Conservation of the glycogen metabolism pathway underlines a pivotal function of storage polysaccharides in Chlamydiae - Nature
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
ARTICLE
https://doi.org/10.1038/s42003-021-01794-y OPEN
Conservation of the glycogen metabolism pathway
underlines a pivotal function of storage
polysaccharides in Chlamydiae
Matthieu Colpaert 1, Derifa Kadouche1,7, Mathieu Ducatez1,7, Trestan Pillonel 2, Carole Kebbi-Beghdadi2,
Ugo Cenci1, Binquan Huang1,6, Malika Chabi1, Emmanuel Maes3, Bernadette Coddeville1, Loïc Couderc3,
Hélène Touzet4, Fabrice Bray5, Catherine Tirtiaux1, Steven Ball1, Gilbert Greub 2 & Christophe Colleoni 1 ✉
The order Chlamydiales includes obligate intracellular pathogens capable of infecting mam-
1234567890():,;
mals, fishes and amoeba. Unlike other intracellular bacteria for which intracellular adaptation
led to the loss of glycogen metabolism pathway, all chlamydial families maintained the
nucleotide-sugar dependent glycogen metabolism pathway i.e. the GlgC-pathway with the
notable exception of both Criblamydiaceae and Waddliaceae families. Through detailed gen-
ome analysis and biochemical investigations, we have shown that genome rearrangement
events have resulted in a defective GlgC-pathway and more importantly we have evidenced a
distinct trehalose-dependent GlgE-pathway in both Criblamydiaceae and Waddliaceae families.
Altogether, this study strongly indicates that the glycogen metabolism is retained in all
Chlamydiales without exception, highlighting the pivotal function of storage polysaccharides,
which has been underestimated to date. We propose that glycogen degradation is a man-
datory process for fueling essential metabolic pathways that ensure the survival and virulence
of extracellular forms i.e. elementary bodies of Chlamydiales.
1 University of Lille, CNRS, UMR8576-UGSF-Unité de Glycobiologie Structurale et Fonctionnelle, Lille, France. 2 Institute of Microbiology, University of
Lausanne and University Hospital Center, Lausanne, Switzerland. 3 University of Lille, CNRS, Inserm, CHU Lille, Institut Pasteur de Lille, US 41 - UMS 2014 -
PLBS, Lille, France. 4 University of Lille, CNRS, Centrale Lille, UMR 9189 - CRIStAL - Centre de Recherche en Informatique Signal et Automatique de Lille,
Lille, France. 5 University of Lille, CNRS, USR 3290—MSAP—Miniaturisation pour la Synthèse, l’Analyse et la Protéomique, Lille, France. 6Present address:
State Key Laboratory for Conservation and Utilization of Bio-Resources in Yunnan/School of Agriculture, Yunnan University, Kunming, China. 7These authors
contributed equally: Derifa Kadouche, Mathieu Ducatez. ✉email: christophe.colleoni@univ-lille.fr
COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio 1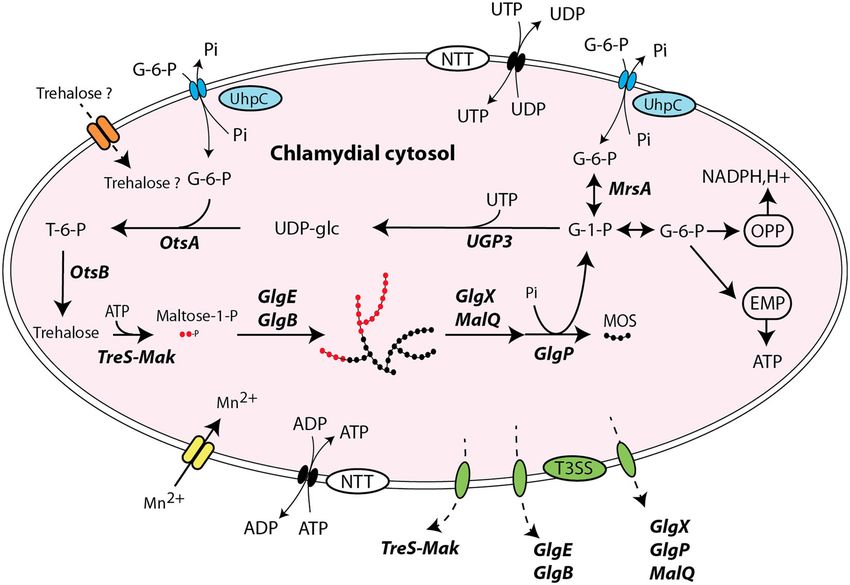
ARTICLE COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y
C
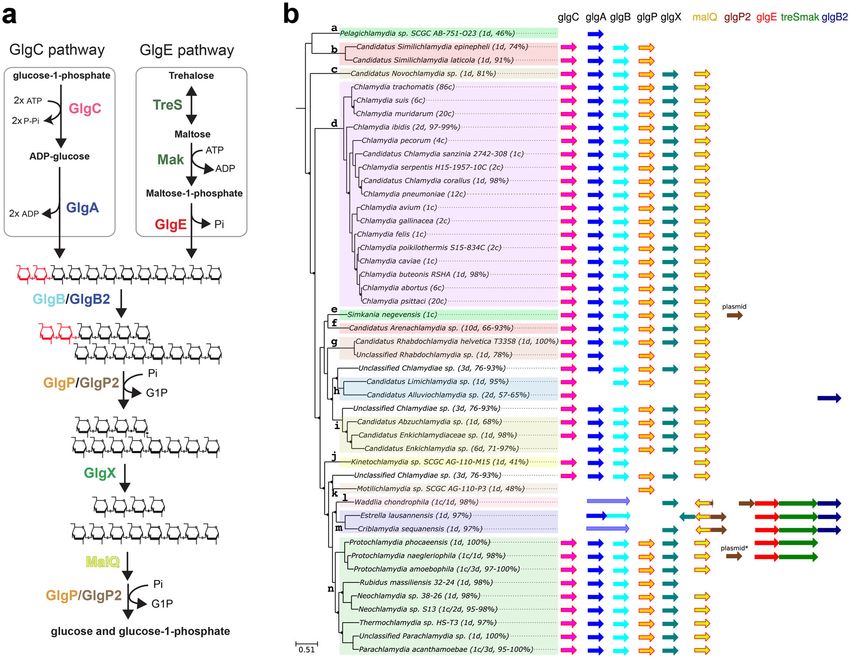
hlamydiae forms with Planctomycetes and Verrucomi- Mycobacteria, the latter is produced either from the condensation
crobia phyla a very ancient monophyletic group of bac- of glucose-1-P and ADP-glucose catalyzed by a glycosyl trans-
teria known as PVC, which has been recently enriched ferase called GlgM or from the interconversion of trehalose (α-α-
with additional phyla1. The Chlamydiales consists of members of 1,1-linked D-glucose) to maltose followed by the phosphorylation
the Chlamydiaceae family that includes etiological agents of of maltose, which is catalyzed by trehalose synthase (TreS) and
human and animal infectious diseases and at of least eight maltose kinase (Mak) activities, respectively17. At the exception of
additional families commonly named “chlamydia-related bac- Actinobacteria (i.e., mycobacteria and Streptomycetes), TreS is
teria” or “environmental” chlamydia2,3. usually fused to maltokinase (Mak) that phosphorylates maltose
All Chlamydiales display an obligate intracellular lifestyle due into maltose-1-phosphate19. Subsequently, maltosyl-1-phosphate
to a massive genome reduction and biphasic development, which transferase (GlgE) mediates the formation of α-1,4-linked poly-
includes two major morphological and physiological distinct mers by transferring the maltosyl moiety onto the non-reducing
stages: the elementary body (EB), a non-dividing and infectious end of a growing α-1,4-glucan chain. As in the GlgC pathway,
form adapted to extracellular survival and the reticulate body branching enzyme (GlgB) then introduces α-1,6 linkages to give
(RB), a replicating form located within a membrane surrounded rise to a highly branched α-glucan. The GlgC pathway is found in
inclusion (for review, see ref. 4). Following entry into a susceptible approximately one-third of the sequenced bacteria and is by far
cell, the EBs differentiate into RBs within the inclusion. During the most widespread and best studied; the GlgE pathway has been
the intracellular stage, RBs secrete many effector proteins through identified in 14% of the genomes of α-, β-, γ-proteobacteria while
the type III secretion system and express a wide range of trans- 4% of bacterial genomes possess both GlgC- and GlgE-
porters in order to manipulate host metabolism and uptake all the pathways19,20.
