The induction of multiple cell cycle events precedes target-related neuronal death
←
→
Page content transcription
If your browser does not render page correctly, please read the page content below
Development 121, 2385-2395 (1995) 2385
Printed in Great Britain © The Company of Biologists Limited 1995
The induction of multiple cell cycle events precedes target-related neuronal
death
Karl Herrup* and Jonathan C. Busser
Alzheimer Research Laboratory, Department of Neurology, Case Western Reserve Medical School, 10900 Euclid Avenue,
Cleveland, OH 44106, USA
*Author for correspondence: e-mail: kxh26@po.cwru.edu
SUMMARY
Unexpected nerve cell death has been reported in several D, proliferating cell nuclear antigen and bromodeoxyuri-
experimental situations where neurons have been forced to dine incorporation – are elevated in the granule cells before
re-enter the cell cycle after leaving the ventricular zone and they die. Although lurcher Purkinje cells die during a
entering the G0, non-mitotic stage. To determine whether similar developmental period, no compelling evidence for
an association between cell death and unscheduled cell any cell cycle involvement in this instance of pre-pro-
cycling might be found in conjunction with any naturally grammed cell death could be found. While application of
occurring developmental events, we have examined target- the TUNEL technique (in situ terminal transferase end-
related cell death in two neuronal populations, the granule labeling of fragmented DNA) failed to label dying granule
cells of the cerebellar cortex and the neurons of the inferior cells in either mutant, light and electron microscopic obser-
olive. Both of these cell populations have a demonstrated vations are consistent with the interpretation that the death
developmental dependency on their synaptic target, the of these cells is apoptotic in nature. Together, the data
cerebellar Purkinje cell. Two mouse neurological mutants, indicate that target-related cell death in the developing
staggerer (sg/sg) and lurcher (+/Lc), are characterized by central nervous system is associated with a mechanism of
intrinsic Purkinje cell deficiencies and, in both mutants, cell death that involves an apparent loss of cell cycle
substantial numbers of cerebellar granule cells and inferior control.
olive neurons die due to the absence of trophic support
from their main postsynaptic target. We report here that Key words: mammalian CNS, cell cycle, cell death, cerebellum,
the levels of three independent cell cycle markers – cyclin lurcher, staggerer, neuronal death
INTRODUCTION death (see refs in Oppenheim et al. (1990) and Williams and
Herrup (1988)). It may seem paradoxical for cell death to be a
During the development of the nervous system, the final normal component of the growth of the nervous system, but
number of nerve cells in any one adult structure represents the target-related cell death is now appreciated as an important part
combined effects of additive processes such as cell division of the developmental program. Indeed, in some of the best
and subtractive processes such as cell death. Cell division studied neuronal populations, over 50% of the originally
typically occurs in the ventricular and subventricular zones – generated neurons normally die. These deaths are referred to
transient, specialized cell layers lining the inner surface of the as target-related because the extent of the neuron loss is a
developing neural tube. At specific times during embryogene- function of the access of the cells to their target cell popula-
sis, groups of young neuroblasts leave these zones and begin tion (be it muscle, gland or other neurons). If the target is
migration to their adult position in the nervous system. Once experimentally removed before or during a well-defined
they migrate from the ventricular zone, they leave the cell cycle critical period, the normal amount of nerve cell loss is
and enter a postmitotic state (known as G0) that persists increased.
throughout their phenotypic maturation. While the molecular Work from many laboratories has shown that these devel-
and biochemical mechanisms that serve to maintain this opmental cell deaths are not passive ‘witherings’ of malnour-
quiescent mitotic state are only beginning to be understood, ished cells, but rather the result of an active process that
they must be exceedingly reliable since CNS cancers of involves the recruitment of entire ensembles of genes for its
neuronal origin are virtually unknown. completion. For example, the target-related cell death that
Perhaps because there is little or no ability for an organism occurs in the developing chick lateral motor column can be
to add nerve cells later, many neuronal cell populations adjust blocked by agents that interrupt neuromuscular activity
their size by a process that involves overproduction early in (Pittman and Oppenheim, 1978), or RNA or protein synthesis
development followed by elimination of excess neurons by cell (Oppenheim et al., 1990). Similarly, the loss of cultured sym-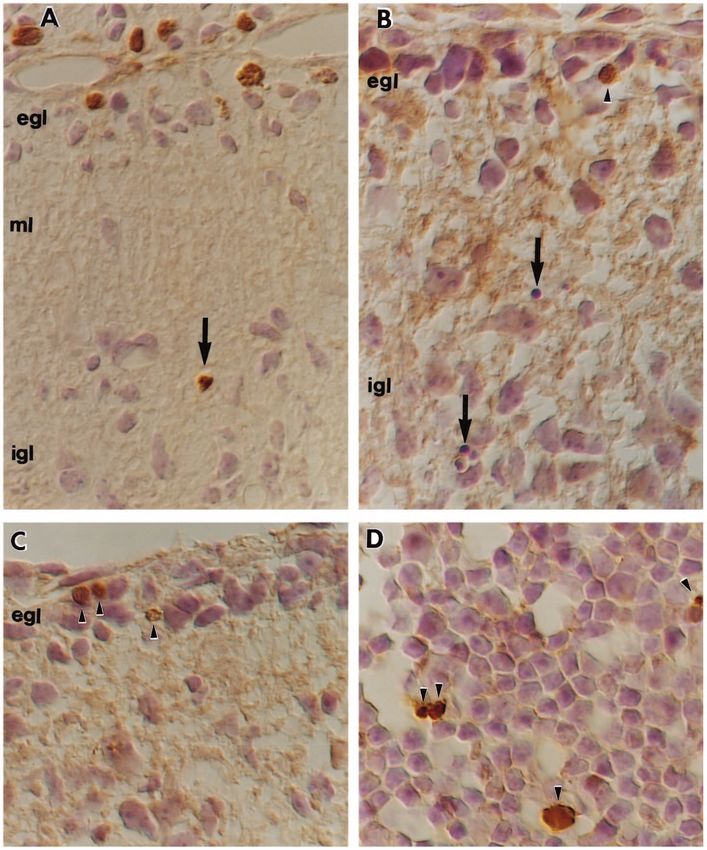
2386 K. Herrup and J. C. Busser
pathetic neurons that occurs in response to withdrawal of NGF division and cell death in the nervous system has been provided
can be stopped by the addition of drugs that block protein or by Freeman et al. (1994) who showed that the death of cultured
mRNA synthesis (Martin et al., 1988; Scott and Davies, 1990). sympathetic neurons, triggered by NGF withdrawal, leads to
The specific genes that are involved in the cell death process the induction of cyclin D1 mRNA (a G1/S phase marker). This
are beginning to be identified in both vertebrate and inverte- last observation suggests that the linkage between unscheduled
brate models. In the nematode C. elegans, the regulation of cell division and cell death might apply to other trophic factor-
developmental cell death has been associated with the activity sensitive cell deaths during normal nervous system develop-
of nearly a dozen genes (see Driscoll and Chalfie, 1992, for ment.
