Analysis of ovarian transcriptomes reveals thousands of novel genes in the insect vector Rhodnius prolixus - Nature
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
www.nature.com/scientificreports
OPEN Analysis of ovarian transcriptomes
reveals thousands of novel genes
in the insect vector Rhodnius
prolixus
Vitor Lima Coelho1, Tarcísio Fontenele de Brito1, Ingrid Alexandre de Abreu Brito1,
Maira Arruda Cardoso1, Mateus Antonio Berni1, Helena Maria Marcolla Araujo1,3,
Michael Sammeth2,4 & Attilio Pane1*
Rhodnius prolixus is a Triatominae insect species and a primary vector of Chagas disease. The genome
of R. prolixus has been recently sequenced and partially assembled, but few transcriptome analyses
have been performed to date. In this study, we describe the stage-specific transcriptomes obtained
from previtellogenic stages of oogenesis and from mature eggs. By analyzing ~ 228 million paired-
end RNA-Seq reads, we significantly improved the current genome annotations for 9206 genes.
We provide extended 5′ and 3′ UTRs, complete Open Reading Frames, and alternative transcript
variants. Strikingly, using a combination of genome-guided and de novo transcriptome assembly we
found more than two thousand novel genes, thus increasing the number of genes in R. prolixus from
15,738 to 17,864. We used the improved transcriptome to investigate stage-specific gene expression
profiles during R. prolixus oogenesis. Our data reveal that 11,127 genes are expressed in the early
previtellogenic stage of oogenesis and their transcripts are deposited in the developing egg including
key factors regulating germline development, genome integrity, and the maternal-zygotic transition.
In addition, GO term analyses show that transcripts encoding components of the steroid hormone
receptor pathway, cytoskeleton, and intracellular signaling are abundant in the mature eggs,
where they likely control early embryonic development upon fertilization. Our results significantly
improve the R. prolixus genome and transcriptome and provide novel insight into oogenesis and early
embryogenesis in this medically relevant insect.
Triatomine bugs (Hemiptera, Reduviidae, Triatominae) include hematophagous species that are widespread in
Latin America and have been firmly connected to the diffusion of Chagas disease. This life-threatening illness is
caused by Trypanosoma cruzi, a protozoan parasite that is transferred from the insect vector to a human host due
to the blood-feeding habit of the Triatomine species. Among these, R. prolixus represents a prominent arthropod
vector of the trypanosome. Between 6 and 7 million people are affected by Chagas disease in Latin American
countries1. However, several studies also reported the spread of Chagas disease in the United States of America,
Canada, Europe, Australia, and J apan2–5. The sequencing and partial assembly of the R. prolixus genome were
recently achieved and the latest RproC3 version is available in V ectorBase6,7. With 702 Mb of genomic DNA
sequenced, it was estimated that 95% of the genome, including 15,738 genes, has been covered6. Despite its medi-
cal relevance, few genomic resources are available for R. prolixus. Over the past decade, transcriptomic analy-
ses were carried out either using expressed sequence tags or, more recently, with high-throughput sequencing
techniques8–10. However, bioinformatic approaches coupled with molecular biology assays have recently shown
that many genes are missing from the current version of the genome11–13. These studies highlight substantial
limitations in the current R. prolixus genome annotations, which can hinder genetic, molecular and evolutionary
studies, and underscore the importance of generating more detailed maps of the genome.
R. prolixus is a hemimetabolous insect that develops through five nymph stages before reaching the sexually
mature adult stage14,15. Oogenesis in adult females is promoted by blood meals and each female can produce up
1
Institute of Biomedical Sciences (ICB), Federal University of Rio de Janeiro, Rio de Janeiro, Brazil. 2Institute
of Biophysics Carlos Chagas Filho (IBCCF), Federal University of Rio de Janeiro, Rio de Janeiro, Brazil. 3Instituto
Nacional de Ciência e Tecnologia em Entomologia Molecular (INCT-EM), Rio de Janeiro, Brazil. 4Department of
Applied Sciences, Institute of Bioanalysis, Coburg University, Coburg, Germany. *email: apane@icb.ufrj.br
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 1
Vol.:(0123456789)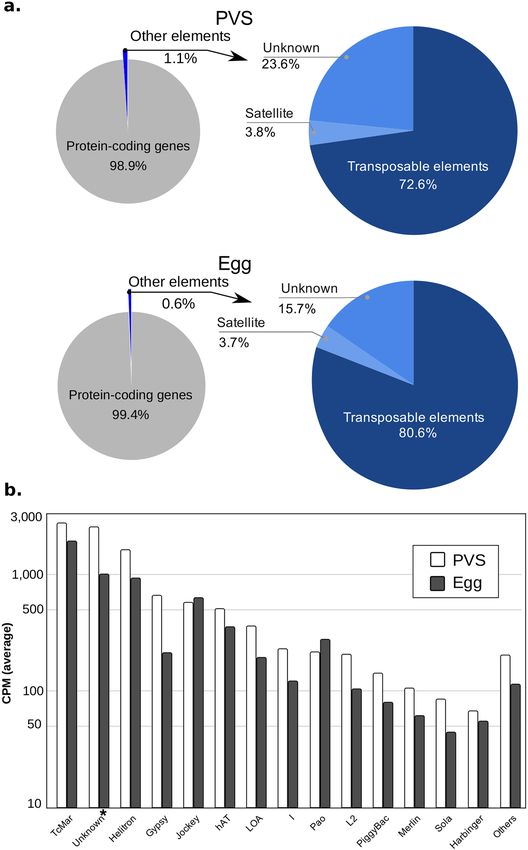
www.nature.com/scientificreports/
to 80 eggs16. Females harbor two ovaries, each containing 6–8 ovarioles simultaneously17,18. The anterior region
of the ovariole is formed by a lanceolate structure known as the tropharium that harbors the trophocytes and
pro-oocytes19. Trophocytes are polyploid cells and are arranged in a syncytium around a central cavity known
as the trophic core, while oocytes arrested in meiosis I are observed at the posterior of the tropharium. Fol-
lowing a blood meal and the reactivation of oogenesis, the oocytes are progressively surrounded by mitotically
active follicle cells to form mature egg c hambers14,15,17,20,21. After budding off from the tropharium, egg chambers
remain connected to the tropharium through cytoplasmic bridges called trophic cords18. Oogenesis can be
divided into three phases: pre-vitellogenesis, vitellogenesis, and choriogenesis. During previtellogenesis, RNAs,
proteins, and nutrients generated by the trophocytes are collected in the trophic core and then transported via
the trophic cords to the growing egg chambers18,19,22–24. While the growth of the oocyte is slow in the previtel-
logenic phase, it is rapidly promoted during vitellogenesis by fat body-derived vitellogenins and nutrients that
are directly absorbed by the o ocyte25. Once the oocyte reaches 1 mm in length, the trophic cords are severed and
choriogenesis begins, thus leading to the deposition of a hard chorion surrounding the oocyte. The egg displays
an apparent anterior–posterior polarity with the operculum at the anterior, which is eventually displaced by the
nymphs at eclosion. The dorsal–ventral axis is also readily distinguishable since the ventral region of the egg is
more concave. The chorion surrounds the mature egg in R. prolixus and protects it against dehydration, while
allowing the exchange of gases and liquids as well as f ertilization26–31.
