NGF Enhances CGRP Release Evoked by Capsaicin from Rat Trigeminal Neurons: Differential Inhibition by SNAP-25-Cleaving Proteases
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
International Journal of
Molecular Sciences
Article
NGF Enhances CGRP Release Evoked by Capsaicin from Rat
Trigeminal Neurons: Differential Inhibition by
SNAP-25-Cleaving Proteases
Mariia Belinskaia , Tomas Zurawski, Seshu Kumar Kaza, Caren Antoniazzi , J. Oliver Dolly and Gary W. Lawrence *
International Centre for Neurotherapeutics, Dublin City University, Collins Avenue, D09 V209 Dublin, Ireland;
mariia.belinskaia2@mail.dcu.ie (M.B.); tom.zurawski@dcu.ie (T.Z.); seshukumar.kaza@dcu.ie (S.K.K.);
caren.antoniazzi@dcu.ie (C.A.); oliver.dolly@dcu.ie (J.O.D.)
* Correspondence: gary.lawrence@dcu.ie; Tel.: +353-1-700-7689
Abstract: Nerve growth factor (NGF) is known to intensify pain in various ways, so perturbing
pertinent effects without negating its essential influences on neuronal functions could help the
search for much-needed analgesics. Towards this goal, cultured neurons from neonatal rat trigem-
inal ganglia—a locus for craniofacial sensory nerves—were used to examine how NGF affects the
Ca2+ -dependent release of a pain mediator, calcitonin gene-related peptide (CGRP), that is triggered
by activating a key signal transducer, transient receptor potential vanilloid 1 (TRPV1) with capsaicin
(CAP). Measurements utilised neurons fed with or deprived of NGF for 2 days. Acute re-introduction
of NGF induced Ca2+ -dependent CGRP exocytosis that was inhibited by botulinum neurotoxin type
A (BoNT/A) or a chimera of/E and/A (/EA), which truncated SNAP-25 (synaptosomal-associated
protein with Mr = 25 k) at distinct sites. NGF additionally caused a Ca2+ -independent enhancement
of the neuropeptide release evoked by low concentrations (43 ◦ C), protons and various chemicals including capsaicin
4.0/). (CAP), the component of chilli peppers responsible for causing heat sensation [7]. These
Int. J. Mol. Sci. 2022, 23, 892. https://doi.org/10.3390/ijms23020892 https://www.mdpi.com/journal/ijms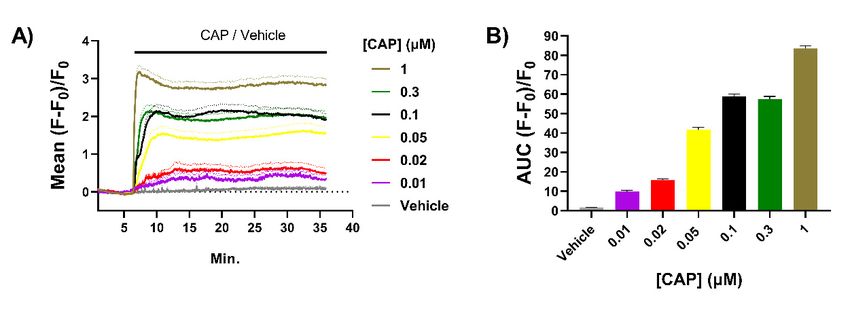
Int. J. Mol. Sci. 2022, 23, 892 2 of 20
activators open a channel pore in TRPV1 to permit an influx of Na+ and Ca2+ . TRPV1 is
the unique receptor for CAP, so the latter is a convenient tool for studying this channel
and neurons that express it [8–10]. Tissue damage after injury or infection enhances the
generation of pain signals by lowering the activation thresholds of nociceptors. This
results in heightened sensitivity to painful stimuli (hyperalgesia) and the production of
pain signals under normally innocuous conditions (allodynia). Inflammation-associated
sensitisation involves the release of pro-algesic affectors from mast cells, macrophages, and
sensory neurons at sites of injury or infection [11]. This normally serves to protect damaged
tissues from further harm but, under certain pathological conditions, inflammation may
persist even after the original trigger has been resolved. The resultant hypersensitivity of
nociceptors can contribute to persistent chronic pain.
TRPV1-mediated nociception is implicated in heat hyperalgesia induced by inflam-
mation [8,9], and has been identified as a target of signalling cascades activated by inflam-
matory mediators such as NGF, which modulates nociceptor sensitivity in addition to its
classical developmental roles in the survival and differentiation of sensory and sympathetic
neurons [4]. NGF levels are elevated in patients with chronic pain conditions such as
osteoarthritis, lower back pain, interstitial cystitis, rheumatoid arthritis, spondylarthritis,
and migraine [4,12]. Inflammatory and certain other cell types synthesise and release NGF
under pathological conditions including, but not limited to, keratinocytes, mast cells and
macrophages [3,4]. Notably, none of these cell types express detectable levels of NGF in
their normal quiescent state in adults [4]. Nerve injury does not induce the upregulation of
NGF expression in sensory neurons themselves [4]. Preclinical studies on rats showed that
intra-plantar injection of NGF induces heat hyperalgesia within minutes attributable to
sensitisation of peripheral nociceptors [13]. There are two receptors for NGF: tropomyosin
receptor kinase A (TrkA) and p75 neurotrophin receptor [3,4], but TrkA-mediated sig-
nalling seems more critical in pain development because p75 knockout mice still develop
acute mechanical and heat hypersensitivity after subcutaneous administration of NGF [14].
Prolonged activation of TrkA signalling enhances the expression of many genes encod-
ing proteins and peptides linked to pain signalling, including TRPV1 and the calcitonin
gene-related peptide (CGRP) [4]. Whilst NGF binding to TrkA promotes neuronal survival
via activation of Ras and the downstream extracellular related kinases 1 and 2 (ERK 1/2),
which involves retrograde transport of signalling endosomes, TrkA elicits a more immedi-
ate and local modulation of nociceptors via stimulation of phosphoinositide 3-kinase and
phospholipase C [4]. Exposure of sensory neurons to NGF results in sensitisation to CAP
within minutes, indicating an early potentiation of TRPV1 without altered gene expres-
sion [15–18]. In this regard, three mechanisms of NGF-induced acute TRPV1 potentiation
have been described: modulation of TRPV1 channel-opening probability [19], prevention
of agonist-induced desensitisation [20], and fast mobilisation of additional channels from
intracellular stores to the neuronal membrane through regulated exocytosis [18,21,22]. All
of those involve NGF-TrkA initiated signalling cascades that culminate in the increased
phosphorylation of TRPV1, but at different sites. Phosphorylation at Y200 increases traffick-
ing of TRPV1 to the cell membrane, whereas phosphate addition to S502 and S801 alters
channel opening probability [18].
Most TRPV1-positive neurons express CGRP, and CAP is an established trigger for
migraine attacks linked to its induction of CGRP release [12,23]. CGRP is a powerful
vasodilator, instigator of neurogenic inflammation plus flare, and its elevated blood levels
during migraine attacks [24] are suggestive of increased release in this prevalent painful
condition. Paradoxically, prolonged activation of TRPV1 with agonists, CAP and civamide,
can reduce headache pain by causing a durable denervation of fibres that express this
channel, but their clinical use has been restricted due to the severity of on-target side
effects of burning pain and lacrimation, whilst clinical trials with TRPV1 antagonists are
ongoing [23]. An alternative migraine treatment that has gained traction is the targeted
blockade of CGRP exocytosis. In fact, botulinum neurotoxin type A (BoNT/A), a potent
and specific inhibitor of transmitter release [25,26] due to truncation and inactivationInt. J. Mol. Sci. 2022, 23, 892 3 of 20
of SNAP-25 (synaptosomal-associated protein with Mr = 25k), received FDA approval
(BOTOX® , a complex of BoNT/A and non-toxic proteins) for treating chronic migraine
but not the episodic form [27]. This came after BOTOX® injections were shown in certain
patients to reduce the frequency and severity of headache episodes in the Phase III Research
Evaluating Migraine Prophylaxis (PREEMPT) series of clinical trials [28,29]. Follow-up
studies reported that patients who respond well to BOTOX® display higher serum levels
of CGRP prior to treatment than non-responders, and after treatment serum levels are
significantly reduced in responders only [30]. However, even in responders, the frequency
and severity of migraine attacks are only partially reduced. It is well established that the
short truncation of SNAP-25 by BoNT/A (it removes only nine residues from its C-terminus)
destabilises SNARE complexes but does not prevent their formation [31]; consequently,
BoNT/A only retards Ca2+ dependent neurotransmitter release [32,33]. The protease of
BoNT/E removes a larger C-terminal fragment (26 residues) and this is enough to prevent
stable (i.e., SDS-resistant) SNARE complexes forming [31,32], but trigeminal ganglion
neurons (TGNs) are insensitive to BoNT/E due to a paucity of glycosylated SV2A and
SV2B, the essential protein component of its high-affinity neuronal receptor [32,34]. This
impediment to the delivery of BoNT/E protease was circumvented by genetic engineering
to create a recombinant chimera,/EA, having the SV2A,B and C binding (HC ) portion
of/A [35] fused to the translocation (HN ) and protease light chain (LC) domains of/E [36].
In terms of the fraction of SNAP-25 cleaved, sensory neurons are almost as susceptible
to/EA [32] as they are to BoNT/A [37], but with a smaller and less functional product
created. Thus, a more complete blockade of CGRP exocytosis evoked by 1 µM CAP was
achieved with BoNT/EA [32] compared to BoNT/A [37]. Hence, with the long-term goal
of improving the therapeutic utility of such neurotoxins (reviewed by Rasetti-Escargueil
and Popoff [38]), herein induction of Ca2+ -dependent CGRP release from sensory neurons
by NGF, and its Ca2+ -independent enhancement of CAP-evoked neuropeptide exocytosis,
were shown to be inhibited by/EA and to a lesser extent by/A.