review), and recent work suggests that the homologous To explore this possibility, we have examined two neuro-
members of the vertebrate genome will soon be known as well logical mutations of the mouse, staggerer (sg/sg) and lurcher
(Boise et al., 1993; Gagliardini et al., 1994; Miura et al., 1993; (+/Lc). Both mutations cause highly predictable cell losses in
Reed, 1994; Smeyne et al., 1993; Yuan et al., 1993; Zhong et the olivocerebellar circuit (Caddy and Biscoe, 1979; Herrup
al., 1993). and Mullen, 1979a; Zanjani et al., 1990) and the sites of gene
Programmed cell deaths such as these tend to proceed in a action have been established for both mutations through the
stereotyped manner, and, when certain characteristic steps are analysis of aggregation chimeras (Herrup, 1983; Herrup and
involved, the process is called apoptosis (Bowen, 1993; Mullen, 1979b, 1981; Wetts and Herrup, 1982a,b; Zanjani et
Schwartz and Osborne, 1993; Schwartzman and Cidlowski, al., 1990). In staggerer, 100% of the cerebellar granule cells
1993). Specifically, in apoptotic cell death, the cell nucleus die due to an absence of trophic support from their postsynap-
condenses and darkens and the cytoplasmic membrane blebs. tic targets, the Purkinje cells. In lurcher, 100% of the Purkinje
These changes occur in advance of any overt changes in the cells die a preprogrammed, cell-autonomous death that in turn
cytoplasmic organelles. Finally, an important signature of the leads to the death of 90% of the granule cells. Purkinje cell
apoptotic process is the early enzymatic fragmentation of the deficiencies in both mutants also cause the loss of substantial
chromosomal DNA into a ‘ladder’ of gel fragments, with each numbers of neurons in the inferior olive. One distinctive dif-
‘rung’ 180 bp larger than the previous. This pattern is inter- ference between the two mutations lies in the phenotypes of
preted as representing the cleavage of chromosomal DNA into the Purkinje cells. In the staggerer, the Purkinje cells never
nucleosome-size units. fully develop, either biochemically (Messer et al., 1990, 1981)
The processes of cell division and cell death would seem to or morphologically (Landis and Sidman, 1978; Sidman, 1968,
be polar opposites, both in terms of their functions and their 1977). The tertiary branchlet spines that are the normal site of
developmental regulation, yet several recent lines of evidence synapse between the granule cell axon (the parallel fiber) and
have raised the possibility that the two processes may be mech- the Purkinje cell dendrite never form. Because of this, granule
anistically related. Part of this evidence comes from the cells never establish adequate contact with their target and this
analysis of engineered mutations in the mouse tumor suppres- leads to their death. In the lurcher, the Purkinje cells develop
sor gene, retinoblastoma (Clarke et al., 1992; Jacks et al., 1992; normally until around the end of the first postnatal week. The
Lee et al., 1992). The normal retinoblastoma protein (RB) has young +/Lc Purkinje cell dendrites do develop spines, but on
been shown in a number of studies to arrest the cell cycle in about postnatal day 7 (P7) their further morphological (Caddy
late G1 (Goodrich et al., 1991). Retinoblastoma-deficient and Biscoe, 1979; Dumesnil-Bousez and Sotelo, 1992) and
mouse embryos do not survive beyond the 16th embryonic day biochemical (Messer et al., 1991; Tano et al., 1992) develop-
and one prominent phenotype of the dying embryos is the ment is arrested, and they begin to die (100% of them are lost
occurrence of massive amounts of apoptotic cell death in the by the end of the second postnatal month). Granule cell loss
CNS just outside of the ventricular zone. The observation of quickly follows though it is incomplete (10% of the normal
ectopic mitotic figures in the same regions as the dying cells population survives in the adult mutant). These mutants thus
led Lee et al. (1992) to hypothesize that once a neuroblast offer examples of two distinct types of cell death: one caused
finishes its normal period of cell division, it commits itself to by a cell-extrinsic failure of trophic support (the death of
a program of maturation that includes the cessation of further granule cells and inferior olive neurons in both mutants), the
cell division. If the control of this process breaks down (as other a cell-autonomous, preprogrammed death (the loss of
might be expected in the absence of a functional RB protein), Purkinje cells in lurcher). We report here that in both mutants
the cell may re-enter the cycle, but, if it does so, it dies. target-related cell death is accompanied by evidence for a re-
The suggestion that cell cycle arrest is important for the entry into the cell cycle.
stable maturation of CNS neurons receives further support
from the analysis of several lines of transgenic mice. SV40 T-
antigen exerts its action in part by sequestering RB thus neu- MATERIALS AND METHODS
tralizing its cell cycle blockade and allowing cell division to
proceed. In opsin:T-antigen transgenic mice, expression of the Identification of mutant genotype
transgene during the differentiation of the photoreceptors leads We identified mutant staggerer animals in litters from +/sg × +/sg
to the induction of DNA synthesis. Rather than proceeding matings based on their ataxia. Animals ranging in age from P10 to
P22 were used. The mutant animals from +/Lc × +/+ matings were
through cell division, however, the photoreceptors die (al- identified at ages from P12 to P30 based on their ataxia. At younger
Ubaidi et al., 1992). Transgenic mice in which T-antigen is ages (P5-P9), the behavioral phenotype is inadequate for genotype
driven by the promoter from the Purkinje cell-specific gene determination, so entire litters were processed with post hoc determi-
pcp-2 reveal similar results. In the presence of T-antigen nation of genotype based on the histological appearance of the cere-
expression, the maturing Purkinje cells die (Vandaele et al., bellum. All animals were maintained in the Case Western Reserve
1991). In vitro support for the association between ectopic cell animal facility in strict compliance with all relevant animal welfare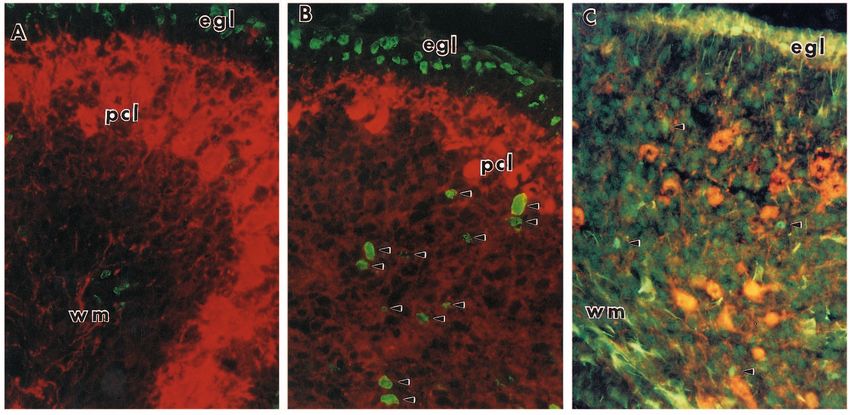
Multiple cell cycle events precede target-related neuronal death 2387
Table 1. Primary antibodies used in the current study guidlines. They were maintained on a 14/10 hour light/dark cycle and
Source of antibody Antigen Dilution Supplier
offered food and water ad libitum.