The genetic and molecular mechanisms that control oogenesis in the model organism Drosophila melanogaster
have been elucidated to a great extent shedding light on key factors in important developmental processes, includ-
ing germline stem cell maintenance and cystoblast differentiation, DNA damage checkpoint and DNA repair
enzymes, mitosis and meiosis, and dorsal–ventral and anterior–posterior axial polarization. Furthermore, in
virtually all animal species, the initial stages of embryogenesis are genetically controlled by maternally-provided
molecules while the zygote is still transcriptionally inactive. Some of the mRNAs and proteins deposited by the
mother in the egg can control crucial developmental decisions. In D. melanogaster, egg chambers of the meroistic
polytrophic ovary are formed by a layer of epithelial cells surrounding the germline, which in turn, comprises 15
nurse cells and the oocyte. The nurse cells provide the gurken, bicoid, and nanos mRNAs, which are transported
and localized into the oocyte, where they control the polarization of the dorsal–ventral and anterior–posterior
axes of the egg chamber and future e mbryo32. TGF-alpha-like molecules, similar to Gurken in D. melanogaster,
also control the establishment of the dorsal–ventral axis in wasps, beetles, and crickets33. In D. melanogaster,
maternally-deposited oskar mRNA accumulates at the posterior of the oocyte, where it determines the nucleation
of the germplasm, a specialized cytoplasm that in turn contributes to the formation of the gonads in offspring34.
In addition, maternal RNAs and proteins mediate the maternal-to-zygotic transition (MZT) during embryo-
genesis, when the developmental program switches from maternal to zygotic c ontrol35. This process entails the
degradation of maternal RNAs and, to a certain extent, is controlled by active mechanisms. For instance, the
Smaug and Brain Tumor enzymes together with a cluster of miRNAs are responsible for maternal mRNA decay
in D. melanogaster, while miR-430 is a key factor in maternal RNA clearance in z ebrafish36,37. Except for a few
model organisms like D. melanogaster and zebrafish, the genetic basis of oogenesis and early embryogenesis is
still poorly understood in other animal species including R. prolixus.
We have recently provided an initial analysis of the transcriptome in previtellogenic stages (PVS) of R. pro-
lixus oogenesis38. In the current study, we combined the PVS datasets with newly generated transcriptomes from
mature eggs resulting in more than 228 million paired-end reads across two biological replicates for each stage.
Our analyses allowed us to dramatically improve the maps of genomic features in the R. prolixus genome and
identify thousands of protein-coding transcripts and repetitive element variants. We used these data to charac-
terize the genetic basis of R. prolixus oogenesis and the complement of maternally provided RNAs that drives
early embryogenesis in R. prolixus. Finally, we have collected our results together with transcriptomic datasets
from other laboratories in a mirror of the UCSC Genome Browser (Kent et al. 2002), that we named Rhodnius
Integrated Omics Browser or RIO browser at the URL www.genome.rio/cgi-bin/hgGateway?db=rproC3.
Results
Improvement of the reference genome annotation with ovarian RNA‑Seq datasets. We con-
ducted a deep interrogation of the previtellogenic stage (PVS) and mature unfertilized eggs (Egg) transcrip-
tomes, yielding in total ~ 228 M (128 M and 100 M, respectively) sequence reads across two biological replicates
for each stage. We previously published a partial analysis of the PVS libraries38, while the Egg transcriptomes
were produced de novo for the current study. Genomic mapping of the RNA-Seq libraries was performed against
the genome assembly RproC3 of R. prolixus6,7. Overall library mapping rates of the PVS and Egg conditions
were on average 96% and 77.5%, respectively. The mapping rate of read-pairs aligning to multiple locations in
the genome (i.e. multi-mapped read-pairs) was 12.6% on average (Supplementary Table S1). The proportion of
the read-pairs unambiguously or uniquely mapped to the genome was similar in PVS and Egg samples, with
85–87% of the total mapped read-pairs. Based on these observations, the uniquely mapped read-pairs were used
for a genome-guided transcriptome assembly to identify novel gene models in the genome and to improve the
current gene annotations of R. prolixus. Preliminary quantification of the genes expressed in PVS and Egg sam-
ples was performed by assigning the uniquely mapped read-pairs to the reference gene annotation available at
VectorBase7. On average, approximately 64% and 42% of uniquely mapped read-pairs of the PVS and Egg sam-
ples were respectively assigned to the reference gene annotation (column “Assigned paired-reads to reference
annotation”, Supplementary Table S1). Yet, a surprisingly high percentage of uniquely mapped read-pairs, on
average 36% for PVS and 58% for Egg, were not attributed to any reference genes or genomic features (Supple-
mentary Table S1). These observations pointed to the fact that genes important for R. prolixus oogenesis might
be missing in the current reference gene annotation. Thus, in order to expand the transcriptome landscape to
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 2
Vol:.(1234567890)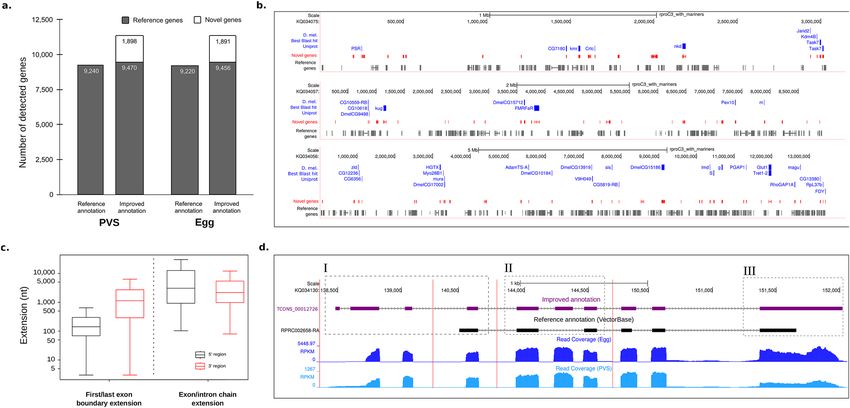
www.nature.com/scientificreports/
Figure 1. Improvement of genome annotations in R. prolixus. (a) Number of genes with at least 40 read-pairs
overlapping the reference and improved transcriptome annotation. (b) Representative R. prolixus super-
contigs with previously annotated genes (grey), novel genes (red), and their D. melanogaster orthologs (blue).
(c) Distribution of the extension lengths at the 5′ (black) and 3′ (red) ends. (d) (I) Extension of the genomic
coordinates of the gene RPRC002658 at the 5′ end due to the prediction of the alternative splice variant
TCONS_00012726 (purple); (II) Prediction of a skipped internal exon compared to the reference transcript
RPRC002658-RA (black); and (III) extension of the genomic coordinates at the 3′ end of the reference exon.
Light and dark blue plots represent the read coverage of the samples PVS and egg, respectively.
include unassigned/unmapped reads, we used the uniquely mapped and unmapped reads as input for genome-
guided transcriptome assembly and de novo transcriptome assembly approaches, r espectively39–41. The expan-
sion of the annotated genetic elements by the genome-guided approach provided 2126 novel putative genes. On
average, the number of genes overlapped by more than 40 read-pairs increased by 23% in PVS and Egg (Fig. 1a).
It is worth noting that we adopted a conservative approach; a cutoff of 10 overlapping read-pairs is generally
used to identify a new gene. For 1659 novel genes (78%), we identified 1204 complete ORFs, 365 (5′ or 3′) partial
ORFs, and 90 internal ORFs (Supplementary Table S3). Although the remaining 476 new transcripts display
small ORFs, we cannot rule out that they represent functional long non-coding RNAs (Supplementary Table S3).
