Long non-coding RNA INXSis a critical mediator of BCL-XSinduced apoptosis
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Nucleic Acids Research Advance Access published July 3, 2014
Nucleic Acids Research, 2014 1
doi: 10.1093/nar/gku561
Long non-coding RNA INXS is a critical mediator of
BCL-XS induced apoptosis
Carlos DeOcesano-Pereira1 , Murilo S. Amaral1 , Kleber S. Parreira1 , Ana C. Ayupe1 ,
Jacqueline F. Jacysyn2 , Gustavo P. Amarante-Mendes2,3 , Eduardo M. Reis1,4 and
Sergio Verjovski-Almeida1,4,*
1
Departamento de Bioquı́mica, Instituto de Quı́mica, Universidade de São Paulo, 05508-900 São Paulo, SP, Brasil,
2
Departamento de Imunologia, Instituto de Ciências Biomédicas, Universidade de São Paulo, 05508-900 São Paulo,
SP, Brasil, 3 Instituto Nacional de Ciência e Tecnologia de Investigação em Imunologia, Universidade de São Paulo,
05508-900 São Paulo, SP, Brasil and 4 Instituto Nacional de Ciência e Tecnologia em Oncogenômica, Universidade
de São Paulo, 05508-900 São Paulo, SP, Brasil
Received November 11, 2013; Revised June 09, 2014; Accepted June 10, 2014
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
ABSTRACT INTRODUCTION
BCL-X mRNA alternative splicing generates pro- BCL-X is a key apoptotic member of the BCL-2 gene fam-
apoptotic BCL-XS or anti-apoptotic BCL-XL gene ily that modulates tumor cell death and growth (1–3). Al-
products and the mechanism that regulates splice ternative splicing of exon 2 in the BCL-X pre-mRNA pro-
shifting is incompletely understood. We identified duces two isoforms, BCL-XL and BCL-XS, which have
been shown to exert antagonistic functions in the apop-
and characterized a long non-coding RNA (lncRNA)
totic pathway (4), however, the mechanism that regulates
named INXS, transcribed from the opposite genomic the splice shifting between these two isoforms is incom-
strand of BCL-X, that was 5- to 9-fold less abundant in pletely understood. BCL-XL, the anti-apoptotic isoform,
tumor cell lines from kidney, liver, breast and prostate is highly expressed in a number of tumors (5) and elevated
and in kidney tumor tissues compared with non- levels of BCL-XL have frequently been associated with ag-
tumors. INXS is an unspliced 1903 nt-long RNA, is gressive disease and/or chemoresistance (2). In contrast,
transcribed by RNA polymerase II, 5 -capped, nuclear BCL-XS pro-apoptotic isoform abundance is low in tumor
enriched and binds Sam68 splicing-modulator. Three cells, and BCL-XS overexpression can sensitize these cells
apoptosis-inducing agents increased INXS lncRNA to chemotherapeutic agents (6–9), and can induce apopto-
endogenous expression in the 786-O kidney tumor sis in melanoma cells (10,11).
cell line, increased BCL-XS/BCL-XL mRNA ratio and The alternative splicing of BCL-X pre-mRNA has been
activated caspases 3, 7 and 9. These effects were shown to involve proteins, such as splicing-modulators or
components of the exon junction complex (12–17), het-
abrogated in the presence of INXS knockdown. Sim- erogeneous nuclear ribonucleoproteins (18,19) and BCL-
ilarly, ectopic INXS overexpression caused a shift in X pre-mRNA cis-element motifs (20,21). Long non-coding
splicing toward BCL-XS and activation of caspases, RNAs (lncRNAs) are crucial players in cancer (22–25) and,
thus leading to apoptosis. BCL-XS protein accumu- although they have been recognized as regulators of gene
lation was detected upon INXS overexpression. In a expression, mainly by recruiting DNA-binding modulatory
mouse xenograft model, intra-tumor injections of an proteins that act on different genes and pathways (26,27),
INXS-expressing plasmid caused a marked reduction the possible regulation of BCL-X splicing by lncRNAs has
in tumor weight, and an increase in BCL-XS isoform, not been investigated.
as determined in the excised tumors. We revealed A number of potentially oncogenic or anti-apoptotic
an endogenous lncRNA that induces apoptosis, sug- lncRNAs, such as PCGEM1 and PANDA, have been iden-
gesting that INXS is a possible target to be explored tified (24,25), yet none of them act on the anti-apoptotic
BCL-2 gene family. Similarly, although four lncRNAs with
in cancer therapies. tumor-suppressive or pro-apoptotic activities have been
well characterized, namely, lincRNA-p21, GAS5, ncRNA
CCND1 and MEG3 (28–31), none of them directly act
on the BCL-2 apoptotic pathway. Regarding gene splicing
* To whom correspondence should be addressed. Tel: +55 11 3091 2173; Fax: +55 11 3091 2186; Email: verjo@iq.usp.br
C The Author(s) 2014. Published by Oxford University Press on behalf of Nucleic Acids Research.
This is an Open Access article distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by-nc/3.0/), which
permits non-commercial re-use, distribution, and reproduction in any medium, provided the original work is properly cited. For commercial re-use, please contact
journals.permissions@oup.com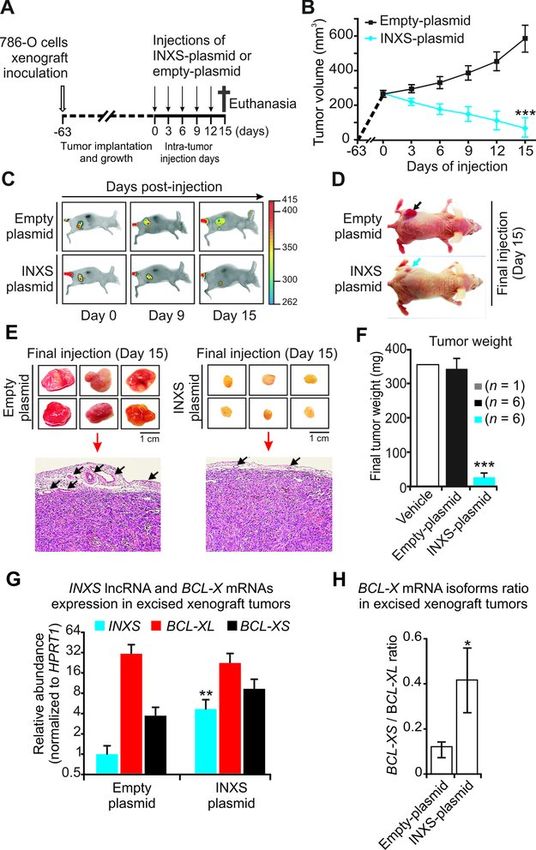
2 Nucleic Acids Research, 2014
modulation, it has been long known to involve the small
nuclear RNAs (snRNAs) (32), which along with the SR
proteins and hnRNPs are components of the spliceosome.
While a computational analysis has revealed an extensive
relationship between long antisense RNAs and alternative
splicing in the human genome (33), only a very limited num-
ber of lncRNAs has been shown to directly modulate al-
ternative mRNA splicing. Thus, an endogenous transcript
antisense to N-myc was shown to form an RNA-RNA du-
plex with the sense mRNA and to cause retention of N-myc
intron 1 (34), however, the functional significance of this al-
ternative splicing was not assessed (34). Related to apopto-
sis, splicing of the Fas protein-coding gene is influenced by
the antisense Saf transcript (35), making the cells highly re-
sistant to Fas-mediated apoptosis, which characterizes Saf
as an anti-apoptotic lncRNA (35) with oncogenic activity.
In a similar manner, a natural antisense transcript regulates
the splicing of Zeb2, and is related to an increase in the pro-
duction of oncogenic Zeb2 protein in human tumors (36).
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
MALAT1 is the only example of an oncogenic lncRNA
(37,38) that indirectly acts on alternative splicing through
the modulation of the SR protein phosphorylation (39).
Here, we show a novel endogenous lncRNA that favors
the accumulation of the pro-apoptotic BCL-XS isoform,
thus having a tumor suppressor activity. This lncRNA is
transcribed from the BCL-X genomic locus in the antisense
direction relative to BCL-X mRNA, and we named it INXS
for intronic BCL-XS-inducing lncRNA. Our results sup-
port a view of INXS as a critical mediator of apoptosis that
integrates damaging environmental conditions with the cel-
lular events that lead to cell death.
MATERIALS AND METHODS
Detailed experimental procedures are available as Supple-
mentary Data.