2. Results
2.1. Exposure to CAP Induces Concentration-Dependent Increases in Intracellular Ca2+ ([Ca2+ ]i )
in Neonatal Rat TGNs In Vitro
Cultured sensory neurons from neonatal rodents have been used extensively to study
the potentiation by NGF of signalling by TRPV1 and related channels [15,21]. Compared to
mature neurons from adult animals, these are relatively easy to isolate and cultivate at high
density; also, ~85% of these neurons maintained in the presence of NGF express TRPV1 [39]
and almost all of these co-express CGRP [37,40]. Binding of CAP causes the opening of
a channel in TRPV1 that conducts Ca2+ (and other cations, mainly Na+ ) into neurons,
and microscopic imaging of DRG neurons (DRGNs) or TGNs loaded with Ca2+ -sensitive
fluorescent dyes is a convenient method to study this nociceptive process [8,32]. Herein,
TGNs were isolated from neonatal rats and cultured in the presence of NGF for 4 days
before experiments. Just prior to confocal microscope imaging, cells were loaded with
Fluo-4 AM as detailed in the Materials and Methods. After washing to remove unloaded
dye, image recordings were begun with continuous superfusion of the recording buffer, and
following a 6 min. period to record baseline signals, TGNs were exposed to CAP for 30 min.
with recording continued throughout. A new sample of naïve cells was used for recording
of responses to each CAP concentration ([CAP]) to avoid results being confounded by
agonist-induced desensitisation. Increases in [Ca2+ ]i were observed in 60% of the TGNs
exposed to 0.01 µM CAP, the lowest concentration tested, rising to 85% of the neurons with
0.05 µM and remaining at about this level for higher [CAP], in good agreement with the
fraction of TGNs found to express TRPV1 by immuno-histochemistry. The mean increase
in fluorescence in responsive cells at each time point was calculated as a fraction of initial
intensity (i.e., before exposure to CAP) and plotted + s.e.m. against time (Figure 1A). This
highlights the escalation of [Ca2+ ]i as [CAP] was raised. Note also that average fluorescence
intensified more rapidly with each increment in [CAP] (Figure 1A). A plot of the areamean increase in fluorescence in responsive cells at each time point was calculated a
fraction of initial intensity (i.e., before exposure to CAP) and plotted + s.e.m. against ti
(Figure 1A). This highlights the escalation of [Ca2+]i as [CAP] was raised. Note also t
average fluorescence intensified more rapidly with each increment in [CAP] (Figure 1
Int. J. Mol. Sci. 2022, 23, 892 A plot of the area under the curve (AUC; Figure 1B) of mean fluorescence induced 4 of 20 by e
[CAP] over time illustrates a clear relationship between [CAP] and sustained increase
[Ca2+]i. These results accord with the [CAP] dose–response obtained by Ca2+-imaging
under the curve (AUC; Figure 1B)
DRGNs [41]. By increasing [Ca2+]of mean fluorescence induced by each [CAP] over time
i in TGNs due to activation of TRPV1 with CAP, exo
illustrates a clear relationship between [CAP] and sustained increases in [Ca2+ ]i . These
tosisresults
of theaccord
pain with
signalling neuropeptide
the [CAP] dose–response CGRP [32]byisCatriggered.
obtained 2+ -imaging Thus,
in DRGNsit is[41].
of interes
elucidate the impact
By increasing [Ca on
2+ ]i inCGRP
TGNs release of factors
due to activation of known
TRPV1 with to modulate TRPV1
CAP, exocytosis of activity,
the su
as NGF.
pain signalling neuropeptide CGRP [32] is triggered. Thus, it is of interest to elucidate the
impact on CGRP release of factors known to modulate TRPV1 activity, such as NGF.
Figure 1. Capsaicin (CAP) induces dose-dependent increases of [Ca2+ ]i in cultured trigeminal
Figure 1. Capsaicin
ganglion (CAP) Neurons
neurons (TGNs). induceswere dose-dependent
cultured for 4 daysincreases
with nerveof growth
[Ca2+]i factor
in cultured trigeminal g
(NGF) before
glionbeing
neurons
loaded(TGNs).
with Fluo-4 Neurons
AM and were cultured
recording for 4intensity
fluorescence days with nervemicroscopy.
by confocal growth factor (NGF) bef
(A) Solid
beinglines
loaded with Fluo-4 AM and recording fluorescence intensity by confocal
indicate the mean fluorescence (F) relative to initial fluorescence (F0 ), which was calculated microscopy.
Solid[(F
lines
− F0indicate the exposed
)/F0 ] for cells mean fluorescence (F) relativeofto
to different concentrations CAPinitial fluorescence
([CAP]) and plotted (F 0), which
against time. was ca
latedBroken 0)/F0indicate
[(F − Flines ] for cells
meanexposed to different
values +s.e.m. (standard concentrations
error of the mean)of nCAP ([CAP])
> 50 cells and[CAP].
for each plotted aga
(B) Area under the curve (AUC) of mean fluorescence traces recorded for
time. Broken lines indicate mean values +s.e.m. (standard error of the mean) n > 50 cells for eeach [CAP] exposure.
[CAP].Column heights
(B) Area and error
under bars indicate
the curve (AUC) mean AUC and
of mean s.e.m, respectively.
fluorescence traces recorded for each [CAP] ex
sure. 2.2.
Column heights and
NGF Withdrawal fromerror
TGNsbars indicate
In Vitro Reducesmean AUC and and
Both Spontaneous s.e.m, respectively.
CAP-Evoked CGRP Release
Immature TGNs are highly dependent for survival on the presence of NGF in the
2.2. NGF Withdrawal
culture medium and from
for TGNs
elicitingInthe
Vitro
highReduces Both
expression of Spontaneous
the signalling and CAP-Evoked
components [4]. CGR
Release
Therefore, it was optimal to cultivate the neurons with this neurotrophin for some time
before its withdrawal and later re-addition in order to examine the effects of re-introducing
Immature TGNs are highly dependent for survival on the presence of NGF in
NGF on CGRP exocytosis, under resting conditions and upon activating TRPV1 with CAP.
culture medium
TGNs wereand for eliciting
cultivated for 2 daysthe in high expression
the presence of NGFof(50the signalling
ng/mL) components
before transfer
Therefore, it wasmedium
into a culture optimal to cultivate
lacking the neurons
this neurotrophin with
and were this neurotrophin
maintained for some ti
for 2 further days
(Figure 2A); anti-NGF antibodies were also added to neutralise any
before its withdrawal and later re-addition in order to examine the effects of re-introd remaining traces.
Note that some TGNs were continuously exposed to NGF for the entire 4 days (‘fed’)
ing NGF on CGRP exocytosis, under resting conditions and upon activating TRPV1 w
to serve as controls. Deprivation of NGF did not significantly change the total amount
CAP.of protein present (27.3 ± 2.7 vs. 24.0 ± 2.4 µg/well [mean ± s.e.m.] in fed or starved
TGNs
cells, were cultivated
respectively; p = 0.38,for 2 days
Figure 2B),inorthe presence
their of NGF
CGRP contents (50 ±
(1293 ng/mL) before
125 [fed] vs. trans
1339 ± 127 [starved] pg/well; p = 0.8, Figure 2C), total TRPV1 expression
into a culture medium lacking this neurotrophin and were maintained for 2 further d (data not shown)
and 2A);
(Figure the proportion
anti-NGF of cells expressing
antibodies were TRPV1alsoremained
added high (~74%) as determined
to neutralise any remainingby trac
immuno-histochemistry. By contrast, spontaneous CGRP release was significantly lower
Noteforthat some TGNs were continuously exposed to NGF for the entire 4 days (‘fed’
the starved neurons (0.88 ± 0.06% of total CGRP, c.f. 1.19 ± 0.09% in fed cells; p = 0.009,
serveFigure
as controls. Deprivation
2D). Likewise, NGF removal of NGF
decreaseddidCGRPnot significantly
exocytosis evoked change theCAP
by 20 nM total(toamoun
protein
7.65 present
± 1.2% of(27.3 ± 2.7 vs.
total CGRP, from24.0
16.78±±2.4 2.6;µg/well
p = 0.007,[mean ± s.e.m.]
Figure 2E). in fedstarving
In summary, or starved ce
respectively; p = 0.38, Figure 2B), or their CGRP contents (1293 ± 125 [fed]CGRP
TGNs of NGF for 2 days had no significant effect on the expression of total protein or vs. 1339 ±
but reduced the fraction of the latter released spontaneously or upon stimulation with a
[starved] pg/well; p = 0.8, Figure 2C), total TRPV1 expression (data not shown) and
fixed low [CAP].
proportion of cells expressing TRPV1 remained high (~74%) as determined by immu
histochemistry. By contrast, spontaneous CGRP release was significantly lower for
starved neurons (0.88 ± 0.06% of total CGRP, c.f. 1.19 ± 0.09% in fed cells; p = 0.009, Fig
2D). Likewise, NGF removal decreased CGRP exocytosis evoked by 20 nM CAP (to 7.6
1.2% of total CGRP, from 16.78 ± 2.6; p = 0.007, Figure 2E). In summary, starving TGNInt. J. Mol. Sci. 2022, 23, x FOR PEER REVIEW 5 of 21
NGF for 2 days had no significant effect on the expression of total protein or CGRP but
Int. J. Mol. Sci. 2022, 23, 892 reduced the fraction of the latter released spontaneously or upon stimulation with a fixed
5 of 20
low [CAP].
Pre-treatment Release experiment
A)
2 days 2 days 30 min 30 min
+ NGF + NGF (Fed) HBS HBS/CAP
- NGF (Starved)
B) C) 1500
1000
500
0
Fed Starved
✱✱
D) E) ✱✱
Figure 2. Deprival of NGF for 2 days does not significantly alter total protein or calcitonin gene-
Figure
related 2. Deprival
peptide of NGF
(CGRP) for 2 days
contents of ratdoes not significantly
cultured alter total
TGNs but reduces protein or calcitonin
both spontaneous gene-
and CAP-evoked
related peptide (CGRP) contents of rat cultured TGNs but reduces both spontaneous
exocytosis of the neuropeptide. (A) Schematic illustrating the experimental protocol. Rat TGNs wereand CAP-
evoked exocytosis of the neuropeptide. (A) Schematic illustrating the experimental protocol. Rat
cultivated initially in the presence of NGF (50 ng/mL) for 2 days before its withdrawal from half
TGNs were cultivated initially in the presence of NGF (50 ng/mL) for 2 days before its withdrawal
of the wells (starved) or retention for the other cohort over the 4 days (fed). The amounts of CGRP
from half of the wells (starved) or retention for the other cohort over the 4 days (fed). The amounts
released were quantified during sequential 30 min. exposures, firstly to HEPES buffered saline (HBS)
of CGRP released were quantified during sequential 30 min. exposures, firstly to HEPES buffered
only ((D),
saline (HBS)spontaneous release), and release),
only ((D), spontaneous then to 20 nMthen
and CAPtoin20HBS
nM (E).