Interestingly, the newly annotated genes often encode putative orthologs of D. melanogaster proteins that are
firmly linked to important developmental processes. Some notable examples in the displayed super-contigs are
the CREB-regulated transcription coactivator (Crtc), the transcription factor zelda (Zld), the type II transmem-
brane protein Star (S), the cadherin kugelei (Kug), the homeodomain transcription factor HTGX, and the BMP
regulator Magu. For easy visualization and navigation, we uploaded our datasets in a mirrored version of the
UCSC genome b rowser42, that we named Rhodnius Integrated Omics or RIO browser (Fig. 1b). To facilitate the
navigation, the RIO browser offers the possibility to use the name or acronym of the D. melanogaster ortholog
to identify and visualize the features, cross-reference databases, functional classification and gene ontology of a
R. prolixus gene.
In our previous study, we noticed that the annotation of certain genes (e.g. the R. prolixus vasa and PIWI
genes) was imprecise as their annotated 5′ and 3′ends did not match the gene boundaries revealed by our RNA-
Seq data38. We, therefore, sought to provide a more detailed map of the gene structure in R. prolixus by genome-
guided assembly of our PVS and Egg datasets. We were able to extend 1960 genes at their 5′ ends, 1473 genes at
their 3′ ends, and 5649 genes at both the 5′ and 3′ ends (Fig. 1c). In addition, we found approximately two new
transcript variants each for 6845 genes of the reference annotation. An assembled transcript was considered
a novel isoform when it displayed (i) a multi-exon structure with at least one junction match with a reference
gene in the same strand, (ii) minimum abundance of 40% of the most abundant isoform (i.e. the major isoform)
of the gene and (iii) Counts per Million (CPM) > 1 at least in one condition. Figure 1c shows two types of gene
extensions, one occurring at the 5′/3′ genomic coordinates of the first/last exon of an annotated gene and the
other that occurs due to the prediction of new chains of exons and introns for the gene (i.e. new isoforms). The
5′ and 3′ ends were extended by a median of 569 bp and 1800 bp, respectively (Supplementary Table S2). By
extending the annotated coding sequence (CDS), we found 78 novel complete Open Reading Frames (ORFs)
in the reference transcriptome. This also improved 1266 5′-incomplete CDSs by adding a start codon and 429
CDSs with a valid stop codon (Supplementary Table S3). A clear example of our transcriptome improvements is
represented by the RPRC002658 gene encoding an uncharacterized R. prolixus protein (Fig. 1d). Our analyses
allowed us to extend the 5′-end of this gene by approximately 2 Kb (Fig. 1d, box I) due to the prediction of the
novel TCONS_00012726 transcript isoform. This alternative splice variant also revealed an extra internal exon
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 3
Vol.:(0123456789)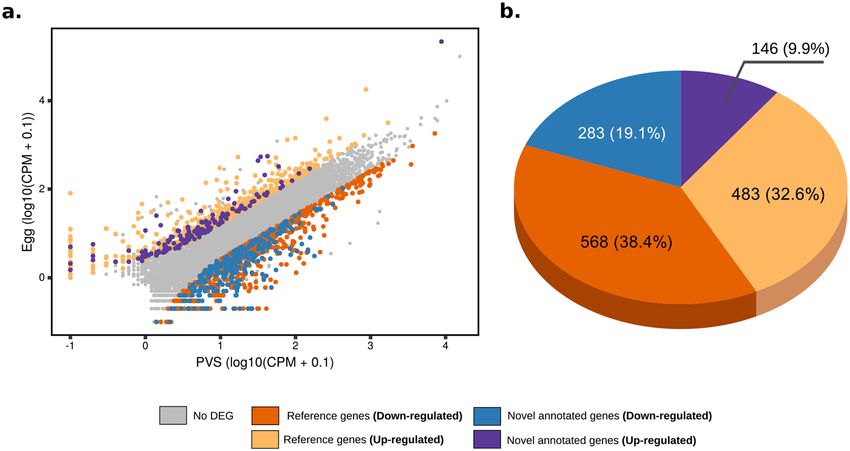
www.nature.com/scientificreports/
Genome-guided assembly De novo assembly Total assigned read pairs
Uniquely mapped read Assigned to improved Assigned to de novo To the reference To the improved
Samples pairs gene annotation Unmapped read pairs transcriptome transcriptome transcriptomea
PVS_1 31,651,452 26,759,942 (83.6%) 1,565,561 871,556 (55.7%) 20,314,277 (63.4%) 27,631,498 (83.2%)
PVS_2 21,166,732 17,784,143 (83.4%) 1,028,565 532,938 (51.8%) 13,931,026 (65.3%) 18,317,081 (82.5%)
Egg_1 17,400,997 15,369,913 (84.8%) 6,360,014 5,337,815 (83.9%) 8,049,949 (44.4%) 20,707,728 (87.2%)
5,918,720
Egg_2 14,258,924 12,500,145 (84.8%) 4,722,559 4,009,233 (84.9%) 16,509,378 (87%)
(40%)
Table 1. Statistics of the read assignment to the reference and expanded transcriptomes. a “Expanded
transcriptome” refers to the improved annotation and final putative de novo transcripts.
that was previously not described in the reference transcript RPRC002658-RA (Fig. 1d, box II). Finally, we were
able to extend the last exon of the gene by approximately 362 bp (Fig. 1d, box III). Other clear examples of the
improvements that we introduced in the R. prolixus are represented by the genes RPRC001867 (CRAL_TRIO_N
domain-containing protein, ortholog gene of D. melanogaster gene cralbp), RPRC004178 (Uncharacterized pro-
tein) and RPRC014580 (ADF-H domain-containing protein, ortholog gene of D. melanogaster gene GMF).
Despite the substantial expansion of the RproC3 transcriptome by genome-guided assembly, increasing the
percentage of the read-pairs overlapping genomic features by up to ~ 40% (Table 1), 28.7% of the reads in the
Egg and 4% in the PVS datasets did not map to any genomic regions (Supplementary Table S1). Since the egg
is not transcriptionally active, we reasoned that the stark difference between the percentage of unmapped reads
in the two datasets might be due to a small number of genes whose transcripts are maternally deposited in the
eggs, where they accumulate at higher levels compared to the PVS stage. To probe this hypothesis, we applied a
de novo assembly approach to the unmapped reads, which initially yielded 38,856 contigs for PVS and 19,390
contigs for Egg samples. After low-expression filtering, sequence deduplication, and quality contig evaluation
steps (see “Methods”), we obtained 1,993 non-redundant protein-coding transcripts (Supplementary Table S5).
However, upon closer inspection, we noticed that three contigs were accountable for > 80% of the unmapped
reads (Supplementary Table S4 and S5; Supplementary Fig. S1). One of these contigs encodes the R. prolixus
Rp30 eggshell p rotein43. This abundant protein has been already biochemically characterized and a partial cDNA
for this protein had already been observed in transcriptomic libraries derived from the R. prolixus digestive tract
and ESTs d atasets8,9,43. Our approach provides the missing sequences of the transcript and the full-length ORF.