Mouse monoclonal BrdU 1:100 Becton-Dickinson Histology
Rat monoclonal BrdU 1:5 Accurate At various times prior to perfusion, the mice were injected subcuta-
Rabbit polyclonal Cyclin D 1:500 UBI
Mouse monoclonal PCNA 1:100 Santa Cruz
neously with 100 µg/g of the DNA precursor, bromodeoxyuridine
Mouse monoclonal Calbindin 1:1000 Sigma (BrdU). All mice were anesthetized with 0.02 cc/g body weight of
Rabbit polyclonal Calbindin 1:1000 Sylvia Christakos Avertin (1.25% 2,2,2-tribromoethanol and 0.625% 2-methyl-2-
Mouse monoclonal Glial fibrillary 1:100 Boehringer-Mannheim butanol in water). They were then killed via trans-cardiac perfusion
acidic protein with 4% paraformaldehyde in 0.1 M phosphate buffer for 15 minutes
Rabbit polyclonal Glial fibrillary 1:100 Accurate following a short PBS rinse. The entire brain was dissected free of
acidic protein the skull and left at 4°C in 4% paraformaldehyde overnight. The next
Mouse monoclonal NeuN (A60) 1:100 Richard Mullen morning the brain was transferred to an 18% sucrose solution in PBS.
Fig. 1. Comparison of the cerebella of wild-type, lurcher and staggerer mice. (A) A midline sagittal section of a wild-type P14 cerebellum.
Note the well-developed folding pattern, the robust internal granule cell layer (IGL) and the thin external granule cell layer (EGL), most
notably on the dorsal surface. Anterior (ANT) is to the right. (B) A midline sagittal section of a +/Lc cerebellum. Note the preservation of the
folding pattern despite a significant size reduction in the IGL. The EGL has nearly disappeared, even on the dorsalmost surface. Anterior
(ANT) is up. (C) A midline sagittal section of a P14 sg/sg cerebellum. This mutation leads to an even more dramatic loss of cells in the IGL
and a loss of the folding pattern than is visible in either of the other two animals. The EGL is thicker than in wild type and will persist longer.
(D) A higher magnification of the cells of the +/+ cerebellar cortex. Large, healthy Purkinje cells are present by P14 and there is virtually no
cell death in the IGL. (E) A similar view of the +/Lc cerebellar cortex. The Purkinje cells are smaller (the dark ones may be in the process of
dying) and there is significant cell death in the IGL, visible as basophilic pycnotic remains (arrows). (F) A similar view of the sg/sg cerebellar
cortex. Purkinje cells are small and disorganized, the granule cells are less dense than in either +/+ or +/Lc and there are large numbers of dying
cells present in the IGL (arrows). Initial magnification: A-C, 100×; D-F, 1600×.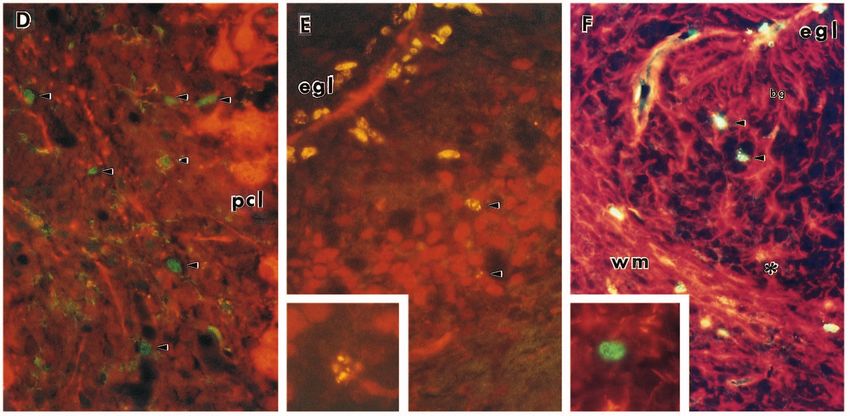
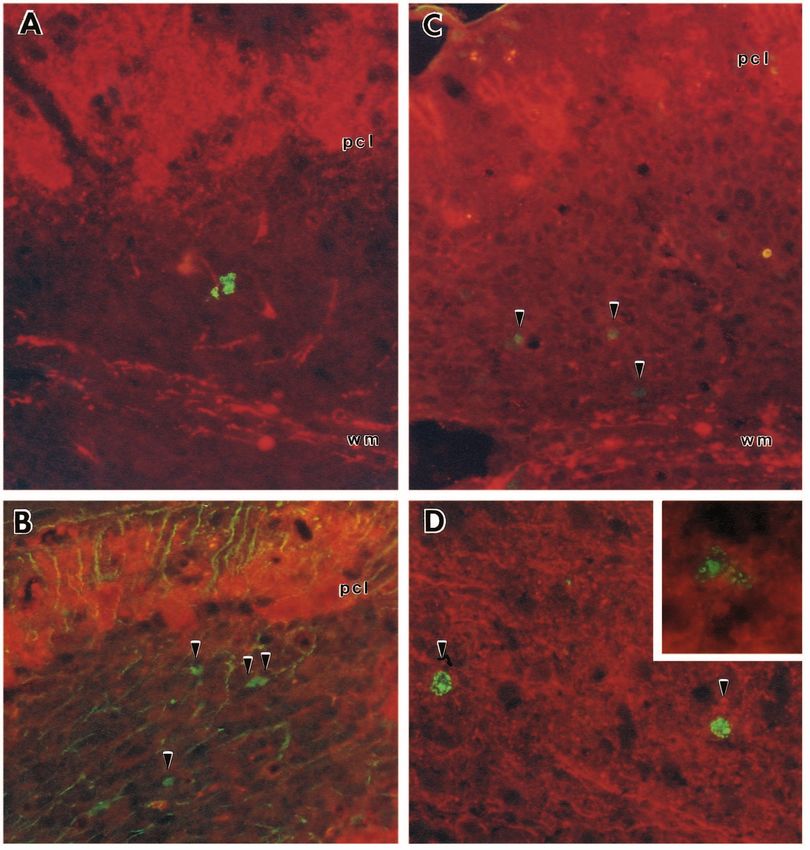
2388 K. Herrup and J. C. Busser Fig. 2. Appearance of cell cycle markers in postmigratory granule cells of the staggerer mutant mouse. (A) P6 wild-type cerebellar cortex immunostained with a Purkinje cell marker (calbindin – revealed with a rhodamine-tagged secondary antibody) and an antibody to BrdU (a DNA synthesis marker – revealed with a fluorescein-tagged secondary). The large Purkinje cells in the Purkinje cell layer (pcl) are stained completely with the calbindin marker. Cells in the mitotic EGL and in the white matter (wm) are labeled, but very few cells in the IGL appear labeled with the BrdU antibody. (B) P6 staggerer cerebellar cortex stained as in A. The EGL cells are labeled with BrdU, as expected, but so are unusual numbers of cells in the IGL (arrowheads). (C) A similar staining paradigm using anti-cyclin D primary antibody in place of the anti-BrdU primary. Again, the P14 EGL shows the strong labeling expected in a mitotic population; the white matter (wm) also shows a number of mitotic cells. The IGL, however, shows an unexpected number of brightly labeled cells (arrowheads). (D) A similar picture is revealed with PCNA antibody. (E) Many of the IGL cells are neurons. Cerebellar tissue from a sg/sg mouse was double labeled with BrdU antibody (fluorescein) and an antibody to the neuron-specific marker NeuN (rhodamine). Note the double-labeled cells in the IGL (arrowhead) and a third cell at higher magnification (inset). (F) Most of the BrdU+ cells are not glia. Double labeling with BrdU (fluorescein) and the glial cell marker, GFAP (rhodamine). Some glial cells are double-labeled with the BrdU marker (*), but most dividing cells in the IGL are not GFAP+ (arrowheads). One such negative cell is shown at higher magnification (inset). bg, Bergmann glial cells. Initial magnification: 1200×.