metabolites required for its replication. At the end of the infection In order to shed light on metabolism of storage polysaccharide
cycle, RBs differentiate back into EBs before they are released into in Chlamydiales, we analyzed 220 genomes (including some
the environment5,6. genomes assembled from metagenomic data) from 47 different
Glycogen metabolism loss appears to be a universal feature of chlamydial species that represent the bulk of currently known
the reductive genome evolution experienced by most if not all chlamydial diversity. A complete GlgE pathway was identified in
obligate intracellular bacterial pathogens or symbionts7,8. Despite five chlamydial species distributed in Criblamydiaceae, Wad-
the more advanced genome reduction experienced by the animal- dliaceae, and Parachlamydiaceae families. In this work, we
specific Chlamydiaceae family (0.9 Mpb) in comparison to other demonstrated that the GlgC pathway is impaired in Cri-
protist-infecting Chlamydiales (2–2.5 Mpb), the glycogen meta- blamydiaceae and Waddliaceae. The complete biochemical
bolism pathway appears surprisingly preserved7. This includes characterization of the GlgE pathway in Estrella lausannensis
the three-enzymatic activities required for glycogen biosynthesis: (family Criblamydiaceae) and Waddlia chondrophila (family
GlgC, GlgA, and GlgB9. ADP-glucose pyrophosphorylase (GlgC) Waddliaceae) isolated, respectively, from water in Spain21,22 and
activity that controls the synthesis and level of nucleotide-sugar, from the tissue of an abortive bovine fetus23,24 is reported. Thus,
ADP-glucose, dedicated solely to glycogen biosynthesis. Glycogen despite the intensive reductive genome evolution experienced by
synthase (GlgA) polymerizes nucleotide-sugar into linear α-1,4- these intracellular bacteria our work shows that glycogen bio-
glucan. GlgA activity has a dual function consisting of a primer- synthesis is maintained in all Chlamydiales and suggests a
independent glucan synthesis and glucan elongation at the hitherto understudied function of storage polysaccharides and
non-reducing end of preexisting polymers10. When the primer oligosaccharides in the developmental cycle of all Chlamydiales.
reaches a sufficient degree of polymerization (DP > 15) to fit the
catalytic site of the glycogen branching enzyme (GlgB), glycogen
branching introduced resulting in the appearance of two non- Results
reducing polymer ends that may be further elongated by GlgA. Two different glycogen metabolic pathways are identified in
The repetition of this process results in an exponential increase in the Chlamydiae phylum. To gain insight into Chlamydiae’s
the number of non-reducing ends leading to a particle with a glycogen metabolism, we analyzed 220 genomes from 47 different
32–40 nm diameter11. chlamydial species. As illustrated in Fig. 1a, the synthesis of linear
Until recently, Waddlia chondrophila (family Waddliaceae) as chains of the synthesis of linearboth ADP-glucose pyropho-
well as all members of Criblamydiaceae could be considered as sphorylase (GlgC) and glycogen synthase (GlgA) activities in the
important exceptions to the universal requirement of Chlamy- GlgC pathway while the GlgE pathway relies on trehalose syn-
diales for glycogen synthesis. Indeed, genome analysis indicated thase (TreS), maltokinase (Mak), and maltosyl-1-phosphate
that ad minima the glgC gene was absent from all these bac- transferase (GlgE). The formation of α-1,6 linkages (i.e.,
teria12–14 and that the function of GlgA was possibly also branching points) and glycogen degradation are catalyzed by a set
impaired. Consequently, based on the absence of glycogen of similar enzymes in both pathways that include glycogen
reported for all knockout glgC mutants in bacteria and plants it branching enzyme isoforms (GlgB/GlgB2) and glycogen phos-
was believed that W. chondrophila was defective in glycogen phorylases isoforms (GlgP/GlgP2), glycogen debranching
synthesis15,16. Using transmission electron microscope analysis, enzymes (GlgX) and glycogen phosphorylases isoforms The
we are now reporting numerous glycogen particles within the genomic database used in this study (https://chlamdb.ch) includes
cytosol of W. chondrophila and Estrella lausannensis (family genomes from both cultured and uncultured Chlamydiae species
Criblamydiaceae) EBs, suggesting either another gene encodes a that cover the diversity of the chlamydiae phylum (Fig. 1b). It
phylogenetically distant protein that overlaps the GlgC activity should be stressed out that several families and genus level-
or an alternative glycogen pathway takes place in these lineages encompass exclusively uncultured Chlamydiae species.
Chlamydiales. As a consequence, derived genomes from metagenomic data have
The recent characterization of an alternative glycogen pathway, been carefully re-annotated and subjected to various quality cri-
the so-called GlgE pathway, in Mycobacterium tuberculosis and teria, such as the proportion of core genes as previously reported3.
streptomycetes prompted us to probe chlamydial genomes with Comparative genomics clearly underlined the high prevalence of
homolog genes involved in this pathway17,18. At variance with the a complete GlgC pathway in most Chlamydiales, including all
nucleotide-sugar based GlgC pathway, the GlgE pathway consists members of the Chlamydiaceae family, which has undergone
of the polymerization of α-1,4-glucan chains from maltose-1-P. In massive genome reduction (identified by the letter “d” on Fig. 1b)
2 COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio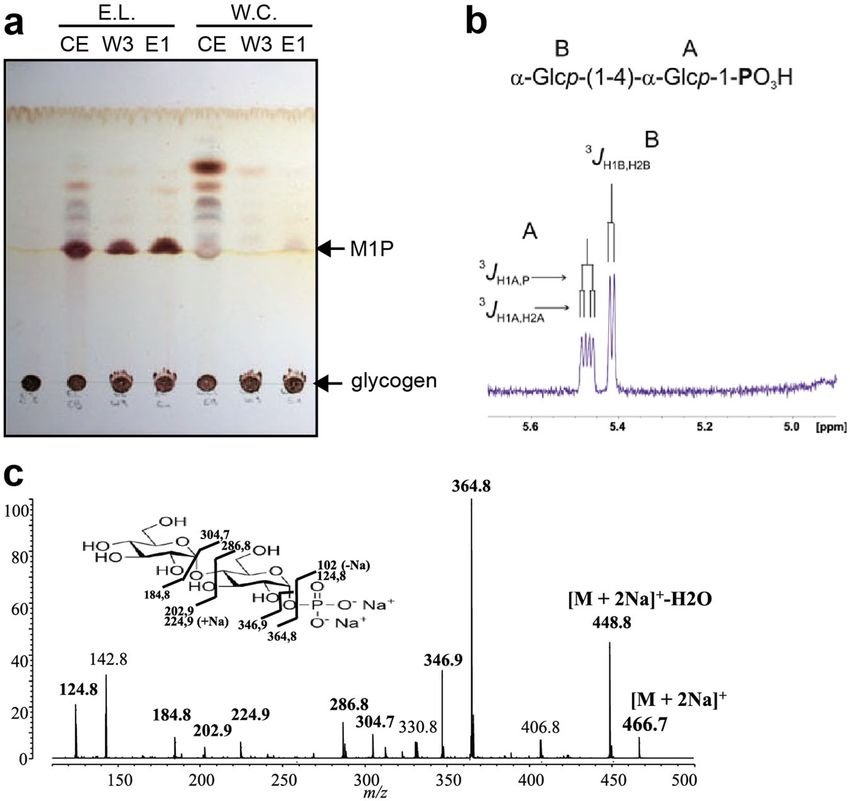
COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y ARTICLE Fig. 1 Comparative genomic analysis of glycogen-metabolizing genes among Chlamydiae phylum. a GlgC- and GlgE-paths represent the main routes of glycogen biosynthesis in prokaryotes. The formation of linear chains of glucosyl units joined by α-1,4 linkages depends on the coupled actions of ADP- glucose pyrophosphorylase (GlgC)/glycogen synthase (GlgA) activities in GlgC-path whereas it relies on the combined actions of trehalose synthase (TreS)/maltokinase (Mak)/maltosyltransferase (GlgE) in GlgE-path. The iteration of glucan synthesis and branching reactions catalyzed by branching enzyme isoforms (GlgB and glgB2) generate a branched polysaccharide. Both α-1,4 and α-1,6 glucosidic linkages are catabolized through synergic actions of glycogen phosphorylase isoforms (GlgP and GlgP2), debranching enzyme (GlgX), and a-1,4-glucanotransferase (MalQ) into glucose-1-phosphate and glucose. b Phylogenic tree of cultured and uncultured Chlamydiae. For each species of the families: a, Ca. Pelagichlamydiaceae; b, Ca. Paralichlamydiaceae; c, Ca. Novochlamydiacae; d, Chlamydiaceae; e, Simkaniaceae; f, Ca. Arenachlamydiaceae; g, Rhabdochlamydiaceae; h, Ca. Limichlamydiaceae; i, Ca. Enkichlamydiaceae; j, Ca. Kinetochlamydiaceae; k, Ca. Motilichlamydiaceae; l, Waddliaceae; m, Criblamydiaceae; n, Parachlamydiaceae, the number of draft (d) or complete (c) genomes and genome completeness expressed in a percentage are indicated between brackets. Homologous genes of the GlgC- and GlgE- pathways are symbolized with colored arrows. The glgP2 gene was identified on the plasmid of S. negevensis and is also present in one of the two available P. neagleriophila genomes. as well as in in the most deeply branching families such as can- estimated to be 71–97% complete, suggesting either the loss of didatus Pelagichlamycidiaceae (“a”) and candidatus Parili- glgC gene or that glgC gene is located in a particular genomic chlamydiaceae (“b”). We noticed that the glg genes are at least 10 region (e.g., next to repeated sequences) that systematically led to kbp apart with a notable exception for glgP and glgC, which are its absence from genome assemblies. Another unexpected result separated by one or two genes in most cases. Caution must be concerns both Waddliaceae (“l”) and Criblamydiaceae (“m”) taken in interpreting the gaps in glycogen metabolism pathways families that encompass Waddlia chondrophila, Estrella lau- of several uncultivated chlamydiae, which likely reflect the fact sannensis, and Criblamydia sequanensis. Genomic rearrange- that many of those genomes are incomplete genomes derived ments caused a sequence of events leading to (i) the deletion of from metagenomic studies (see percentages in brackets in both glgC and glgP genes, (ii) the fusion of glgA with the glgB Fig. 1b). Considering that the GlgC pathway is highly conserved gene, (iii) the insertion of the glgP2 gene encoding a glycogen in nearly all sequenced genomes of the phylum, missing genes phosphorylase isoform at the vicinity of malQ gene. It should be probably reflect missing data rather than gene losses. It is inter- noted that an homolog of glgP2 has also been identified on the esting to note that there is some uncertainty about the presence of plasmids of S. nevegensis and P. naegleriophila. In W. chon- the glgC gene in candidatus Enkichlamydia genome (“j”), as a drophila, another insertion of glgP2 occurred downstream to the complete set of glycogen-metabolizing enzymes were recovered GlgE operon, which may be correlated with partial deletion of except for the gene encoding for ADP-glucose pyrophosphorylase glgP2 at the vicinity of malQ (Fig. 1b). The parsimonious inter- (glgC). This gene is missing from six independent draft genomes pretation of glgC and glgP deletion and glgAglgB fusion is that a COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio 3
ARTICLE COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y
100/1
a b
-/0.98 Planctomycetes_bacterium_GWF2_39_10_an_OHB50689
-/0.93 Candidatus_Brocadia_sinica_an_KXK29055
Deltaproteobacteria_bacterium_RBG_19FT_COMBO_46_12_an_OGP99163
Candidatus_Kuenenia_stuttgartiensis_an_WP_099324375
-/0.83 Spirochaetes_bacterium_DG_61_an_KPJ88710
-/0.96 Actinobacteria_bacterium_RBG_13_35_12_an_OFW47850
-/0.74 Omnitrophica_WOR_2_bacterium_RIFOXYC2_FULL_38_12_an_OGX57864
Omnitrophica_WOR_2_bacterium_RIFCSPHIGHO2_02_FULL_48_11_an_OGX25457
-/0.98 58/0.98 Planctomycetes_bacterium_RBG_16_55_9_an_OHB78517
Planctomycetes_bacterium_GWF2_42_9_an_OHB59987
100/1 Acetomicrobium_thermoterrenum_an_WP_091460818
Synergistales_bacterium_54_24_an_KUK39012
96/1 Estrella_lausannensis_an_WP_098038073
-/0.75 Criblamydia_sequanensis_an_WP_041016796 -/0.99 Planctomycetes_bacterium_GWF2_39_10_an_OHB49437
-/0.72 100/1 100/1 Planctomycetes_bacterium_GWC2_39_26_an_OHB42652
Parachlamydia_sp__C2_an_WP_068470901 -/0.96
Candidatus_Protochlamydia_naegleriophila_an_WP_059059655 68/0.99 Candidatus_Brocadia_sinica_an_WP_052561367
100/1 Elusimicrobia_bacterium_RIFOXYA12_FULL_57_11_an_OGS14391 Elusimicrobia_bacterium_CG02_land_8_20_14_3_00_37_13_an_PIV54013
-/0.99 Elusimicrobia_bacterium_CG1_02_56_21_an_OIO03018 Candidatus_Magnetoovum_chiemensis_an_KJR42025
Planctomycetes_bacterium_GWF2_50_10_an_OHB56553 Geobacteraceae_bacterium_GWC2_48_7_an_OGU00225
100/1 Geoalkalibacter_subterraneus_an_WP_052464481
-/0.78 Geoalkalibacter_ferrihydriticus_DSM_17813_an_KIH78255 Deltaproteobacteria_bacterium_RBG_19FT_COMBO_46_12_an_OGP99164
100/1 Desulfonatronum_thiosulfatophilum_an_WP_092116601 Nitrospira_bacterium_SG8_35_4_an_KPJ97763
Desulfonatronum_lacustre_an_WP_028571164 Chitinispirillum_alkaliphilum_an_KMQ52518_Fibrobacteres
- / 0 . 9 2 64/0.99 Geobacter_sp__M18_an_WP_015718476 100/1 Prosthecochloris_aestuarii_an_WP_012504805
Geobacter_daltonii_an_WP_012646307 Chlorobium_sp__KB01_an_WP_076792731
-/0.99 100/1 Ohtaekwangia_koreensis_an_WP_079686571 Omnitrophica_WOR_2_bacterium_GWF2_38_59_an_OGX23595
-/0.82 Bacteroidetes_bacterium_OLB12_an_KXK25024 Verrucomicrobia_bacterium_13_2_20CM_2_54_15_an_OLB42330
63/1 Flavobacterium_sp__an_PJE44001 -/0.97 Chthoniobacter_flavus_an_WP_006979282
Arcticibacter_svalbardensis_an_WP_016195383 100/1 Verrucomicrobium_sp__3C_an_WP_020494395
98/1 Salinimicrobium_catena_an_WP_093112815 -/0.8 Methylacidiphilum_infernorum_an_WP_012463358
Anditalea_andensis_an_WP_035079135 5 6 / 0 . 7 4 Desulfovibrio_sp__FW1012B_an_WP_009182520
99/1 Verrucomicrobia_bacterium_LP2A_an_WP_024807032
Methylacidiphilum_fumariolicum_an_WP_009060448 -/0.82 Elusimicrobia_bacterium_GWD2_63_28_an_OGR71506
100/1 Pelodictyon_phaeoclathratiforme_an_WP_012509402 Acetomicrobium_hydrogeniformans_an_WP_009200396
-/0.79 Synergistales_bacterium_53_16_an_KUK58635
Chlorobium_phaeobacteroides_an_WP_012473699
100/1 Legionella_wadsworthii_an_WP_031565189 Planctomycetes_bacterium_RBG_16_55_9_an_OHB78518
-/0.74 Legionella_quinlivanii_an_WP_058506242 -/0.91 100/1 Estrella_lausannensis_an_WP_098038072
Waddlia_chondrophila_203299_an_CCB90680 -/0.93 97/1 Criblamydia_sequanensis_an_WP_041016797
-/0.88 100/1 Desulfotignum_phosphitoxidans_an_WP_006964612 Waddlia_chondrophila_an_WP_013182918
Desulfatitalea_sp__BRH_c12_an_KJS30950 100/1 Parachlamydia_sp__C2_an_WP_068470903