The remaining two contigs encode predicted proteins that do not share similarity with known proteins in the
NCBI databases, but are particularly abundant in mature eggs. Additional studies will be required to understand
whether these genes are involved in the maturation of the egg chamber, like Rp30, or they are implicated in early
R. prolixus embryogenesis.
Stage‑specific gene expression during R. prolixus oogenesis. With our expanded version of the R.
prolixus transcriptome, we wondered what genes are expressed during oogenesis in this species. Our quantifi-
cation of the Egg datasets shows that the transcripts of 11,127 genes are present in the mature unfertilized egg
(Supplementary Table S6). In R. prolixus, RNAs and proteins synthesized in the tropharium are transported into
the oocyte via the trophic cord, while the oocyte is transcriptionally quiescent. We then performed differential
expression analysis by comparing the PVS and Egg datasets in an attempt to identify stage- and tissue-specific
gene expression patterns. Differentially expressed genes (DEGs) were selected according to DESeq244 to identify
reliable fold changes between the PVS and Egg datasets, where |fold-change|≥ 2.5 and adjusted p-value ≤ 0.0144
were set as the thresholds for significantly differentially expressed genes (Fig. 2a). Here, we refer to genes as
“down-regulated” or “up-regulated” if their steady-state RNA levels are respectively lower or higher in eggs
compared to PVS stages (Supplementary Table S6). Differential gene expression analysis resulted in 1480 DEGs
(Fig. 2a). Of these, 851 (57.5%) genes appeared to be expressed at high levels or specifically in the PVS stage,
while no expression beyond background levels was detected in Egg datasets. Surprisingly, however, 629 DEGs,
which is 42.5% of the total, appeared to be highly enriched in the Egg samples. Novel putative genes represent
29% of the total DEGs, with 9.9% up-regulated and 19.1% down-regulated in PVS versus Egg stages, respectively
(Fig. 2b).
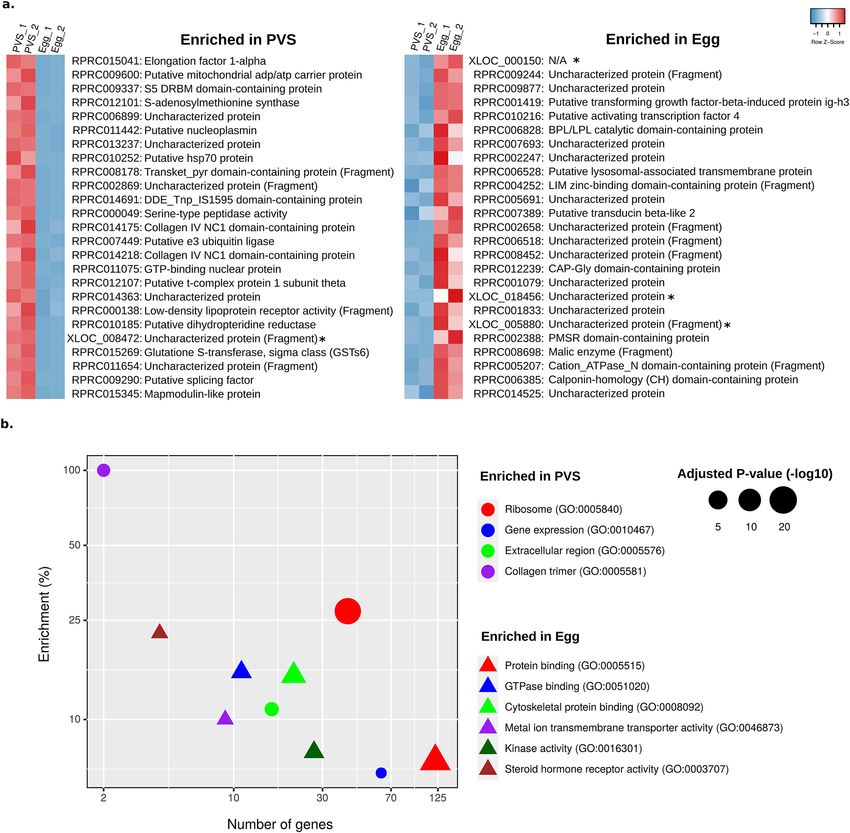
Consistent with our previous s tudy38, among the top 25 genes that are highly represented in the PVS tran-
scriptome, 15 encode ribosomal proteins (Supplementary Fig. S2). After removing these genes from the list,
the most differentially expressed genes in this stage code for components of the transcription and translation
machinery (Fig. 3a). Conversely, we find 13 genes in the top 25 expressed in eggs encoding uncharacterized
proteins along with evolutionarily conserved proteins (Fig. 3a). On top of this list, we find the XLOC_000150
transcript that might represent a long non-coding RNA. Some examples are given by putative orthologs of the
transforming growth factor-beta (TGF-β)-induced protein ig-h3 (RPRC001419), the activating transcription fac-
tor 4 (RPRC010216), a lysosomal-associated transmembrane protein (RPRC006528), and a Peptide Methionine
Sulfoxide Reductase (PMSR) domain-containing protein (RPRC002388), among others. Enrichment analysis of
the differentially expressed genes with g:Profiler45 found 35 and 54 GO terms enriched (Benjamini–Hochberg
FDR ≤ 0.05) in the PVS and Egg samples, respectively (Supplementary Table S7). Figure 3b shows ten major
functionally grouped networks of these enriched GO terms produced by ClueGO46,47. This approach yielded four
groups with 29 enriched terms in PVS (Supplementary Fig. S3a) and 12 groups with 54 enriched terms in Egg
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 4
Vol:.(1234567890)www.nature.com/scientificreports/
Figure 2. Differential expression analysis of previtellogenic stage and mature egg datasets of R. prolixus. (a)
Scatter plot of the gene expression normalized by the average CPM (counts per million, in log10). Y axis shows
Egg datasets, while X displays the PVS datasets. Upregulated reference (pink) and novel (magenta) genes and
downregulated reference (orange) and novel (blue) genes are displayed. Genes with no differential expression
in the two stages are shown (grey). (b) Composition of the total differentially expressed (DE) genes using the
improved gene annotation.
samples (Supplementary Fig. S3b). Our improved transcriptome confirms previous observations that the most
expressed reference genes in R. prolixus PVS encode putative proteins with homology to D. melanogaster proteins
related to ribosome, translation, and s plicing38, but it also adds enriched processes related to the P-bodies, the
extracellular region, and the collagen trimer (Supplementary Table S7; Supplementary Fig. S3a). Processing bod-
ies or P-bodies are eukaryotic cytoplasmic structures that are evolutionarily conserved in organisms as distant
as flies, worms and m ammals48. In D. melanogaster, P-bodies are organized around the nurse cell nuclei within
the egg chambers, where they form a membrane-less organelle known as “nuage”. The nuage and the P-bodies
share a number of enzymatic activities and have been linked to a different molecular mechanisms including RNA
decay and RNA interference. Among the genes enriched in the mature unfertilized eggs, we found protein-coding
genes associated with steroid hormone receptors, cellular organization, kinase activity, transmembrane trans-
port, protein binding, and GTPase binding. These results suggest that the females provides their oocytes with a
transcriptomic repertoire encoding hormone receptors (e.g. Nuclear hormone (NR) LBD domain-containing
protein and ecdysone response nuclear receptor e75a), enzymes (e.g. Diacylglycerol kinases and Guanylate
cyclase) and proteins with GTPase binding domains (e.g. Ran\Rab binding function and Rho GTPase exchange
factor activity) critical for organismal growth, microtubule organization, cell polarity, developmental transition
and regulation during the period of nutrient starvation, and transcriptional quiescence of early embryogenesis.