Multiple cell cycle events precede target-related neuronal death 2389
Fig. 3. Appearance of cell cycle markers in postmigratory cells of the lurcher mutant mouse. Older animals (P22) were used so that BrdU
injections could be done after the EGL had ceased to exist as a germinative region. (A) Cerebellar cortex of a lurcher mutant stained with
calbindin (rhodamine) to reveal the remaining Purkinje cells and anti-BrdU (fluorescein) to reveal the cells in S phase. A cohort of cells in
the IGL is stained with the cell-cycle marker. (B) Calbindin (rhodamine)/cyclin D (fluorescein) double labeling of a similar section. A
similar cluster of mitotic cells plus two others are stained with the cyclin D antibody (arrowheads). (C) Calbindin (rhodamine)/PCNA
(fluorescein) double-labeling of lurcher cerebellar cortex. Three dimly labeled cells can be seen in the IGL. The yellow labeling in the
dendritic field of one of the Purkinje cells in the Purkinje cell layer (pcl) is the autofluorescent material typical of macrophages. (D) Inferior
olive of a P6 sg/sg mouse. Calbindin (rhodamine) also labels the neurons in the inferior olive, a major source of afferents to the Purkinje
cell. 60% of these neurons die in the staggerer mutant and before they do, many can be labeled by short exposures to BrdU followed by
immunostaining with a fluorescein-tagged secondary (arrowheads). The inset is a higher magnification of an olive neuron from a P14 sg/sg.
Initial magnification: 2000×.
After at least 24 hours, the brain was embedded in OCT embedding perature and washed 6 times with PBS. The primary antibody (diluted
media, quick-frozen and sectioned by cryostat at 5 or 6 µm. in 20% goat serum/0.5% Tween/PBS) was applied overnight at room
temperature in a humid chamber. The following day, the sections were
Immunohistochemistry washed 3 times in PBS before the secondary antibody was applied.
Sections were treated in fresh 2 N HCl for 30 minutes at room tem- Sections were exposed to secondary antibodies (at a 1:200 dilution)2390 K. Herrup and J. C. Busser
for 2 hours in a humid chamber at room temperature. Nine different certain regions of the Purkinje cell layer where the +/Lc
primary antibodies were used. Their sources and concentrations are Purkinje cells have died and been removed. The remaining
listed in Table 1. Antibodies to cell cycle markers (BrdU, cyclin D, Purkinje cells are reduced in size from their wild-type coun-
PCNA) were viewed using a fluorescein-tagged secondary. Anti- terparts, but no examples of condensed ‘pycnotic’ Purkinje
bodies to cell-type-specific markers (calbindin, GFAP, NeuN) were cells were found in our material.
viewed using a rhodamine-tagged secondary. The secondary anti-
bodies were from Boehringer-Mannheim or Cappel with the exception Cell cycle markers in the staggerer cerebellum
of the fluorescein-tagged goat anti-rat IgG (non-cross reactive with
mouse IgG) and rhodamine-tagged goat anti-mouse (non-cross BrdU was injected into sg/sg and wild-type control mice 2
reactive with rat) which were from Jackson ImmunoResearch. In hours before killing by trans-cardial perfusion. BrdU is a
some cases, goat anti-BrdU was revealed with a peroxidase-conju- thymidine analog; any cell that is synthesizing DNA will incor-
gated secondary (Fig. 4A). In this case, a Tris-based buffer system porate it into its genome. After histological processing, all of
was used. Following application of the secondary antibody, slides the cells that were in S phase during the brief BrdU pulse will
were rinsed in TBS and the peroxidase revealed with diaminobenzi- stain with the BrdU antibody. When P6 wild-type cerebellar
dine (0.05%) and H2O2 (0.005%). The sections were then lightly tissue from BrdU-injected mice is processed in this manner and
counter-stained with quarter-strength Hematoxylin. stained with a fluorescein-tagged secondary antibody, the
End labeling of nicked DNA (TUNEL) expected cell types are labeled (Fig. 2A). These include the
Cryostat sections were hydrated in double-distilled water, and dividing cells in the mitotic zone of the EGL, a few cells in the
endogenous peroxidase was quenched by incubating the slides in 2% IGL (presumably astrocytic cells) and numerous cells in the
H2O2 for 5 minutes. Sections were equilibrated in reaction buffer (30 white matter (presumably the developing oligodendrocytes).
mM Tris pH 7.2/140 mM sodium cacodylate/2 mM cobalt chloride) As illustrated in Fig. 2B, if BrdU is injected into a P6 staggerer
for 25 minutes then incubated with 0.25 U/µl TdT and 5 µM biotiny- mouse 2 hours before killing, the dividing cells in the EGL are
lated-dUTP (both from Boehringer-Mannheim) in reaction buffer for labeled as expected, but there is a noticeable increase in the
60 minutes at 37°C in a humid chamber. The reaction was stopped by number of cells in the IGL that have incorporated BrdU into
incubation in 2× SSC for 15 minutes and a water rinse. The sections DNA (Fig. 2B, arrowheads).
were then blocked using 2% BSA in double-distilled water for 10 To explore the possibility that this incorporation of BrdU
minutes followed by a 5 minute wash in TBS. The sections were then
incubated in peroxidase-conjugated streptavidin (Boehringer-
might represent an artifact associated with DNA breakdown
Mannheim) diluted 1:20 in TBS for 60 minutes at room temperature. and repair in the dying staggerer granule cells, sections were
After washing in TBS for 5 minutes, the end labeling was visualized stained with two independent cell cycle markers. Cyclin D is
with diaminobenzidine (0.05%) and H2O2 (0.005%). The sections a nuclear protein found in three different isoforms whose con-
were then lightly counter-stained with quarter-strength Hematoxylin. centration oscillates with the cell cycle and which is thought
All results were viewed on either a Zeiss Axiophot (courtesy of Dr to regulate the progression of a cell from G1 to S phase
H. Willard) or a Leica DM RB microscope. The fluorescent pictures (reviewed in Sherr, 1993). Proliferating cell nuclear antigen
were taken using a dual rhodamine/fluorescein cube. (PCNA) is a subunit of the DNA polymerase holoenzyme
whose concentration rises during early S phase and remains
high through G2 phase (Bravo et al., 1987). If a wild-type cere-
RESULTS bellum is stained with cyclin-D antibody, the cells of the EGL
are stained and very few others (not shown). Staining of wild-
Review of normal development type cerebella with antibody to PCNA reveals a similar
Most of the development of the mouse cerebellum occurs post- situation. Staining of P14 staggerer cerebella with either cyclin
natally. Nearly the entire granule cell population is generated D (Fig. 2C) or PCNA (Fig. 2D), however, reveals a picture
after birth and there is substantial cytological and biochemical identical to that found with the BrdU labeling, namely, an
maturation of all of the other cellular elements including the unusually large number of cells in the IGL that are evidently
Purkinje cells, Golgi II neurons, astrocytes and the specialized in various phases of the cell cycle from very late G1 through
glial cells known as Bergmann glial cells. In the wild-type, by S phase.