100/1 Bdellovibrionales_bacterium_RIFOXYD1_FULL_53_11_an_OFZ82072_ Candidatus_Protochlamydia_massiliensis_an_WP_032124802
Bdellovibrionales_bacterium_RIFOXYC1_FULL_54_43_an_OFZ54055_ Planctomycetes_bacterium_GWF2_50_10_an_OHB56554
-/0.97 -/0.98 Tangfeifania_diversioriginum_an_WP_073169723 100/1 Pelobacter_propionicus_an_WP_011737073
Alkalispirochaeta_alkalica_an_WP_018526923 Geobacter_metallireducens_an_WP_004513920
100/1 Chloroflexi_bacterium_RBG_16_60_22_an_OGO43732 100/1 Salinimicrobium_catena_an_WP_093112816
Chloroflexi_bacterium_RBG_16_56_11_an_OGO31222 -/0.98
100/1 Nitrosomonas_sp__Nm51_an_WP_090330979 Leeuwenhoekiella_palythoae_an_WP_072982091
I
-/0.91 Nitrosomonas_aestuarii_an_WP_090699605 Pontibacter_chinhatensis_an_WP_092099346
100/1 Nitrosococcus_watsonii_an_WP_013221180 -/0.99 80/1 -/0.71 Flammeovirgaceae_bacterium_311_an_WP_061984987
Candidatus_Nitrosoglobus_terrae_an_WP_096526708 93/1 Ohtaekwangia_koreensis_an_WP_079686037
100/1 Betaproteobacteria_bacterium_SG8_41_an_KPK19593 95/1 99/1 Dyadobacter_crusticola_an_WP_031527357
Betaproteobacteria_bacterium_RIFCSPLOWO2_02_FULL_62_79_an_OFZ90144 Flavisolibacter_tropicus_an_WP_066401371
Candidatus_Contendobacter_odensis_an_WP_081756447 Phaeodactylibacter_xiamenensis_an_WP_044224133
-/0.91 Variovorax_paradoxus_an_WP_021006725 -/0.93 Spirochaeta_africana_an_WP_041396969
-/0.93 Sulfuricella_denitrificans_an_WP_009205429
-/0.84 Betaproteobacteria_bacterium_RIFCSPLOWO2_02_FULL_65_24_an_OGA28709 Desulfovibrionaceae_bacterium_CG1_02_65_16_an_OIO01428
-/0.95 Thiohalomonas_denitrificans_an_WP_092994834 Sulfobacillus_thermosulfidooxidans_an_WP_028962253
-/1 Legionella_wadsworthii_an_WP_031565191
-/0.74 Sulfurifustis_variabilis_an_BAU48057
-/0.7 Nitrospinae_bacterium_CG11_big_fil_rev_8_21_14_0_20_56_8_an_PIQ95900 Bdellovibrio_sp__SKB1291214_an_WP_088616901_
Nitrococcus_mobilis_an_WP_005004716 100/0.99 Sorangium_cellulosum_an_KYF78085
-/0.96 69/0.99 Chondromyces_apiculatus_an_WP_044250042
Deltaproteobacteria_bacterium_SM23_61_an_KPK91810 90/0.99
-/0.99 Deltaproteobacteria_bacterium_RBG_13_60_28_an_OGP70080 Sorangiineae_bacterium_NIC37A_2_an_OQX66705
Deltaproteobacteria_bacterium_CG2_30_66_27_an_OIP35186 Haliangium_ochraceum_an_WP_012829192
50/0.99 Dehalococcoidia_bacterium_SG8_51_3_an_KPK23489 Nitriliruptor_alkaliphilus_an_WP_052667796
-/0.94 Chloroflexi_bacterium_RBG_13_53_26_an_OGO03925 100/1
-/0.78 Planktothrix_an_WP_026788709
-/0.93 1 0 0 / 1Dehalococcoidia_bacterium_SM23_28_2_an_KPK47014 - / 0 . 9 6 Arthrospira_sp__PCC_8005_an_WP_008052613
100/1 Chloroflexi_bacterium_13_1_40CM_66_19_an_OLC21315 -/0.99 Oscillatoria_sp__PCC_10802_an_WP_017720816
-/0.9 Actinobacteria_bacterium_13_2_20CM_2_66_6_an_OLC00509 Spirulina_major_an_WP_072623136
-/0.6 Actinobacteria_bacterium_13_1_40CM_66_12_an_OLC15073
Anaeromyxobacter_sp__RBG_16_69_14_an_OFX21687 1 0 0 / 1- / 0 . 9 6 Leptolyngbya_sp__hensonii_an_WP_075596519
-/0.99 Sphaerobacter_thermophilus_an_WP_041399600 95/1 Richelia_intracellularis_an_CDN16121
Nitrolancea_hollandica_an_WP_008480772 100/1 100/1 Fischerella_muscicola_an_WP_016861395
-/0.98 8 4 / 0 . 9 9 Elusimicrobia_bacterium_RIFOXYA2_FULL_53_38_an_OGS29251 Rivularia_sp__PCC_7116_an_AFY54611
- / 0 . 7 4- / 0 . 6 Elusimicrobia_bacterium_RIFCSPLOWO2_12_FULL_59_9_an_OGS04265 -/0.91 -/0.93 Kamptonema_an_WP_007355980
Nitrospira_moscoviensis_an_WP_053381139 -/0.96 Synechococcus_an_WP_099813207
-/0.6 Chloroflexi_bacterium_RBG_16_54_18_an_OGO34720 Deltaproteobacteria_bacterium_SM23_61_an_KPK91809
-/0.96 Acidobacteriaceae_bacterium_KBS_96_an_WP_020722338 52/0.99 bacterium_F11_an_OVE75537
-/0.79 Opitutus_terrae_an_WP_012373931 Planctomycetes_bacterium_GWF2_41_51_an_OHB49974
Acidobacteria_bacterium_13_1_40CM_2_56_11_an_OLD71809 -/0.4
70/- Crinalium_epipsammum_an_WP_015204971 -/0.6 Deltaproteobacteria_bacterium_GWC2_65_14_an_OGP33450
100/1 Coleofasciculus_chthonoplastes_an_WP_006100233 Deltaproteobacteria_bacterium_13_1_40CM_4_68_19_an_OLC78335
Microcoleus_sp__PCC_7113_an_WP_015183597 Dehalococcoidia_bacterium_SM23_28_2_an_KPK47010
Acidobacteria_bacterium_RIFCSPLOWO2_02_FULL_59_13_an_OFW26834 -/0.4 Deltaproteobacteria_bacterium_RBG_13_65_10_an_OGP84727
-/0.5 Synechococcus_an_WP_011429175 Gemmatimonas_sp__SM23_52_an_KPK83041
Betaproteobacteria_bacterium_RIFCSPLOWO2_12_FULL_65_14_an_OGA61708 1 0 0 / 1 Elusimicrobia_bacterium_GWA2_69_24_an_OGR56405
-/0.76 Acidobacterium_ailaaui_an_WP_026443321 Paludisphaera_borealis_an_WP_076350284
87/0.99 Thermorudis_peleae_an_WP_038040328 -/0.4 Desulfatitalea_sp__BRH_c12_an_KJS30952
Thermomicrobium_roseum_an_WP_015922872 -/0.76 Chloroflexus_an_WP_012257046
-/0.88 Candidatus_Thiosymbion_oneisti_an_WP_089729968 -/0.94 Thermorudis_peleae_an_WP_038040329
Leptospirillum_ferriphilum_an_WP_051613765 -/0.85 Actinobacteria_bacterium_13_2_20CM_2_66_6_an_OLC00511
100/1 Chondromyces_crocatus_an_WP_050435754
Chondromyces_apiculatus_an_WP_044250040 Sphaerobacter_thermophilus_an_WP_012871575
93/1 100/1 Chloroflexi_bacterium_RBG_16_50_9_an_OGO24493
Labilithrix_luteola_an_AKV00723
Sandaracinus_amylolyticus_an_WP_053238684 Chloroflexi_bacterium_RBG_13_53_26_an_OGO01422
52/0.97 Haliangium_ochraceum_an_WP_012829193 Candidatus_Koribacter_versatilis_an_WP_011522289
-/0.99 100/1 Truepera_radiovictrix_an_WP_013177912 Chloroflexi_bacterium_G233_an_WP_098503461
Rubrivirga_marina_an_WP_095508616 81/1 81/0.99 Pseudomonas_sp__MOIL14HWK12_I2_an_WP_027599229
Chloroflexus_islandicus_an_WP_066786530 100/1 Granulibacter_bethesdensis_an_WP_072548311
Ralstonia_solanacearum_an_OAK89975 6 1 / 0 . 9 9
-/0.91 73/1 Belnapia_moabensis_an_WP_043335788
-/0.95 Bordetella_flabilis_an_ANN77580 Rhizobiales_bacterium_YIM_77505_an_WP_037015305
-/0.8 Pigmentiphaga_sp__NML080357_an_WP_087782425 Azospirillum_brasilense_an_WP_014240599
Acidiphilium_cryptum_an_WP_012040420 81/1
-/0.98 96/1 Rhizobiales_bacterium_GAS191_an_WP_092918464 Rhodopseudomonas_palustris_an_WP_011159182
Bosea_sp__Root483D1_an_WP_082613402 100/1 Rhodospirillum_rubrum_an_WP_011389295
100/1 99/1 Betaproteobacteria_bacterium_RIFCSPLOWO2_02_64_14_an_OGA09598
77/0.97 Roseomonas_gilardii_an_WP_075799010 100/1
Noviherbaspirillum_humi_an_WP_089398766 -/0.9 Acidovorax_avenae_an_WP_053842426
Methyloceanibacter_caenitepidi_an_WP_082025375 Nitrosococcus_oceani_an_WP_002811514
100/1 Rhodocista_sp__MIMtkB3_an_WP_075772009 100/1 delta_proteobacterium_ML8_D_an_OPL17032
Rhodospirillales_bacterium_URHD0088_an_WP_051474484 Desulfobacterales_bacterium_GWB2_56_26_an_OGR16935
93/1 57/0.84 Azospirillum_sp__B506_an_WP_042704400 -/0.89 Acidobacteria_bacterium_13_1_40CM_65_14_an_OLC40998