Key ovarian genes of D. melanogaster are conserved in R. prolixus and expressed during
oogenesis. We sought to determine the evolutionary conservation and stage-specific expression levels of
the R. prolixus orthologs of D. melanogaster genes that exert critical roles in oogenesis and early embryogen-
esis (Fig. 4). Therefore, we focussed on six cellular and developmental processes: germline stem cells, DNA
damage checkpoint and repair, mitosis/meiosis, germ granules/pole cells, axial polarization, and RNA decay
37,49–51
. For each process, we selected well-characterized D. melanogaster genes, searched for the putative R. pro-
lixus orthologs using BLAST tools, and measured their expression levels in PVS and Egg conditions using our
RNA-Seq datasets (Fig. 4 and Supplementary Table S8). Interestingly, we find that factors involved in D. mela-
nogaster germline stem cell maintenance and cystoblast differentiation are conserved in R. prolixus even though
this cell type is not observed in the adult ovary. The expression of the R. prolixus orthologs of pumilio (pum)
(RPRC000720), decapentaplegic (dpp) (RPRC000401), mad (RPRC008357), benign gonial cell neoplasm (bgcn)
(RPRC008118), medea (RPRC003748), glass bottom boat (gbb) (RPRC013357), zero-population-growth (zpg)
(RPRC014101), hedgehog (hh) (RPRC012384) and Notch (RPRC008058) is readily detected in both PVS and
Egg datasets (Fig. 4a)51–55. The differentiation of the D. melanogaster germline stem cells requires the activity of
bag-of-marbles (bam), a partner of BGCN in cystoblast differentiation. However, we did not find a bam ortholog
in the R. prolixus genome, or in the improved transcriptome. Interestingly, dpp transcripts are detected only in
PVS and do not accumulate in the egg.
We also examined the expression profile of genes involved in meiosis and mitosis, including cdk1
(RPRC009342), cyclin B (cycB) (RPRC012735), twine (RPRC008472), greatwall (RPRC005266), polo
(RPRC002498), orb (RPRC006637), orb2 (RPRC001021), calcineurin B (RPRC008393), sarah (RPRC006424),
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 5
Vol.:(0123456789)www.nature.com/scientificreports/
Figure 3. Functional categories enriched in previtellogenic stage and mature eggs. (a) Left: Top 25 differentially
expressed genes (non-ribosomal-related protein) ordered by average normalized expression (CPM) for PVS,
and the encoded putative protein according to the Uniprot database. Right: Top 25 most differentially expressed
genes at the Egg stage. Novel genes (identifiers with suffix “XLOC” and asterisks, *) were assigned to protein
products by BLAST sequence similarity. “N/A” indicates that a significant hit was not obtained (e-value ≤ 0.05).
(b) Enriched GO terms representing 10 functionally related groups identified by ClueGO in the PVS (circle) and
Egg (triangle) conditions. Enrichment (y-axis) is given by the number of genes in the input related to a GO term
(x-axis) divided by the total number of genes associated with this GO term in %.
verthandi (RPRC005603), stromalin (RPRC005579), SMC1 (RPRC009822) and SMC3 (RPRC009208)56–60 . All
of the genes appear to be expressed in PVS and deposited in the mature eggs with one notable exception of cycB.
We find two putative orthologs of cycB in the R. prolixus genome, RPRC012735 and RPRC001188, whose puta-
tive protein products share 43% and 37% amino acid sequence identity, respectively, with the D. melanogaster
cycB gene product (Supplementary Table S8). Yet, neither is expressed at detectable levels in R. prolixus ovaries,
nor do we find additional putative orthologs in our improved transcriptome. Given that cycB expression is
downregulated in endocycling cells like D. melanogaster nurse cells, and because most of the PVS tissues in
R. prolixus are occupied by giant polyploid nuclei, it is possible that the expression of cycB genes in R. prolixus
ovaries is below detectable levels.
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 6
Vol:.(1234567890)www.nature.com/scientificreports/
Figure 4. Gene expression levels of the R. prolixus orthologs of D. melanogaster genes involved in oogenesis and
early embryogenesis. (a) Genes involved in maintaining germline stem cells. (b) Genes related to DNA damage
checkpoint and repair processes. (c) Genes associated with mitosis and meiosis processes. (d) Essential genes
for the germ granules and pole cells (e) Genes related to the axial polarization process. (f) Genes associated with
RNA decay process. Y-axis displays expressions levels are normalized by transcripts per million (TPM).
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 7
Vol.:(0123456789)www.nature.com/scientificreports/
A set of evolutionarily conserved genes is dedicated to the DNA damage checkpoint and repair in organ-
isms ranging from yeast to mammals. DNA damage can be introduced by UV light and mutagenic agents, but
DNA double-strand breaks (DSBs) introduced by mei-W68, the D. melanogaster ortholog of yeast spo11, are
also required for meiotic r ecombination50,61–68. DNA damage and DSBs are sensed and repaired by a set of genes
including mei-41 (RPRC004298), chk-2 (RPRC005512), wee1 (RPRC013702), mei-w68 (RPRC004962), hus1-
like (RPRC004826), methuselah1 (mthl1) (RPRC001933), spn-A (RPRC006488), spn-B (RPRC010106), spn-D
(RPRC000377), Rad9 (RPRC001146), p53 (RPRC003641), ras85D (RPRC008553), apc (RPRC002106), and pten
(RPRC014866). The putative orthologs of these genes are present in the R. prolixus genome and the corresponding
transcripts can be readily detected both in early oogenesis as well as in mature eggs (Fig. 4b).
In D. melanogaster, the germ plasm is a specialized cytoplasm that accumulates at the posterior of the oocyte.
At the beginning of cellularization during embryogenesis, the germplasm is incorporated in the first cells appear-
ing at the posterior of the embryo. These cells, known as pole cells, will later migrate during gastrulation and join
gonadal somatic precursor cells to initiate the formation of the future gonad. Although there is no evidence of a
germplasm and germ granules in R. prolixus, we do observe the expression of the putative orthologs of the follow-
ing D. melanogaster genes that have been firmly connected to germplasm assembly and pole cell specification and
migration: exuperantia (exu) (RPRC001415), tudor (RPRC012896), valois (RPRC005681), vasa (RPRC009661),
germ cell-less (gcl), (RPRC010622), wunen (RPRC004485), wunen2 (RPRC009835), staufen (RPRC010019), mago
nashi (RPRC010985), pym (RPRC009783), eIF4AIII (RPRC002208), bruno (RPRC007498), rump (RPRC014343),
lost (RPRC014809), and tre1 (RPRC011268)69–75. All of the corresponding R. prolixus genes seem to be expressed
early in oogenesis and accumulate in the mature eggs. The steady-state RNA levels of the putative lost ortholog
in R. prolixus (Transcripts per Million or TPM > 1000) suggest that this gene exerts a prominent role during R.
prolixus oogenesis. As expected, we did not find a R. prolixus ortholog of the oskar gene, that is known to be
restricted to Diptera, while, consistent with our previous findings, the R. prolixus vasa gene is expressed in PVS
and the transcripts are deposited in mature chorionated e ggs38.
Axial polarization in D. melanogaster is dictated by maternal transcripts encoding the Bicoid and Gurken
proteins, whose localized translation sets the anterior–posterior and dorsal–ventral axes, respectively, of the egg
and future embryo76,77. Neither of these two genes appears to be conserved in R. prolixus, although we found a
spitz/tgf-a gene (RPRC009386) that is expressed in both the PVS and Egg datasets. Members of the TGF-alpha
family were shown to control the formation of the dorsal–ventral axis in crickets, wasps, and b eetles78. We also
find putative orthologs of nanos (RPRC002927) and torso-like (RPRC006513), but not of torso, which are involved
in the specification of the anterior–posterior axis in fruit flies, aphids and honeybees79.