postnatal day 14, most neurogenesis has been completed in the
external granule cell layer (EGL), the size and density of the Dividing cells are granule cells
internal granule cell layer (IGL) have begun to assume normal The presence of cycling cells in the staggerer IGL is not unex-
adult values and the folia are well defined (Fig. 1A). By com- pected. Astrocyte proliferation occurs during the early
parison, development in the two mutants lags considerably. postnatal period and, as mentioned above, the wild-type IGL
The staggerer cerebellum is significantly smaller than its age- does contain an occasional BrdU-positive cell. There is also
matched control (Fig. 1C). A clear IGL is present, but its gliosis in the staggerer (Hirano and Dembitzer, 1976) which
density is reduced from normal values and there are many should serve to increase further the apparent number of
examples of dying, ‘pycnotic’ cells in this layer (arrows, Fig. dividing cells. To show that the cells incorporating BrdU were
1F). The Purkinje cells are small, malpositioned and reduced neurons, we double stained staggerer cerebella with antibody
in number from the wild-type (Fig. 1F, compare with the +/+ to the NeuN antigen, a neuron-specific nuclear antigen (Mullen
in Fig. 1D). The lurcher cerebellum is also reduced in size et al., 1992). Many of the BrdU-positive cells were
compared to normal at this age (Fig. 1B) with a granule cell BrdU+/NeuN+ (Fig. 2E, arrowheads), identifying them as
layer that is reduced in density. In lurcher, as in staggerer, neurons. To emphasize this point further, we double stained
numerous ‘pycnotic’ granule cells are found scattered through- alternate sections for the glial cell marker, GFAP and BrdU.
out the IGL (arrows, Fig. 1E). Though Purkinje cells are in Some of the cells in the IGL were indeed double labeled (Fig.
evidence at this relatively young age, there are also gaps in 2F, asterisk), but some labeled only with BrdU (Fig. 2F, arrow-Multiple cell cycle events precede target-related neuronal death 2391
heads and inset). Since 100% of the granule cells in the no Purkinje cell was found in S phase, i.e., none was labeled
staggerer mutant will eventually die a target-related cell death, with both calbindin and BrdU. We take this as evidence that
the finding that significant numbers of normally postmitotic the loss of Purkinje cells in the lurcher mouse is not preceded
IGL granule cells re-enter the cell cycle means that ectopic cell by a re-entry into the cell cycle.
division must precede the neurons’ demise.
The nature and timing of the neuronal deaths
Cell cycle markers in the lurcher cerebellum To determine the timing of the granule cell death with respect
The lurcher mutant also loses granule cells to target-related to the cell cycle, animals were injected with BrdU, killed after
cell death, and we found similar numbers of BrdU+-, cyclin varying survival times and then stained with BrdU antibody
D+- and PCNA+-positive cells in the IGL of this mutant (Fig. using a peroxidase-linked secondary antibody. As illustrated in
3A-C). As in staggerer, some of these dividing cells could be Fig. 4A, a five hour time interval between administration of the
identified as granule cells since they were NeuN+/BrdU+ (not thymidine analog and killing was sufficient to label pycnotic
shown). These observations in lurcher are less conclusive, granule cells in the IGL. The cell indicated by the arrow in this
however, since 10% of the lurcher granule cells will survive figure must have been in S phase no more than 5 hours before
into adulthood. It is, therefore, impossible to be certain that any dying. To enhance the kinetic picture, three staggerer and three
one labeled granule cell will die. control (sg/+ or +/+) pups from a single litter were injected
with BrdU on postnatal day 17. The animals were killed by
Association of cell cycle markers with cell death in perfusion 5, 10 or 20 hours later. Serial sections were obtained
the inferior olive from all six animals and ten sections from each animal were
A second major projection to the cerebellar Purkinje cells stained with antibody to BrdU. In each section, the number of
comes from the cells of the inferior olivary nucleus located on pycnotic cells, BrdU-labeled cells and double labeled (pycnotic
the ventral surface of the caudal brain stem. Similar to the and BrdU-labeled) cells in the internal granule cell layer was
granule cells, the inferior olive neurons have a developmental counted. The results are presented in Fig. 5.
target-dependency on the cerebellar Purkinje cells. Given the These data reveal several important aspects of the cell death
Purkinje cell deficiencies in the two mutants, it is not surpris- process. First, of the cells in the IGL that incorporate BrdU, a
ing that 70% of the olive neurons in lurcher and 60% in significant fraction go on to die (at any one time, up to 30% of
staggerer die postnatally. We also found evidence for cell the pycnotic cells are found to be labeled with BrdU). Second,
cycle progression prior to cell death in the inferior olive. Fig. cell death (pycnosis) occurs reasonably quickly after BrdU
3D illustrates that cells in the sg/sg inferior olive complex incorporation. From Fig. 5A it appears that the death of the
could be double-labeled with BrdU and calbindin (also a labeled cohort of cells peaks at about 10 hours. Third, the small
marker of inferior olivary neurons – Christakos et al., 1989). amount of cell death that occurs during normal cerebellar
Staining of the +/Lc animals revealed a similar picture, but no development is also accompanied by an induction of DNA
such cells were found in age-matched, wild-type controls. synthesis and the kinetics of the process appear quite similar.
Since olivary neuron loss is not complete in either mutant, it The total number of pycnotic granule cells in a normal animal
is not possible to know with certainty that ectopic cell cycling is of course much smaller than the number found in staggerer.
in any individual cell will precede its death. Nonetheless, Finally, Fig. 5B suggests that the pycnotic process itself may
coupled with the results from the staggerer granule cells, the be quite different in staggerer and normal animals. In the
evidence suggests that target-related neuronal death is corre- control animals (circles), the BrdU-labeled pycnotic remnants
lated with an unscheduled re-entry into the cell cycle in this are cleared rapidly (of the total number of BrdU-labeled cells,
second cell type as well. fewer than 10% are pycnotic after 20 hours). In the mutants,
however, the labeled cells must be cleared quite slowly as the
Neuronal death is not always associated with fraction of the labeled cells that are pycnotic actually increases
ectopic cell cycles up to 20 hours.
The association of abortive cell division and target-related cell The evidence for unscheduled re-entry into the cell cycle
death led us to question whether this cell cycle re-entry might preceding the death of the granule cells and inferior olive
precede all instances of nerve cell death. To address this neurons implies that these normally postmitotic cells are
question, we have examined the Purkinje cell population of the undergoing major alterations in their chromatin structure
lurcher. All of the lurcher Purkinje cells die between birth and during some of the most active times in their maturation
P90 (Caddy and Biscoe, 1979). A series of lurcher animals was process. This in turn suggested to us that the cells might be
injected with BrdU shortly before killing. Cryostat sections of vulnerable to the degradative processes that underlie
cerebellum were then stained with anti-BrdU antibody to apoptotic cell death. To test this hypothesis, we applied the
reveal dividing cells and with anti-calbindin to identify all TUNEL technique of DNA end-labeling with terminal
Purkinje cells. At least one dying Purkinje cells would be deoxynucleotidyl transferase (TdT) to cerebellar sections
expected in each section*. Five to ten sections from each of from wild-type and mutant animals at P14. Unexpectedly,
six lurcher mice of different postnatal ages were examined, but none of the pycnotic granule cells in the IGL of either mutant
was labeled with this technique (Fig. 4B,C). This includes the
pycnotic remains of the dead granule cells that are common
*This estimate is based on the number of sections per half cerebellum (~250) and on the
assumptions that the original 100,000 Purkinje cells (per half cerebellum) will die over a at these ages (Fig. 4B, arrows). Rarely, a TUNEL-labled cell
period of 10 weeks, leaving morphologically visible traces for 8 hours. As this method could be found in the staggerer or lurcher EGL (Fig. 4B,C,
of calculation assumes a linear decay of Purkinje cells, it is surely an underestimate. The
true rate of death during the period examined (the second postnatal week) is likely to be
arrowheads), but the significance of these observations is
five to ten fold greater (Caddy and Biscoe, 1979). uncertain. TUNEL labeling was easily detected in the2392 K. Herrup and J. C. Busser
Fig. 4. Timing of granule
cell death and
involvement of apoptotic
mechanisms.