100/1 Rubrobacter_xylanophilus_an_WP_011563308 70/0.91 100/1 Granulicella_pectinivorans_an_WP_089843144
94/0.99 Rubrobacter_radiotolerans_an_AHY45474 98/1 Acidobacteria_bacterium_KBS_146_an_WP_051628343
Caenispirillum_salinarum_an_WP_009541642 Gammaproteobacteria_bacterium_RIFCSPLOWO2_02_FULL_56_15_an_OGT78108
-/0.99 -/0.96 Rhodopseudomonas_palustris_an_WP_041807137 -/0.87
-/0.85 Candidatus_Rokubacteria_bacterium_RIFCSPLOWO2_02_FULL_73_56_an_OGL07526 Singulisphaera_sp__GP187_an_WP_074306456
100/1 Candidatus_Rokubacteria_bacterium_13_1_20CM_2_70_7_an_OLE39328 Cystobacter_fuscus_an_WP_095987764
-/0.93 75/0.98 Ornithinimicrobium_pekingense_an_WP_097186925
Candidatus_Rokubacteria_bacterium_13_1_20CM_4_70_14_an_OLD75031 100/1
-/0.99 Candidatus_Rokubacteria_bacterium_RIFCSPLOWO2_12_FULL_71_19_an_OGL20528 88/0.99 Actinomycetales_bacterium_JB111_an_WP_087061270
58/0.73 Omnitrophica_bacterium_RIFCSPHIGHO2_12_FULL_44_12_an_OGW91399 61/1 Kineosporia_sp__R_H_3_an_WP_088318884
-/0.97 Candidatus_Magnetoovum_chiemensis_an_KJR42939 -/0.6 Cutibacterium_acnes_an_WP_023487572
Terrimicrobium_sacchariphilum_an_WP_075079925 64/0.99 Sporichthya_polymorpha_an_WP_051098396
Candidatus_Accumulibacter_aalborgensis_an_SBT09740 Actinopolymorpha_alba_an_WP_020579298
99/1 Streptomyces_scabrisporus_an_WP_078979218 -/0.98 Nocardioides_terrae_an_SFB71474
II
98/1 -/0.87 Streptomyces_jeddahensis_an_WP_078066827 100/1 -/0.9 Thermomonospora_chromogena_an_WP_093257445
-/0.76 Sporichthya_polymorpha_an_WP_028984880 92/0.95 Marinactinospora_thermotolerans_an_WP_078762835
57/0.89 -/0.91 Actinopolymorpha_singaporensis_an_WP_092656372 -/0.99 Actinomadura_echinospora_an_SEF79781
Angustibacter_sp__Root456_an_WP_056672890 55/0.87 Actinomadura_oligospora_an_WP_084338939
Actinomadura_hibisca_an_WP_067481742 54/0.91
Frankia_sp__DC12_an_WP_045876178 -/0.98 Pseudonocardia_thermophila_an_WP_073455812
100/1 95/1 Actinoplanes_awajinensis_an_WP_067696287
100/1 Tsukamurella_tyrosinosolvens_an_WP_068526334
Rhodococcus_sp__NCIMB_12038_an_WP_087561062 75/0.99 Allokutzneria_albata_an_WP_030429025
74/0.99
Amycolatopsis_saalfeldensis_an_WP_091627791 Candidatus_Rokubacteria_bacterium_RIFCSPLOWO2_02_FULL_73_56_an_OGL07525 0.2
90/0.98 Micromonospora_haikouensis_an_WP_091280303 bacterium_JKG1_an_WP_029215667
Thermobaculum_terrenum_an_WP_012874288 -/0.75 Acidobacteria_bacterium_13_1_20CM_3_53_8_an_OLE54927
-/0.98 60/0.91 Nannocystis_exedens_an_SFE16387
97/1 Gemmatirosa_kalamazoonesis_an_AHG91353
Sorangiineae_bacterium_NIC37A_2_an_OQX66829
Gemmatimonas_sp__SM23_52_an_KPK83069
100/1 Desulfobulbaceae_bacterium_BRH_c16a_an_KJS01348
98/1 Desulfobacterales_bacterium_GWB2_56_26_an_OGR16942
Deltaproteobacteria_bacterium_RIFOXYD12_FULL_57_12_an_OGQ94265
100/1 Singulisphaera_sp__GP187_an_SIN92619
Singulisphaera_acidiphila_an_WP_015249074 0.09
_an_AMV19242
Isosphaera_pallida_an_WP_013564039
-/0.5 Acidobacteria_bacterium_RIFCSPLOWO2_02_FULL_67_36_an_OFW08021
65/0.98 Acidobacteria_bacterium_RIFCSPLOWO2_02_FULL_67_21_an_OFW13256
Fig. 2 Phylogenetic analysis of GlgE and TreS-Mak. Both phylogenetic trees of GlgE (a) and TreS-Mak (b) were performed with Phylobayes under the
C20 + Poisson model. We then mapped onto the nodes ML bootstrap values obtained from 100 bootstrap repetitions with LG4X model (left) and Bayesian
posterior probabilities (right). Bootstrap values >50% are shown, while only posterior probabilities >0.6 are shown. The trees are midpoint rooted. The
Chlamydiales are displayed in purple. The scale bar shows the inferred number of amino-acid substitutions per site.
single-deletion event led to the loss of DNA fragment bearing glgP Streptomycetes), respectively. For Tres-Mak phylogeny (Fig. 2b),
and glgC genes between glgA and glgB. However, despite many chlamydial Tres-Mak sequences cluster together, suggesting a
variations, the genomic configuration compatible with this par- common origin, however, with a low statistical support (pp =
simonious hypothesis was never observed in extant Chlamydiae 0.93). Although the origin of GlgE operon cannot be pinpointed
(Supplementary Table 1). Rather, such genomic rearrangements in our phylogenetic analysis, conceivable scenarios are that either
are associated with a novel glycogen pathway based on the GlgE (i) the GlgE operon reflects vestigial metabolic function of the
operon described in mycobacteria and also observed in Prototo- ancestral chlamydiae and then has been lost in most families or
chlamydia naegleriophila and Protochlamydia phocaeensis (syn. (ii) this operon was acquired by a lateral gene transfer event from
Parachlamydia C2). All three genes are clustered in the classical a member of the PVC phylum by the common ancestor of
unfused glgE-treSmak-glgB2 operon arrangement in Waddliaceae Parachlamydiaceae, Waddliaceae, and Criblamydiaceae families.
and Criblamydiaceae, while the glgB2 gene is missing in the
Parachlamydiaceae operons (Fig. 1b). The occurrence of the GlgE
pathway restricted to Parachlamydiaceae, Waddliaceae, and Cri- The classical GlgC pathway is not functional in E. lausannensis
blamydiaceae families beg the question of its origin in Chlamy- and W. chondrophila. To further investigate whether his-tagged
diales. To get some insight on this issue, phylogenetic trees of recombinant proteins GlgA–GlgB of E. lausannensis and W.
TreS-Mak and GlgE have been inferred using the phylobayes chondrophila are functional, glycogen synthase activities at the N-
method (Fig. 2). The GlgE phylogeny shows that even if the terminus domain were assayed by measuring the incorporation of
Chlamydiae sequences are split into two with W. chondrophila on labeled 14C-glucosyl moiety from ADP- or UDP-14C-glucose
one side and the other sequences on the other side, which reflects onto glycogen and by performing a specific nondenaturing PAGE
likely lateral gene transfer events with other bacteria, chlamydial or zymogram to visualize glycogen synthase activities. After
glgE sequences might still be monophyletic since the only strongly separation on native-PAGE containing glycogen, recombinant
supported node (marked as red star) with a posterior probability proteins were incubated in the presence of 1.2 mM ADP-glucose
(pp) higher than 0.95 (pp = 0.99) unifies all chlamydiae sequen- or UDP-glucose, glycogen synthase activities are visualized as
ces (Fig. 2a), which has also been confirmed using the LG model dark activity bands after soaking gels in iodine solution (Fig. 3).