Finally, we investigate the expression profiles of genes associated with the RNA decay pathway. The early
phases of embryonic development in D. melanogaster and other organisms are driven by maternally contributed
RNAs and proteins. However, the maternal program must be erased at the onset of MZT so that the embryo
can follow a zygotically-driven transcription program. The degradation of maternal RNAs in D. melanogaster
is mostly guided by the protein products of a group of genes comprising smaug (RPRC007649), brain tumor
(RPRC007254), not1 (RPRC005691), not3 (RPRC003870), not10 (RPRC001957), cnot4 (RPRC001525), pop2
(RPRC004491), regena (RPRC003111), caf1 (RPRC015316), early girl (RPRC004083), wispy (RPRC007755),
twin/CCR4 (RPRC006991), and string (RPRC008472)80–85. In addition, zelda (RPRC000020) encodes a transcrip-
tion factor that is pivotal for the onset of zygotic transcription 86. We find that all of these genes are expressed
in PVS and their transcripts are detected in mature eggs. Unexpectedly, the levels of smaug, wispy, and zelda are
low and might suggest that other mechanisms are in place during early R. prolixus embryogenesis to degrade
maternal RNAs and prompt zygotic transcription. We did not find R. prolixus orthologs for the following key D.
melanogaster genes in the genome or in our updated transcriptome: fs(1)Yb, swallow, trunk, barentsz, atr/tefu,
cup, and matrimony50,87–89. Despite the extensive conservation of crucial genes between D. melanogaster and
R. prolixus, we also detect important differences, which might explain, at least in part, the striking differences
between oogenesis and embryogenesis in these two insect species.
Expression levels of transposable and repetitive sequences during R. prolixus oogene-
sis. Transposable elements (TE) are mobile sequences that often constitute a significant proportion of animal
and plant genomes. While the biological function of these selfish sequences is still debated, it is well estab-
lished that transposon mobilization is often connected to DNA damage, insertional mutagenesis, and genome
instability90–92. RepeatMasker annotation of RproC3, available at VectorBase, shows that more than half of R.
prolixus mobilome is dominated by repetitive elements (longer than 0.5 Kb) of the type Tc1-mariner (28%), Heli-
tron (14%), Tc1 (10%) and a class of repetitive elements labeled as Unknown (9.8%) (Supplementary Table S9).
The prevalence of Tc1-mariner elements has been reinforced using de novo and library-based approaches for
transposon detection, revealing that approximately 75% of the transposons dispersed in the R. prolixus genome
belong to the Tc1-Mariner family93. Given the incomplete assembly of the genome and the challenges imposed
by the repetitive nature of these elements, it is likely that the mobilome of R. prolixus is not completely char-
acterized. Furthermore, the expression levels of these sequences have not been investigated yet in R. prolixus.
We therefore employed the available data together with our stage-specific RNA-Seq datasets to determine the
expression levels of all known transposon families in R. prolixus oogenesis. As for the protein-coding genes, we
also compared the early (PVS) and late (Egg) stages of oogenesis to identify transposons potentially displaying
differential activity in these two stages (Fig. 5). First, we calculated the RNA-Seq data coverage of the trans-
posable elements by assigning all of the mapped read-pairs to the RproC3 RepeatMasker annotation and our
improved gene annotation. Read-pairs assigned to more than one feature or genomic element class (e.g., protein-
coding gene, transposable element, simple repeats) were considered ambiguous and, therefore, not included in
the pie charts (less than 7% of the total assigned read-pairs) (Fig. 5a). In agreement with their relative abundance
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 8
Vol:.(1234567890)www.nature.com/scientificreports/
Figure 5. RNA levels of transposable and repetitive elements. (a) Left: pie chart shows the assignment of the
multi- and uniquely-mapped paired-end reads to the protein-coding gene annotation (gray) and repetitive
element regions (blue). Right: pie chart shows a more detailed distribution of mappings for satellite simple
repeats, transposable elements, and unknown classes of repetitive elements. (b) Cumulative quantification of
expression for each class of transposable elements longer than 0.5 Kb (kilobases) for the conditions PVS (white)
and egg (gray).
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 9
Vol.:(0123456789)www.nature.com/scientificreports/
Identifier Stage Dfam classification E-value Score Rep. 1 (CPM) Rep. 2 (CPM)
DN6993_c2_g2_i2 PVS R1 (DF0004710) 3.6e−08 32.2 119.3 1714.6
DN6822_c0_g1_i1 PVS Mariner (DF0006767) 4.3e−38 129.6 705.3 611.9
DN6491_c0_g1_i1 PVS Satellite (DF0001947) 4.3e−17 61.4 345.8 462.5
DN6980_c0_g1_i1 PVS R1 (DF0004887) 2.40e−09 35.6 114.9 544.3
DN4130_c0_g1_i1 PVS Mariner (DF0004831) 1.8e−14 52.5 202.9 257.9
DN6969_c2_g5_i1 PVS L2 (DF0006592) 9.9e−16 55.1 209.1 216.3
DN6874_c0_g1_i3 PVS R1 (DF0004775) 4.3e−33 113.5 57.1 307.8
DN6975_c2_g1_i1 PVS RTE-BovB (DF0004797) 6.2e−09 34.8 96.7 129.9
DN4053_c0_g1_i1 PVS Unknown (DF0005939) 4.4e−14 51 187.7 1.8
DN6127_c0_g1_i2 PVS DNA transposon TIR (DF0005060) 5.8e−10 38.2 173.2 0
DN5699_c0_g1_i1 PVS RTE-BovB (DF0005461) 1.5e−08 31.7 73.9 81.8
DN2864_c3_g2_i1 Egg Unknown (DF0004525) 9.6e−44 148.4 32,006 28,084
Table 2. Putative repetitive elements and transposons non-redundant sequence variants.
in the genome, the Tc1-mariner family of transposons appeared to have the highest expression levels both in
PVS and in mature eggs (Fig. 5b). High expression levels are also observed for a class of “unknown” transposons
and the Helitron family. When comparing the steady-state levels in PVS versus Egg datasets, we observed that
some classes of DNA transposons, LTR and Non-LTR retrotransposons, have greater coverage in the previtel-
logenic stage (like the “unknown” TE class and gypsy), while others have greater coverage in the mature unferti-
lized egg (like Tc1-mariner and helitron). Despite these differences, however, it seems that all of the transposon
transcripts that are generated in the tropharium are efficiently transported to the growing oocyte. We were also
interested in identifying novel repetitive element (RE) variants, more specifically transposons. To do so, we
performed homology searches based on hidden Markov model (HMM) profiles of transposable elements94,95
against our de novo assembled contigs. In total, homology searches resulted in 167 contigs with significant
hits of non-rRNA repetitive elements. To reduce the sequence redundancy between stages, we aligned the new
repetitive elements with each other, resulting in 20 non-redundant putative REs (Supplementary Table S10). We
also aimed to identify whether our transposons were already characterized in the library of RproC3 repetitive
elements available at VectorBase or found in published Tc1-mariner consensus sequences93. Interestingly, we
found 12 non-redundant transposon consensus sequences with no significant similarity or with low alignment
coverage with RproC3 repetitive elements6,7 or Tc1-mariner consensus sequences93 (Table 2). Abundances of
these sequences are represented by the number of estimated reads normalized by the total of estimated reads
(estimated CPM) for each replicate (“Rep.”) of the PVS and Egg de novo contigs. Our study clearly shows that a
variety of transposons belonging to known or uncharacterized families are expressed in early oogenesis and their
transcripts accumulate in the mature eggs.