CAPAtinthe
HBSend of At
(E). experiments,
the end of cells
were solubilised
experiments, cellswith
were1% (v/v) Triton
solubilised withX-100 in HBS,
1% (v/v) Tritonthen total
X-100 inprotein (B) total
HBS, then and CGRP
proteincontents
(B) and (C)
were determined,
CGRP contents (C)aswere
detailed in Materials
determined, and Methods.
as detailed Data are
in Materials andpresented
Methods.asData
mean s.e.m., n ≥as12,
are+presented
mean
N = 3.+Asterisks
s.e.m., n ≥summarise
12, N = 3. Asterisks
the resultssummarise
of unpairedthe results
t-tests ofWelch’s
with unpaired t-tests with
correction, ** pWelch’s cor-
< 0.01, shown
rection, ** p < 0.01, shown only
only for significant differences. for significant differences.
2.3. Depriving
2.3. Depriving TGNs
TGNs ofof NGF
NGF Reduces
Reduces the
the Amount
Amount of of CGRP
CGRP Release
Release Evoked
Evoked by by CAP
CAP
To glean
To glean further
further insight
insight into
into the
the consequences
consequences of of NGF
NGF removal,
removal, thethe starved
starved and
and fed
fed
neurons were
neurons werestimulated
stimulatedwithwitha range
a range of increasing
of increasing [CAP],
[CAP], using
using a slightly
a slightly modified
modified pro-
protocol
tocol (Figure
(Figure 3A)3A) to introduce
to introduce a second
a second 3030min
minincubation
incubationwith withHEPES
HEPESbuffered
buffered saline
saline
(HBS) prior to CAP stimulation. This extra step was to accommodate a
(HBS) prior to CAP stimulation. This extra step was to accommodate a further manipula-further manipulation
which
tion is detailed
which later;later;
is detailed otherwise, the procedure
otherwise, remained
the procedure the same
remained theassame
in Figure
as in 2A. In both
Figure 2A.
fed and starved TGNs, stimulation with as little as 0.01 µM CAP produced
In both fed and starved TGNs, stimulation with as little as 0.01 µM CAP produced a de- a detectable
amount amount
tectable of CGRPofrelease
CGRP above
releasethe baseline
above (i.e., spontaneous)
the baseline level and
(i.e., spontaneous) increments
level in
and incre-
[CAP] in
ments produced increases increases
[CAP] produced that reachedthata reached
peak at 0.1 µM (Figure
a peak at 0.1 µM3B).(Figure
Not only 3B).did further
Not only
raising [CAP] fail to augment the amount of CGRP released from either fed or starved
cells, in both cases a decline was observed with the higher [CAP]. Importantly, at all [CAP]
tested the amount of CGRP release observed for NGF-starved cells was always less than
the corresponding quantity seen with the fed neurons. The inclining phases of each doseInt. J. Mol. Sci. 2022, 23, x FOR PEER REVIEW 6 of 21
did further raising [CAP] fail to augment the amount of CGRP released from either fed or
Int. J. Mol. Sci. 2022, 23, 892
starved cells, in both cases a decline was observed with the higher [CAP]. Importantly, at
6 of 20
all [CAP] tested the amount of CGRP release observed for NGF-starved cells was always
less than the corresponding quantity seen with the fed neurons. The inclining phases of
each dose response curve (0.01 to 0.1 µM [CAP]) were fit with four-parameter logistic
response curve (0.01 to 0.1 µM [CAP]) were fit with four-parameter logistic functions to
functions to determine the maximum fraction of CGRP released from fed cells (37.3 ± 7.1%
determine the maximum fraction of CGRP released from fed cells (37.3 ± 7.1% of total
of total content); this was almost 1.5-times higher than the corresponding maximum
content); this was almost 1.5-times higher than the corresponding maximum evoked from
evoked from starved cells (25.2 ± 2.1%) (Figure 3B). Notably, NGF withdrawal induced a
starved cells (25.2 ± 2.1%) (Figure 3B). Notably, NGF withdrawal induced a small but just
small but just significant increase in EC50 values (32.6 ± 1.7 vs. 40.9 ± 1.8, p = 0.045). Thus,
significant increase in EC50 values (32.6 ± 1.7 vs. 40.9 ± 1.8, p = 0.045). Thus, depriving
depriving neonatal rat TGNs of NGF for 48 h in vitro reduces their ability to exocytose
neonatal rat TGNs of NGF for 48 h in vitro reduces their ability to exocytose CGRP in
CGRP in response to CAP and their apparent sensitivity to the TRPV1 agonist.
response to CAP and their apparent sensitivity to the TRPV1 agonist.
Pre-treatment Release experiments
A)
2 days 2 days 30 min 30 min 30 min
+ NGF + NGF (Fed) HBS HBS ± NGF HBS/CAP
- NGF (Starved) or
60 mM K+ HBS
CAP-evoked CGRP release (% of total)
B) C)
✱
60 mM K+ -evoked CGRP release
No NGF
D) 2.5 E) 30
+ NGF
2.0
(% of total)
20
1.5
1.0
10
0.5
0.0 0
Fed Starved Fed Starved
Figure 3. NGF withdrawal for 2 days from cultured TGNs reduces CGRP release stimulated by
Figure 3. NGF withdrawal for 2 days from cultured TGNs reduces CGRP release stimulated by
CAP; acute NGF induces exocytosis and enhances that stimulated with low [CAP]. (A) Timeline for
CAP; acute NGF induces exocytosis and enhances that stimulated with low [CAP]. (A) Timeline for
pre-treatment
pre-treatmentand andexperimental
experimentalmanipulations
manipulations ofof TGNs
TGNs to to
determine
determine thethe
effect of NGF
effect starvation
of NGF and
starvation
its
andbrief re-introduction
its brief (100 ng/mL)
re-introduction on CGRP
(100 ng/mL) releaserelease
on CGRP under requisite conditions.
under requisite (B) Dose-response
conditions. (B) Dose-
relationship between [CAP]
response relationship betweenand[CAP]
CGRPandrelease
CGRP expressed as a % of total.
release expressed as a % Asterisks
of total.show significant
Asterisks show
significant differences
differences between
between starved starved
and and NGF
NGF acutely acutely
treated treated
cells (* pInt. J. Mol. Sci. 2022, 23, 892 7 of 20
2.4. Brief Re-Exposure to NGF Induces CGRP Exocytosis from Starved TGNs and Augments the
Amount Released in Response to Subsequent Stimulation with CAP
It has been established that brief exposure of previously-starved TGNs to NGF en-
hances CAP-evoked currents and increases [Ca2+ ]i by augmenting TRPV1 activity [15,21].
So, the experimental protocol was modified to examine the consequence of acute NGF re-
exposure of starved cells for CAP-evoked CGRP release. This involved a 30 min incubation
of the cells with NGF (100 ng/mL) just prior to stimulating them with the various [CAP]
described above (Figure 3A,B). Such an acute exposure to NGF did not significantly change
the total CGRP content (Figure 3C), but it did elicit a two-fold increment in CGRP release
compared to the spontaneous level in starved cells incubated with HBS alone (Figure 3D,
p = 0.02). By contrast, in cells that had been continuously fed with NGF, the omission
(or inclusion) of NGF during the 30 min recording period did not change the amount of
spontaneous release (1.3% vs. 1.3% of total CGRP; Figure 3D). Presumably, the signalling
events underpinning the aforementioned NGF-induced CGRP release in starved cells arose
from the sudden return of a depreciated activity that was maintained constitutively in
cells continuously fed with NGF. Immediately after the short application of NGF to the
starved TGNs, they were exposed to a range of [CAP], from 10 nM–1 µM. Remarkably,
acute treatment with NGF reversed the starvation-associated reduction in CGRP release
evoked by low [CAP] (10 and 25 nM, p = 0.001 and p = 0.036 respectively, Figure 3B), was
partially effective for intermediate [CAP] (35–50 nM), and caused a minor increase in CGRP
release evoked from starved cells by higher [CAP] (0.1–1 µM). Fitting the inclining phase
with a logistic function, as described before, confirmed that acute exposure of starved TGNs
to NGF did not increase the maximum fraction of total CGRP that could be released upon
stimulation with CAP (~25%) (Figure 3B). In contrast, the EC50 for [CAP] was lowered from
40.9 ± 1.8 nM (starved cells without acute NGF) to 31.6 ± 3.6 nM (starved cells after acute
exposure to NGF), marking a restoration to the sensitivity displayed by TGNs fed continu-
ously with NGF (EC50 = 32.6 ± 1.5 nM). Acute exposure to NGF selectively enhanced CGRP
release triggered by activation of TRPV1 because the amount evoked by depolarisation
with 60 mM K+ was not altered significantly in either fed or starved cells (Figure 3E).
2.5. NGF Requires Extracellular Ca2+ for Inducing CGRP Release but Not Its Enhancement of
20 nM CAP-Evoked CGRP Exocytosis
Whilst CAP-evoked CGRP release from cultured TGNs is strictly dependent on the
presence of extracellular Ca2+ [37], it seems that this is not the case for the enhancement by
brief exposure to NGF of CAP-evoked TRPV1 currents and [Ca2+ ]i accumulation [17,42].