(Supplementary Data 1). The phylogeny analysis highlights Enzymatic assays and zymogram analyses show that the
that GlgE sequences can be classified into classes I and II, com- glycogen synthase domain of the chimeric GlgA–GlgB of W.
prising Chlamydiales and Actinomycetales (i.e., mycobacteria, chondrophila (hereafter GlgA–GlgB-WC) is functional but highly
4 COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio
COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y ARTICLE
tagged recombinant proteins of GlgE of Estrella lausannensis
(hereafter GlgE-EL) and Waddlia chondrophila (hereafter
GlgE-WC) were expressed and further characterized (Supple-
mentary Fig. 2). As described in previous studies, GlgE of
Mycobacteria mediates the reversible reaction consisting of the
release of maltose-1-phosphate in the presence of orthopho-
sphate and α-glucan polysaccharide. Both GlgE-EL and GlgE-
WC were incubated in presence of glycogen from rabbit liver
and orthophosphate. After overnight incubation, reaction
products were analyzed on thin-layer chromatography and
sprayed with oricinol-sulfuric acid (Fig. 4a). A fast migration
product capable of interacting with orcinol-sulfuric acid was
clearly synthesized in crude extract (CE), in washing # 3 (W3),
and in the purified enzyme fraction (E1) of the GlgE-EL
sample. A barely visible product is only observed in the pur-
ified fraction (E1) of GlgE-WC. The compound produced by
GlgE-EL in presence of glycogen and orthophosphate was
further purified through different chromatography steps and
Fig. 3 Zymogram analysis of glycogen synthase activities. Total crude subjected to mass spectrometry and proton-NMR analyses
extracts of the recombinant proteins of GlgA of Escherichia coli (GlgA_E. (Fig. 4b, c). The combination of these approaches confirms
coli), GlgA–GlgB of E. lausannensis (GlgA-GlgB_EL), and W. chondrophila that GlgE of E. lausannensis as well as W. chondrophila
(GlgA-GlgB_WC) were separated by native-PAGE containing 0.6% (w/v) (Supplementary Fig. 3) catalyzes the formation of a compound
glycogen. The native gels were then incubated with 1.2 mM ADP-glc or with a molecular weight of 422 Da (Fig. 4c) corresponding to
1.2 mM UDP-glc. Glycogen synthase activities are seen after iodine staining α-maltose-1-phosphate, as shown on the proton and phos-
as dark bands. phorus spectra (Fig. 4b). In order to carry out enzymatic
characterization of GlgE activities, identical purification pro-
specific for ADP-glucose (0.70 nmol of incorporated glucose. cesses were scaled up to purify enough M1P, free of inorganic
min−1.mg−1) and has little to no activity using UDP-glucose as phosphate and glucan.
substrate. As predicted, the activity of the truncated glycogen
synthase in E. lausannensis was not detected on activity gels or
during enzymatic assays (Supplementary Fig. 1a). Kinetic parameters of GlgE activity of E. lausannensis in the
We further investigated whether the branching activity domain biosynthetic direction. Because the his-tagged recombinant
at the carboxyl terminal of chimeric protein GlgA–GlgB of W. GlgE-WC expresses very poorly and the specific activity of GlgE-
chondrophila (GlgA–GlgB-WC) was functional. To check this, the WC was ten times lower than GlgE-EL, kinetic parameters were
same chimeric GlgA–GlgB-WC sample previously analyzed was determined in the synthesis direction, i.e., the transfer (amount)
incubated with ADP-glucose (3 mM) and maltoheptaose (10 mg. of maltosyl moieties onto non-reducing ends of glucan chains,
mL−1) overnight. Subsequently, the appearance of branching exclusively for GlgE-EL. Transfer reactions are associated with the
point (i.e., α-1,6 linkages) onto growing linear glucans can be release of inorganic phosphate that can be easily monitored
specifically observed by the resonance of protons onto carbon 6 at through the sensitive malachite green assay. Thus, under variable
4.9 ppm using proton-NMR analysis. However, as depicted in M1P concentrations and using fixed concentrations of glycogen
Supplementary Fig. 1c, we did not observe any signal, suggesting or maltoheptaose, the GlgE-EL activity displays allosteric beha-
that branching enzyme activity domain is not functional despite vior indicating positive cooperativity, which has been corrobo-
an active glycogen synthase domain. This result is consistent with rated with Hill coefficients that were above 1 (Fig. 5a, b). In
several reports indicating that the amino-acid length at the N- agreement with this, the molecular weight of native GlgE-EL
terminus of branching enzyme affects its catalytic properties25–27. determined either by size exclusion chromatography or by native-
In regard to this information, the glycogen synthase domain PAGE containing different acrylamide concentrations (5, 7.5, 10,
extension located at the N-terminus prevents, probably, the and 12.5%) indicates an apparent molecular weight of 140–180
branching enzyme activity of GlgA–GlgB. Thus α-1,6 linkages or kD, respectively, corresponding to the formation of dimer species
branching points are likely to be the result of the GlgB2 isoform while no monomer species of 75 kD were observed (Fig. 5e, f).
activity found in both instances. Altogether, these data strongly The enzyme exhibited S0.5 values for M1P that vary from 0.16 ±
suggest that the classical GlgC pathway is not functional in both 0.01 mM to 0.33 ± 0.02 mM if DP7 and glycogen are glucan
Waddliaceae and Criblamydiaceae families. acceptors, respectively. However, using M1P at saturating con-
centration, GlgE-EL displays Michaelis kinetics (nH close to 1)
indicating a non-cooperative reaction (Fig. 5c, d). In such
GlgE-like genes of E. lausannensis and W. chondrophila
experimental conditions, the apparent Km values for glycogen and
encode α-maltose-1-phosphate: 1,4-α-D-glucan 4-α-D-
DP7, 2.5 ± 0.2 mg.mL−1 and 3.1 ± 0.2 mM, respectively, were
maltosyltransferase. Based on phylogenetic analysis of GlgE,
similar to the apparent Km value of glycogen synthase (GlgA) that
both GlgE of mycobacteria (Actinobacteria) and Chlamydiales
synthesizes α-1,4 linkages from ADP-glucose29.
are phylogenetically distant (Fig. 2a). GlgE of M. tuberculosis
displays 43–40% of identity with GlgE-like sequences of E.
lausannensis and W. chondrophila, respectively. Because GlgE De novo glycogen synthesis: GlgE activity enables the initiation
activity belongs to the large and diversified Glycosyl Hydrolase and elongation of glucan. At variance with eukaryotic glycogen
13 family consisting of carbohydrate active enzymes with quite synthase, prokaryotic glycogen synthase (GlgA) does not require
diverse activities such as α-amylases, branching enzymes, the presence of a short α-1,4-glucan or primer to initiate glycogen
debranching enzymes28, we undertook to demonstrate that biosynthesis10. In absence of GlgA and GlgC activity in E. lau-
these enzymes displayed catalytic properties similar to those sannensis and in the absence of GlgC and thus of ADP-glucose
previously described for GlgE of mycobacteria. Histidine- supply in W. chondrophila, this raised the question of the ability
COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio 5ARTICLE COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y
Fig. 4 Characterization of compounds released by recombinant GlgE of Estrella lausannensis. a Both histidine-tagged recombinant GlgE-EL and GlgE-WC
proteins were purified and incubated in presence of glycogen and inorganic phosphate. The overnight reaction products from crude extract (CE), third
washing step (W3) purified enzymes (E1) were subjected to thin-layer chromatography analysis. Orcinol-sulfuric spray reveals a notable production of M1P
with recombinant GlgE-EL, which is less visible with recombinant GlgE-WC. b Part of 1D-1H-NMR spectrum of maltoside-1-phosphate. α-anomer
configuration of both glucosyl residues was characterized by their typical homonuclear vicinal coupling constants (3JH1A,H2A and 3JH1B,H2B) with values of
3.5 and 3.8 Hz, respectively. A supplementary coupling constant was observed for α-anomeric proton of residue A as shown the presence of the
characteristic doublet at 5.47 ppm. This supplementary coupling constant is due to the heteronuclear vicinal correlation (3JH1A,P) between anomeric proton
of residue A and phosphorus atom of a phosphate group, indicating that phosphate group was undoubtedly O-linked on the first carbon of the terminal
reducing glucosyl unit A. The value of this 3JH1A,P was measured to 7.1 Hz (Table 1). c MS-MS sequencing profile of M1P. The molecular ion [M + 2Na]+ at
m/z 466.7 corresponding to M1P + 2 sodium was fractionated in different ions. Peak assignments were determined according to panel incrusted in (c).
electrophoresis. We noticed that the C1 phosphate group
Table 1 1Hchemical shifts (ppm) of anomeric protons and prevented the labeling of M1P with fluorescent probe. Never-
their first vicinal coupling constants (J Hz) recorded at 300 theless, the level of maltose released from M1P due to the
spontaneous dephosphorylation during the experiment was
K in D2O displayed on Fig. 4b.