(A) Cerebellar cortex
from a staggerer mouse
injected 5 hours before
killing with BrdU and
stained with anti-BrdU
antibody followed by a
peroxidase-conjugated
secondary antibody. The
latter was revealed with
DAB substrate leaving a
brown reaction product.
A cell, labeled with
BrdU, has begun to die
as indicated by its
pycnotic appearance
(arrow). The external
granule cell layer (egl)
contains several cells
that were in S phase. ml,
molecular layer; igl,
internal granule cell
layer. (B) TUNEL
labeling of apoptotic cell
nuclei in staggerer
cerebellum. Note that
although several
pycnotic remnants are
visible in this photograph
(arrows), no cell in the
IGL is labeled with
brown DAB reaction
product. The field chosen
illustrates the rare
occurance of a labeled
cell in the EGL.
(C) TUNEL labeling of
apoptotic cell nuclei in
the lurcher mouse. As in
the previous image, no
IGL cells can be shown
to be dying by apoptosis
in this mutant. Again, the
EGL has examples of
rare TUNEL-positive
cells. (D) By contrast,
the dying thymocytes of
the neonatal thymus are easily labeled with the terminal transferase enzyme (arrowheads) indicating that apoptosis can be demonstrated in our
material in this classic example of genetically preprogrammed cell death. Initial magnification: 5000×.
neonatal thymus tissue used as a control (Fig. 4D). The DISCUSSION
absence of TUNEL labeling should be weighed against other
morphological evidence strongly favoring the apoptotic The data presented here add to the growing body of evidence
nature of the granule cell deaths. The dense clumped appear- suggesting that when a postmitotic neuron re-enters the cell
ance of the pycnotic cells shown in Figs 1 and 4 is one such cycle it dies. The most straight-forward example of this
piece of evidence. Further, previous ultrastructural examina- relationship is the cerebellar granule cell population of the
tions of degenerating staggerer granule cells have revealed staggerer. The double-immunostaining of sg/sg granule cells
nuclear condensation in advance of cytoplasmic alterations with BrdU and the NeuN neuronal nuclear antigen demon-
(Landis and Sidman, 1978; Sotelo and Changeux, 1974) and strates clearly that normally postmitotic neurons in the
clear examples of cytoplasmic membrane blebbing (see Fig. staggerer IGL are induced by the conditions in the mutant
23 in Landis and Sidman, 1978). cerebellum to begin DNA synthesis. That this synthesisMultiple cell cycle events precede target-related neuronal death 2393
Fig. 5. The kinetics of granule cell
A B death in staggerer cerebellum.
staggerer and control pups from a
60% 60% single litter were injected with BrdU
and killed after varying delays. Cell
% BrdU labeled cells that are
50% 50% counts were performed and the results
% pycnotic cells that are
expressed as (A) the percentage of the
40%
total number of pycnotic cells that
BrdU labled
40%
were labeled with BrdU or (B) the
pycnotic
percentage of the total BrdU-labeled
30% 30% cells that were pycnotic. Control
counts (+/+ or sg/+) are represented
20% 20% by circles. Small unfilled circles are
the individual data points; large filled
10% 10% circles represent the mean values.
Counts from staggerer animals are
represented by triangles. Small
5 10 15 20 25
unfilled triangles are the individual
5 10 15 20 25
data points; large filled triangles
Hours after injection Hours after injection represent the mean values.
actually represents an attempt to re-enter the cell cycle is that terms such as S phase and G2 may have no true mecha-
supported by the observation of increased expression of two nistic meaning.
independent cell cycle markers, cyclin D and PCNA. Since the During the preparation of this manuscript an experiment
natural history of the mutant phenotype allows us be certain conducted over a decade ago in the chick was brought to our
that all of the sg/sg granule cells will ultimately die, we may attention. In an attempt to determine whether the axonal
conclude that this unscheduled re-entry into the cell cycle is process of a dorsal root ganglion neuron reaches its peripheral
associated with the death of these cells. The observations of a target before it succumbs to target-related neuronal death, Carr
similar association between cell division and cell death in and Simpson (1982) killed chick embryos at various times after
lurcher granule cells and in the inferior olivary neurons in both [3H]thymidine injection. Their hypothesis was that a recently
mutants implies that the same mechanism is working in these generated neuron could not have had the time to send its axon
cells as well. This latter conclusion is less definitive, however, to its target field. Thus the time between genesis and naturally
since in each of these populations, a fraction of the cells occurring cell death would provide an indication of whether,
survives into adulthood making it impossible to know with and for how long, the cell had direct contact with its target. To
certainty that any one cell that we observe to have re-entered the authors’ surprise, they found that 24% of the degenerating
the cell cycle will finally die. cells of the ganglion were labeled within 2 hours of injection.
It is likely that the cell cycle events we are observing The percentage of labeled pycnotic cells increased until 24
represent cells re-entering rather than failing to leave the cell hours (reaching 40% to 50%) after which it declined. Further,
cycle. The most clearcut evidence of this are the cells of the while target ablation (wing bud removal) increased the total
inferior olivary complex. These cells normally undergo their amount of pycnosis, it did not significantly change the per-
final cell division on embryonic day 11 (E11, Taber-Pierce, centage of dying cells that were labeled at one time after
1973), a full two weeks before we find evidence for cell cycle injection. The interpretation of these experiments is compli-
activity. Further, normal granule cells become postmitotic cated by the fact that legitimate neurogenesis is occuring in the
before they leave the EGL, requiring more than 24 hours to same population as the cell deaths. Nonetheless, the qualitative
complete G2 phase, divide and migrate across the molecular and quantitative agreement of the two data sets hints that the
layer to the IGL. correlation of cell cycle re-entry with target-related cell death
We can also estimate that the death of the ectopically may be found in a variety of cell types of both the central and
dividing cells must occur either late in S phase or early in G2. peripheral nervous systems.
2 hours after BrdU injection, anti-BrdU labeling can be found The persistent association of unscheduled cell division and
in the pycnotic remnants of dying granule cells, implying that cell death in the nervous system (al-Ubaidi et al., 1992;
these cells were recently in S phase. This result is similar to Freeman et al., 1994; Lee et al., 1992; Vandaele et al., 1991;
that of Lee et al. (1994) in their study of dying cells in Yonish-Rouach et al., 1993) raises the question of why a
retinoblastoma knockout mice. Indeed, our kinetic studies (Fig. neuron would die if it attempted to divide. Heintz (1993) has
5) suggest that, once DNA synthesis begins, the average suggested that maturing neurons re-tool their signal transduc-
granule cell becomes pycnotic after approximately 10 hours. tion machinery such that, upon receiving a cell division signal,
Finally, while mitotic figures are easily detected in the outer their response is switched to an ‘alternate effector pathway’.