Thus, the pertinent question of whether NGF-induced CGRP release requires Ca2+ was
addressed using the protocol in Figure 4A. As described previously, TGNs were NGF starved
before measuring spontaneous release (during the first of three 30 min. incubation periods)
and then split into three cohorts. Two of these were exposed during a second 30 min. period
to NGF, one of them in the presence of Ca2+ whilst the other was incubated without added
Ca2+ and with 2 mM EGTA included to chelate any traces. The third batch was incubated
without NGF but with Ca2+ . After completing the latter stage, all three were then stimulated
(a third 30 min. incubation) with 20 nM CAP in the presence of Ca2+ (which, as noted above,
is essential for triggering CAP-evoked CGRP release). The absence of Ca2+ during acute
exposure to NGF did not prevent its induction of ERK1/2 phosphorylation (Figure 4B).
Indeed, a ~three-fold increase in the ratio of p-ERK1/2: total ERK 1/2 was observed relative
to the proportion in continuously-starved cells (p = 0.02; Figure 4C), confirming effective
activation of the NGF-TrkA signalling pathway. In fact, the latter ratio obtained in the
absence of Ca2+ was indistinguishable from the proliferation of p-ERK 1/2 induced by
NGF in the presence of Ca2+ (Figure 4C). By contrast, the absence of Ca2+ abolished the
NGF-induced small increase in CGRP release relative to the spontaneous level (Figure 4D).
Despite this, an approximately two and a half-fold enhancement by NGF of 20 nM CAP-
evoked Ca2+ -dependent CGRP release remained unaffected by the presence or absence of
Ca2+ during the 30 min. that the cells were exposed to the neurotrophin (Figure 4E).Ca2+ abolished the NGF-induced small increase in CGRP release relative to the spontane-
ous level (Figure 4D). Despite this, an approximately two and a half-fold enhancement by
NGF of 20 nM CAP-evoked Ca2+-dependent CGRP release remained unaffected by the
Int. J. Mol. Sci. 2022, 23, 892 presence or absence of Ca2+ during the 30 min. that the cells were exposed to the neuro-
8 of 20
trophin (Figure 4E).
Pre-treatment Release experiments
2 days 2 days 30 min 30 min 30 min
+ NGF - NGF HBS HBS ± NGF HBS/CAP
or
Ca2+-free HBS + NGF
✱
✱✱
- + + Acute NGF 2.0
+ + - Ca2+
Mr (k) 1.5
p-ERK 1/2
37
1.0
total ERK 1/2
37 0.5
25 SNAP-25
0.0
F
F
N e
G
G
e
S - fr
N
N
F
G
H a 2+
no
+
S
+
S,
B
C
H
B
B
H
✱✱
20 nM CAP-evoked CGRP release
Spontaneous or NGF-induced
20 ✱✱
✱✱✱✱ ✱✱✱
CGRP release (% of total)
15
(% of total)
10
5
0
F
F
N e
F
F
G
G
e
N e
G
G
S - fr
e
N
N
F
S - fr
N
N
F
G
2+
+
no
G
H a 2+
o
+
S
n
a
S
+
S,
B
C
+
S,
B
H
C
B
H
B
B
H
B
H
H
Figure 4. Extracellular Ca2+ 2+ is required for NGF to raise CGRP release but not for its enhancement of
Figure 4. Extracellular
2+ Ca is required for NGF to raise CGRP release but not for its enhancement
CAP-evoked Ca -dependent exocytosis. (A) NGF-starved TGNs were exposed in sequence for three
of CAP-evoked Ca2+-dependent exocytosis. (A) NGF-starved TGNs were exposed in sequence for
30 min.
three periods
30 min. as follows.
periods In theInfirst
as follows. theperiod, all cells
first period, all were exposed
cells were to HBS
exposed to only. For period
HBS only. 2, the
For period
cells 2+
2, thewere
cells split
wereinto
splitthree
intocohorts and incubated
three cohorts with Cawith
and incubated /HBS
Ca2+modified
/HBS modifiedas follows: CohortCohort
as follows: 1, HBS
only;
1, HBSCohort 2, Ca2+ /HBS
only; Cohort 2, Ca2+containing 100 ng/mL
/HBS containing NGF; Cohort
100 ng/mL NGF; 3, HBS containing
Cohort 100 ng/mL
3, HBS containing 100NGF
ng/mLbut
NGF
with Cabut2+with Ca2+ replaced
replaced by 2 mM by EGTA.2 mM EGTA. 3,
In period Inall
period 3, all incubated
cells were cells werewith Ca2+ /HBS
incubated withcontaining
Ca2+/HBS
containing
20 nM CAP.20(B) nM InCAP. (B) Inset
a separate a separate set of experiments,
of experiments, TGNs were
TGNs were processed processed
as far as periodas 2far as period
before being
2lysed
before being
in 1x LDSlysed
buffer,inand
1x LDS buffer,
the ERK and the ERK was
phosphorylation phosphorylation
determined by was determined
Western blottingbyasWestern
detailed
blotting as detailed
in the Materials andinMethods.
the Materials and Methods.
(C) Histogram (C) Histogram
showing the ratio ofshowing the ratiofor
signal intensity of p-ERK
signal inten-
1/2 to
sity for p-ERK 1/2 to total ERK 1/2 (mean + s.e.m., n ≥ 3, N = 2), determined from the requisite im-
total ERK 1/2 (mean + s.e.m., n ≥ 3, N = 2), determined from the requisite immuno-reactive bands
muno-reactive bands detailed in panel B. (D) Spontaneous or NGF-induced CGRP release during
detailed in panel B. (D) Spontaneous or NGF-induced CGRP release during incubation period 2
incubation period 2 and (E) 20 nM CAP-evoked CGRP release in period 3 calculated by subtracting
andamount
the (E) 20 nM CAP-evoked
released CGRP release
during incubation in period
1, both 3 calculated
expressed as a % of bytotal
subtracting the amount
CGRP content; released
n = 9, N = 3.
during incubation 1, both expressed as a % of total CGRP content; n = 9,
For all histograms, one-way ANOVA was used followed by Bonferroni’s post hoc test, and signifi- N = 3. For all histograms,
one-way
cance ANOVA
indicated was
with used followed
asterisks; by Bonferroni’s
* p < 0.05, ** p < 0.01, ***post
p < hoc test,
0.001, ****and
pInt. J. Mol. Sci. 2022, 23, 892 9 of 20
evidenced by the reduction in the amount of intact SNAP-25 and the appearance of a faster-
migrating immuno-reactive band that was not observed in neurotoxin-free control cells
(Figure 5B, C). Relative to control starved cells, BoNT/A-treated TGNs displayed a small
increment in total CGRP content (Figure 5D) but this change was not significant (p = 0.22).
On the other hand, basal neuropeptide release from the cells without NGF was nearly two-
fold lower (Figure 5E, grey bars; 0.77 ± 0.11 vs. 0.40 ± 0.03% of total CGRP, mean ± s.e.m.;
p = 0.005) compared to toxin-free control counterparts, revealing that spontaneous CGRP
release largely entails SNAP-25-mediated exocytosis. Likewise, the NGF-induced elevation
of CGRP release was significantly (p = 0.0003) lower in BoNT/A-intoxicated neurons than
in toxin-free controls, confirming that this too arises from SNARE-dependent exocytosis
(Figure 5E, blue bars). As before, acute treatment of starved TGNs with NGF induced a
significant increment in the subsequent release evoked by 20 nM CAP (Figure 5F, c.f. control
grey and NGF-treated blue bar, p = 0.006). BoNT/A reduced both CAP-evoked CGRP
release from starved cells (Figure 5F, grey bars, p = 0.001) and its enhancement by acute treat-
Int. J. Mol. Sci. 2022, 23, x FOR PEER ment
REVIEWwith NGF (Figure 5F, blue bars, p = 0.005). By contrast, BoNT/A did not reduce CGRP10 of 21
release stimulated by 1 µM CAP (Figure 5G, grey bars), and the enhanced level induced
after pre-treatment with NGF was not lowered significantly (Figure 5G, blue bars).
Pre-treatment Release experiments
2 days 2 days 30 min 30 min 30 min
+ NGF - NGF HBS HBS/NGF HBS/CAP
± BoNT/A
Mr (k) BoNT/A
- +
37
syntaxin-1
25 SNAP-25
SNAP-25A
✱✱
✱✱ ✱✱✱
✱
✱✱✱
✱✱ ✱✱ ✱
✱✱
Figure 5. BoNT/A blocked NGF-induced CGRP release and -enhancement of 20 nM CAP-evoked CGRP
Figure 5. BoNT/A blocked NGF-induced CGRP release and -enhancement of 20 nM CAP-evoked
release. (A) After 2 days in the presence of 50 ng/mL NGF, TGNs were starved of the neurotrophin as
CGRP release. (A) After 2 days in the presence of 50 ng/mL NGF, TGNs were starved of the neuro-
trophin as detailed before, without or with the inclusion of 100 nM BoNT/A during this latter step.
The release experiment was performed as described in Figure 3A. (B) At the end of the protocol, one
well each of BoNT/A-treated and non-treated cells were solubilised in 1× LDS and subjected to West-
ern blotting. PVDF membranes were cut horizontally midway between the 25 k and 37 k molecular
weight markers. The upper portion was exposed to antibodies reactive with syntaxin-1 (mouse mon-Int. J. Mol. Sci. 2022, 23, 892 10 of 20
detailed before, without or with the inclusion of 100 nM BoNT/A during this latter step. The release
experiment was performed as described in Figure 3A. (B) At the end of the protocol, one well each of
BoNT/A-treated and non-treated cells were solubilised in 1× LDS and subjected to Western blotting.
PVDF membranes were cut horizontally midway between the 25 k and 37 k molecular weight markers.
The upper portion was exposed to antibodies reactive with syntaxin-1 (mouse monoclonal, 1:2000) and
the lower piece probed with an antibody recognising both intact and BoNT/A-truncated SNAP-25
(mouse monoclonal, 1:3000). (C) The amount of cleaved SNAP-25 in BoNT/A-treated cells was
calculated as a % (mean + s.e.m., N = 3) of the total SNAP-25 (intact + BoNT/A product). (D) Total
CGRP (pg/well). (E–G) Histograms showing: (E) spontaneous CGRP release during the second
30 min. incubation into HBS without (grey bars) or induced by 100 ng/mL NGF (blue bars), (F) during
the third period evoked by 20 nM CAP (minus the spontaneous release), and (G) during the third
incubation with 1 µM CAP minus the spontaneous release. In all cases, CGRP release is expressed as a
% of total CGRP (mean + s.e.m., N = 3, n = 9). Asterisks summarise the results of unpaired one-tailed
Welch tests applied to the data plotted in panels E, F, and G, * p < 0.05, ** p < 0.01, *** p < 0.001.