RNA extraction and strand-specific reverse transcription-
polymerase chain reaction (RT–PCR) Figure 1. Identification and characterization of INXS as an intronic an-
Total RNA was isolated from cultured cells with Tri- tisense lncRNA downregulated in tumor cell lines. (A) Structure of the
BCL-X gene locus on chromosome 20 with the BLC-X protein-coding
zol (Invitrogen) and purified with an RNAspin Mini kit mRNA being transcribed from the minus genomic strand and the anti-
(GE Healthcare) according to the manufacturer’s instruc- sense unspliced INXS lncRNA transcribed from the intronic region on the
tions, with an extended treatment with DNase I for 1 opposite strand. Gray boxes in INXS indicate transcript portions extended
h. Total RNA of excised xenograft tumors was extracted by RACE-PCR and sequencing. Small blue arrows next to INXS indicate
PCR primers positions. (B) Antisense transcription [AS] of INXS lncRNA
from paraffin-embedded tumor samples with the miRNeasy was detected by strand-specific RT-PCR in 786-O, DU145 and HepG2 cell
kit for formaline-fixed, paraffin-embedded (FFPE) tissues lines. Sense transcription [S] was not detected in this locus region. C1 and
(217504, Qiagen) according to the manufacturer’s instruc- C2 are negative controls. (C) INXS expression levels across a panel of tu-
tions. Total RNA was quantified on ND-1000 (NanoDrop) mor (light blue) and non-tumor (dark blue) cell lines from kidney, liver,
and its integrity was assessed on a Bioanalyzer (Agilent). breast and prostate. Absolute quantification of INXS was obtained for
each cell line and the relative abundance is shown with respect to the ab-
For measuring INXS in the strand-specific assays (Fig- solute amount measured in the CaKi-1 cell line (175 molecules per ng of
ure 1B and C), reverse transcription (RT) was performed total RNA). (D) BCL-XS (black) and BCL-XL (red) mRNA isoform lev-
with SuperScript III (Invitrogen) using 3 g of total RNA els across a panel of tumor (T) and non-tumor (NT) cell lines from kidney,
and a gene-specific primer listed in the Supporting Infor- liver, breast and prostate. Expression levels are relative to the expression
of the BCL-XS isoform in the CaKi-1 cell line. The data are the mean ±
mation. Controls for RT without primer (-primer) or with- SD of three independent experiments. *(P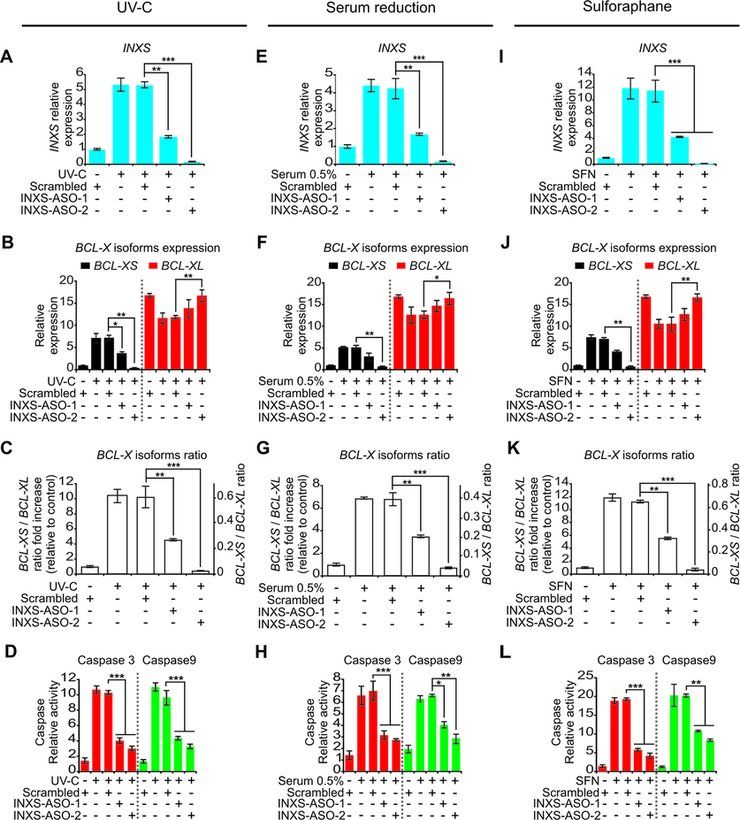
Nucleic Acids Research, 2014 3
RT-qPCR of BCL-X (Figure 1A). The antisense directionality of this
transcript relative to mRNAs encoded in the BCL-X locus
For measuring INXS lncRNA and protein-coding mRNAs,
was determined by strand-oriented RT-PCR in three hu-
reverse transcription was performed with the SuperScript
man cell lines (Figure 1B), namely, 786-O kidney tumor,
III (Invitrogen) using 1 g of total RNA and oligo-dT
DU145 prostate tumor and HepG2 liver tumor cell lines,
primer, followed by qPCR as described in the Supplemen-
and the identity of the PCR product was confirmed by se-
tary Data.
quencing. It is worth noting that no transcript was observed
in the sense controls (Figure 1B), indicating that no DNA
RACE-PCR contaminant was present, and also indicating that the levels
of BCL-X pre-mRNA that accumulate in these cell lines are
For RACE-PCR we used the Human Fetal Kidney
undetectable. An in silico analysis with the CPC Coding Po-
Marathon-Ready cDNA (Clontech, No. 639323) commer-
tential Calculator tool (41) showed no coding potential for
cial library prepared from poly(A) RNA extracted from
the transcript, and we named it INXS, for intronic BCL-
a pool of 59 spontaneously aborted fetal kidneys. Gene-
XS-inducing lncRNA.
specific primers are listed in the Supplementary Data.
Transient plasmid or oligonucleotide transfections
INXS is less abundant in a set of different tumor cell lines and
Cells were transfected with pCEP4-INXS plasmid using in kidney tumor tissues compared with non-tumors
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
FuGENE HD (Promega) for the INXS overexpression as-
Tumor cell lines from kidney, liver, breast and prostate
says. For INXS knockdown assays, cells were transfected
showed an endogenous INXS expression that was 5- to 9-
with modified antisense oligonucleotides (ASOs) using lipo-
fold lower compared with non-tumor cell lines derived from
fectamine RNAimax (Invitrogen). See details of ASOs titra-
the same tissues (Figure 1C). In parallel, we measured the
tions under Supplementary Data.
relative abundance of BCL-X protein-coding mRNA in all
studied cell lines, and found that the BCL-X expression
Apoptosis assay and fluorimetric determination of caspase had the opposite pattern, being higher in the tumor than
activity in the non-tumor cell lines studied (Figure 1D). More im-
portantly, the BCL-XS pro-apoptotic isoform was relatively
Cell viability was determined by staining with Annexin V
lower in tumor than in non-tumor cell lines derived from all
FITC and propidium iodide (Invitrogen) by flow cytometry
four different tissues that we have studied (Figure 1D, black
(Beckman Coulter FC500 MPL). Caspase activities were
bars).
measured with specific fluorigenic substrates and the respec-
Because the INXS tumor/non-tumor abundance ratio in
tive caspase inhibitors (Kamiya Biomedical); see details un-
kidney cell lines was the lowest among the cell lines of dif-
der Supplementary Data.
ferent origins that were tested, we chose the kidney tumor
cell line 786-O as a model for the characterization of the in-
Nude mouse xenograft assays duction of endogenous INXS expression, as described fur-
ther below. In this cell line the absolute level of INXS cor-
Generation of human kidney tumor 786-O cell mouse
responds to ∼5 copies per cell, whereas BCL-XS is 2-fold
xenografts and intra-tumor injections of pCEP4-empty
higher (12 copies per cell), and BCL-XL is 35-fold higher
or pCEP4-INXS plasmids dissolved in TurboFect in vivo
than INXS (167 copies per cell).
transfection solution (Fermentas) are described in detail
Next, we analyzed INXS endogenous expression in the
under Supplementary Data.
post-nephrectomy kidney samples of 13 patients with re-
nal cell carcinoma, and observed quite variable levels of
RESULTS INXS (Supplementary Figure S1A). Interestingly, in all pa-
tient samples, when the INXS level of the non-tumor tissue
INXS is a long non-coding antisense transcript
was set to 1, we found that the INXS levels were 2- to 60-
To identify lncRNAs with a possible involvement in the reg- fold lower in the tumor tissue compared with the matched
ulation of cell apoptosis, we searched the public databases non-tumor (Supplementary Figure S1B). Again, BCL-X
for ESTs and mRNAs that mapped to gene loci encoding protein-coding mRNA relative abundance had the opposite
proteins from the BCL-2 family. A detailed analysis of the pattern, being significantly higher in the tumor than in the
BCL-X genomic locus on human chromosome 20 revealed paired non-tumor (Supplementary Figure S1C), and BCL-
at least 70 unspliced ESTs covering 1291 bp in that locus XS pro-apoptotic isoform was similarly lower in tumor tis-
(Figure 1A, thin black box on upper strand), which over- sues compared with matched non-tumor tissues (Supple-
lapped the 5 -UTR portion of BCL-X. We performed 5 - mentary Figure S1C, black bars) in all 13 kidney cancer pa-
and 3 -RACE assays using a human fetal kidney cDNA li- tient samples.