Discussion
R. prolixus is a primary vector of Trypanosoma cruzi, the etiologic agent of Chagas disease. Sequencing of the R.
prolixus genome in recent years together with studies on specific pathways or gene sets provided an important
resource to understand the biology of this insect6,96 . In this study, we sought to improve genome annotations
and use the improved transcriptome to gain insight into R. prolixus oogenesis and the maternal contribution to
early embryonic development. Using a combination of genome-guided and de novo transcriptome assembly, we
were able to increase the number of putative protein-coding genes in R. prolixus from the 15,738 that are cur-
rently annotated in the genome (RproC3 version) to 17,864. Many of these genes likely code for critical factors
in R. prolixus development as they include histone modification enzymes and chromatin remodeling factors,
homeotic genes, signaling molecules and kinases, and metabolic enzymes, many of which are evolutionarily
conserved. Furthermore, we significantly improved the 5′ and 3′ boundaries of previously annotated genes, thus
shedding light on their 5′ and 3′ UTRs. Importantly, for 2,709 of these genes, we revealed the complete ORF and
the encoded full-length putative protein. Finally, the paired-end nature of our libraries allowed us to determine
transcript variants generated by alternative splicing events for 6845 genes.
Based on the updated transcriptome, we investigated the complement of genes expressed in early stages of
R. prolixus oogenesis that drive egg chamber formation and oocyte maturation, as well as the atlas of transcripts
that are maternally deposited in mature unfertilized eggs. The latter genes are essential during the first stages of
embryogenesis, when zygotic transcription is not yet active. The morphology and architecture of the ovarioles
differ profoundly in R. prolixus and D. melanogaster. While in D. melanogaster the oocytes are produced from ger-
mline stem cells located at the anterior tip of the ovariole, this cell type seems to be restricted to nymph stages in
R. prolixus97. Instead, the anterior region of the ovariole harbors actively dividing trophocytes in this s pecies17–19.
Despite this difference, we find orthologs of Drosophila genes involved in germline stem cell specification and
maintenance, like hts, α-spec, bgcn and dpp, in early stages of R. prolixus oogenesis. Particularly surprising is the
expression of hts and a-spec, given that the products of these genes in D. melanogaster mark the spectrosome/
fusome, a cytoplasmic structure that initially labels the germline stem cells, but once the cystoblast differentiates
and start diving, it forms a bridge connecting the germ cells within the mitotic c yst98. The spectrosome/fusome
exerts an important role in oocyte specification in the fruit fly. Thus, it will be of great interest to characterize
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 10
Vol:.(1234567890)www.nature.com/scientificreports/
its localization and function during R. prolixus oogenesis, where the pro-oocytes are already specified in earlier
nymph stages and germline stem cells are not detected in the adult ovary. Our analysis also shows that many D.
melanogaster genes controlling critical processes like meiosis/mitosis, DNA damage checkpoint and repair, germ
plasm assembly and pole cell formation, maintenance and migration and RNA decay are conserved in R. prolixus.
Remarkably, the R. prolixus orthologs of several genes, whose expression in D. melanogaster is restricted to early
oogenesis like hts, α-spec, bgcn, and mei-W68, are instead expressed in PVS and their transcripts deposited in
mature eggs. This might represent a consequence of the particular structure of R. prolixus egg chambers and
ovarioles. The egg chamber is exclusively formed by somatic follicle cells surrounding the oocyte and is con-
stantly communicating with the tropharium through the trophic cords until choriogenesis begins. Differently,
the egg chamber in the fruit fly is formed by a layer of follicle cells surrounding the germline, where the germline
comprises the oocyte and fifteen nurse cells. Once the egg chamber buds off the germarium in D. melanogaster,
it remains connected to the germarium through stalk cells, but the supply of RNAs and proteins for the growth
and patterning of the oocyte is provided by the adjacent nurse cells through ring canals. Therefore, RNAs and
factors expressed in early stages of oogenesis (i.e. tropharium/germarium) can be found in the mature eggs of
R. prolixus, but not in those of D. melanogaster. The presence of transcripts encoding components of the RNA
decay pathway, including smaug, wispy, and the CCR4/Not complex, suggest that maternal RNAs are actively
degraded in R. prolixus to promote the maternal-to-zygotic transition. As expected, some patterning genes
like gurken and bicoid, which are responsible for axial polarization in D. melanogaster are not detected in the
R. prolixus genome given that they are restricted to Diptera33. However, we identified a putative ortholog of D.
melanogaster spitz, which encodes a TGF-α protein, expressed in R. prolixus oogenesis. Members of this fam-
ily, which include Gurken, have been connected to the establishment of the dorsal–ventral polarity in crickets,
wasps, and beetles33. Also, we did not find an obvious ortholog of the germ plasm factor oskar, although a oskar
gene was identified in cricket, a hemimetabolous insect like R. prolixus99. Another patterning gene expressed in
R. prolixus oogenesis is nanos. Nanos is responsible for the specification of the posterior segments in a range of
distantly related insect s pecies78,100,101. Although these observations suggest that the mechanisms controlling axial
polarization and germplasm formation are partially conserved in R. prolixus, functional studies are required to
gain further insights into these processes.
Our data show that the transcripts encoding a variety of ribosomal proteins rank among the top 25 genes that
are expressed early during oogenesis, but are under-represented in mature unfertilized eggs. It is possible that in
R. prolixus ribosomal proteins rather than the corresponding transcripts are deposited in the egg and used in early
stages of embryogenesis. This strategy might guarantee that ribosomal proteins are readily available to rapidly
assemble ribosomes. Surprisingly, we found 629 genes to be highly represented in the Egg samples, but not in
the PVS. It seems reasonable to assume that these genes are activated in the tropharium in response to the blood
meal, but their expression is not sustained over time. The transport of the corresponding transcripts to develop-
ing oocytes would, therefore, progressively deplete the tropharium of these RNAs while they accumulate in the
egg. Among the top 20 of DEGs with high expression in mature eggs, we find a putative ortholog of Yellow-g2
(RPRC009244). This gene has been associated with the resistance of mosquito eggs to desiccation as well as to
the production of cuticle pigments in R. prolixus102. At least two genes might be activated in oogenesis to control
the oxidative stress caused by the blood feeding habits of R. prolixus. In fact, RPRC010216 encodes a putative
ortholog of the cyclic AMP-dependent transcription factor ATF-4, a key regulator in metabolic and redox pro-
cesses as well as a crucial factor in the integrated stress response, and RPRC002388 encoding a predicted PMSR-
containing protein. Peptide Methionine Sulfoxide Reductase (PMSR) was associated with the repair of proteins
and peptides that have been damaged by oxidizing environments103. It is well-established that the blood-feeding
habit of R. prolixus generates oxidative species. Hence, RPRC002388 might exert a crucial role in the repair of
proteins during R. prolixus oogenesis and early embryonic development. Another gene that might be linked to
the heme metabolism is RPRC001419 encoding a putative ortholog of the venom periostin-like protein 1 from
Pristhesancus plagipennis, also an assassin bug like R. prolixus, but preying on other insects and not on mammals.
Pristhesancus injects the venom into the prey to paralyse it and liquify the tissues to facilitate the feeding. Finally,
some promising genes need to be validated by other molecular biology techniques. For instance, RRPC009877
coding for a putative ortholog of a proline-proline-glutamic acid (PPE) protein from Mycobacterium. The PPE
protein family has been connected to cell wall remodeling and virulence in bacteria. It is noteworthy however,
that the neighboring gene in the contig, namely RPRC009884, also encodes for a putative bacterial protein. It will
be important to determine whether these genes are truly present in the R. prolixus genome or they are bacterial
genes accidentally assembled in the insect genome.