EGL, virtually none is found in the IGL, and those that we do Thus, a signal that induces cell division in a ventricular zone
find could easily be dividing astrocytes (no cell type-specific cell induces a different set of genes in a mature neuron and this
marker has been used). Together these data suggest that induction leads to programmed cell death. The results reported
granule cells die during late S- or early G2 phase. Our estimates here suggest a modification of this hypothesis. Three different
are, of course, limited by the possibility that cell cycle regula- cell cycle markers are all activated in our model system in
tion is so disorganized in these normally postmitotic neurons response to target-deprivation: cyclin D (a G1/S phase cell2394 K. Herrup and J. C. Busser
cycle protein), PCNA (a DNA polymerase subunit expressed bcl-2-related gene that functions as a dominant regulator of apoptotic cell
from late G1 to late G2/early M phase) and DNA synthesis (as death. Cell 74, 597-608.
measured by BrdU incorporation). Our interpretation of this Bowen, I. D. (1993). Apoptosis or programmed cell death? Cell Biol. Int. Rep.
17, 365-380.
data is that, for granule cells and inferior olivary neurons, some Bravo, R., Frank, R., Blundell, P. and MacDonald-Bravo, H. (1987). PCNA
aspect of the cell division process itself is toxic. Perhaps the is the auxiliary protein of DNA polymerase-delta. Nature 326, 515-520.
drain on the pool of nucleotide precursors, or abnormal ionic Caddy, K. W. T. and Biscoe, T. J. (1979). Structural and quantitative studies
fluxes (e.g., calcium; see Whitaker and Patel, 1990), cannot be on the normal C3H and lurcher mutant mouse. Phil. Trans. Roy. Soc. (Lond.)
B 287, 167-201.
tolerated by the cell. Equally likely, however, is that the dis- Carr, V. M. and Simpson, S. B. J. (1982). Rapid appearance of labeled
organization of chromatin that must accompany DNA replica- degenerating cells in the dorsal root ganglia after exposure of chick embryos
tion in an unprepared cell may lead to genetic imbalances that to tritiated thymidine. Devel. Brain Res. 2, 157-162.
cannot be tolerated. In this regard, it is noteworthy that Christakos, S., Gabrielides, C. and Rhoten, W. (1989). Vitamin D-dependent
mutations have been identified that define specific cell cycle calcium binding proteins: chemistry, distribution, functional considerations
and molecular biology. Endocrine Rev. 10, 3-26.
checkpoints in both S phase and in G2 phase of the cell cycle Clarke, A., Maandag, E., van Roon, M., van der Lugt, N., van der Valk, M.,
(Earnshaw and Pluta, 1994; Hartwell and Weinert, 1989; Hooper, M., Berns, A. and te Riele, H. (1992). Requirement for a
Weinert et al., 1994). The apparent functions of the wild-type functional Rb-1 gene in murine development. Nature 359, 328-330.
gene products are to halt further progression of the cell cycle Driscoll, M. and Chalfie, M. (1992). Developmental and abnormal cell death
in C. elegans. Trends in Neurosci. 15, 15-19.
until the genome has been properly prepared, and cells lacking Dumesnil-Bousez, N. and Sotelo, C. (1992). Early development of the Lurcher
these functions will die if they process through cell division. cerebellum: Purkinje cell alterations and impairment of synaptogenesis. J
In the CNS, it may be that the DNA damage sustained by a Neurocytol. 21, 506-29.
mature neuron when it tries to divide is simply incompatible Earnshaw, W. C. and Pluta, A. F. (1994). Mitosis. BioEssays 19, 639-643.
with the proper regulation of gene expression and thus leads to Freeman, R., Estus, S. and Johnson, E. (1994). Analysis of cell cycle-related
gene expression in postmitotic neurons: selective induction of cyclin D1
death. This might be especially true in the nervous system during programmed cell death. Neuron 12, 343-355.
where a disproportionately large fraction of the total genome Gagliardini, V., Fernandez, P.-A., Lee, R., Drexler, H., Rotello, R.,
is transcribed (Bantle and Hahn, 1976; He and Rosenfeld, Fishman, M. and Yuan, J. (1994). Prevention of vertebrate neuronal death
1991; Milner and Sutcliffe, 1983). by the crmA gene. Science 263, 826-828.
Whatever the ultimate mechanism by which unscheduled Goodrich, D., Wang, N., Qian, Y., Lee, E. and Lee, W. (1991). The
retinoblastoma gene product regulates progression through the G1 phase of
cell division causes a postmitotic neuron to die, there is a the cell cycle. Cell 67, 293-302.
growing recognition that the correlation between cell death Hartwell, L. H. and Weinert, T. A. (1989). Checkpoints: controls that ensure
and cell division is potentially significant for a wide variety the order of cell cycle events. Science 246, 629-634.
of neurodegenerative diseases. Neurons of the CNS are He, X. and Rosenfeld, M. (1991). Mechanisms of complex transcriptional
regulation: implications for brain development. Neuron 7, 183-196.
unusual in that they fail to develop tumors. Perhaps this is Heintz, N. (1993). Cell death and the cell cycle: a relationship between
because any ‘second hit’ in a nerve cell leads not to a clone transformation and neurodegeneration? Trends Biochem. Sci. 18, 157-159.
of tumor cells, but rather to a single dead neuron. This Herrup, K. (1983). Role of staggerer gene in determining cell number in
suggests, in turn, that late onset, degenerative disorders might cerebellar cortex. I. Granule cell death is an indirect consequence of staggerer
result from autocrine or paracrine stresses that force suscepti- gene action. Devel. Brain Res. 11, 267-274.
Herrup, K. and Mullen, R. J. (1979a). Regional variation and absence of large
ble populations into the cell cycle and hence to die. The T- neurons in the cerebellum of the staggerer mouse. Brain Res. 172, 1-12.
antigen transgenic mice provide additional perspectives on Herrup, K. and Mullen, R. J. (1979b). Staggerer chimeras: intrinsic nature of
dominantly inherited neurodegenerative disease. One source Purkinje cell defects and implications for normal cerebellar development.
of blood cell tumors is chromosomal translocations that Brain Res. 178, 443-457.
Herrup, K. and Mullen, R. J. (1981). Role of staggerer gene in determining
juxtapose an immune system regulatory element and an cell number in the cerebellar cortex: a quantitative analysis of staggerer
oncogene. A comparable event with a neuron-specific chimeric mice. Devel. Brain Res. 1, 475-485.
enhancer might produce a population-specific cell death. Hirano, A. and Dembitzer, H. (1976). The fine structure of the astrocytes in
These observations suggest new avenues of both genetic and the adult staggerer. J. Neuropathol. Exp. Neurol. 35, 63-74.
pathological exploration of the etiology of several important Jacks, T., Fazeli, A., Schmitt, E., Bronson, R., Goodell, M. and Weinberg,
R. (1992). Effects of an Rb mutation in the mouse. Nature 359, 295-300.
human neurodegenerative diseases. Landis, D. M. D. and Sidman, R. L. (1978). Electron microscopic analysis of
postnatal histogenesis in the cerebellar cortex of staggerer mutant mice. J.
The authors gratefully acknowledge Dr Sylvia Christakos for the Comp. Neurol. 179, 831-863.
supply of calbindin antiserum and Dr Richard Mullen for the NeuN Lee, E. Y.-H. P., Chang, C.-Y., Hu, N., Wang, Y.-C. J., Lai, C.-C., Herrup,
antibody. We are especially grateful for the helpful conversations with K., Lee, W.-H. and Bradley, A. (1992). Mice deficient for Rb are nonviable
Dr Virginia McMillan Carr concerning her earlier studies in the chick. and show defects in neurogenesis and haematopoiesis. Nature 359, 288-
This work was supported by a grant from the NIH (NS-20591). 294.