2.7. Chimera/EA Inhibits CGRP Release Elicited by High [CAP]
The/EA chimera was tested to ascertain whether it could cause a greater blockade than
Int. J. Mol. Sci. 2022, 23, x FOR PEER REVIEW 11 of 2
BoNT/A of CGRP release evoked by 1 µM CAP and its enhancement by acute NGF, using a
protocol identical to the one described above for BoNT/A (Figure 5A). TGNs were starved
of NGF and simultaneously incubated with 100 nM/EA before being briefly exposed to
NGF (or controlresulted in the cleavage
HBS) followed of more than
by stimulation with85% of the
1 µM CAP.SNAP-25
Exposurepresent
to/EA(Figure 6A, B), as re
resulted
flected by the notably faster-migrating product (SNAP-25
in the cleavage of more than 85% of the SNAP-25 present (Figure 6A, B), as reflectedE) than that produced b
by the notably BoNT/A (SNAP-25Aproduct
faster-migrating ; c.f. Figure 5B). As )found
(SNAP-25 for BoNT/A, total CGRP content wa
E than that produced by BoNT/A
slightly, but not significantly, increased in EA-treated TGNs (Figure 6C) but both sponta
(SNAP-25A ; c.f. Figure 5B). As found for BoNT/A, total CGRP content was slightly, but
neous CGRP exocytosis (Figure 6D, grey bars) and its elevation by NGF (Figure 6D, blu
not significantly, increased in EA-treated TGNs (Figure 6C) but both spontaneous CGRP
bars) were markedly reduced. Notably, pre-treatment with/EA reduced the 1 µM CAP
exocytosis (Figure 6D, grey bars) and its elevation by NGF (Figure 6D, blue bars) were
evoked CGRP release from NGF starved cells (Figure 6E, grey bars), unlike BoNT/A
markedly reduced. Notably, pre-treatment with/EA reduced the 1 µM CAP-evoked CGRP
Moreover, the enhancement of 1 µM CAP-evoked CGRP release observed in control cell
release from NGF starved cells (Figure 6E, grey bars), unlike BoNT/A. Moreover, the
after acute exposure to NGF (Figure 6E, control grey bar vs. control blue bar, p = 0.006
enhancement of 1 µM CAP-evoked CGRP release observed in control cells after acute
was absent in TGNs pre-treated with/EA. Consequently, this chimera clearly blocked re
exposure to NGF (Figure 6E, control grey bar vs. control blue bar, p = 0.006) was absent in
sponses to 1 µM CAP and its enhancement by NGF to a much greater extent than fo
TGNs pre-treated
BoNT/Awith/EA. Consequently,
(Figure 6E, c.f. Figurethis chimera
5G). These clearly
results blocked
confirm responses to 1 µM
the involvement of SNARE
CAP and its enhancement
dependent membrane trafficking in nociceptor sensitisation by NGF, a key6E,
by NGF to a much greater extent than for BoNT/A (Figure mediator o
c.f. Figure 5G).inflammatory
These results confirm the involvement of SNARE-dependent membrane
pain.
trafficking in nociceptor sensitisation by NGF, a key mediator of inflammatory pain.
/EA
Mr (K) - +
37
syntaxin-1
25 SNAP-25
SNAP-25E
✱✱✱
✱✱
✱ ✱✱✱✱
✱✱✱✱ ✱✱✱✱
Figure 6. Chimera/EA effectively inhibits 1 µM CAP-evoked CGRP release from starved TGNs, and
Figure 6. Chimera/EA effectively inhibits 1 µM CAP-evoked CGRP release from starved TGNs, an
its enhancement by NGF. TGNs were starved and incubated with 100 nM/EA using a protocol iden
tical to that described previously for BoNT/A (Figure 5A). (A) Western blotting with anti-SNAP-2
antibodies confirms the disappearance of intact SNAP-25 and the appearance of a much faster-m
grating product in cells exposed to/EA (+) but not control (−). (B) Histogram displaying the % oInt. J. Mol. Sci. 2022, 23, 892 11 of 20
its enhancement by NGF. TGNs were starved and incubated with 100 nM/EA using a protocol
identical to that described previously for BoNT/A (Figure 5A). (A) Western blotting with anti-
SNAP-25 antibodies confirms the disappearance of intact SNAP-25 and the appearance of a much
faster-migrating product in cells exposed to/EA (+) but not control (−). (B) Histogram displaying the
% of SNAP-25 cleaved, which was calculated as in Figure 5C. (C) Total amounts (pg/well) of CGRP,
determined as before, in control and/EA-treated cells. (D) Amounts of spontaneous CGRP release
(% total) into HBS only, (grey bars), and that during incubation with 100 ng/mL NGF (blue bars), and
(E) upon stimulation with 1 µM CAP (minus the spontaneous release) (mean + s.e.m. N = 3, n = 9).
Unpaired one-tailed Welch test was applied to the data plotted in panels (C–E), * p < 0.05, ** p < 0.01,
*** p < 0.001, **** p < 0.0001.
3. Discussion
Amongst the factors released during chronic inflammation that contribute to persis-
tent intransigent pain, NGF signalling has emerged as a prime candidate for therapeutic
interventions [3,4]. NGF-sequestering monoclonal antibodies showed promise, but clinical
trials had to be resumed with restricted dose protocols after serious adverse effects were
detected in the original tests, apparently, due to interference with NGF’s roles in bone
density maintenance and non-noxious sensation [3,4,43]. Thus, methods to mitigate NGF
signalling that selectively target its pain-promoting pathways are needed. Exposure of
sensory neurons to NGF causes an increase in their excitability and sensitivity to noxious
molecules [4,15–17,19,44], with an involvement of membrane trafficking of nociceptive
receptors to the neuron surface ([18,21]; Figure 7A). Moreover, the noxious chemical CAP
activates its unique receptor TRPV1 [8,9], and induces increases in [Ca2+ ]i [8,10] that trigger
the exocytosis from receptive neurons of pro-inflammatory neuropeptides, substance P and
CGRP ([37]; Figure 7A). In this regard, it is relevant to note that CGRP release from TGN
fibres that densely innervate the meninges is strongly implicated in migraine, albeit not
in all cases [45], and CAP is a recognised trigger of this painful condition (see Introduc-
tion). TRPV1-containing neurons are also clearly implicated in neurogenic inflammation,
which can be induced by CAP injection. On the other hand, repeated application of the
vanilloid prevents its induction of neurogenic inflammation due to targeted denervation of
CAP-sensitive neurons [46].
The results of this in vitro study provide new insights into sensitisation of neonatal
rat TGNs by NGF in relation to CAP-evoked CGRP exocytosis, as well as the different
abilities of/A and/E proteases to inhibit the process (see later). Exposing NGF-fed TGNs to
[CAP] from 20 to 100 nM yielded concentration-dependent increases in the amounts of both
[Ca2+ ]i (Figure 1A,B) and CGRP release (Figure 3B), which accords with reports of a four to
five fold increment in [Ca2+ ]i in FURA2-AM loaded cultures of neonatal rat DRGNs [41].
However, as reported by others using adult rat TGNs [47], further raising [CAP] to 300 nM
and 1 µM caused a progressive reduction in the amount of CGRP exocytosed relative to
the maximum level obtained with 100 nM (Figure 3B), unlike [Ca2+ ]i which continues to
accumulate in response to [CAP] up to 1 µM (Figure 1A,B) and as previously reported
for DRGNs [41]. This suggests that the fall-off in CGRP release at high [CAP] (0.3 to
1 µM) is due to a curtailment of the stimulation of exocytosis at the higher [Ca2+ ]i rather
than any reduction in the responsiveness to high CAP concentrations arising from TRPV1
desensitisation, for example.of of
sensory
of
sensory ious
sensory
neurons molecules
neurons
neurons
to to
NGFto
NGF [4,15–17,19,44],
NGF
causes
causes
causes
anan
increase
an with
increase
increase
in inan
their
in involvement
their
their
excitability ofand
excitability membrane
excitability
and sensitivity
and trafficking
sensitivity
sensitivity
to to
nox-
to
nox- of no
nox-
from TGN fibres that densely innervate the meninges is strongly implicated in migraine,
ious
ious
ious
molecules tive
molecules
molecules receptors
[4,15–17,19,44],
[4,15–17,19,44],to the
[4,15–17,19,44], withneuron
with
with
anan surface
involvement
aninvolvement ([18,21];
involvement of of Figure
membrane
of
membrane
membrane7A). Moreover,
trafficking
trafficking
trafficking
of of the
nocicep-
of noxious che
nocicep-
nocicep-
albeit not in all CAP casesactivates
[45], and CAP
itssurface
unique is areceptor
recognised
TRPV1 trigger
[8,9], of this
and painful
induces condition
increases in [Ca(see
2+]i [8,10
tive
tive
receptors
tive
receptors
receptors
to totheto
theneuron
the
neuron
neuron
surfacesurface
([18,21];
([18,21];
([18,21];
Figure
Figure
Figure
7A).7A). 7A).
Moreover,
Moreover,
Moreover, thethe
noxious
the
noxious
noxious chemical
chemical
chemical
xOR
PEER
FOR
PEER
PEER
REVIEW
REVIEW
REVIEW Introduction).
CAPCAPCAP
activates
TRPV1-containing
trigger
activates
activates
itsits
unique
itsthe
unique exocytosis
unique
receptor
receptor
neurons
receptor from
TRPV1
TRPV1
TRPV1
are
[8,9],
also
receptive
[8,9], 12
[8,9],
and and12
of clearly
neurons
12
of
21 induces
induces
and of
induces
implicated
ofincreases
21 21increases
increases
in in
in
pro-inflammatory
[Cain
[Ca
neurogenic
2+[Ca
]i2+[8,10]
]i2+[8,10]
[8,10]
that
that
in-
]ineuropeptides
that
flammation,
trigger
trigger
trigger which
thethe stance
exocytosis
the can
exocytosis
exocytosis befrom
P from
and induced
CGRP
from
receptive byneurons
([37];
receptive
receptiveCAP
Figure
neuronsinjection.