brary, sequenced the products and detected an unspliced Thus, there is a concomitant lower expression of INXS
1903 nt-long transcript, which extended the ESTs evidence lncRNA and of BCL-XS pro-apoptotic mRNA isoform
by 537 nt and 75 nt at the 5 - and 3 -ends, respectively (Fig- both in the tumor cell lines that have been studied and in the
ure 1A, thin gray boxes on the upper strand). The full- kidney patient tumor samples that have been analyzed com-
length transcript spanned beyond exons 1 and 2, overlap- pared with non-tumor, suggesting that INXS might play a
ping intron 1 and some of the genomic regions upstream role in BCL-X splicing regulation.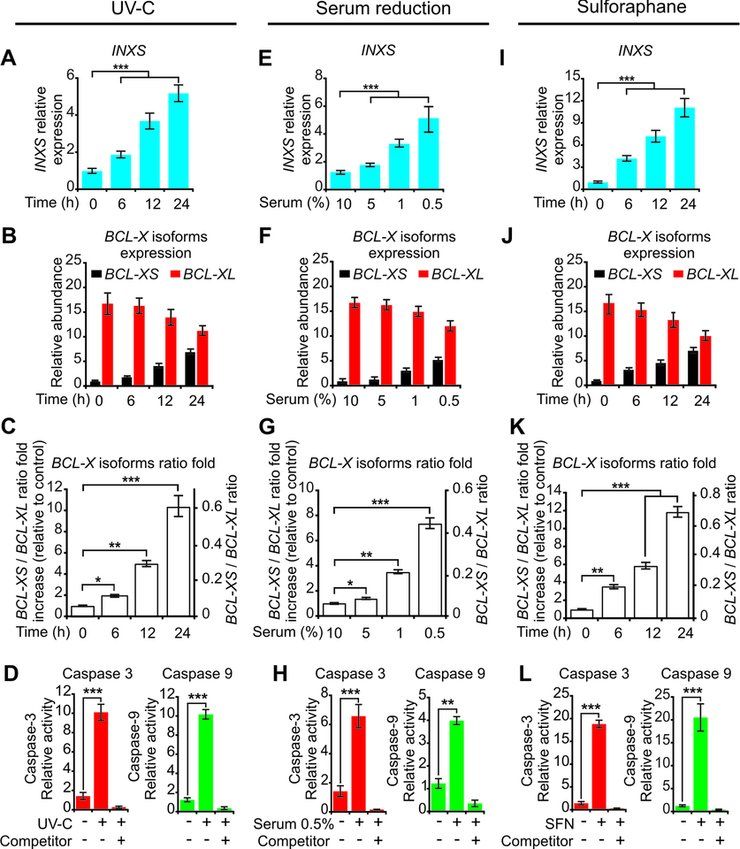
4 Nucleic Acids Research, 2014
Figure 2. INXS knockdown in the non-tumor kidney cell line RC-124 re-
duces the BCL-XS/BCL-XL ratio. (A) INXS knockdown in the RC-124
cell line using two distinct INXS-knockdown ASOs (INXS-ASO-1 and -
2) reduces by 60% the endogenous levels of INXS compared to a control
scrambled oligonucleotide transfection. (B) The levels of BCL-XS (black)
and BCL-XL (red) mRNA isoforms in the same cells were measured by
RT-qPCR. (C) BCL-XS/BCL-XL mRNA ratio in these cells was signifi-
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
cantly reduced from 0.25 to 0.15 upon transfection with INXS-ASO-1 and
-2. The data are the mean ± SD of three independent experiments. *(P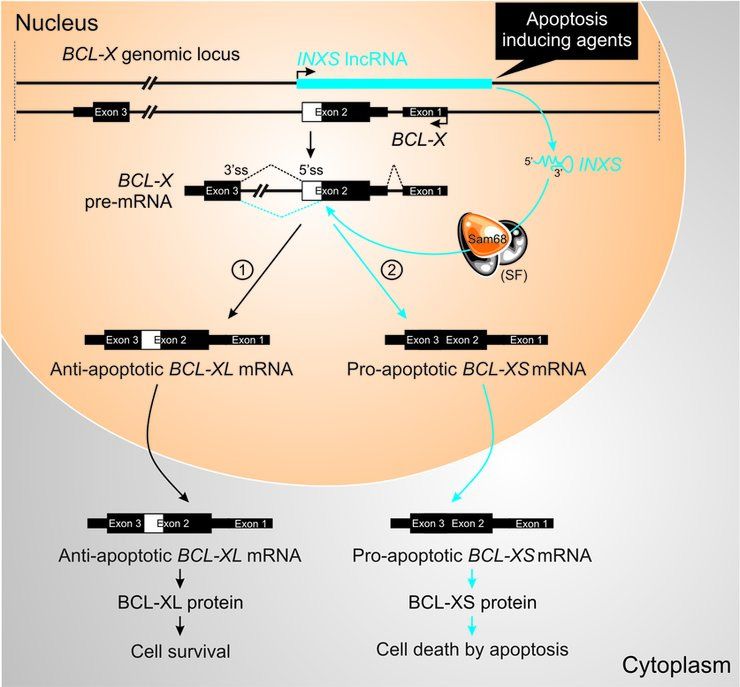
Nucleic Acids Research, 2014 5
from the extrinsic apoptotic pathway was not activated by
UV-C exposure (Supplementary Figure S4A).
A similar pattern of increased INXS expression was ob-
tained 24 h after reducing the serum in the culture medium
in a dose-dependent manner from 10% to 5%, 1% and 0.5%
(Figure 4E), which resulted in similar BCL-X isoforms shift
patterns, and in the activation of caspases (Figure 4F–H,
Supplementary Figures S3B and S4B).
Treatment with the anti-cancer agent sulforaphane (SFN)
induces cell-cycle arrest and apoptosis and causes a reduc-
tion in anti-apoptotic BCL-XL expression (44). After 24 h
exposure of 786-O cells to SFN (50 M), there was a 10-fold
increase in INXS expression (Figure 4I), which was again
accompanied by similar changes in BCL-X isoforms and
in the activation of caspases (Figure 4J–L, Supplementary
Figures S3C and S4C).