appreciated by performing incubations with denatured enzymes
3J 3J
(Fig. 6a, h). Incubation experiments show that both GlgE
Residue H1 H1,H2 H1,P activities possess either an α-1,4-glucanotransferase or maltosyl-
B α-Glcp(1–>4) 5.403 3.8 – transferase activities depending on the presence of M1P. When
A α-Glcp(1–>P) 5.459 3.5 7.1 M1P is omitted, GlgE activities harbor an α-1,4-glucanotransfer-
ase activity exclusively with glucans composed of six or seven
glucose units (DP6 or DP7). Interestingly, after 1 h or overnight
of GlgE activities to substitute for GlgA with respect to the incubation, DP6 or DP7 are disproportionated with one or two
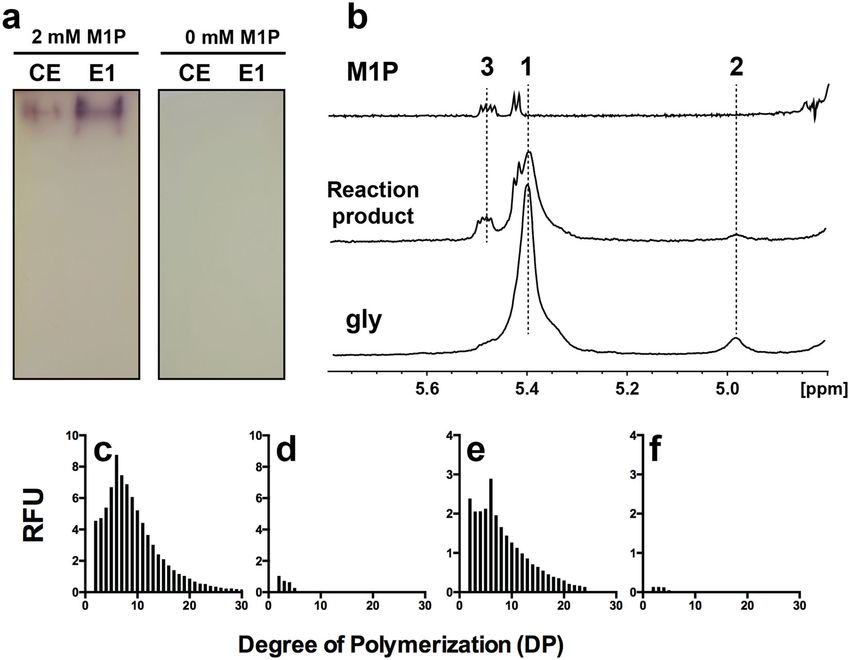
priming of glycogen biosynthesis. To establish whether GlgE maltosyl moieties leading to the release of shorter (DPn-2)
activities are able to prime glucan synthesis, both his-tagged and longer glucans (DPn + 2) (Fig. 6g, n and Supplementary
GlgE-EL (3.51 nmol of Pi released.min−1) and GlgE-WC Figs. 4 and 5). The limited number of transfer reactions
(1.38 nmol of Pi released.min−1) were incubated with 1.6 mM emphasizes probably a side reaction of GlgE activities. The
M1P in the presence of 5 mM of various glucan chains with a α-glucanotransferase activity can be also appreciated on native-
degree of polymerization (DP) of 1 to 7. Identical incubation PAGE containing glycogen. Chain length modification of external
experiments were conducted with GlgE recombinant proteins glucan chains of glycogen results in increase of iodine interactions
except M1P was omitted in order to appreciate α-1,4-glucano- visualized as a brownish activity band (Supplementary Fig. 6a).
transferase or disproportionnating activity (Fig. 6 and Supple- After 1 h of incubation (Supplementary Figs. 4 and 5), both GlgE
mentary Figs. 4 and 5). activities enable the transfer the maltosyl moiety of M1P onto the
After incubation, the reduced-ends of glucan chains were glucan primer with a DP ≥ 3 (Fig. 6d–f, k–m and Supplementary
labeled with fluorescent charged probe (APTS) and separated Figs. 4 and 5). Interestingly, for a longer period of incubation
according to their degree of polymerization by capillary time, both GlgE activities behave either like processive or
6 COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbioCOMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y ARTICLE Fig. 5 Kinetic parameters of recombinant GlgE-EL. GlgE activity was assayed spectrophotometrically by monitoring the release of inorganic orthophosphate (Pi). Data are presented as individual data points of n independent experiments. M1P saturation plots for GlgE-EL were determined in the presence of 10 mM of maltoheptaose (DP7) (n = 3) (a) or 10 mg.mL−1 of glycogen (n = 3) (b). At low M1P concentrations (panels), GlgE-EL activity behaves as allosteric enzyme with Hill coefficients (nH) of 1.6 and 1.5, respectively (fit shown as the solid line giving r2 = 0.98). The S0.5 (M1P) values for GlgE-EL were determined at 0.33 ± 0.02 mM and 0.16 ± 0.01 mM in the presence of DP7 and glycogen, respectively. In the presence of 2 mM M1P, both DP7 (n > 3) (c) and glycogen (n = 2) (d) saturation plots are conformed to the Michaelis–Menten behavior (nH close to 1) with Km values of 3.1 ± 0.2 mM and 2.5 ± 0.2 mg.mL−1, respectively. The apparent molecular weight of GlgE-EL was determined by native-PAGE (e) and size exclusion chromatography (Superose 6 Increase GL 10/300) (f) at 140.9 and 180 kDa, respectively, suggesting a dimer of GlgE (76 kD). distributive enzymes depending on the initial degree of we can hypothesize that the low affinity of short glucan primers polymerization of the glucan primer. The processive behavior (DP < 4) for glucan-binding sites favors probably iterative of GlgE enzymes was unexpected since GlgE activity has been transferase reactions onto the same acceptor glucan (i.e., reported to operate a double displacement reaction (i.e., Ping- processive-like mode) resulting in the synthesis of long glucan Pong mechanism) involving the release of (2 + n) glucan prior to chains whereas glucan primers with DP ≥ 4 compete strongly for the next reaction18. As depicted in Fig. 6, the synthesis of very the binding site leading to a distributive mode. The discrepancy long glucan chains, up to 32 glucose residues suggests that both between GlgE-EL and GlgE-WC to synthesize long glucan chains GlgE-EL and GlgE-WC undergo processive-like elongation in the absence (Fig. 6b, i) or in the presence of glucose (DP1) activities in the presence of maltose (DP2) or maltotriose (Fig. 6c, j) might be explained by a higher amount of free maltose (DP3). In contrast, when both GlgE activities are incubated in observed in denatured GlgE-WC samples (Fig. 6a) by comparison presence of glucan primers with DP ≥ 4, the latter add and to denatured GlgE-EL samples (Fig. 6h). Despite having taken all immediately release a glucan primer (DP) with an increment of precautions (same M1P preparation, buffer pH 7), spontaneous two glucose moieties (DPn + 2) that leads to a distributive dephosphorylation of M1P occurred more significantly in GlgE- elongation behavior. The mechanism underlying the switch WC samples. We therefore conclude that initial traces of maltose between processive-like and distributive elongation activities in GlgE-WC samples facilitate the synthesis of long glucan chains reflects, probably, a competition of glucan primers for the in the absence (Fig. 6b) or in the presence of glucose (DP1) glucan-binding site in the vicinity of the catalytic domain. Thus, (Fig. 6c). To test this hypothesis, crude extract (CE) and purified COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio 7
ARTICLE COMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y
Fig. 6 FACE analyses of enzymatic reaction products of GlgE activity of
1×105
WC 1×105
EL W. chondrophila, WC and E. lausannensis, EL. a, h Spontaneous
a h dephosphorylation of M1P during overnight incubation was estimated by
M1P M1P incubating denatured GlgE enzymes in buffer containing 1.6 mM M1P. The
5×104 2 5×104
transfer of maltosyl moieties from M1P at 1.6 mM onto non-reducing ends
of glucan acceptors (5 mM) were determined in absence of glucan
2 acceptor, DP0 (b, i) or in presence of glucose, DP1 (c, j), maltotriose, DP3
0 0 (d, k), maltotetraose, DP4 (e, l), and maltoheptaose, DP7 (f, m). g, n α-1,4-
5 10 15 20 25 5 10 15 20 25
1×105
glucanotransferase activities of GlgE were determined by incubating 5 mM
1×105
b i of maltoheptaose, DP7, without maltose-1-phosphate. Numbers on the top
of fluorescence peaks represent the degree of polymerization of glucan
M1P+DP0 M1P+DP0 chains.
5×104 5×104
32
0 0 through interaction with iodine molecules) is detected by soaking
5 10 15 20 25 5 10 15 20 25
the gel in iodine solution. As depicted in Fig. 7a, the synthesis of
1×107 1×107
glucan chains catalyzed by GlgE-EL appears exclusively as dark-
1×105 c 1×105
j blue activity bands inside native-PAGE incubated with 2 mM
M1P+DP1 M1P + DP1
Relative Fluorescence Units (RFU)
M1P and not in the absence of M1P.