7A).
neurons
of of Inofthis On
pro-inflammatory the other
regard,
pro-inflammatory
pro-inflammatory hand, repeated
it isneuropeptides,
relevant to notesub-
neuropeptides,
neuropeptides, appli-
that
sub-CGRP r
sub-
Int. J. Mol. Sci. 2022, 23, 892 12 of 20
cation
stanceof
stance the
stance
P and vanilloid
P and from
PCGRP
andCGRPCGRPTGNprevents
([37]; fibres
([37];
([37];
Figure
Figure its
that
Figure
7A).induction
densely
7A).In
7A).
In
thisIn
this of
regard,
this neurogenic
innervate
regard, it isitthe
regard, relevant inflammation
ismeninges
isit relevant
relevant
to to
noteis strongly
to
note
note
that
that due
CGRP
thatCGRP CGRPto
implicatedtargeted
release
release in mig
release
of of
sensory
of
sensory
sensory
neurons
neurons
neurons
to to
NGFto
NGF denervation
NGF from
causes
causesfrom
causesfrom
ananTGN TGNof
TGN
increase
an CAP-sensitive
albeit
fibres
fibres
increase
increasefibres
in in that
their
innot
that
their in
densely
that
their all
densely neurons
excitabilitycases
densely
excitability
excitability
and[45],
innervate
innervate
and [46].
innervateand
the the
sensitivity
and CAP
meninges
the
sensitivity
sensitivityis
meninges
meninges
to to a
nox-torecognised
is
nox- strongly
is
nox-strongly
is stronglytrigger
implicated of
implicated
implicatedthis
in in painful
migraine,
in
migraine, conditio
migraine,
ious
ious
ious
molecules
molecules
molecules[4,15–17,19,44],
[4,15–17,19,44],
[4,15–17,19,44], albeit
with albeit
with albeit
with
an not
an not in
notallIntroduction).
in
involvement
an in
all
involvementcases
all
involvementcases
cases
of [45],
of [45],
membrane
of TRPV1-containing
[45],
and and
membrane CAP
and
membrane CAP CAP
is trafficking
ais recognised
trafficking
trafficking ofneurons
ais recognised
a of
recognised
nocicep-
of trigger
nocicep-
nocicep-areofalso
trigger
triggerof
this
ofclearly
this
painful
this implicated
painful
painful
condition
condition in(see
condition
(seeneurogen
(see
tive
tive
receptors
tive
receptors
receptors
to totheto
theneuron
theneuron
neuron Introduction).
surface Introduction).
surface Introduction).
surface
([18,21];
([18,21];
([18,21];flammation,
TRPV1-containing
Figure TRPV1-containing
Figure
Figure
7A). 7A). 7A). which
TRPV1-containing
Moreover,
Moreover, can
Moreover,neurons
the be
neurons
the induced
neurons
noxious
the are
noxiousare also
noxiousareby
also
chemical CAP
also
clearly
chemical
chemical injection.
clearly
clearly
implicated On
implicated
implicatedthe
in in other hand,
neurogenic
in
neurogenic
neurogenic repeated
in-in-in-
CAPCAPCAP
activates
activates
activates
itsits
unique
its
unique
unique
receptor flammation,
receptor flammation,
receptor
TRPV1flammation,
TRPV1
TRPV1 [8,9],cation
which
[8,9],which
[8,9],
and which
and of
can can
induces
and the
be
can
induces bevanilloid
induced
inducesbe
induced
induced
increases
increasesbyprevents
increasesby
in CAP by
inCAP
[Ca inCAP
[Ca
2+ its
[Ca
]i2+ induction
injection.
injection.
injection.
[8,10]
]i2+[8,10] On
]i [8,10]
that On
that of
the
On
that neurogenic
the
other
the
other
other
hand,
hand,inflammation
hand,
repeated
repeated
repeated
appli-due to tar
appli-
appli-
trigger
trigger
trigger
thetheexocytosis
theexocytosis
exocytosis from
from
from cation
receptivecation
receptive cation
receptiveofneurons
neuronsof
theof
thedenervation
vanilloid
the
neurons vanilloid
of vanilloid
of prevents
pro-inflammatory
of of
pro-inflammatory CAP-sensitive
prevents
prevents
pro-inflammatory itsits
induction
its
induction
neuropeptides, neurons
induction
neuropeptides,of of
neurogenic
neuropeptides, of [46].
neurogenic
neurogenic
sub-sub- inflammation
sub- inflammation
inflammationduedueto
dueto
targeted
to
targeted
targeted
stance
stance
stance
P and
P and
PCGRP
and CGRPCGRP
([37];
([37];
([37];
Figure
Figure denervation
Figure
7A).denervation
7A).denervation
In
7A).In
this In
this of
regard,
this of
CAP-sensitive
regard,of
CAP-sensitive
CAP-sensitive
regard,
it isit relevant
isit relevantneurons
is relevant neurons
to to neurons
noteto
note [46].
note
that [46].
that [46].
CGRP
thatCGRPCGRPrelease
release
release
from
from
from
TGNTGNTGN
fibres
fibres
fibres
thatthat
densely
that
densely
densely
innervate
innervate
innervatethethe meninges
the
meninges
meninges is strongly
is strongly
is stronglyimplicated
implicated
implicated in inmigraine,
in
migraine,
migraine,
albeit
albeit
albeit
notnot
in
notin
allin
all
cases
all
cases
cases
[45],
[45],
[45],
andandCAP
and CAPCAP
is ais recognised
ais recognised
a recognised trigger
trigger
trigger
of of
this
of
this
painful
this
painful
painfulcondition
condition
condition (see(see
(see
Introduction).
Introduction).
Introduction). TRPV1-containing
TRPV1-containing
TRPV1-containing neurons
neurons
neurons areare also
are
also
also
clearly
clearly
clearly
implicated
implicated
implicated in in neurogenic
in
neurogenic
neurogenic in-in-in-
flammation,
flammation,
flammation, which
which
which
cancan be
canbe
induced
be
induced
induced
byby CAPby
CAP CAP
injection.
injection.
injection.
OnOn the
On the
other
the
otherother
hand,
hand,hand,
repeated
repeated
repeatedappli-
appli-
appli-
cation
cation
cation
of of
theof
the
vanilloid
the
vanilloid
vanilloid
prevents
prevents
prevents
itsits
induction
its
induction
inductionof ofneurogenic
of
neurogenic
neurogenic inflammation
inflammation
inflammation due dueto
due to
targeted
to
targeted
targeted
denervation
denervation
denervation of of
CAP-sensitive
of
CAP-sensitive
CAP-sensitive neurons
neurons
neurons
[46].
[46].[46].
Figure 7. NGF induces a minor increase in Ca 2+- and SNARE-dependent CGRP release, whereas it
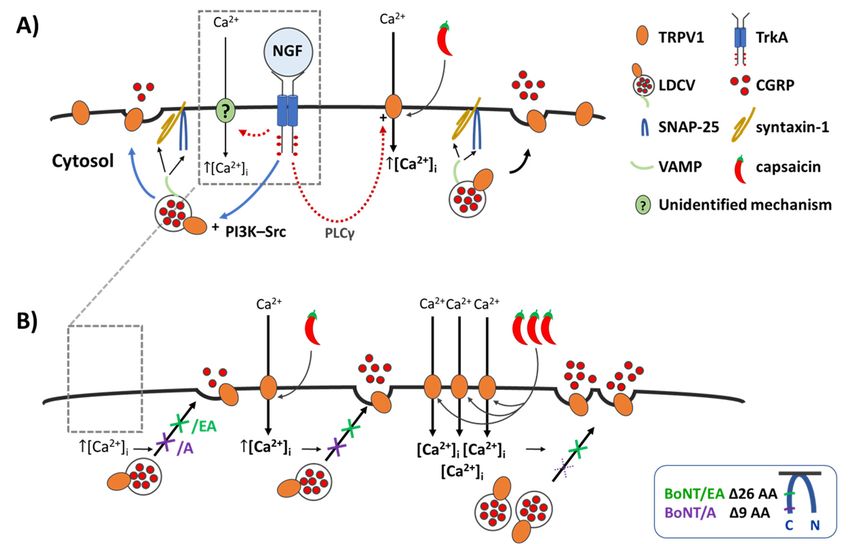
Figure 7. NGF induces a minor increase in Ca2+ - and SNARE-dependent CGRP release, whereas it
greatly enhances the CAP-evoked exocytosis which is blocked by BoNT/A and/EA at low [CAP] but
greatly enhances the CAP-evoked exocytosis which is blocked by BoNT/A and/EA at low [CAP]
only abolished byFigure BoNT/EA 7. NGF at induces
higher [CAP]. a minor (A) Illustrates
increase in Ca2+the - and effect of acute NGF on
SNARE-dependent CGRP exo- whe
but only abolished by BoNT/EA at higher [CAP]. (A) Illustrates the effect of acute NGF CGRP on CGRP release,
cytosis
Figure
from
Figure
Figure
7. NGF
control
7.from
NGF
7. NGFgreatly
induces
neonatal
induces enhances
induces
aneonatal
minor
rat
a minor
a minor
TGNs
the
increase
starved
CAP-evoked of the
exocytosis neurotrophinwhich is blocked for 2 days,
by and
BoNT/A (B) in
and/EATGNs pre-
itat it
low it [CA
exocytosis control ratincrease
increase
TGNs instarved
in
Cain Ca
2+- Caand
2+ -ofand
2+ -the
SNARE-dependent
and SNARE-dependent
SNARE-dependent
neurotrophin forCGRP
2 CGRP CGRP
days, release,
release,
and release,
(B)whereas
inwhereas
whereas
TGNs
treated
greatly with
greatly
greatly BoNT/A
enhances
enhances only
enhances thethe or /EA.
abolished
CAP-evoked
the
CAP-evoked (A)
CAP-evokedby NGF
BoNT/EA
exocytosis binds
exocytosis
exocytosis atto
which its
higher
which receptor
[CAP]. TrkA,
(A) activates
Illustrates the the signalling
effect of acute cascades
NGF
butbutbutCGR
on
pre-treated with BoNT/A or /EA. (A) NGF binds to which
itsisreceptor
blocked
is blocked
is blockedby by
TrkA, BoNT/A
by
BoNT/A
BoNT/A
activates and/EA
and/EA
theand/EA at low
at low
signalling at[CAP]
low[CAP]
cascades[CAP]
shown
only only [18],
abolished
only and by
abolished
abolished induces
cytosis
by
BoNT/EA
by Ca2+
from
BoNT/EA
BoNT/EA atinflux
control
higher
at higher
at by
neonatal
higher an
[CAP]. unidentified
[CAP]. rat
[CAP].