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
INXS is required for caspase activation caused by apoptosis-
inducing agents
The transfection of 786-O cells with two different ASOs
complementary to INXS lncRNA significantly abrogated
the increase in INXS levels caused by the three apoptosis-
inducing agents, namely, UV-C exposure (Figure 5A),
serum reduction to 0.5% (Figure 5E) or SFN treatment
(Figure 5I). Under INXS knockdown, the increase in the
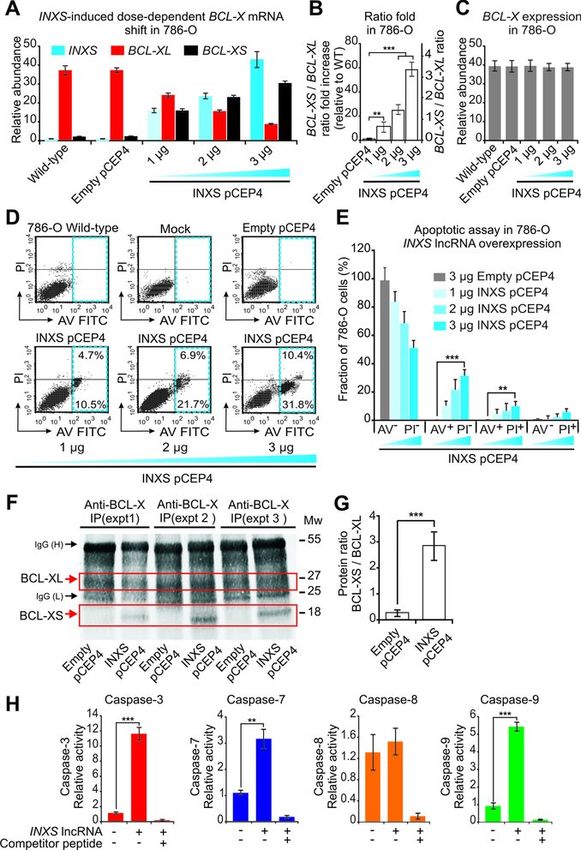
Figure 4. INXS endogenous expression is increased by apoptosis-inducing pro-apoptotic BCL-XS isoform was equally abrogated un-
agents and is associated with increased BCL-XS mRNA and caspases acti- der these three cell treatments (Figure 5B, F and J, black
vation. (A, E, I) Induced expression of INXS lncRNA in 786-O kidney tu-
mor cells by exposure (A) to 40 J/m2 UV-C light for 40 s at time zero, (E) to
bars), and the shift in the BCL-XS/BCL-XL ratio toward
serum reduction for 24 h or (I) to sulforaphane anti-cancer drug (SFN) for the pro-apoptotic BCL-XS isoform was significantly re-
up to 24 h. (B, F, J) Levels of BCL-XS (black) and BCL-XL (red) mRNA duced in all three conditions (Figure 5C, G and K). As a
isoforms in the same cells. (C, G, K) BCL-XS/BCL-XL mRNA ratio in consequence of INXS knockdown, the activation of both
the cells. (D, H, L) Activation of caspase 3 and caspase 9 in 786-O cells effector caspase 3 (Figure 5D, H and L, red bars) and ini-
at 24 h after the exposure to (D) 40 J/m2 UV-C, (H) 0.5% serum or (L)
SFN. (See Supplementary Figure S4 for other caspases.) *(P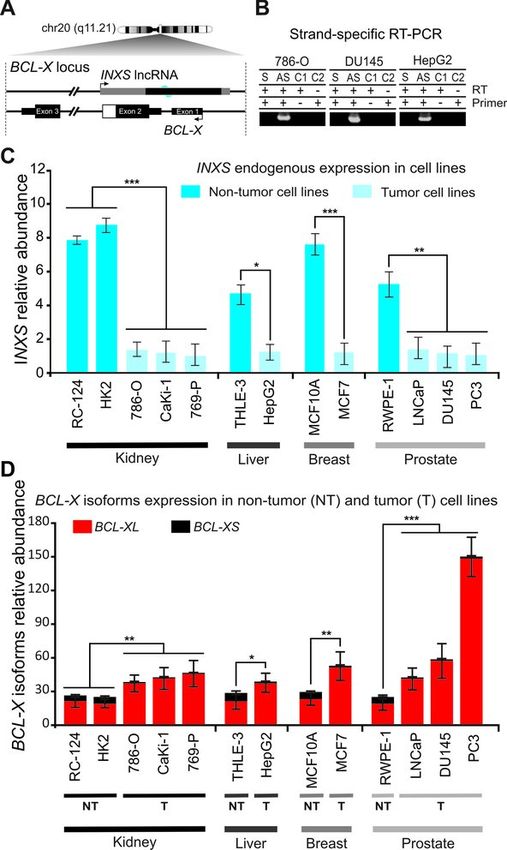
6 Nucleic Acids Research, 2014
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
Figure 5. INXS knockdown abrogates the effect of apoptosis-inducing
agents. (A, E, I) Abrogated induction of INXS expression in the presence
of two distinct INXS-knockdown ASOs (INXS-ASO-1 or -2), with cells
exposed (A) to 40 J/m2 UV-C light for 40 s at time zero and incubated for
24 h, (E) to serum reduction (0.5 % serum) for 24 h or (I) to sulforaphane
anti-cancer drug (SFN) for 24 h. (B, F, J) The increase in BCL- XS (black)
and the decrease in BCL-XL (red) mRNA isoforms induced by the apop-
totic agents are abrogated in the presence of two distinct INXS-knockdown
ASOs (INXS-ASO-1 or -2). (C, G, K) The increase in BCL-XS/BCL-XL
mRNA ratio is significantly abrogated in the INXS-knockdown treated
cells. (D, H, L) Abrogated activation of caspase 3 and caspase 9 in the Figure 6. Overexpression of INXS induces BCL-XS mRNA and protein
presence of two distinct INXS-knockdown ASOs (INXS-ASO-1 or -2). and promotes apoptosis. (A) The levels of BCL-XS (black) and BCL-XL
(See Supplementary Figure S4 for other caspases.) In all panels, the data (red) mRNA isoforms were measured by RT-qPCR in 786-O kidney tumor
are the mean ± SD of three independent experiments. *(P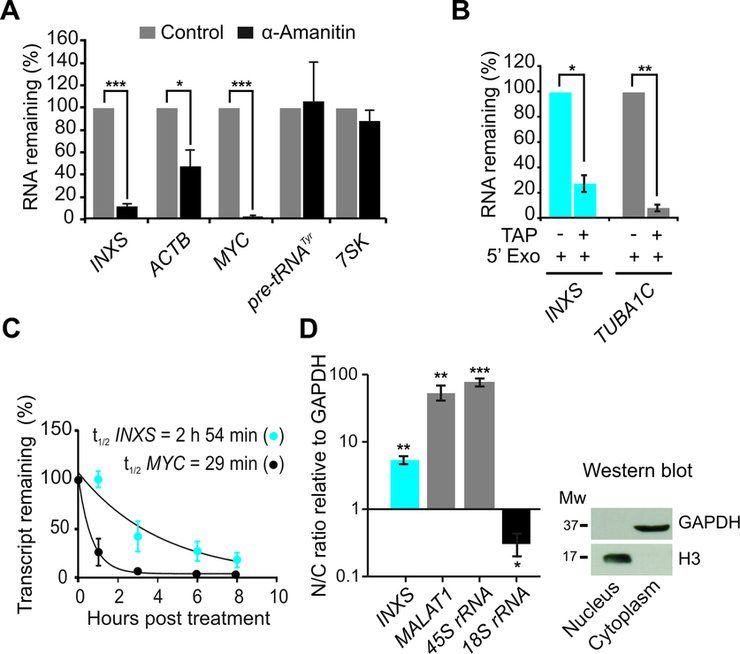
Nucleic Acids Research, 2014 7
expression; no change in the total BCL-X mRNA (Figure a 22-fold increase in INXS lncRNA compared with the
6C) was observed. At the highest INXS lncRNA levels, the endogenous (Supplementary Figure S8A), and an 18-fold
pro-apoptotic BCL-XS isoform was clearly predominant increase in BCL-XS pro-apoptotic mRNA isoform (Sup-
(∼80% of all BCL-X mRNA in the cell). plementary Figure S8B), with a concomitant 2.5-fold re-
The efficiency of INXS lncRNA ectopic overexpression duction in BCL-XL mRNA (Supplementary Figure S8C).
in affecting BCL-X mRNA alternative splicing was tested At 15 h after transfection a significant 18-fold increase in
in two additional tumor cell lines, namely, the breast tu- the BCL-XS/BCL-XL ratio from 0.06 to 1.2 was obtained
mor MCF7 cell line (Supplementary Figure S6A–C) and (Supplementary Figure S8D).
the prostate tumor PC3 cell line (Supplementary Figure Thus, transfected cells collected at 15 h were lysed, BCL-
S6F–H), showing similar patterns of dose-dependent BCL- X protein isoforms were immunoprecipitated with anti-
XS mRNA increase upon ectopic overexpression of INXS BCL-X antibody and the IP fraction was analyzed by west-
lncRNA, with a concomitant BCL-XL decrease and a sus- ern blot. A distinct BCL-XS band was detected only in
tained level of total BCL-X. the INXS-overexpressing cells but not in the control (Fig-
Having determined that INXS overexpression alters the ure 6F). The densitometric intensity ratio between BCL-
BCL-X splicing pattern in 786-O, MCF7 and PC3 cell lines, XS and BCL-XL signals significantly increased from 0.2
we next examined its effect on cell apoptosis. Cells overex- in the control cells (where the background intensity sig-
pressing INXS lncRNA showed a dose-dependent increase nal was used as a proxy for BCL-XS abundance) to 2.8 in
in apoptosis; the population of apoptotic 786-O kidney the INXS-overexpressing cells (Figure 6G). The amount of
cells labeled with Annexin V FITC increased from 15% in BCL-XL isoform apparently did not decrease after 15 h of
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
786-O cells transfected with 1 g of pCEP4-INXS plas- INXS overexpression (Figure 6F), suggesting that the BCL-
mid (Figure 6D, left bottom panel) to ∼42% in cells trans- XL protein turnover is longer than the BCL-XL mRNA
fected with 3 g of plasmid (Figure 6D, right bottom panel). turnover (Supplementary Figure S8C). It is noteworthy that
The apoptosis-inducing effect of INXS overexpression was BCL-XS protein abundance in non-apoptotic human cell
very reproducible among three biological replicates (Figure lines is relatively low, and was shown to be below western
6E), and the average fraction of apoptotic cells (AV+ PI− blot detection limits in melanoma (10,46) or lung adeno-
plus AV+ PI+ ) in the population significantly increased from carcinoma (8) cell lines. The difficulties in detection of the
∼15% in 786-O cells transfected with 1 g of plasmid to BCL-XS protein isoform was also observed here in 786-O
∼40% in cells transfected with 3 g of INXS-expressing kidney tumor cells, which led us to use the IP approach.
plasmid (Figure 6E). Apoptosis induced by INXS lncRNA The overexpression of INXS lncRNA caused a significant
overexpression as detected by Annexin V-FITC labeling activation of caspases 3, 7 and 9, the major caspases in the
was reproduced in the two other tumor cell lines tested, mitochondrially mediated apoptosis pathway, but had no
namely, the MCF7 breast tumor (Supplementary Figure effect on initiator caspase 8 of the death receptor extrinsic
S6D and E) and PC3 prostate tumor (Supplementary Fig- pathway (Figure 6H).