5×104 5×104
Altogether, these results suggest that GlgE activities are able to
synthesize de novo a sufficient amount of long linear glucans
32
from maltose-1-phosphate. We cannot exclude the role of
0
5 10 15 20 25
0
5 10 15 20 25
maltose in the initiation process of glucan synthesis as glucan
1×107 1×107
acceptor since spontaneous dephosphorylation of M1P is
d k unavoidable. We further carried out a series of experiments that
4×105 4×105
consisted to synthesize in vitro high molecular branched glucans
M1P+DP3 M1P+DP3 by incubating both recombinant glycogen branching enzyme of
2×105 2×105 W. chondrophila (GlgB-WC: Supplementary Fig. 6b) and GlgE-
32 32
EL in the presence of M1P. After overnight incubation, the
appearance of α-1,6 linkages or branching points were directly
0 0
5 10 15 20 25 5 10 15 20 25 measured by subjecting incubation product on proton-NMR
1×107 4 1×107 4 analysis (Fig. 7b). In comparison with M1P and glycogen as
2×106 e 2×106 l controls, proton-NMR spectrum of incubation products shows a
typical profile of glycogen-like with signals at 5.6 and 4.9 ppm of
M1P+DP4 M1P+DP4 proton involved in α-1,4 and α-1,6 linkages. This branched
1×106 6 1×106 6 polysaccharide material was further purified and incubated with a
8 8 commercial isoamylase type debranching enzyme (Megazyme)
0 0
that cleaves off α-1,6 linkages or branching points. Released linear
5 10 15 20 25 5 10 15 20 25 glucan chains were labeled with APTS and separated according to
4×106 4×106 the degree of polymerization by capillary electrophoresis. The
f 5
5 m chain length distribution (CLD) of synthesized polysaccharides
3×106 3×106
7 M1P+DP7 M1P+DP7 (Fig. 7c) was compared with glycogen from bovine liver (Fig. 7e).
7 As control, the amounts of free linear glucans were estimated by
2×106 2×106
9
9 analyzing the APTS-labeled samples not incubated with com-
1×106 11 1×106 11 mercial debranching enzyme (Fig. 7d, f). In absence of notable
17 17 amount of free glucan chains (Fig. 7d), the in vitro synthesized
0 0
5 10 15 20 25 5 10 15 20 25 polysaccharide harbors a typical CLD similar to animal glycogen
7 4×106
7 with monomodal distribution and maltohexaose (DP6) as most
4×106
g n abundant glucan chains. Altogether, these results confirm that
3×106 3×106
DP7 DP7 GlgE activities display an in vitro function similar to that of
2×106 5 2×106 5 glycogen synthase (GlgA) for initiating and elongating the
9 9 growing glycogen particles.
1×106 1×106
11
11
0 0
5 10 15 20 25 5 10 15 20 25
Expression of bifunctional TreS-Mak of Estrella lausannensis.
time (min) To our knowledge, the characterization of the bifunctional TreS-
Mak activity has not yet been reported in the literature. The his-
tagged TreS-Mak protein purified on nickel columns displays a
molecular weight of 115 kDa on SDS-PAGE (Supplementary
GlgE proteins (E1) of E. lausannensis were loaded onto Fig. 7a) while in solution recombinant TreS-Mak formed a
nondenaturing polyacrylamide electrophoresis (native-PAGE). homodimer with an apparent molecular weight of 256 kDa as
After migration, slices of polyacrylamide gel were incubated analyzed by superose 6 column chromatography (Supplementary
overnight in buffers containing 0 mM (control) or 2 mM M1P Fig. 7b). This contrasts with the hetero-octameric complex
(Fig. 7a). The synthesis of long glucan chains with DP > 15 composed of four subunits of TreS and four subunits of Mak
(minimum number of glucose units required for detection (≈490 kDa) observed in Mycobacterium smegmatis in which
8 COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbioCOMMUNICATIONS BIOLOGY | https://doi.org/10.1038/s42003-021-01794-y ARTICLE Fig. 7 De novo synthesis of branched polysaccharides. a Recombinant GlgE activity of Estrella lausannensis from crude extract of E. coli (CE) and purified on nickel affinity column (E1) were loaded on nondenaturing polyacrylamide gel. After migration, slices of native-PAGE were incubated in TRIS/acetate buffer containing 2 mM of maltose-1-phosphate (M1P) over 16 h at 25 °C. The synthesis of de novo glucan chains is visualized as dark-blue bands due to the formation of glucan–iodine complexes. Similar in vitro experiments were conducted by adding GlgB activity of W. chondrophila to a TRIS/acetate buffer containing GlgE activity and 2 mM of M1P. After overnight incubation, reaction mixture was subjected to 1H-NMR analysis. b Part of 1H-NMR spectra of maltose-1-phosphate (M1P), non-purified reaction mixture, and glycogen (gly) from bovine liver in D2O. Peak #1 (5.45–5.3 ppm) and peak #2 (4.98 ppm) represent the signals of protons involved, respectively, in α-1,4 and α-1,6 linkages while peak #3 (5.47 ppm) represents the characteristic doublet of doublet signals of α-anomeric proton located on C1 of maltose-1-phosphate. The appearances of peak #2 and peak #3 in incubation product indicate the formation of branched polysaccharides composed of α-1,4 and α-1,6 linkages. The presence of peak #3 suggests that M1P was not completely polymerized by GlgE activity of EL. α-Polysaccharides were then purified (see “Methods” for details) and incubated with a commercial isoamylase type debranching enzyme. After overnight incubation, the linear glucan chains released from α-polysaccharides (c) and glycogen from bovine liver used as reference (e) were separated according to the degree of polymerization by capillary electrophoresis coupled with a fluorescent labeling of reduced-ends. As control, α- polysaccharide (d) and glycogen (f) samples were directly labeled and analyzed by capillary electrophoresis in order to estimate the content of free-linear glucan chains. homotetramers of TreS forms a platform to recruit dimers of Mak (0.29 μmol maltose. min−1.mg−1) and dropped by 2.8-fold when via specific interaction domain30,31. the ATP concentration reaches up to 20 mM (0.15 μmol maltose. We first confirmed that the N-terminus TreS domain is min−1.mg−1). Finally, the apparent Km value for trehalose was functional by measuring the interconversion of trehalose into determined at 42.3 ± 2.7 mM in the presence of 1 mM MnCl2 and maltose (see “Methods” for details). Previous reports indicated that 0 mM ATP (Fig. 8c). This is consistent with the apparent Km values TreS activities are partially or completely inhibited with 10 mM of for trehalose (50–100 mM) reported in the literature for TreS divalent cation while a concentration of 1 mM has positive effects. activity in various species32. We further focused on the activity of The effect of Mn2+ cation on the activity of TreS domain was the maltokinase domain that catalyzes the phosphorylation of inferred at 200 mM of trehalose. As depicted in Fig. 8a, the activity maltose in presence of ATP and releases M1P and ADP. The latter of the TreS domain increases only slightly by 1.1-fold from 0 to was monitored enzymatically via the pyruvate kinase assay in order 1 mM of Mn2+ (0.37 μmol maltose. min−1.mg−1) whereas a notice- to express the Mak activity domain as μmol of ADP released.min−1. able decrease of TreS activity (0.24 μmol maltose. min−1.mg−1) is mg−1 of protein. The pH and temperature optima were, obtained at 10 mM of Mn2+. As reported in the literature, the TreS respectively, determined at 42 °C and pH 8 (Supplementary Fig. activity is also associated with the release of glucose during the 7c, d). Interestingly, the activity of the Mak domain is functional interconversion of trehalose into maltose. Because TreS activity is within a wide range of temperature that reflects, probably, the fused with the Mak domain in E. lausannensis, we tested the effect of temperature of free-living amoebae or animal hosts. Kinase activities a wide range of concentration of ATP concentration on the are reported for their requirement in divalent cation in order to interconversion of trehalose (Fig. 8b). Although no revelant effect of stabilize the negatively charged phosphate groups of phosphate ATP was observed on TreS activity at 1 mM (0.43 μmol maltose. donors such as ATP. Therefore, TreS-Mak activity was inferred in min−1.mg−1), TreS activity decreased by 0.6-fold at 3–10 mM ATP the presence of various divalent cations (Fig. 8e). As expected, the COMMUNICATIONS BIOLOGY | (2021)4:296 | https://doi.org/10.1038/s42003-021-01794-y | www.nature.com/commsbio 9
You can also read

















