(A) TGNs
(A)
Illustrates
(A)Illustratesmechanism
starved
Illustrates
the of
the the
effect
the
effect (?).
effect
of of Elevated
neurotrophin
acuteof
acute
acute
NGF for
NGF NGF
onintracellular
2 ondays,
CGRPon
CGRP and
CGRP
exo- exo- Ca
(B)exo-
2+ TGN
in
shown [18], andtreated induceswith Ca2+BoNT/A influx by or an
/EA. unidentified
(A) mechanism (?). Elevated intracellular Ca 2+
([Ca ]i)cytosis
triggers thecontrol
fusion ofrat large dense core of NGF
vesicles binds to its receptor
(LDCVs) forvia TrkA,
2SNARE-complexesactivates the signalling
(VAMP, pre- ca
2+
cytosis
cytosis from from from
control
control neonatal
neonatal
neonatal rat
TGNs rat
TGNs TGNs
starved
starved
starved the
of the
ofneurotrophin
the
neurotrophin
neurotrophin for
2 days,
for days,
2 days,
and and(B)
and (B)
in(B)TGNs
in TGNs
in TGNs
pre- pre-
([Ca2+ ]i ) triggers the fusion
shown of large dense core vesicles (LDCVs) via SNARE-complexes (VAMP,
or[18], and induces Ca influx by an unidentified mechanism (?). Elevated intracellula
2+
syntaxin-1
treated
treated
treatedand
with with SNAP-25),
BoNT/A
with BoNT/A
BoNT/A orthereby,
/EA.or
/EA. (A)
/EA. (A)NGFcausing
(A)NGF NGF
binds exocytotic
bindsbinds
to its
to itsto
receptor
its release
receptor
receptor
TrkA, ofTrkA,
TrkA, CGRP
activates
activatesand
activates surface
thethesignalling
the
signallingdelivery
signalling cascades of
cascades
cascadesves-
iclesyntaxin-1 and SNAP-25),
([Ca thereby, causing exocytotic release of CGRP and surface delivery of vesicle
2+]i) triggers
constituents.
shownshownshown
[18],[18],and
[18], This
and induces
and acute
induces
induces 2+ the
Capotentiation
2+ influx
Ca Ca influx
2+ fusion
influx
by by anby anofan large
NGF
unidentified
unidentified dense
can
unidentified involvecore
mechanism
mechanismvesicles
the (?).
mechanism (LDCVs)
phosphatidylinositol
(?).
Elevated
(?).
Elevated
Elevatedvia SNARE-complexes
intracellular
intracellular 3-kinase—
intracellular CaCa 2+ Ca2+ 2+ (V
constituents.
Src([Ca ([Ca
2+ ]i) ]triggers
([Ca
(PI3K-Src) 2+ 2+ This
i) ]triggers
syntaxin-1
i)pathway,
triggers acute
thethe fusion
thefusion
which and
potentiation
fusion
of ofSNAP-25),
largeof
large
promotes by
large
dense NGF
dense thereby,
dense
core can
core
trafficking causing
involve
vesicles
core vesicles
vesicles exocytotic
the
(LDCVs)
of LDCVs, (LDCVs)
(LDCVs) viavia
and release
phosphatidylinositol of
SNARE-complexes
via CGRP
SNARE-complexes
insertionSNARE-complexes and
3-kinase—Src
of their TRPV1 surface
(VAMP,(VAMP,
(VAMP, delivery
chan-
Figure
Figure
Figure
7. NGF7. NGF
7. NGF
induces
induces
induces
a minor
a minor
a minor
increase
increase
increase
in
(PI3K-Src)
syntaxin-1 in
Cain
syntaxin-1 Ca - Ca
and
2+pathway,
syntaxin-1 and2+
and icle
- 2+
and
SNAP-25),
and constituents.
-SNARE-dependent
and SNARE-dependent
SNARE-dependent
which
SNAP-25),
SNAP-25), promotes
thereby,
thereby, This
thereby, acute
CGRP CGRP CGRP
trafficking
causing
causing
causing potentiation
release,
release,
exocytotic release,
of
exocytotic whereas
LDCVs,
exocytotic by
whereas
release
releaseNGF
whereas
and
release CGRP
of can
itof itarrows)
itCGRP
insertion involve
of CGRP and thesurface
ofsurface
and their
and phosphatidylinositol
TRPV1
surface delivery
deliverychannels
deliveryof ves-
of ves-
of ves- 3-ki
nels into the plasmalemma by Ca 2+ -regulated exocytosis (blue c.f. [18,21]. Additionally, the
greatly
greatly
greatly
enhances
enhances
enhances
thethe
CAP-evoked
the
CAP-evoked
CAP-evoked exocytosis
exocytosis
into exocytosis
icleicle the whichwhich
plasmalemma
constituents.
icle
constituents. Src
which
is
constituents. (PI3K-Src)
blocked
is blocked
is blocked
by
byacute
Ca by pathway,
2+
BoNT/A
by
BoNT/A
BoNT/A
-regulated and/EAwhich
and/EA
and/EAatpromotes
exocytosislow
at low
at[CAP]
low trafficking
[CAP]
(blue [CAP]
but but
arrows) but of LDCVs,
c.f. [18,21]. on and insertion
Additionally, of their
the TRPV1
phospholipase C γThis This
(PLCγ) acute
This acute
potentiation
cascade potentiation
potentiation
leadsbyto byNGFbyNGF NGF
cancan
sensitisation involve
caninvolve
involve
of the
TRPV1thephosphatidylinositol
the
phosphatidylinositol
phosphatidylinositol
already the 3-kinase—3-kinase—
plasmalemma3-kinase—
only
onlyabolished
only abolished
abolished
by by
BoNT/EA
by
BoNT/EA
BoNT/EA
at higher
at higher
atphospholipase
higher
Src [CAP].
Src [CAP].
[CAP].
(PI3K-Src)
Src (A)(A)
(PI3K-Src)
(PI3K-Src) nels
Illustrates
(A)
pathway,
C into the
Illustrates
Illustrates
pathway,
γ pathway,
(PLCγ) thethe
which plasmalemma
effect
whichtheeffect
which
cascade effect
of leads
promotes of
acute
promotes of
acute
promotes by
acute
NGF Ca
NGF
trafficking
to
2+-regulated exocytosis (blue arrows) c.f. [18,21]. Additional
NGF
on
trafficking onCGRP
trafficking
sensitisation ofonCGRP CGRP
LDCVs,
of exo-
LDCVs,
of
of exo-
LDCVs,
TRPV1exo-
and andinsertion
andinsertion
already insertion
of
on their
of
the their
of their
TRPV1 TRPV1
plasmalemma TRPV1 chan-chan- chan-
(red dashed arrows) [18]. The outcome
phospholipase Cby γ (PLCγ)
of these cascade
compositeleads to
influences
sensitisation
of NGF ofc.f.
on TRPV1
TRPV1 already
is that
onthe
when
the plasmal
cytosis
cytosis
cytosis
from from from
control
control
control
neonatal
neonatal
neonatal
ratrat
TGNs
rat
TGNsTGNs
nels
(red starved
nels starved
into
nels starved
into
dashed the of
into
thethe
of the
ofneurotrophin
plasmalemma
the the
neurotrophin
plasmalemma
arrows) neurotrophin
plasmalemma
[18]. by
The byfor
Ca for
Ca
2+2-regulated
outcome days,
for
Ca
2+2-regulated
days,
2+ 2-regulated
days,
and
of and(B)
and(B)
in
exocytosis
these (B)
TGNs
in TGNs
exocytosis in(blue
exocytosis
composite TGNs
pre-pre-
(blue pre-
(blue
arrows)
arrows)
influences arrows)
c.f. c.f.
of [18,21].
[18,21].
NGF [18,21].
onAdditionally,
Additionally,
TRPV1Additionally,
is that the the
the channel is activated (red by CAP
dashed arrows)[Ca [18].
2+ ]I is The raised even more
outcome of cascades
these than normally
composite [15,18,21]
influences of and this
NGF on TRPV1further is that
treated
treated
treated
with with BoNT/A
with
BoNT/A
BoNT/A
or or
/EA.
or
/EA.
(A)
/EA.
(A)NGF
(A)
NGF NGF
bindsbinds
phospholipase binds
phospholipaseto its
phospholipaseto its
to
receptor
C its
receptor
γC receptor
(PLCγ)
γ
C TrkA,
(PLCγ)
γ TrkA,
(PLCγ) TrkA,
activates
cascade activates
cascade activates
cascade
leads theleads
leads the
to
2+ signalling
the
tosignalling
signalling
sensitisation
to
sensitisationcascades
sensitisation ofcascades
of
TRPV1of
TRPV1 TRPV1already
already
already
on on
theontheplasmalemma
theplasmalemma
plasmalemma
when
enhances the CGRPchannel releaseis activated
(Figure by2). CAP(B) [Ca
The ] is
proteases
I raised2+]of even
BoNT/A more than
2+ and/EA
normally
delete [15,18,21]
9 (purple and this
arrow)and andthis f
shown
shown
shown[18], [18],and
[18],
andinduces
and
induces
induces 2+ influx
CaCa 2+ influx
Ca 2+ influx
by(red
(red by
an byan
unidentified
dashed
(red an
dashed
dashed the
unidentified
unidentified
arrows)
arrows) channel
mechanism
arrows) mechanism
[18].[18].