ure S6I and J). The apoptotic phenotype was further confirmed by im-
The above-described phenotype was not observed when munofluorescence imaging with an anti-active caspase 3 an-
we overexpressed ANRASSF1, a similar but unrelated un- tibody, which showed a stronger signal in the 786-O cell
spliced lncRNA that is endogenously transcribed antisense line transfected with pCEP4-INXS vector when compared
to the RASSF1A gene in prostate cancer cells (45). Upon with the control (Supplementary Figure S9). These results
overexpression of ANRASSF1 in 786-O kidney tumor cells, are entirely in agreement with the role of BCL-XS protein
no change in BCL-XS or BCL-XL isoform levels was de- in activating the mitochondrially mediated apoptosis path-
tected (Supplementary Figure S7A and B). In addition, no way (10).
apoptosis was observed in ANRASSF1-overexpressing cells
using the Annexin V-PI assay (Supplementary Figure S7C),
providing further support that the apoptotic phenotype was
INXS interacts with Sam68 splicing-modulator complex
the result of a specific effect elicited by INXS lncRNA over-
expression. Because the Sam68 splicing-modulator complex regulates
the alternative splicing of BCL-X pre-mRNA (12), we per-
formed a native Ribonucleoprotein immunoprecipitation
Overexpression of INXS increases BCL-XS protein and ac-
(RIP) assay with an anti-Sam68 antibody to determine if
tivates caspases 3, 7 and 9
INXS lncRNA binds to Sam68-containing ribonucleopro-
The effect of INXS lncRNA overexpression on BCL-X tein (RNP) complexes. The immunoprecipitated proteins
mRNA isoforms was tested at the protein level. Because were digested, and the protein-bound RNAs were measured
too many dying cells were accumulated 24 h after trans- with RT-PCR. In this assay, the INXS lncRNA was de-
fection with the INXS-expressing plasmid (3 g plasmid), tected as bound to Sam68-containing RNP complexes (Fig-
which reduced our ability to collect viable cells for the west- ure 7A). As positive controls, BCL-XL and BCL-XS mR-
ern blot assays, we first determined the shortest time of NAs were also detected as bound to the Sam68-containing
overexpression at which to perform the western blots. By RNP complex (Figure 7A), in agreement with previous re-
measuring the time course of INXS lncRNA and BCL- ports (12). The negative control BCL-2 mRNA (12), which
XS mRNA increase following the transient transfection of is known to not bind to Sam68, was not detected in the im-
786-O cells (Supplementary Figure S8A and B), we found munoprecipitate (Figure 7A). As expected, Sam68 was de-
that as early as 15 h after transfection there was already tected in the immunoprecipitate fraction of the anti-Sam68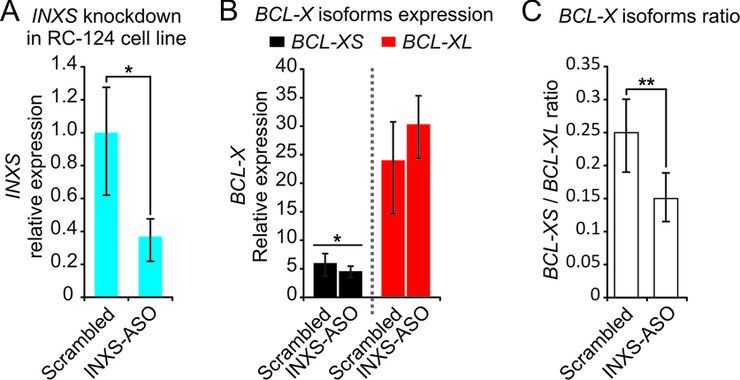
8 Nucleic Acids Research, 2014
RIP assay, by a western blot with the same anti-Sam68 an-
tibody (Figure 7A).
Overexpression or knockdown of INXS does not affect the
splicing of other Sam68 target mRNAs
We monitored the effects of INXS overexpression on splic-
ing of two additional Sam68 target mRNAs that have been
identified in the literature, namely, the proto-oncogenes cy-
clin D1(CCDN1-v1) (47) and SF2/ASF (SRSF1-v1 and -
v2) (48). As shown in Figure 7B, overexpression of INXS
did not affect the splicing of either cyclin D1(CCDN1-v1)
or SF2/ASF (SRSF1-v1 and -v2). We also monitored the
effect of INXS knockdown on these Sam68 mRNA tar-
gets and observed no effect on splicing of these genes (Fig-
ure 7C).
INXS induces tumor regression in vivo
To determine the potential of the INXS lncRNA to affect
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
tumor growth in vivo, ectopic overexpression of INXS was
tested in nude mice 786-O kidney tumor cells xenograft as-
says. Tumor cells were inoculated subcutaneously into 12
female athymic nude mice on day –63 (Figure 8A), and
when the tumors reached approximately the same volume
of 250 mm3 (day zero) the animals were randomly sepa-
rated into two groups that were injected either with INXS
or empty pCEP4 vectors. Intra-tumor injections of the
plasmid-containing transfectant solution started at day zero
and proceeded at every third day for 15 days (Figure 8A).
The tumor volume for each animal was measured with the
caliper at each injection day (Figure 8B). At day 15, in all
animals injected with INXS lncRNA plasmid the average
tumor volume was reduced to 70 mm3 (Figure 8B), an av-
erage 8-fold tumor regression as compared to the tumors of
animals injected with empty vector (570 mm3 ) (Figure 8B).
In parallel, we monitored tumor size through near-
infrared optical imaging. Animals were injected in the tail
vein with a fluorescently labeled Epidermal Growth Fac-
tor (EGF) marker and scanned at the beginning (day 0), in
the middle (day 9) and at the end of the injection period
(day 15). Images of two representative animals are shown
(Figure 8C). A considerable reduction in tumor volume was
observed in the animals from the group that received the
pCEP4-INXS vector (Figure 8C, lower panels), while tu-
mor growth over time was evident in the control group in-
oculated with empty vector (Figure 8C, upper panels). On
day 15, the tumor size difference could be clearly visualized
(Figure 8D). All mice were euthanized on day 15; the tu-
mors were removed and photographed (Figure 8E). The tu-
mors were formalin-fixed, paraffin-embedded and observed
by light microscopy with hematoxylin and eosin stain (Fig-
Figure 7. INXS interacts with the Sam68 splicing-modulator complex. (A) ure 8E). In the INXS-injected tumors the vascularization
Native RIP (RNA-binding protein immunoprecipitation) assay with anti-
Sam68 antibody was performed, followed by RT-PCR with primers for the was considerably reduced as compared with controls (Fig-
indicated genes. A negative control, from RNA-IP with immunoglobulin ure 8E).
G (IgG) was included. For INXS lncRNA, a strand-specific primer was The excised tumors were weighed and a marked 13-fold
used for RT. For the positive controls (BCL-XL and BCL-XS mRNAs) reduction in the mean tumor weight was observed in the
and the negative control (BCL-2 mRNA), oligo-dT primer was used for
RT. The protein fraction from the native RIP assay with anti-Sam68 an-
INXS-injected tumors that remained (27 mg) compared
tibody was analyzed by western blot, which was developed with the same with the empty-plasmid-injected tumors from the controls
antibody. A negative control sample, from RIP with IgG, was included. (339 mg) (Figure 8F). An additional animal that had a tu-
(B) INXS overexpression was performed in the 786-O kidney tumor cell mor injected with only transfection solution (vehicle) had a
line with 3 g of INXS-expressing plasmid for 24 h and the alternative final tumor weight of 351 mg (Figure 8F).