Lee, E. Y.-H. P., Hu, N., Yuan, S.-S. F., Cox, L. A., Bradley, A., Lee, W.-H.
and Herrup, K. (1994). Dual roles of the RB protein in cell cycle regulation
and neuron differentiation. Genes Dev. 8, 2008-2021.
REFERENCES Martin, D., Schmidt, R., DeStefano, P., Lowry, O., Carter, J. and Johnson,
E. (1988). Inhibitors of protein synthesis and RNA synthesis prevent
al-Ubaidi, M. R., Hollyfield, J. G., Overbeek, P. A. and Baehr, W. (1992). neuronal death caused by nerve growth factor deprivation. J. Cell Biol. 106,
Photoreceptor degeneration induced by the expression of simian virus 40 829-844.
large tumor antigen in the retina of transgenic mice. Proc. Natl Acad. Sci. Messer, A., Eisenberg, B. and Plummer, J. (1991). The Lurcher cerebellar
USA 89, 1194-1198. mutant phenotype is not expressed on a staggerer mutant background. J.
Bantle, J. and Hahn, W. (1976). Complexity and characterization of Neurosci. 11, 2295-302.
polyadenylated RNA in the mouse brain. Cell 8, 139-150. Messer, A., Plummer-Siegard, J. and Eisenberg, B. (1990). Staggerer mutant
Boise, L. H., González-García, M., Postema, C. E., Ding, L., Lindsten, T., mouse Purkinje cells do not contain detectable calmodulin mRNA. J.
Turka, L. A., Mao, X., Nuñez, G. and Thompson, C. B. (1993). bcl-x, a Neurochem. 55, 293-302.Multiple cell cycle events precede target-related neuronal death 2395 Messer, A., Savage, M. and Carter, T. P. (1981). Thymidine kinase activity is cascade’ in cerebellar cortex of staggerer mutant mice. Brain Res. 67, 519- reduced in the developing staggerer cerebellum. J. Neurochem. 37, 1610-2. 526. Milner, R. and Sutcliffe, J. (1983). Gene expression in rat brain. Nucleic Acids Taber-Pierce, E. (1973). Time of origin of neurons in the brain stem of the Res. 11, 5497-5520. mouse. Prog. Brain Res. 40, 53-65. Miura, M., Zhu, H., Rotello, R., Hartwieg, E. and Yuan, J. (1993). Induction Tano, D., Napieralski, J., Eisenman, L., Messer, A., Plummer, J. and of apoptosis in fibroblasts by IL-1 beta-converting enzyme, a mammalian Hawkes, R. (1992). Novel developmental boundary in the cerebellum homolog of the C. elegans cell death gene ced-3. Cell 75, 653-660. revealed by zebrin expression in the lurcher (Lc/+) mutant mouse. J. Comp. Mullen, R., Buck, C. and Smith, A. (1992). NeuN, a neuronal specific nuclear Neurol. 323, 128-36. protein in vertebrates. Development 116, 201-211. Vandaele, S., Nordquist, D. T., Feddersen, R. M., Tretjakoff, I., Peterson, Oppenheim, R., Prevette, D., Tytello, M. and Homma, S. (1990). Naturally A. C. and Orr, H. T. (1991). Purkinje cell protein-2 regulatory regions and occurring and induced cell death in the chick embryo in vivo requires protein transgene expression in cerebellar compartments. Genes Dev. 5, 1136-48. and RNA synthesis. Dev. Biol. 138, 104-113. Weinert, T., Kiser, G. and Hartwell, L. (1994). Mitotic checkpoint genes in Pittman, R. H. and Oppenheim, R. W. (1978). Neuromuscular blockade budding yeast and the dependence of mitosis on DNA replication and repair. increases motoneurone survival during normal cell death in the chick Genes Dev. 8, 652-665. embryo. Nature 271, 364-366. Wetts, R. and Herrup, K. (1982a). Interaction of granule, Purkinje and Reed, J. C. (1994). Bcl-2 and the regulation of programmed cell death. J. Cell inferior olivary neurons in lurcher chimeric mice. II. Granule cell death. Biol. 124, 1-6. Brain Res. 250, 358-362. Schwartz, L. M. and Osborne, B. A. (1993). Programmed cell death, Wetts, R. and Herrup, K. (1982b). Interaction of granule, Purkinje and olivary apoptosis and killer genes. Immunol. Today 14, 582-590. neurons in lurcher chimeric mice. I. Qualitative studies. J. Embryol. Exp. Schwartzman, R. A. and Cidlowski, J. A. (1993). Apoptosis: The Morphol. 68, 87-98. biochemistry and molecular biology of programmed cell death. Endocrin. Whitaker, M. and Patel, R. (1990). Calcium and cell cycle control. Rev. 14, 133-151. Development 108, 525-542. Scott, S. and Davies, A. (1990). Inhibition of protein synthesis prevents cell Williams, R. W. and Herrup, K. (1988). Control of cell number in the nervous death in sensory and parasympathetic neurons deprived of neurotrophic system. Ann. Rev. Neurosci. 11, 423-453. factor in vitro. J. Neurobiol. 21, 630-638. Yonish-Rouach, E., Grunwald, D., Wilder, S., Kimchi, A., May, E., Sherr, C. (1993). Mammalian G1 cyclins. Cell 73, 1059-1065. Lawrence, J. J., May, P. and Oren, M. (1993). p53-Mediated cell death: Sidman, R. L. (1968). Development of interneuronal connections in brains of Relationship to cell cycle control. Mol. Cell Biol. 13, 1415-1423. mutant mice. In Physiological and Biochemical Aspects of Nervous Yuan, J., Shaham, S., Ledoux, S., Ellis, H. M. and Horvitz, H. R. (1993). Integration, (ed. Carlson, S. D.), pp. 163-193. Englewood Cliffs, N.J.: The C. elegans cell death gene ced-3 encodes a protein similar to mammalian Prentice Hall. interleukin-1beta-converting enzyme. Cell 75, 641-652. Sidman, R. L. (1977). Experimental neurogenetics. In Genetics of Zanjani, H., Mariani, J. and Herrup, K. (1990). Cell loss in the inferior olive Neurological Disorders (ed. Sidman, R. L. and Matthysse, S. W.), pp. 19-46. of the staggerer mutant mouse is an indirect effect of the gene. J. Neurogenet. New York: Raven. 6, 229-241. Smeyne, R., Vendrell, M., Hayward, M., Baker, S., Miao, G., Schilling, K., Zhong, L., Sarafian, T., Kane, D. J., Charles, A. C., Mah, S. P., Edwards, R. Robertson, L., Curran, T. and Morgan, J. (1993). Continuous c-fos H. and Bredesen, D. E. (1993). bcl-2 inhibits death of central neural cells expression precedes programmed cell death in vivo. Nature 363, 166- induced by multiple agents. Proc. Natl Acad. Sci. USA 90, 4533-4537. 169. Sotelo, C. and Changeux, J.-P. (1974). Transsynaptic degeneration ‘en (Accepted 5 May 1995)
You can also read

















