The is
mechanism
[18].
The activated
(?).
outcome
The (?).
outcome Elevated
(?).
outcome ofbythese
Elevated
ofCAP
Elevated
these
of [Ca
intracellular
these
composite
compositeI is raised
intracellular
intracellular
composite 2+ even
Cainfluences
Ca
influences Ca 2+ more than
influences of NGF
of NGF
of NGF
on normally
on
TRPV1
on
TRPV1TRPV1is [15,18,21]
that
is that
is when
that
when when
26 further
(green enhances
arrow) CGRP release
residues
enhances from (Figure
SNAP-25 2). (B) The
(Insert), proteases
respectively, of BoNT/A
preventing and/EAtheand/EA delete
fusion of9 LDCVs;
(purple this
([Ca
([Ca ]i)2+]triggers
2+([Ca i)2+]triggers
i) triggers
thethe
fusion
thefusion
fusion
of of
large
of
large
large
dense
the dense
the dense
channel
the core
channelcore
channelvesicles
core
is vesicles
vesicles
activated
is activated
is activated byCGRP
(LDCVs)
(LDCVs)
(LDCVs)
by
CAP by via
CAP release
CAP
[Cavia[Ca
2+ [Cais(Figure
SNARE-complexes
via
]SNARE-complexes
I2+ raised
is]Iraised
I2+
2).
]SNARE-complexes
is raised
even(B)
even The
even
more(VAMP, proteases
(VAMP,
more (VAMP,
more
thanthan normally
than ofnormally
BoNT/A
normally [15,18,21]
[15,18,21]
[15,18,21]
and and delete
this
and this 9further
further
this (purple
furtherarrow
syntaxin-1
syntaxin-1
syntaxin-1 and and
SNAP-25),
and
SNAP-25),
SNAP-25),
thereby,
thereby, arrow)
blocks
thereby,
causing the
causing and
causing minimal26
exocytotic
exocytotic(green
26
exocytotic CGRP
(green
release arrow)
release
release
of residues
exocytosis
arrow)CGRP
of residues
CGRP
of from
elicited from SNAP-25
by NGF
SNAP-25 (Insert),
(arrow
(Insert), respectively,
with crosses,
respectively, preventing
left) and
preventing its the
the fusion
enhancement
fusion of LDCV
enhances
enhances
enhances CGRP CGRP CGRP
release
release
release
(Figure
(Figure 2).CGRP
(Figure and
2).
(B) 2).and
(B)
Thesurface
and
(B)
Thesurface
surface
proteases
The delivery
proteasesdelivery
proteasesdelivery
of ofofBoNT/A
ofves-
BoNT/A ofBoNT/A
ves-
of and/EA
ves- and/EA
and/EAdelete
delete
delete
9 (purple
9 (purple
9 (purplearrow)
arrow)arrow)
and and and
icleicle
constituents.
icle
constituents.
constituents.
ThisThis
acute
This
acute
acute
potentiationof
theLDCVs;
potentiation
potentiation
of by (green
by
release NGF this
by
NGF blocks
blocks
NGF
can
evokedcan
involve
can the
the
involve
by minimal
minimal
involve
20the the
nM CGRP
phosphatidylinositol
the exocytosis
phosphatidylinositol
CAP phosphatidylinositol
( (Insert),
) (arrow elicited
elicited
3-kinase—
3-kinase—
with by NGF
by
3-kinase— NGF(arrow
crosses, (arrowwith
centre). with
Strongercrosses,
crosses, left)and
left) andits enhanc
26 26
(green
26
(green arrow)arrow)
arrow)
residues
residues
residues
from from from
SNAP-25
SNAP-25
SNAP-25 (Insert),
(Insert),
respectively,
respectively,
respectively, preventing
preventing
preventingthethe fusion
the
fusion
fusion ofstimulation
of LDCVs; LDCVs;
of LDCVs; of
thisthisthis
SrcSrc
(PI3K-Src)
Src
(PI3K-Src)
(PI3K-Src)
pathway,
pathway,
pathway,which
which
which
promotes
promotes
promotes
its
blocks trafficking
blocks trafficking
enhancement
blocks
the trafficking
the
minimal
the of
of of
minimal LDCVs,
of
the
the
minimal
CGRPLDCVs,
of LDCVs,
release
release
CGRP CGRP and and
insertion
and
evoked
evoked
exocytosis
exocytosis insertion
exocytosis insertion
byby
elicited of20
20
elicited their
of nM
nM
elicited
by their
of
by
NGF their
TRPV1
CAP
CAP
by
NGF TRPV1
NGFTRPV1
(
(arrow chan-
(arrow)
) chan-chan-
(arrow
(arrow
(arrow
withwith with
with
crosses,
with
crosses, crosses,
crosses,
crosses,
left) left)
and centre).
left)
andcentre).
its
and its Stronger
enhancement
its Stronger
enhancement
enhancement stimulat
TRPV1 with 1 µM CAP ( ) induces a lot more Ca influx ([Ca ]i, ([Ca ]i, ([Ca ]i; Figure 1)
2+ 2+ 2+ 2+
nelsnels
into
nels
into
the
into
the
plasmalemma
the
plasmalemma
plasmalemma by by
Caby2+-regulated
Ca 2+-regulated
Ca 2+-regulated
stimulation exocytosis
exocytosis
exocytosis
TRPV1
ofmoderate
TRPV1 (blue
(blue
with
with (blue
arrows)
arrows)
120
201by arrows)
µM c.f.
CAP
CAP c.f.
[18,21].
(c.f.
([18,21].
( [18,21].
Additionally,
Additionally,
Additionally,
)(arrow
induces
)) induces a the
alotthe
lot the
more
more Ca
Ca 2+2+ influx
influx ([Ca
([Ca 2+ ([Ca2+2+
2+]]i, ,([Ca ](purple
i,of
]([Ca 2+]i; Fig
whichof of the of
the
release
causes the
release
arelease
evoked
evoked
evoked
by by
increase µM
nM 20
nM CAP
innM CAP
CGRP CAP )release
((arrow
) (arrow with
but with with
crosses,
overcomes crosses,
crosses,
centre).
centre).
the centre).
Stronger
inhibition Stronger
Stronger
bystimulation
stimulation
i stimulation
BoNT/A i , of of
phospholipase
phospholipase
phospholipase Cγ C (PLCγ)
γ
C (PLCγ)
γ (PLCγ)
cascade
cascade
cascade
leadsleads leads
to to
sensitisation
to
sensitisation
2+ ] ; Figure 1) sensitisation
which of of
TRPV1
causes of
TRPV1
a TRPV1already
moderate already
already
on on
increasethe ontheplasmalemma
the
in plasmalemma
plasmalemma
CGRP release but overcomes the inhibition by BoNT/A
cross([Ca
TRPV1 with
TRPV1TRPV1 broken
i with which
lines, causes
right) a
( (while/EA moderate increase
(green a cross, in CGRP
right) release
Caremains but overcomes
effective ]iin the inhibition
]idiminishing byCGRP1) 1) (
of with with
1composite
µM
1composite
µM
1CAP
µM CAP CAP
( influences of)lines,
induces
)NGF
induces
)NGF
inducesa onlot lot
more
a islot
moreismore
Ca Ca
2+ influx
2+ influx
2+ influx
([Ca
([Ca
2+([Ca
]i, ([Ca
2+ ]i, ([Ca
2+ ]i, ([Ca
2+ , ([Ca
2+ , ([Ca
2+ ]i, ([Ca
2+ ]i; Figure
2+ ]i; Figure
2+ ]i; Figure
1)
(red(red
dashed
(red
dashed
dashed
arrows)
arrows)
arrows)
[18].
[18].
The
[18].
Theoutcome
Theoutcome
outcome of these
of these
these
compositecross influences
withinfluences
broken NGF
of of on
right)on
TRPV1 TRPV1 TRPV1
while/EA that that
is when
that
(green whenwhen
cross, right) remains effective in diminishing
BoNT/A
which whichwhich (purple cross with broken in lines, right) while/EA (green cross, right) remains effective in
]causes
iscausescauses
aeven
moderate
aeven
moderate
amore
moderate increase
increase
increase CGRP
in CGRP
in CGRP
release
release
release
but but
overcomes
butovercomes
overcomesthethe
inhibition
the
inhibition
inhibitionby by BoNT/A
by
BoNT/A
BoNT/A (purple
(purple
(purple
thethe
channel
the
channel
channel
is activated
is activated
is activated
by byCAPby
CAPCAP
[Ca[Ca2+ ][Cais
I2+ ]I2+
raised
is Iraised
raised
even more more
than than normally
than normally
normally [15,18,21]
[15,18,21]
[15,18,21] and and this
and this
further
this
further
further
cross cross
diminishingcross
with withbroken
with broken
CGRP broken
lines,
lines,
lines,
release.right)
right)
Acuteright)
while/EA
while/EA
while/EA
sensitisation(green(green(green
bycross,cross,
NGF cross,
right)
of right)
right)
TRPV1remains
remains
remains
effective
effective
selectively effective
in diminishing
enhances in diminishing
in diminishing
neuropeptide CGRPCGRP CGRP
enhances
enhances
enhances
CGRPCGRPCGRP
release
release
release
(Figure
(Figure
(Figure
2). 2).
(B)2).
(B)
The (B)
The proteases
The proteases
proteasesof BoNT/A
of BoNT/A
of BoNT/A and/EAand/EA
and/EA
deletedelete
delete
9 (purple
9 (purple
9 (purplearrow)arrow)arrow)
and and and
26 26
(green
26
(green
(green
arrow)
arrow)
arrow)
residues
residues
residues
fromfromfrom
SNAP-25 exocytosis
SNAP-25
SNAP-25 (Insert), stimulated
(Insert),
(Insert),
respectively, by
respectively,
respectively,low [CAP]
preventing
preventing
preventing (You can also read

















