splicing isoforms of two Sam68 target mRNAs that have been identified in
the literature, namely, CCDN1-v1 and SRSF1-v1 and -v2, were measured
by RT-qPCR. (C) INXS knockdown was performed in the RC-124 kid-
ney non-tumor cell line as in Figure 2, and the levels of CCDN1-v1 and
SRSF1-v1 and -v2 were measured by RT-qPCR. The data are the mean ±
SD of three independent experiments.Nucleic Acids Research, 2014 9
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
Figure 8. Overexpression of INXS induces tumor regression in vivo. (A) Schematic representation showing the subcutaneous inoculation of mice with 786-
O human kidney tumor cells (open arrow, on day –63), followed by a waiting period until all the tumors had implanted and grown to reach the approximate
same volume of 250 mm3 (on day zero), when the injection of animals began (vertical arrows). Intra-tumor injections of INXS-plasmid or empty-plasmid
were performed every third day over a period of 15 days. (B) Tumor volume was monitored by caliper on the days of injection in six different animals from
each of two groups, which were injected either with INXS-plasmid (blue) or control empty-plasmid (black). The data are the mean ± SD of measurements
from the six animals of each injection group. (C) An EGF-labeled dye marker was detected in the tumor by in vivo scanning with a near-infrared optical
imaging system. Only one representative mouse is shown for each group, namely, INXS-plasmid or control empty-plasmid. (D) Pictures of the scanned
animals, taken on day 15; note the difference in size between the tumors of the two animals, as indicated by arrows. (E) On day 15, all six animals from
each of the two groups were euthanized, and their tumors were excised and photographed. Subsequently, tumors were formalin-fixed, paraffin-embedded,
stained and observed by light microscopy. Arrows point to tumor peripheral vascularization. (F) Average weights of the excised tumors for each of the
two injection groups, INXS-plasmid (blue) or control empty-plasmid (black). The data are the mean ± SD of measurements from the six animals of each
injection group. An additional animal whose tumor was injected with only transfection solution is shown (vehicle, white bar). (G) INXS lncRNA (blue),
BCL-XL mRNA (red) and BCL-XS mRNA (black) expression levels measured by RT-qPCR in the excised xenograft tumors. (H) BCL-X mRNA isoforms
ratio in the excised xenograft tumors. *(P10 Nucleic Acids Research, 2014
Total RNA was extracted from the excised paraffin-
embedded tumor samples, and INXS lncRNA and BCL-
X mRNA isoforms were measured. A significant 5-fold
higher level of INXS lncRNA was detected in the remaining
tumor cells from the INXS-plasmid-injected tumors com-
pared with the tumor cells from the empty-plasmid-injected
controls (Figure 8G). In these INXS-injected excised tu-
mors the BCL-XS mRNA was increased and the BCL-
XL mRNA was reduced as compared with controls (Fig-
ure 8G). It is worth noting that even though the measure-
ments were carried out with cells that did not die along the
15 days of INXS-plasmid injections, the BCL-XS/BCL-XL
ratio was significantly increased 4-fold in these surviving tu-
mor cells (Figure 8H). Those cells that had died along the 15
days of INXS-plasmid injections, causing the dramatic re-
duction in tumor size, were the ones likely expressing high
levels of INXS. Taken together, these results show that the
INXS lncRNA can reduce tumor size in vivo, thus having a
tumor suppressor activity.
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
DISCUSSION
We identified a novel endogenous lncRNA with an apop- Figure 9. Proposed model of action of INXS lncRNA. BCL-X pre-mRNA
totic role, acting post-transcriptionally in trans at the level of undergoes either a BCL-XL anti-apoptotic or BCL-XS pro-apoptotic al-
ternative processing (4). We propose that the control of BCL-X pre-mRNA
BCL-X pre-mRNA splicing. INXS lncRNA is transcribed alternative splicing, between pathway [1] toward BCL-XL anti-apoptotic
from the same locus as BCL-X, and its expression levels isoform and pathway [2] toward BCL-XS pro-apoptotic isoform, depends
were lower in all tested cancer cell lines, compared with im- on INXS endogenous lncRNA. Apoptosis inducing agents, such as UV-C
mortalized non-tumor cell lines from the same tissues of light exposure, serum starvation or anti-cancer drugs, lead to an increased
origin and in kidney tumor tissue compared with adjacent expression of endogenous INXS. Sam68 increases the level of the pro-
apoptotic BCL-XS isoform, while its absence leads to the accumulation
non-tumor from clear cell renal cell carcinoma patient sam- of BCL-XL (12). We propose that the augmented levels of INXS would
ples. This suggests that down-regulation of INXS lncRNA favor the positioning of Sam68 splicing-modulator and of possible addi-
might be one of the factors conferring tumor resistance to tional splice factors (SF) of the splicing machinery near the distal donor
apoptosis. 5 splice site on the pre-mRNA (5 ss, dotted blue line) and favor the splic-
ing predominantly through pathway 2, thus leading to BCL-XS protein
INXS is one of thousands of unspliced lncRNAs that are synthesis and to apoptosis.
transcribed from intronic regions of 74% of the protein-
coding genes (49), making them a part of the widespread
non-coding transcription that is estimated to arise from at
least 75% to 90% of the human genome (50,51). INXS- pose the model mechanism shown in the scheme of Figure 9,
mediated apoptosis, induced by BCL-XS, exemplifies a in which we postulate that INXS endogenous expression is
novel functional role exerted by a member of the class of required to shift BCL-X pre-mRNA splicing toward BCL-
unspliced intronic antisense lncRNAs; this class has been XS pro-apoptotic isoform and can be induced by apoptotic
so far implicated in gene regulation at the transcriptional agents. When INXS is absent or expressed at low levels, such
level through the recruitment of polycomb repressor com- as in cancer tissues, the predominant output of BCL-X pre-
plex 2 (PRC2) by intronic antisense Kcnq1ot1 lncRNA to mRNA splicing is the anti-apoptotic BCL-XL isoform (Fig-
the Kcnq1 locus to regulate 10 genes at this imprinted gene ure 9, pathway 1). In contrast, augmented levels of INXS
cluster (52), and the recruitment of PRC2 by intronic an- (Figure 9, pathway 2) would interfere with the positioning
tisense ANRASSF1 lncRNA to cause the location-specific and/or accessibility of splice factors to the alternative dis-
cis-acting regulation of the tumor suppressor RASSF1A tal 5 donor splice site on intron 2 of the pre-mRNA and
gene (45). Acting in trans at the post-transcriptional level, favor the positioning of the splicing machinery to increase
the unspliced intronic antisense lncRNA Saf was shown to the abundance of the BCL-XS isoform, leading to an in-
have an anti-apoptotic oncogenic function by modifying the crease in BCL-XS protein and to cell apoptosis. Sam68 is
splicing of the Fas gene (35). It is conceivable that the char- not required for the accumulation of BCL-XL isoform as
acterization of additional members of the class of unspliced determined by Paronetto et al. (12), and a recent paper by
intronic antisense transcripts will lead to novel functional Bielli et al. (53) shows that the transcription factor FBI-1
roles of lncRNAs acting at the post-transcriptional level. reduces Sam68 binding to BCL-X mRNA and that the ab-
Our results showed that increased levels of the unspliced sence of Sam68 favors the anti-apoptotic BCL-XL isoform.
antisense lncRNA INXS induces apoptosis by shifting BCL-X alternative splicing has emerged as a target for
BCL-X pre-mRNA splicing toward the pro-apoptotic BCL- molecular therapy in cancer treatment (4,6,8,46), follow-
XS isoform, leading to accumulation of BCL-XS protein. ing the early demonstration by Boise et al. (4) that BCL-
We also obtained evidence that INXS interacts with Sam68- X long and short isoforms exhibit opposing anti-apoptotic
containing splicing-modulator complexes. Thus, we pro- or pro-apoptotic functions, and act as dominant regulatorsNucleic Acids Research, 2014 11
of apoptotic cell death. The in vivo therapeutic potential of ACKNOWLEDGEMENTS
splice-switching oligonucleotides has been consistently ex-
We thank Ricardo J. Giordano, Universidade de São Paulo,
plored in many diseases (50,54), including the switching of
Emmanuel Dias-Neto, The A.C. Camargo Cancer Hospital
BCL-X splicing in cancer (46); nevertheless, there are still
and Helder I. Nakaya, Emory University, for suggestions
hurdles that must be overcome, such as eventually achieving
and criticism.
an improved cell-killing efficiency to overcome the present
inability of these splice-switching oligonucleotides alone to
cause apoptosis (6,8,46). The possibility seems to be open FUNDING
for a very effective gene-specific splice shifting toward pro- Fundação de Amparo à Pesquisa do Estado de São
apoptotic BCL-XS that is accompanied by apoptosis, via a Paulo (FAPESP) [to S.V.A. and to E.M.R.]; FAPESP and
targeted induction of INXS lncRNA expression. We spec- Conselho Nacional de Desenvolvimento Cientı́fico e Tec-
ulate that the INXS 1.9 kb transcript is effective in caus- nológico (CNPq) through the Instituto Nacional de Ciência
ing apoptosis because this endogenous lncRNA would have e Tecnologia em Oncogenômica. C.D.P., M.S.A., K.S.P.
the potential to adopt a complex secondary/tertiary struc- and A.C.A. received fellowships from FAPESP, and M.S.A.
ture that could facilitate binding and possibly recruitment received a fellowship from CNPq prior to the one from
of splicing modulators, such as Sam68 protein, in a man- FAPESP. S.V.A. and E.M.R. received investigator fellow-
ner that might have been evolutionarily selected as part of ship awards from CNPq. Fundingfor open access charge:
a concerted mechanism. Overexpression or knockdown of Fundação de Amparo à Pesquisa do Estado de São Paulo
INXS did not affect the splicing of two other genes that (FAPESP) and Conselho Nacional de Desenvolvimento
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
are known targets of Sam68, namely, cyclin D1 (47) and Cientı́fico e Tecnológico (CNPq) through the Instituto Na-
SF2/ASF (48), suggesting that INXS does not exert a global cional de Ciência e Tecnologia em Oncogenômica.
role on Sam68-mediated alternative splicing, and that the Conflict of interest statement. None declared.
specificity might arise from a portion of INXS eventually
establishing a direct RNA/RNA base pair with the target
BCL-X pre-mRNA. Further studies on modulating the en- REFERENCES
dogenous expression of INXS and on determining the de- 1. Cory,S. and Adams,J.M. (2002) The Bcl2 family: regulators of the
tailed molecular mechanism of action of INXS are war- cellular life-or-death switch. Nat. Rev. Cancer, 2, 647–656.
