R, et al. Human non-
ciency virus infection in elite controllers. Nat Med 2015; neutralizing HIV-1 envelope monoclonal antibodies limit
21:132–9. the number of founder viruses during SHIV mucosal infec-
82. Sips M, Krykbaeva M, Diefenbach TJ, et al. Fc receptor- tion in rhesus macaques. PLoS Pathog 2015; 11:1–38.
mediated phagocytosis in tissues as a potent mechanism for 97. Ahmed AA, Keremane SR, Vielmetter J, Bjorkman PJ.
preventive and therapeutic HIV vaccine strategies. Mucosal Structural characterization of GASDALIE Fc bound to the
Immunol 2016; 9:1584–95. activating Fc receptor FcγRIIIa. J Struct Biol 2016; 194:78–89.
83. Huang Y, Ferrari G, Alter G, et al. Diversity of antiviral IgG 98. Li W, Zhu Z, Chen W, Feng Y, Dimitrov DS. Crystallizable
effector activities observed in HIV-infected and vaccinated fragment glycoengineering for therapeutic antibodies de-
subjects. J Immunol 2016; 197:4603–12. velopment. Front Immunol 2017; 8:1554.
84. Kellner C, Otte A, Cappuzzello E, Klausz K, Peipp M. 99. Lund J, Tanaka T, Takahashi N, Sarmay G, Arata Y, Jefferis R.
Modulating cytotoxic effector functions by Fc engineering A protein structural change in aglycosylated IgG3 correlates
to improve cancer therapy. Transfus Med Hemother 2017; with loss of huFc gamma R1 and huFc gamma R111 binding
44:327–36. and/or activation. Mol Immunol 1990; 27:1145–53.
S30 • jid 2021:223 (Suppl 1) • Rossignol et al100. Beck A, Reichert JM. Marketing approval of mogamulizumab: a triumph for 109. Rothenberger MK, Keele BF, Wietgrefe SW, et al. Large number of rebounding/
glyco-engineering. MAbs 2012; 4:419–25. founder HIV variants emerge from multifocal infection in lymphatic tissues after
101. Boesch AW, Kappel JH, Mahan AE, et al. Enrichment of high affinity sub- treatment interruption. Proc Natl Acad Sci U S A 2015; 112:E1126–34.
classes and glycoforms from serum-derived IgG using FcγRs as affinity ligands. 110. Chaillon A, Gianella S, Dellicour S, et al. HIV persists throughout deep tis-
Biotechnol Bioeng 2018; 115:1265–78. sues with repopulation from multiple anatomical sources. J Clin Invest 2020;
102. Li T, DiLillo DJ, Bournazos S, Giddens JP, Ravetch JV, Wang LX. Modulating 130:1699–712.
IgG effector function by Fc glycan engineering. Proc Natl Acad Sci U S A 2017; 111. Stefic K, Chaillon A, Bouvin-Pley M, et al. Probing the compartmentalization of
114:3485–90. HIV-1 in the central nervous system through its neutralization properties. PLoS
103. Pereira NA, Chan KF, Lin PC, Song Z. The “less-is-more” in therapeutic anti- One 2017; 12:1–22.
bodies: afucosylated anti-cancer antibodies with enhanced antibody-dependent 112. Wang Z, Simonetti FR, Siliciano RF, Laird GM. Measuring replication competent
cellular cytotoxicity. MAbs 2018; 10:693–711. HIV-1: advances and challenges in defining the latent reservoir. Retrovirol 2018;
104. Houde D, Peng Y, Berkowitz SA, Engen JR. Post-translational modifications dif- 15:1–9.
ferentially affect IgG1 conformation and receptor binding. Mol Cell Proteomics 113. Ho YC, Shan L, Hosmane NN, et al. Replication-competent noninduced pro-
2010; 9:1716–28. viruses in the latent reservoir increase barrier to HIV-1 cure. Cell 2013;
105. Thomann M, Reckermann K, Reusch D, Prasser J, Tejada ML. Fc-galactosylation 155:540–51.
modulates antibody-dependent cellular cytotoxicity of therapeutic antibodies. 114. Hake A, Pfeifer N. Prediction of HIV-1 sensitivity to broadly neutralizing antibodies
Mol Immunol 2016; 73:69–75. shows a trend towards resistance over time. PLoS Comput Biol 2017; 13:1–23.
106. Chung AW, Crispin M, Pritchard L, et al. Identification of antibody glycosylation 115. Magaret CA, Benkeser DC, Williamson BD, et al. Prediction of VRC01 neutral-
Downloaded from https://academic.oup.com/jid/article/223/Supplement_1/S22/6135673 by guest on 10 May 2021
structures that predict monoclonal antibody Fc-effector function. AIDS 2014; ization sensitivity by HIV-1 gp160 sequence features. PLoS Comput Biol 2019;
28:2523–30. 15:e1006952.
107. Mastrangeli R, Palinsky W, Bierau H. Glycoengineered antibodies: towards the 116. Yu W-H, Su D, Torabi J, et al. Predicting the broadly neutralizing antibody suscep-
next-generation of immunotherapeutics. Glycobiol 2018; 29:199–210. tibility of the HIV reservoir. JCI Insight 2019; 4: e130153.
108. Gagez AL, Cartron G. Obinutuzumab: a new class of anti-CD20 monoclonal an- 117. Rawi R, Mall R, Shen CH, et al. Accurate prediction for antibody resistance of
tibody. Curr Opin Oncol 2014; 26:484–91. clinical HIV-1 isolates. Sci Rep 2019; 9:1–12.
Antibodies for HIV-1 Cure Strategies • jid 2021:223 (Suppl 1) • S31You can also read

















