Our data also reveal that a variety of transposable elements are expressed in the early stages of oogenesis and
their transcripts accumulate in the developing oocyte. It was previously shown that approximately 6% of the R.
prolixus genome is made up of transposable sequences, with the Tc1-mariner elements comprising 75% of the
mobilome. In accordance, we find that the Tc1-mariner transposons display the highest steady-state RNA levels
in both PVS and Egg samples compared to other transposon families. However, the helitron elements also appear
to display comparable expression levels to Tc1-mariner together with a class of “unknown” repetitive sequences,
even though they are less represented in the genome. Overall, transposable and repetitive sequences account
for only 1.1% and 0.6% of the unambiguous reads in PVS and Egg stages, respectively. We previously showed
that key components of the piRNA pathway are conserved in R. prolixus and are important for female adult
fertility38. Together with the observation that R. prolixus ovaries express p iRNAs38, these data suggest that the
piRNA pathway efficiently downregulates transposable and repetitive sequences in R. prolixus oogenesis. For all
of the transposons, the RNA levels in PVS and Egg stages are generally comparable, thus pointing to a mechanism
whereby transposon transcripts are expressed in the trophocytes and transported from the tropharium onto the
oocyte through the trophic cords. Although our analysis does not allow us to draw conclusions on the transcrip-
tional activity of the transposons in the follicle cells versus the germline in R. prolixus, the fact that transcripts
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 11
Vol.:(0123456789)www.nature.com/scientificreports/
corresponding to all known transposon families are detected both in PVS and in Egg stages demonstrates that
none of these families is restricted to the follicle cells. It remains to be elucidated whether transposons can be
transcribed in the follicle cells and their transcripts transferred to the oocyte during oogenesis or rather the
transcription of these repetitive sequences occurs both in somatic and germ cells.
Chagas disease is a main threat to human health worldwide, but cures or vaccines for this illness are not yet
available. The most promising strategies to reduce its diffusion rely on targeting triatomine vectors. Our study
sheds light on oogenesis, early embryogenesis, and adult female fertility in R. prolixus, a primary vector of the
Chagas disease in Central and South America. Together with substantial improvements to genome annota-
tion in this species and novel bioinformatic resources, we provide a framework for future genetic, genomic
and functional studies that might pave the way to the development of novel R. prolixus population control or
replacement strategies.
Methods
R. prolixus handling, total RNA extraction, and library preparation. The R. prolixus colony was
raised at 28 °C and 75% relative humidity and regularly fed white rabbit blood at 3-week intervals in the labora-
tory of Insect Biochemistry at the Institute of Medical Biochemistry at the Federal University of Rio de Janeiro,
Brazil. All animal care and experimental protocols were conducted in accordance with the guidelines of the
Committee for Evaluation of Animal Use for Research (Universidade Federal do Rio de Janeiro, CAUAP-UFRJ)
and the NIH Guide for the Care and Use of Laboratory Animals (ISBN 0-309-05377-3). Protocols were approved
by CAUAP-UFRJ under registry #IBQM155/13. Dedicated technicians in the animal facility localized at the
Instituto de Bioquímica Médica Leopoldo de Meis (UFRJ) carried out all protocols related to rabbit husbandry
under strict guidelines, with supervision of veterinarians to ensure appropriate animal handling. Approxi-
mately, 100 mature chorionated eggs were dissected in ice-cold Phosphate Buffered Saline (PBS) from 10/15
adult females two weeks after a blood meal, and RNA-Seq libraries were produced as previously d escribed38.
Briefly, total RNA was extracted with TRIzol Reagent (Life Technologies) according to manufacturer instruc-
tions, treated with TURBO DNA-free kit (Ambion), and subjected to paired-end RNA sequencing (RNA-Seq)
library production (Illumina). The libraries were generated and sequenced on HiSeq Illumina platforms at Lac-
tad Facility (University of Campinas, Brazil) as previously described38. PVS libraries were generated previously
and are available from NCBI Sequence Read Archive (SRA)38.
Pre‑processing and genomic mapping of the RNA‑Seq reads. Different tools were used in order to
ensure the qualities of the reads we worked on as well as map them to Rhodnius’ genomic sequence. FastQC104
was adopted to check the quality of the libraries in regard to per-base sequence quality, sequence duplication
level, overrepresented sequences, GC content, and presence of Illumina adapters. Raw read sequences were pre-
processed by filtering out read sequences with less than 20 bp in length (-length 20); removing Ns from the read
ends (-trim-n); trimming Illumina adapters (-illumina) and low-quality bases (-q 20) by Trim Galore! version
0.6.1105,106. A genome index was built with the genome assembly of R. prolixus RproC37 and R. prolixus Tc1-
mariner consensus sequences93 for the first-pass genomic mapping. Read alignment was performed by STAR
(Spliced Transcripts Alignment to a Reference) version 2.7.2b107 reporting read-pairs with at most 30 multiple
alignments (-outFilterMultimapNmax) and a ratio between mismatches and mapped bases less or equal to 0.1
(-outFilterMismatchNoverLmax). To rescue unmapped reads in this first-pass genomic mapping, we trimmed
these sequences for poly-A/T tails (tail length > 4) and we filtered by a minimum length (> 25nt) and GC-content
(> 20%) using prinseq-lite v0.20.4108 and re-mapped again (Supplementary Table S1).
Transcriptome improvements in the reference annotation. Novel genes and genomic coordinate
updates of reference genes were identified using the genomic mappings by Cufflinks. Uniquely mapped read-
pairs were used to improve the transcriptome landscape by genome-guided transcriptome assembly using Cuf-
flinks pipeline version 2.2.141 with the following parameters: at least 20 mapped read-pairs to support an assem-
bled transcript (-min-frags-per-transfrag 20), novel isoform abundance ≥ 40% of the major isoform abundance
(-min-isoform-fraction 0.4), reference assembly RproC3 (Rhodnius-prolixus-CDC_SCAFFOLDS_RproC3.fa)
was used for bias detection (-b) and to exclude artifacts (e.g. repetitive elements) (-s); and the reference gene
annotation of RproC3 (Rhodnius-prolixus-CDC_BASEFEATURES_RproC3.3.gtf) was used to guide the assem-
bly, to compare with reference genes and to classify the assembled elements. We considered the assembled ele-
ments with a complete match of intron chain (class code “ = ”), potentially novel isoform (class code “j”), and
unknown/intergenic transcript (class code “u”). Novel isoform abundances were calculated by Salmon v1.2.1109
and assembled isoforms with estimated CPM < 1 (Counts per Million) on both previtellogenic and egg stage
were removed from the transcriptome assembly. Transdecoder v5.5110 was applied to predict ORF candidates in
the reference transcripts and to compare with the predicted ORFs in the improved transcriptome (Supplemen-
tary Tables S2 and S3).
Discovery of novel transcripts and repetitive elements by de novo assembly. RNA-Seq
sequences that failed to map to the reference genome were used by the tool Trinity to assemble transcripts
and repetitive elements. Unmapped read-pairs were used for de novo transcriptome assembly with the Trinity
method v2.5.140 with default parameters to identify novel transcripts that were not previously detected due to
incomplete sequencing and assembly of the R. prolixus genome for each development stage, PVS and egg. De
novo assembled transcripts were post-processed to filter out potential artifacts based on abundance and quality.
First, expression levels of the de novo assembled transcripts were quantified by Salmon v1.2.1109 and isoforms
with TPM < 10 were removed. Next, deduplication of redundant contig sequences was performed by CD-HIT-
Scientific Reports | (2021) 11:1918 | https://doi.org/10.1038/s41598-021-81387-1 12
Vol:.(1234567890)You can also read

















