2. Kelly,P.N. and Strasser,A. (2011) The role of Bcl-2 and its
ranted. pro-survival relatives in tumourigenesis and cancer therapy. Cell
Most importantly, the effect of INXS lncRNA on the Death Diff., 18, 1414–1424.
BCL-X splicing may eventually have less off-target effects 3. Tait,S.W. and Green,D.R. (2010) Mitochondria and cell death: outer
than the anti-cancer drugs, which are known to induce membrane permeabilization and beyond. Nat. Rev. Mol. Cell Biol.,
11, 621–632.
the opposing anti-apoptotic splice variants of many other 4. Boise,L.H., Gonzalez-Garcia,M., Postema,C.E., Ding,L.,
apoptotic genes, while causing a shift toward pro-apoptotic Lindsten,T., Turka,L.A., Mao,X., Nunez,G. and Thompson,C.B.
BCL-XS splicing (55). In this respect, it has been shown (1993) bcl-x, a bcl-2-related gene that functions as a dominant
that most of the 20 tested anti-cancer drugs favor the anti- regulator of apoptotic cell death. Cell, 74, 597–608.
apoptotic splicing event on the Fas gene (55), while shift- 5. Plati,J., Bucur,O. and Khosravi-Far,R. (2011) Apoptotic cell signaling
in cancer progression and therapy. Integrative Biol., 3, 279–296.
ing BCL-X splicing by different degrees toward the pro- 6. Mercatante,D.R., Bortner,C.D., Cidlowski,J.A. and Kole,R. (2001)
apoptotic isoform. Modification of alternative splicing of Bcl-x pre-mRNA in prostate
In a broader perspective, the identification of INXS as the and breast cancer cells. analysis of apoptosis and cell death. J. Biol.
first lncRNA that acts on splicing and induces apoptosis, Chem., 276, 16411–16417.
7. Minn,A.J., Boise,L.H. and Thompson,C.B. (1996) Bcl-x(S)
calls the attention to the importance of looking at the func- anatagonizes the protective effects of Bcl-x(L). J. Biol. Chem., 271,
tional roles and at the modes of induction of other lncR- 6306–6312.
NAs possibly acting on splicing events that occur in apop- 8. Taylor,J.K., Zhang,Q.Q., Wyatt,J.R. and Dean,N.M. (1999)
totic genes. Moreover, our INXS-plasmid injection experi- Induction of endogenous Bcl-xS through the control of Bcl-x
ments in a mouse xenograft model serve as a proof of con- pre-mRNA splicing by antisense oligonucleotides. Nat. Biotechnol.,
17, 1097–1100.
cept that tumor regression could be effectively obtained in 9. Sumantran,V.N., Ealovega,M.W., Nunez,G., Clarke,M.F. and
vivo. It is tempting to assume that controlling and/or condi- Wicha,M.S. (1995) Overexpression of Bcl-XS sensitizes MCF-7 cells
tionally increasing INXS expression in tumor cells could ef- to chemotherapy-induced apoptosis. Cancer Res., 55, 2507–2510.
fectively induce apoptosis and limit tumorigenesis, and that 10. Plotz,M., Gillissen,B., Hossini,A.M., Daniel,P.T. and Eberle,J. (2012)
Disruption of the VDAC2-Bak interaction by Bcl-x(S) mediates
the INXS lncRNA may represent a target to be explored in efficient induction of apoptosis in melanoma cells. Cell Death Diff.,
a molecular-based cancer therapy. 19, 1928–1938.
11. Hossini,A.M., Eberle,J., Fecker,L.F., Orfanos,C.E. and Geilen,C.C.
(2003) Conditional expression of exogenous Bcl-X(S) triggers
apoptosis in human melanoma cells in vitro and delays growth of
ACCESSION NUMBER melanoma xenografts. FEBS Lett., 553, 250–256.
12. Paronetto,M.P., Achsel,T., Massiello,A., Chalfant,C.E. and Sette,C.
INXS Accession number in GenBank: KC505631 (2007) The RNA-binding protein Sam68 modulates the alternative
splicing of Bcl-x. J. Cell Biol., 176, 929–939.
13. Massiello,A., Roesser,J.R. and Chalfant,C.E. (2006) SAP155 Binds to
ceramide-responsive RNA cis-element 1 and regulates the alternative
SUPPLEMENTARY DATA 5 splice site selection of Bcl-x pre-mRNA. FASEB J., 20, 1680–1682.
14. Montes,M., Cloutier,A., Sanchez-Hernandez,N., Michelle,L.,
Supplementary Data are available at NAR Online. Lemieux,B., Blanchette,M., Hernandez-Munain,C., Chabot,B. and12 Nucleic Acids Research, 2014
Sune,C. (2012) TCERG1 regulates alternative splicing of the Bcl-x gene Saf transcribed from the opposite strand of Fas. Hum. Mol.
gene by modulating the rate of RNA polymerase II transcription. Genet., 14, 1465–1474.
Mol. Cell. Biol., 32, 751–762. 36. Beltran,M., Puig,I., Pena,C., Garcia,J.M., Alvarez,A.B., Pena,R.,
15. Zhou,A., Ou,A.C., Cho,A., Benz,E.J. Jr. and Huang,S.C. (2008) Bonilla,F. and de Herreros,A.G. (2008) A natural antisense transcript
Novel splicing factor RBM25 modulates Bcl-x pre-mRNA 5 splice regulates Zeb2/Sip1 gene expression during Snail1-induced
site selection. Mol. Cell. Biol., 28, 5924–5936. epithelial-mesenchymal transition. Genes Dev., 22, 756–769.
16. Moore,M.J., Wang,Q., Kennedy,C.J. and Silver,P.A. (2010) An 37. Ji,P., Diederichs,S., Wang,W., Boing,S., Metzger,R., Schneider,P.M.,
alternative splicing network links cell-cycle control to apoptosis. Cell, Tidow,N., Brandt,B., Buerger,H., Bulk,E. et al. (2003) MALAT-1, a
142, 625–636. novel noncoding RNA, and thymosin beta4 predict metastasis and
17. Michelle,L., Cloutier,A., Toutant,J., Shkreta,L., Thibault,P., survival in early-stage non-small cell lung cancer. Oncogene, 22,
Durand,M., Garneau,D., Gendron,D., Lapointe,E., Couture,S. et al. 8031–8041.
(2012) Proteins associated with the exon junction complex also 38. Tripathi,V., Shen,Z., Chakraborty,A., Giri,S., Freier,S.M., Wu,X.,
control the alternative splicing of apoptotic regulators. Mol. Cell. Zhang,Y., Gorospe,M., Prasanth,S.G., Lal,A. et al. (2013) Long
Biol., 32, 954–967. noncoding RNA MALAT1 controls cell cycle progression by
18. Garneau,D., Revil,T., Fisette,J.F. and Chabot,B. (2005) regulating the expression of oncogenic transcription factor B-MYB.
Heterogeneous nuclear ribonucleoprotein F/H proteins modulate the PLoS Genet., 9, e1003368.
alternative splicing of the apoptotic mediator Bcl-x. J. Biol. Chem., 39. Tripathi,V., Ellis,J.D., Shen,Z., Song,D.Y., Pan,Q., Watt,A.T.,
280, 22641–22650. Freier,S.M., Bennett,C.F., Sharma,A., Bubulya,P.A. et al. (2010) The
19. Revil,T., Pelletier,J., Toutant,J., Cloutier,A. and Chabot,B. (2009) nuclear-retained noncoding RNA MALAT1 regulates alternative
Heterogeneous nuclear ribonucleoprotein K represses the production splicing by modulating SR splicing factor phosphorylation. Mol.
of pro-apoptotic Bcl-xS splice isoform. J. Biol. Chem., 284, Cell, 39, 925–938.
21458–21467. 40. Louro,R., Nakaya,H.I., Amaral,P.P., Festa,F., Sogayar,M.C., da
20. Lee,J., Zhou,J., Zheng,X., Cho,S., Moon,H., Loh,T.J., Jo,K. and Silva,A.M., Verjovski-Almeida,S. and Reis,E.M. (2007) Androgen
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015
Shen,H. (2012) Identification of a novel cis-element that regulates responsive intronic non-coding RNAs. BMC Biol., 5, 4.
alternative splicing of Bcl-x pre-mRNA. Biochem. Biophys. Res. 41. Kong,L., Zhang,Y., Ye,Z.Q., Liu,X.Q., Zhao,S.Q., Wei,L. and
Commun., 420, 467–472. Gao,G. (2007) CPC: assess the protein-coding potential of transcripts
21. Massiello,A., Salas,A., Pinkerman,R.L., Roddy,P., Roesser,J.R. and using sequence features and support vector machine. Nucleic Acids
Chalfant,C.E. (2004) Identification of two RNA cis-elements that Res., 35, W345–W349.
function to regulate the 5 splice site selection of Bcl-x pre-mRNA in 42. Consortium,E.P., Bernstein,B.E., Birney,E., Dunham,I., Green,E.D.,
response to ceramide. J. Biol. Chem., 279, 15799–15804. Gunter,C. and Snyder,M. (2012) An integrated encyclopedia of DNA
22. Nie,L., Wu,H.J., Hsu,J.M., Chang,S.S., Labaff,A.M., Li,C.W., elements in the human genome. Nature, 489, 57–74.
Wang,Y., Hsu,J.L. and Hung,M.C. (2012) Long non-coding RNAs: 43. Dani,C., Blanchard,J.M., Piechaczyk,M., El Sabouty,S., Marty,L.
versatile master regulators of gene expression and crucial players in and Jeanteur,P. (1984) Extreme instability of myc mRNA in normal
cancer. Am. J. Trans. Res., 4, 127–150. and transformed human cells. Proc. Natl. Acad. Sci. U.S.A., 81,
23. Cheetham,S.W., Gruhl,F., Mattick,J.S. and Dinger,M.E. (2013) Long 7046–7050.
noncoding RNAs and the genetics of cancer. Brit. J. Cancer, 108, 44. Yeh,C.T. and Yen,G.C. (2005) Effect of sulforaphane on
2419–2425. metallothionein expression and induction of apoptosis in human
24. Huarte,M. and Rinn,J.L. (2010) Large non-coding RNAs: missing hepatoma HepG2 cells. Carcinogenesis, 26, 2138–2148.
links in cancer? Hum. Mol. Genet., 19, R152–R161. 45. Beckedorff,F.C., Ayupe,A.C., Crocci-Souza,R., Amaral,M.S.,
25. Hung,T., Wang,Y., Lin,M.F., Koegel,A.K., Kotake,Y., Grant,G.D., Nakaya,H.I., Soltys,D.T., Menck,C.F., Reis,E.M. and
Horlings,H.M., Shah,N., Umbricht,C., Wang,P. et al. (2011) Verjovski-Almeida,S. (2013) The intronic long noncoding RNA
Extensive and coordinated transcription of noncoding RNAs within ANRASSF1 recruits PRC2 to the RASSF1A promoter, reducing the
cell-cycle promoters. Nat. Genet., 43, 621–629. expression of RASSF1A and increasing cell proliferation. PLoS
26. Guttman,M. and Rinn,J.L. (2012) Modular regulatory principles of Genet., 9, e1003705.
large non-coding RNAs. Nature, 482, 339–346. 46. Bauman,J.A., Li,S.D., Yang,A., Huang,L. and Kole,R. (2010)
27. Kung,J.T., Colognori,D. and Lee,J.T. (2013) Long noncoding RNAs: Anti-tumor activity of splice-switching oligonucleotides. Nucleic
past, present, and future. Genetics, 193, 651–669. Acids Res., 38, 8348–8356.
28. Mourtada-Maarabouni,M., Pickard,M.R., Hedge,V.L., Farzaneh,F. 47. Paronetto,M.P., Cappellari,M., Busa,R., Pedrotti,S., Vitali,R.,
and Williams,G.T. (2009) GAS5, a non-protein-coding RNA, Comstock,C., Hyslop,T., Knudsen,K.E. and Sette,C. (2010)
controls apoptosis and is downregulated in breast cancer. Oncogene, Alternative splicing of the cyclin D1 proto-oncogene is regulated by
28, 195–208. the RNA-binding protein Sam68. Cancer Res., 70, 229–239.
29. Huarte,M., Guttman,M., Feldser,D., Garber,M., Koziol,M.J., 48. Valacca,C., Bonomi,S., Buratti,E., Pedrotti,S., Baralle,F.E., Sette,C.,
Kenzelmann-Broz,D., Khalil,A.M., Zuk,O., Amit,I., Rabani,M. et al. Ghigna,C. and Biamonti,G. (2010) Sam68 regulates EMT through
(2010) A large intergenic noncoding RNA induced by p53 mediates alternative splicing-activated nonsense-mediated mRNA decay of the
global gene repression in the p53 response. Cell, 142, 409–419. SF2/ASF proto-oncogene. J. Cell Biol., 191, 87–99.
30. Wang,X., Arai,S., Song,X., Reichart,D., Du,K., Pascual,G., 49. Nakaya,H.I., Amaral,P.P., Louro,R., Lopes,A., Fachel,A.A.,
Tempst,P., Rosenfeld,M.G., Glass,C.K. and Kurokawa,R. (2008) Moreira,Y.B., El-Jundi,T.A., da Silva,A.M., Reis,E.M. and
Induced ncRNAs allosterically modify RNA-binding proteins in cis Verjovski-Almeida,S. (2007) Genome mapping and expression
to inhibit transcription. Nature, 454, 126–130. analyses of human intronic noncoding RNAs reveal tissue-specific
31. Zhang,X., Rice,K., Wang,Y., Chen,W., Zhong,Y., Nakayama,Y., patterns and enrichment in genes related to regulation of
Zhou,Y. and Klibanski,A. (2010) Maternally expressed gene 3 transcription. Genome Biol., 8, R43.
(MEG3) noncoding ribonucleic acid: isoform structure, expression, 50. Djebali,S., Davis,C.A., Merkel,A., Dobin,A., Lassmann,T.,
and functions. Endocrinology, 151, 939–947. Mortazavi,A., Tanzer,A., Lagarde,J., Lin,W., Schlesinger,F. et al.
32. Burge,C.B., Tuschl,T. and Sharp,P.A. (1999) In: Gesteland,R.F., (2012) Landscape of transcription in human cells. Nature, 489,
Cech,T.R. and Atkins,J.F. (eds.). The RNA World. Cold Spring 101–108.
Harbor Laboratory Press, Cold Spring Harbor, pp. 525–560. 51. Birney,E., Stamatoyannopoulos,J.A., Dutta,A., Guigo,R.,
33. Morrissy,A.S., Griffith,M. and Marra,M.A. (2011) Extensive Gingeras,T.R., Margulies,E.H., Weng,Z., Snyder,M.,
relationship between antisense transcription and alternative splicing Dermitzakis,E.T., Thurman,R.E. et al. (2007) Identification and
in the human genome. Genome Res., 21, 1203–1212. analysis of functional elements in 1% of the human genome by the
34. Krystal,G.W., Armstrong,B.C. and Battey,J.F. (1990) N-myc mRNA ENCODE pilot project. Nature, 447, 799–816.
forms an RNA-RNA duplex with endogenous antisense transcripts. 52. Pandey,R.R., Mondal,T., Mohammad,F., Enroth,S., Redrup,L.,
Mol. Cell. Biol., 10, 4180–4191. Komorowski,J., Nagano,T., Mancini-Dinardo,D. and Kanduri,C.
35. Yan,M.D., Hong,C.C., Lai,G.M., Cheng,A.L., Lin,Y.W. and (2008) Kcnq1ot1 antisense noncoding RNA mediates lineage-specific
Chuang,S.E. (2005) Identification and characterization of a novelNucleic Acids Research, 2014 13
transcriptional silencing through chromatin-level regulation. Mol. 54. Spitali,P. and Aartsma-Rus,A. (2012) Splice modulating therapies for
Cell, 32, 232–246. human disease. Cell, 148, 1085–1088.
53. Bielli,P., Busa,R., Di Stasi,S.M., Munoz,M.J., Botti,F., 55. Shkreta,L., Froehlich,U., Paquet,E.R., Toutant,J., Elela,S.A. and
Kornblihtt,A.R. and Sette,C. (2014) The transcription factor FBI-1 Chabot,B. (2008) Anticancer drugs affect the alternative splicing of
inhibits SAM68-mediated BCL-X alternative splicing and apoptosis. Bcl-x and other human apoptotic genes. Mol. Cancer Ther., 7,
EMBO Rep., 15, 419–427. 1398–1409.
Downloaded from http://nar.oxfordjournals.org/ by guest on November 1, 2015You can also read

















































